
Survey of Oomycetes Associated with Root and Crown Rot of Almond in Spain and Pathogenicity of Phytophthora niederhauserii and Phytopythium vexans to ‘Garnem’ Rootstock

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
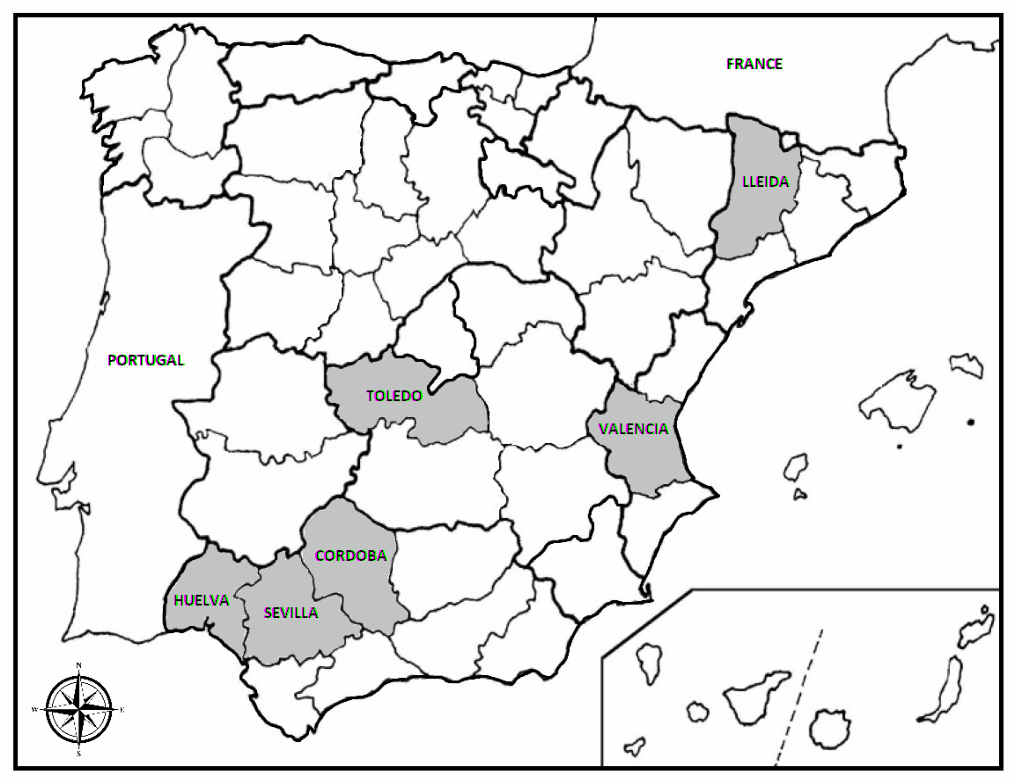
2.1. Field and Nursery Survey and Sampling
2.2. Isolation of Oomycetes from Plant and Soil Samples
2.3. DNA Isolation, Amplification and Sequencing of Oomycetes
2.4. Pathogenicity Tests
2.5. Data Analysis
3. Results
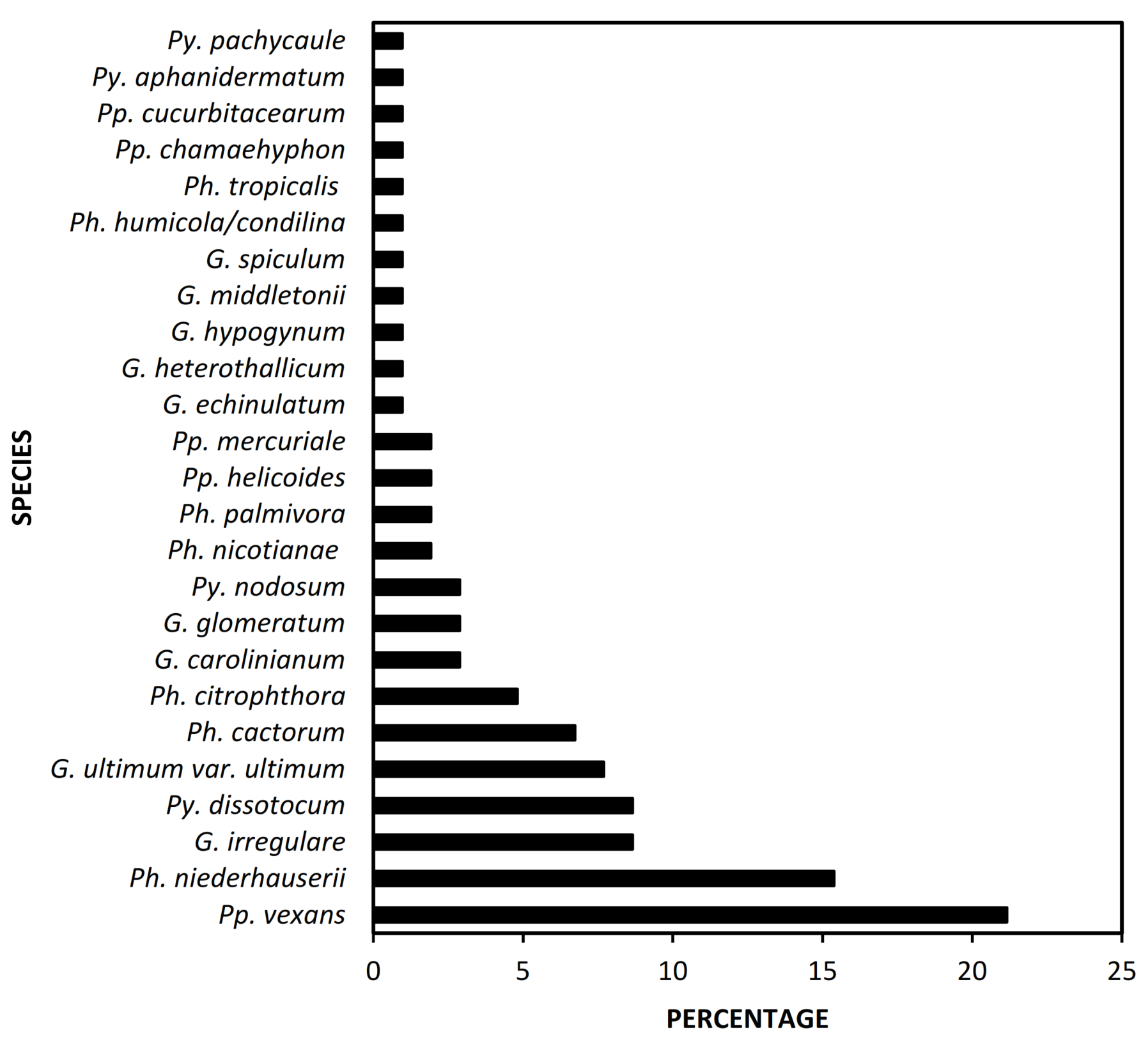
3.1. Occurrence of Oomycetes in Almond Samples
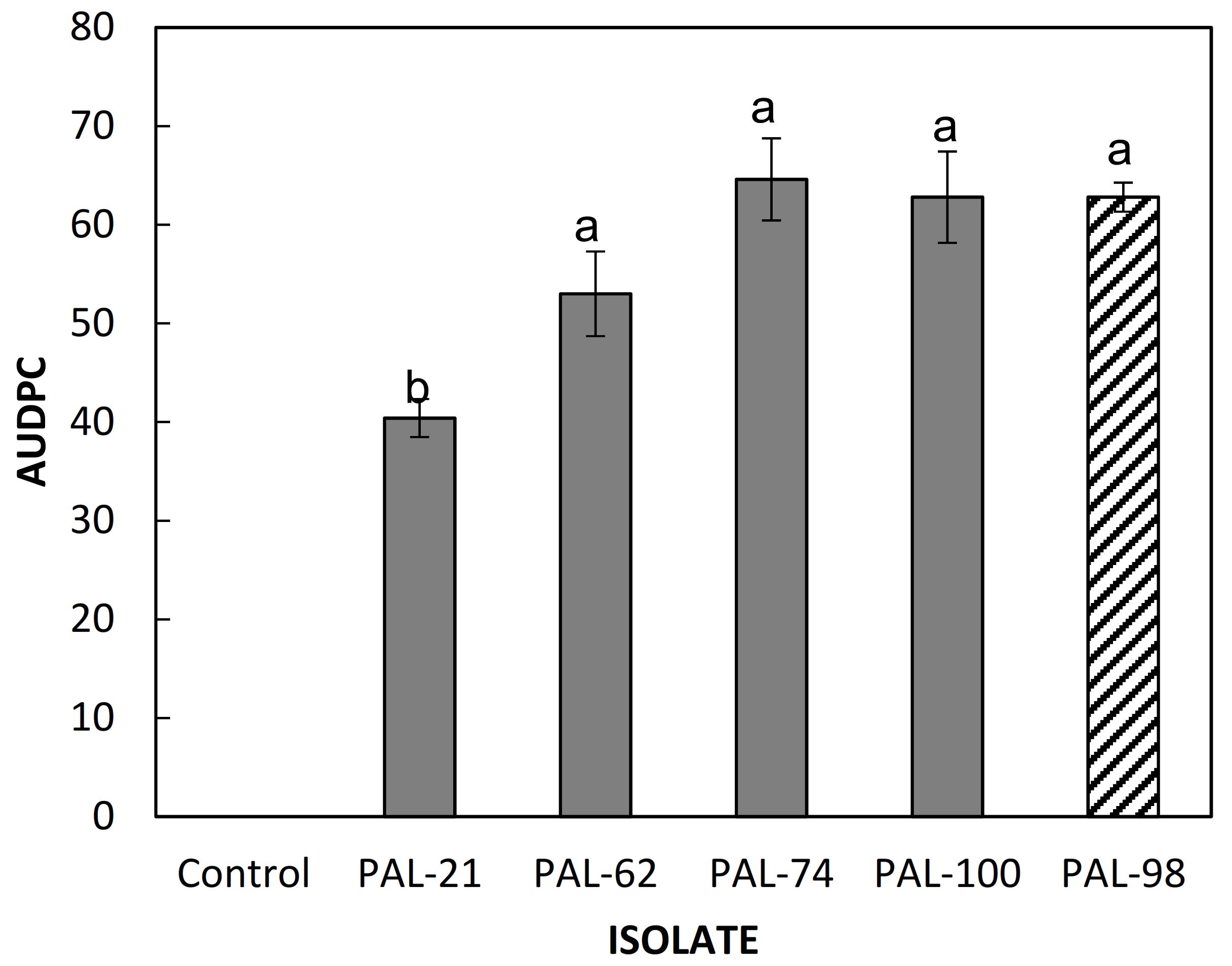
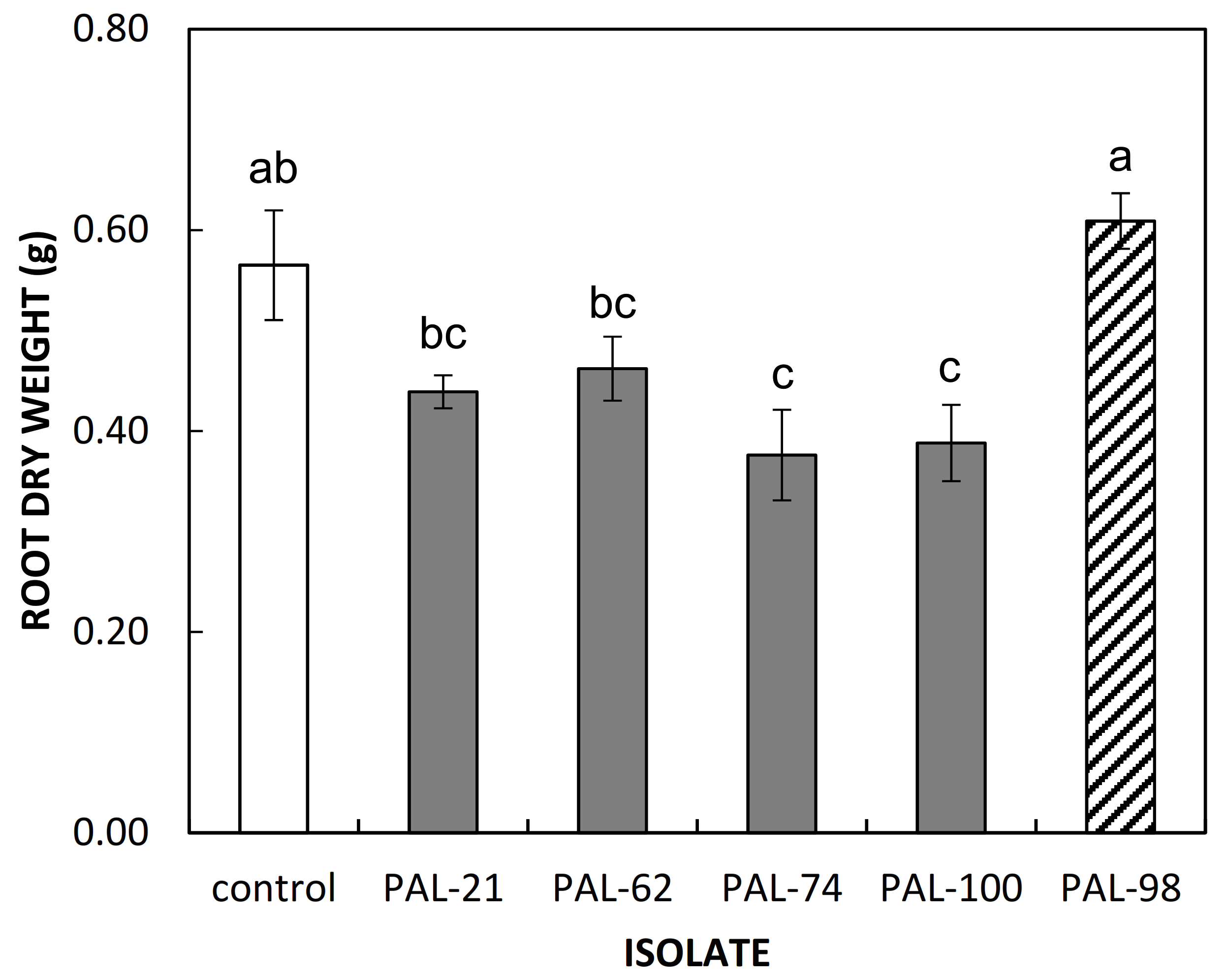
3.2. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gradziel, T.; Curtis, R.; Socias i Company, R. Production and Growing Regions. In Almonds: Botany, Production and Uses; Socias i Company, R., Gradizel, T., Eds.; CABI: Wallingford, UK, 2017; pp. 70–86. [Google Scholar]
- Felipe, A. El Almendro I: El Material Vegetal, 1st ed.; Mira Editores: Zaragoza, Spain, 2000; pp. 1–461. [Google Scholar]
- Miarnau, X.; Torguet, L.; Batlle, I.; Alegre, S. El cultivo del almendro en alta densidad. Rev. Frutic. 2016, 49, 68–87. [Google Scholar]
- Torguet, L.; Maldonado, M.; Miarnau, X. Importancia y control de las enfermedades en el cultivo del almendro. Agri. Rev. Agropec. Gan. 2019, 1026, 72–77. [Google Scholar]
- Rubio-Cabetas, M.; Felipe, A.; Reighard., G. Rootstock Development. In Almonds: Botany, Production and Uses; Rafel Socias i Company, Gradizel, T., Eds.; CABI: Wallingford, UK, 2017; pp. 209–227. [Google Scholar]
- Iglesias, I.; Carbó, J. Situació actual, característiques i comportament agronòmic dels portaempelts de presseguer. Dossier Tècnic. 2006, 17, 3–18. [Google Scholar]
- Pinochet, J. ‘Replantpac’ (Rootpac® R), a plum–almond hybrid rootstock for replant situations. HortScience 2010, 45, 299–301. [Google Scholar] [CrossRef] [Green Version]
- Teviotdale, B.; Michailides, T.; Pscheidt, J. Compendium of Nut Crop Diseases in Temperate Zones; American Phytopathological Society: Portland, OR, USA, 2002; pp. 18–429. [Google Scholar]
- Pérez-Sierra, A.; Mora-Sala, B.; León-Santana, M.; García-Jiménez, J.; Abad-Campos, P. Enfermedades causadas por Phytophthora en viveros de plantas ornamentales. Bol. Sanid. Veg. Plagas 2012, 38, 143–156. [Google Scholar]
- Wang, Y.; Tyler, B.; Wang, Y. Defense and counterdefense during plant-pathogenic oomycete infection. Annu. Rev. Microbiol. 2019, 73, 667–696. [Google Scholar] [CrossRef]
- Pinto de Torres, A.; Mircetich, S. Pythium ultimun Trow causante de pudrición del tronco en plántulas de frutales de carozo y en almendros y damascos de dos años. Agric. Téc. 1976, 36, 171–173. [Google Scholar]
- Azizi, Z.; Amini, J.; Shaykh-Eslami, M.; Abasi, S. Pathogenicity of some isolates of Pythium and Phytophthora on detached shoots and seedlings of almond. Iran. J. Plant. Pathol. 2013, 49, 33–39. [Google Scholar]
- Javadi, N.; Sharifnabi, B. Phytopythium litorale, the Causal Agent of Almond Root and Crown rot in Iran. Proceedings of 22nd Iranian Plant Protection Congress, Karaj, Iran, 27–30 August 2016; College of Agriculture and Natural Resources: Karaj, Iran, 2016; pp. 27–30. [Google Scholar]
- Browne, G.; Ott, N.; Fichtner, E. First Report of Phytopythium helicoides causing root rot on peach rootstock in California. Plant. Dis. 2019, 103, 2968. [Google Scholar] [CrossRef]
- Browne, G.; Schmidt, L.; Brar, G. First report of Phytophthora niederhauserii causing crown rot of almond (Prunus dulcis) in California. Plant. Dis. 2015, 99, 1863. [Google Scholar] [CrossRef]
- Wicks, T.; Lee, T. Phytophthora crown rot of almond trees. Aust. J. Agric. Res. 1986, 37, 277–287. [Google Scholar] [CrossRef]
- Wicks, T. Susceptibility of almond and cherry rootstocks and scions to Phytophthora species. Aust. J. Exp. Agric. 1989, 29, 103–109. [Google Scholar] [CrossRef]
- Browne, G.; Viveros, M. Diverse symptoms and tree losses caused by Phytophthora spp. in California almonds. Acta Hortic. 1997, 470, 570–575. [Google Scholar] [CrossRef]
- Sahragard, N.; Banihashemi, Z. Evaluation of resistance of some almond genotypes and cultivars to Phytophthora cactorum. Iran. J. Plant. Pathol. 2006, 42, 97–99. [Google Scholar]
- Pérez-Sierra, A.; León, M.; Álvarez, L.; Alaniz, S.; Berbegal, M.; García-Jiménez, J.; Abad-Campos, P. Outbreak of a new Phytophthora sp. associated with severe decline of almond trees in eastern Spain. Plant. Dis. 2010, 94, 534–541. [Google Scholar] [CrossRef]
- Kurbetli, I.; Değirmenci, K. First report of Phytophthora taxon niederhauserii causing decline of almond in Turkey. New Dis. Rep. 2011, 23, 14. [Google Scholar] [CrossRef] [Green Version]
- Abad, G.; Abad, J.; Cacciola, S.; Pane, A.; Faedda, R.; Moralejo, E.; Pérez-Sierra, A.; Abad-Campos, P.; Álvarez-Bernaola, L.; Bakonyi, J.; et al. Phytophthora niederhauserii sp. nov., a polyphagous species associated with ornamentals, fruit trees and native plants in 13 countries. Mycologia 2014, 106, 431–447. [Google Scholar] [CrossRef]
- Kurbetli, İ.; Yılmaz, A. Almond decline caused by Phytophthora megasperma in southeastern Anatolian region of Turkey. J. Turk. Phytopathol. 2016, 45, 13–20. [Google Scholar]
- Çiftçi, O.; Türkölmez, Ş.; Derviş, S.; Serçe, Ç. First report of canker and root rot of almond caused by Phytophthora plurivora in Turkey. Plant. Dis. 2016, 100, 1507. [Google Scholar] [CrossRef]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Ulubaş Serçe, Ç. First report of Phytophthora chlamydospora causing root and crown rot on almond (Prunus dulcis) trees in Turkey. Plant. Dis. 2016, 100, 1796. [Google Scholar] [CrossRef]
- Browne, G.; Ott, N.; Forbes, H.; Yaghmour, M.; Milliron, L. First Report of Phytophthora chlamydospora causing crown and root rot on almond in California. Plant. Dis. 2020, 104, 2033. [Google Scholar] [CrossRef]
- Jeffers, S.; Martin, S. Comparison of two media selective for Phytophthora and Pythium species. Plant. Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Erwin, D.; Ribeiro, O. Phytophthora Disease Worldwide; American Phytopathology Society Press: Saint Paul, MN, USA, 1996; pp. 1–562. [Google Scholar]
- Linderman, R.; Zeitoun, F. Phytophthora cinnamomi causing root rot and wilt of nursery-grown native western azalea and salal. Plant. Dis. Report. 1977, 61, 1045–1048. [Google Scholar]
- Collado-Romero, M.; Mercado-Blanco, J.; Olivares-García, C.; Valverde-Corredor, A.; Jiménez-Díaz, R. Molecular variability within and among Verticillium dahliae vegetative compatibility groups determined by fluorescent amplified fragment length polymorphism and polymerase chain reaction markers. Phytopathology 2006, 96, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.; Drenth, A.; Duncan, J.; Wagels, G.; Brasier, C. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungi ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: Millbrae, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; López-García, C.; León, M.; García-Jiménez, J.; Abad-Campos, P.; Jung, T. Previously unrecorded low-temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Jönsson, U.; Jung, T.; Rosengren, U.; Nihlgård, B.; Sonesson, K. Pathogenicity of Swedish isolates of Phytophthora quercina to Quercus robur in two different soils. New Phytol. 2003, 158, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.; Madden, L. Introduction to plant disease epidemiology, 1st ed; John Wiley & Sons: New York, NY, USA, 1990; pp. 192–194. [Google Scholar]
- Bielsa, B.; Rubio-Cabetas, M. La elección del patrón en la almendricultura moderna. Rev. Frut. 2018, 65, 92–113. [Google Scholar]
- Drenth, A.; Sendall, B. Practical Guide to Detection and Identification of Phytophthora; CRC for Tropical Plant Protection: Brisbane, Australia, 2001; pp. 1–41. [Google Scholar]
- Yang, J.; Ruegger, P.; McKenry, M.; Becker, J.; Borneman, J. Correlations between root-associated microorganisms and peach replant disease symptoms in a California Soil. PLoS ONE 2012, 7, e46420. [Google Scholar] [CrossRef]
- Mircetich, S. The role of Pythium in feeder roots of diseased and symptomless peach trees. Phytopathology 1971, 61, 357–360. [Google Scholar] [CrossRef]
- Langenhoven, S.; Halleen, F.; Spies, C.; Stempien, E.; Mostert, L. Detection and quantification of black foot and crown and root rot pathogens in grapevine nursery soils in the Western Cape of South Africa. Phytopathol. Mediter. 2018, 57, 519–537. [Google Scholar] [CrossRef]
- Rodríguez-Padrón, C.; Siverio, F.; Pérez-Sierra, A.; Rodríguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythium vexans recovered from avocado orchards in the Canary Islands, including Phytophthora niederhauserii as a new pathogen of avocado. Phytopathol. Mediterr. 2018, 57, 89–106. [Google Scholar] [CrossRef]
- Jabiri, S.; Lahlali, R.; Bahra, C.; Amraoui, M.; Tahiri, A.; Amiri, S. First report of Phytopythium vexans associated with dieback disease of apple trees in Morocco. J. Plant. Pathol. 2020, 102, 1319. [Google Scholar] [CrossRef]
- Ivors, K.; Abad, Z.; Benson, D. Evaluating the pathogenicity of Pythium vexans isolates from Fraser fir in North Carolina. Plant. Health Prog. 2008, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Kurbetli, İ.; Karaca, G.; Aydoğdu, M.; Sülü, G. Phytophthora species causing root and collar rot of pomegranate in Turkey. Eur. J. Plant. Pathol. 2020, 157, 485–496. [Google Scholar] [CrossRef]
- Moralejo, E.; Pérez-Sierra, A.; Álvarez, L.; Belbahri, L.; Lefort, F.; Descals, E. Multiple alien Phytophthora taxa discovered on diseased ornamental plants in Spain. Plant. Pathol. 2009, 58, 100–110. [Google Scholar] [CrossRef]
- Prigigallo, M.; Mosca, S.; Cacciola, S.; Cooke, D.; Schena, L. Molecular analysis of Phytophthora diversity in nursery-grown ornamental and fruit plants. Plant. Pathol. 2015, 64, 1308–1319. [Google Scholar] [CrossRef] [Green Version]
- Smither, M.; Jones, A. Pythium species associated with sour cherry and the effect of P. irregulare on the growth of mahaleb cherry. Can. J. Plant. Pathol. 1989, 11, 1–8. [Google Scholar] [CrossRef]
- Schmidt, L.; Browne, G. Characterization of Pythium species associated with Prunus replant disease. In Phytopathology; The American Phytopathological Society: Saint Paul, MN, USA, 2013; Volume 103, p. 128. [Google Scholar]
- McGehee, C.; Raudales, R.; Elmer, W. First report of Pythium dissotocum causing Pythium root rot on hydroponically grown lettuce in Connecticut. Plant. Dis. 2018, 102, 2043. [Google Scholar] [CrossRef]
- Owen-Going, N.; Sutton, J.; Grodzinski, B. Relationships of Pythium isolates and sweet pepper plants in single-plant hydroponic units. Can. J. Plant. Pathol. 2003, 25, 155–167. [Google Scholar] [CrossRef]
- Huo, C.; Cao, J.; Wu, K.; Chen, Y.; Zhao, Z. First Report of Pythium dissotocum causing root rot on hydroponically grown spinach in China. Plant. Dis. 2020, 104, 3084. [Google Scholar] [CrossRef]
- Bhunjun, C.; Phillips, A.; Jayawardena, R.; Promputtha, I.; Hyde, K. Importance of molecular data to identify fungal plant pathogens and guidelines for pathogenicity testing based on Koch’s Postulates. Pathogens 2021, 10, 1096. [Google Scholar] [CrossRef]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete interactions with plants: Infection strategies and resistance principles. Microbiol. Mol. Biol. Rev. 2015, 79, 263–280. [Google Scholar] [CrossRef] [Green Version]
- El Hadrami, A.; El-Bebany, A.; Yao, Z.; Adam, L.; El Hadrami, I.; Daayf, F. Plants versus fungi and oomycetes: Pathogenesis, defense and counter-defense in the proteomics era. Int. J. Mol. Sci. 2012, 13, 7237–7259. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Code | Geographic Origin | Source | Rootstock/Scion | Acc. No. |
|---|---|---|---|---|---|
| Globisporangium carolinianum | PAL-106 | Valencia | Soil | MZ921970 | |
| G. carolinianum | PAL-13 | Lleida | Soil | MZ921972 | |
| G. carolinianum | PAL-69 | Valencia | Soil | MZ922023 | |
| G. echinulatum | PAL-44 | Toledo | Soil | MZ921949 | |
| G. glomeratum | PAL-107 | Valencia | Soil | MZ921971 | |
| G. glomeratum | PAL-14 | Lleida | Soil | MZ921973 | |
| G. glomeratum | PAL-41 | Lleida | Soil | MZ922001 | |
| G. heterothallicum | PAL-42 | Toledo | Soil | MZ922002 | |
| G. hypogynum | PAL-68 | Valencia | Soil | MZ921955 | |
| G. irregulare | PAL-45 | Toledo | Soil | MZ922003 | |
| G. irregulare | PAL-46 | Toledo | Soil | MZ922004 | |
| G. irregulare | PAL-47 | Toledo | Soil | MZ922005 | |
| G. irregulare | PAL-48 | Toledo | Soil | MZ922006 | |
| G. irregulare | PAL-49 | Toledo | Soil | MZ922007 | |
| G. irregulare | PAL-51 | Toledo | Soil | MZ922009 | |
| G. irregulare | PAL-52 | Toledo | Soil | MZ922010 | |
| G. irregulare | PAL-90 | Huelva | Soil | MZ922042 | |
| G. irregulare | PAL-91 | Sevilla | Root | ‘Rootpac-40′/nd | MZ922043 |
| G. middletonii | PAL-15 | Lleida | Soil | MZ921965 | |
| G. speculum | PAL-50 | Toledo | Soil | MZ921950 | |
| G. ultimum var. ultimum | PAL-17 | Lleida | Soil | MZ921974 | |
| G. ultimum var. ultimum | PAL-58 | Valencia | Root | ‘GF-677′/’Lauranne’ | MZ922015 |
| G. ultimum var. ultimum | PAL-61 | Valencia | Root | ‘GF-677′/’Lauranne’ | MZ922017 |
| G. ultimum var. ultimum | PAL-63 | Valencia | Soil | MZ922018 | |
| G. ultimum var. ultimum | PAL-79 | Huelva | Soil | MZ922030 | |
| G. ultimum var. ultimum | PAL-84 | Huelva | Soil | MZ922035 | |
| G. ultimum var. ultimum | PAL-92 | Sevilla | Soil | MZ922044 | |
| G. ultimum var. ultimum | PAL-94 | Sevilla | Soil | MZ922046 | |
| Phytophthora cactorum | PAL-10 | Lleida | Soil | MZ921975 | |
| Ph. cactorum | PAL-12 | Lleida | Soil | MZ921977 | |
| Ph. cactorum | PAL-24 | Lleida | Soil | MZ921983 | |
| Ph. cactorum | PAL-28 | Lleida | Soil | MZ921984 | |
| Ph. cactorum | PAL-35 | Lleida | Soil | MZ921994 | |
| Ph. cactorum | PAL-38 | Lleida | Soil | MZ921997 | |
| Ph. cactorum | PAL-4 | Lleida | Soil | MZ921999 | |
| Ph. citrophthora | PAL-16 | Lleida | Soil | MZ921978 | |
| Ph. citrophthora | PAL-18 | Lleida | Soil | MZ921979 | |
| Ph. citrophthora | PAL-19 | Lleida | Soil | MZ921980 | |
| Ph. citrophthora | PAL-2 | Lleida | Soil | MZ921981 | |
| Ph. citrophthora | PAL-20 | Lleida | Soil | MZ921982 | |
| Ph. humicola/condilina | PAL-5 | Lleida | Soil | MZ922008 | |
| Ph. nicotianae | PAL-57 | Valencia | Soil | MZ922014 | |
| Ph. nicotianae | PAL-71 | Valencia | Soil | MZ922025 | |
| Ph. niederhauserii | PAL-11 | Lleida | Soil | MZ921964 | |
| Ph. niederhauserii | PAL-34 | Lleida | Soil | MZ921969 | |
| Ph. niederhauserii | PAL-60 | Valencia | Root | ‘GF-677′/’Lauranne’ | MZ921953 |
| Ph. niederhauserii | PAL-62 | Valencia | Root | ‘GF-677′/’Lauranne’ | MZ921954 |
| Ph. niederhauserii | PAL-1 | Lleida | Soil | MZ921961 | |
| Ph. niederhauserii | PAL-100 | Córdoba | Root | ‘GF-677′/nd | MZ921962 |
| Ph. niederhauserii | PAL-105 | Valencia | Root | ‘Garnem/’Lauranne’ | MZ921963 |
| Ph. niederhauserii | PAL-21 | Lleida | Soil | MZ921966 | |
| Ph. niederhauserii | PAL-26 | Lleida | Soil | MZ921967 | |
| Ph. niederhauserii | PAL-3 | Lleida | Soil | MZ921968 | |
| Ph. niederhauserii | PAL-6 | Lleida | Soil | MZ921952 | |
| Ph. niederhauserii | PAL-7 | Lleida | Soil | MZ921956 | |
| Ph. niederhauserii | PAL-74 | Valencia | Soil | MZ921957 | |
| Ph. niederhauserii | PAL-77 | Valencia | Soil | MZ921958 | |
| Ph. niederhauserii | PAL-78 | Valencia | Root | ‘GF-677′/nd | MZ921959 |
| Ph. niederhauserii | PAL-8 | Lleida | Soil | MZ921960 | |
| Ph. palmivora | PAL-66 | Valencia | Soil | MZ922021 | |
| Ph. palmivora | PAL-72 | Valencia | Soil | MZ922026 | |
| Ph. tropicalis | PAL-101 | Córdoba | Soil | MZ921976 | |
| Phytopythium chamaehyphon | PAL-99 | Córdoba | Soil | MZ922051 | |
| Pp. cucurbitacearum | PAL-53 | Valencia | Root | ‘Garnem’/nd | MZ921951 |
| Pp. helicoides | PAL-76 | Valencia | Root | ‘Garnem’/’Penta’ | MZ922029 |
| Pp. helicoides | PAL-96 | Sevilla | Root | ‘GF-677′/nd | MZ922048 |
| Pp. mercuriale | PAL-86 | Huelva | Soil | MZ922037 | |
| Pp. mercuriale | PAL-87 | Huelva | Soil | MZ922038 | |
| Pp. vexans | PAL-22 | Lleida | Soil | MZ921985 | |
| Pp. vexans | PAL-23 | Lleida | Root | ‘GF-677′/’Constantí’ | MZ921986 |
| Pp. vexans | PAL-25 | Lleida | Root | ‘GF-677′/’Vairo’ | MZ921987 |
| Pp. vexans | PAL-27 | Lleida | Root | ‘GF-677′/’Marinada’ | MZ921988 |
| Pp. vexans | PAL-29 | Lleida | Root | ‘Garnem’/’Soleta’ | MZ921989 |
| Pp. vexans | PAL-32 | Lleida | Soil | MZ921992 | |
| Pp. vexans | PAL-33 | Lleida | Soil | MZ921993 | |
| Pp. vexans | PAL-36 | Lleida | Soil | MZ921995 | |
| Pp. vexans | PAL-37 | Lleida | Soil | MZ921996 | |
| Pp. vexans | PAL-40 | Lleida | Soil | MZ922000 | |
| Pp. vexans | PAL-59 | Valencia | Root | ‘GF-677′/’Lauranne’ | MZ922016 |
| Pp. vexans | PAL-70 | Valencia | Soil | MZ922024 | |
| Pp. vexans | PAL-73 | Valencia | Soil | MZ922027 | |
| Pp. vexans | PAL-75 | Valencia | Soil | MZ922028 | |
| Pp. vexans | PAL-82 | Huelva | Soil | MZ922033 | |
| Pp. vexans | PAL-83 | Huelva | Soil | MZ922034 | |
| Pp. vexans | PAL-85 | Huelva | Soil | MZ922036 | |
| Pp. vexans | PAL-88 | Huelva | Soil | MZ922039 | |
| Pp. vexans | PAL-89 | Huelva | Soil | MZ922040 | |
| Pp. vexans | PAL-9 | Lleida | Soil | MZ922041 | |
| Pp. vexans | PAL-93 | Sevilla | Soil | MZ922045 | |
| Pp. vexans | PAL-98 | Córdoba | Root | ‘GF-677′/nd | MZ922050 |
| Pythium aphanidermatum | PAL-39 | Lleida | Soil | MZ921998 | |
| Py. dissotocum | PAL-30 | Lleida | Soil | MZ921990 | |
| Py. dissotocum | PAL-31 | Lleida | Soil | MZ921991 | |
| Py. dissotocum | PAL-54 | Valencia | Soil | MZ922011 | |
| Py. dissotocum | PAL-55 | Valencia | Soil | MZ922012 | |
| Py. dissotocum | PAL-65 | Valencia | Soil | MZ922020 | |
| Py. dissotocum | PAL-80 | Huelva | Soil | MZ922031 | |
| Py. dissotocum | PAL-81 | Huelva | Soil | MZ922032 | |
| Py. dissotocum | PAL-95 | Sevilla | Soil | MZ922047 | |
| Py. dissotocum | PAL-97 | Sevilla | Soil | MZ922049 | |
| Py. nodosum | PAL-56 | Valencia | Soil | MZ922013 | |
| Py. nodosum | PAL-64 | Valencia | Soil | MZ922019 | |
| Py. nodosum | PAL-67 | Valencia | Soil | MZ922022 | |
| Py. pachycaule | PAL-43 | Toledo | Soil | MZ921948 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beluzán, F.; Miarnau, X.; Torguet, L.; Armengol, J.; Abad-Campos, P. Survey of Oomycetes Associated with Root and Crown Rot of Almond in Spain and Pathogenicity of Phytophthora niederhauserii and Phytopythium vexans to ‘Garnem’ Rootstock. Agriculture 2022, 12, 294. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020294
Beluzán F, Miarnau X, Torguet L, Armengol J, Abad-Campos P. Survey of Oomycetes Associated with Root and Crown Rot of Almond in Spain and Pathogenicity of Phytophthora niederhauserii and Phytopythium vexans to ‘Garnem’ Rootstock. Agriculture. 2022; 12(2):294. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020294
Chicago/Turabian StyleBeluzán, Francisco, Xavier Miarnau, Laura Torguet, Josep Armengol, and Paloma Abad-Campos. 2022. "Survey of Oomycetes Associated with Root and Crown Rot of Almond in Spain and Pathogenicity of Phytophthora niederhauserii and Phytopythium vexans to ‘Garnem’ Rootstock" Agriculture 12, no. 2: 294. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020294