
Effects of Solar Radiation on Dry Matter Distribution and Root Morphology of High Yielding Maize Cultivars

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
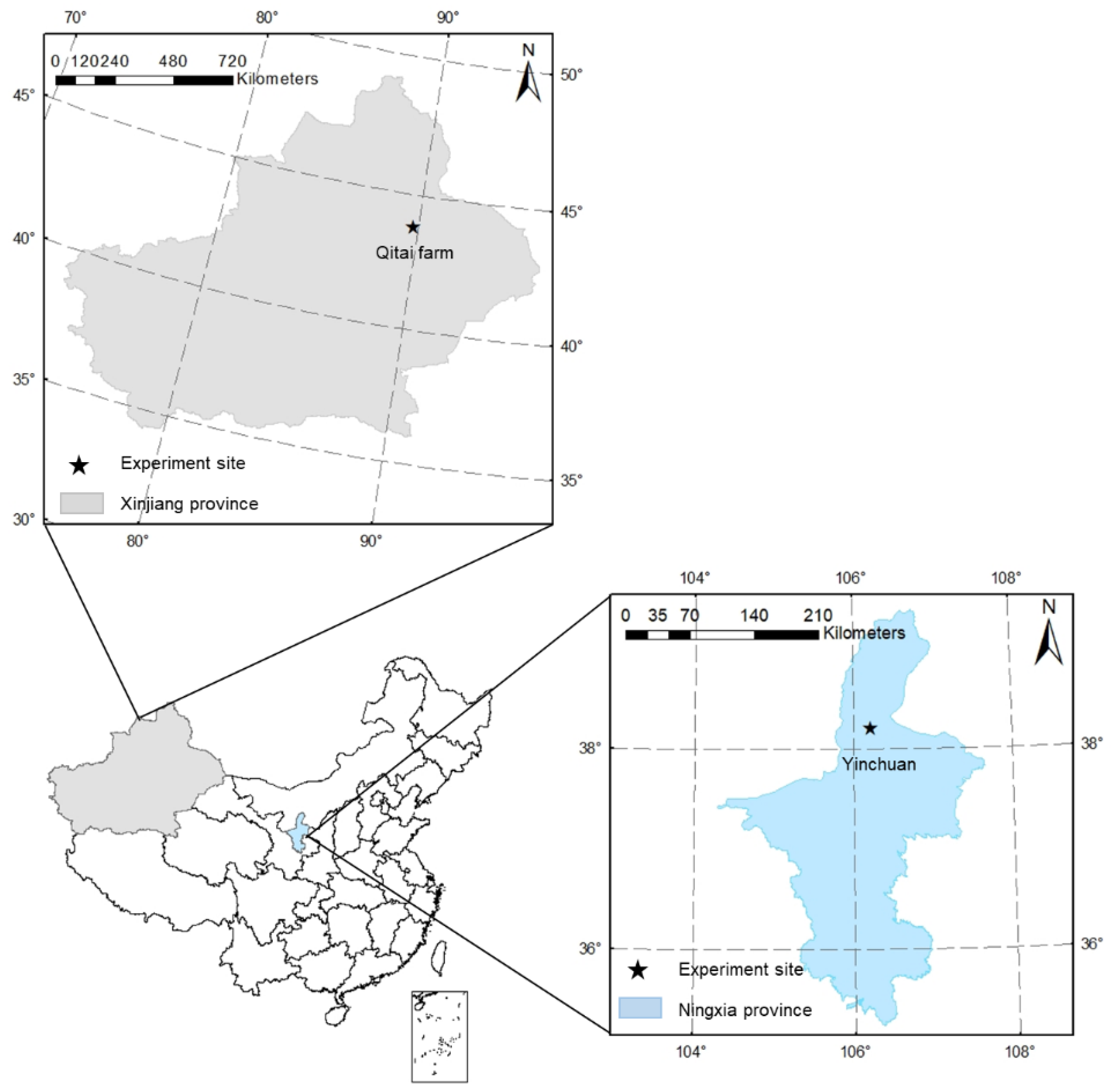
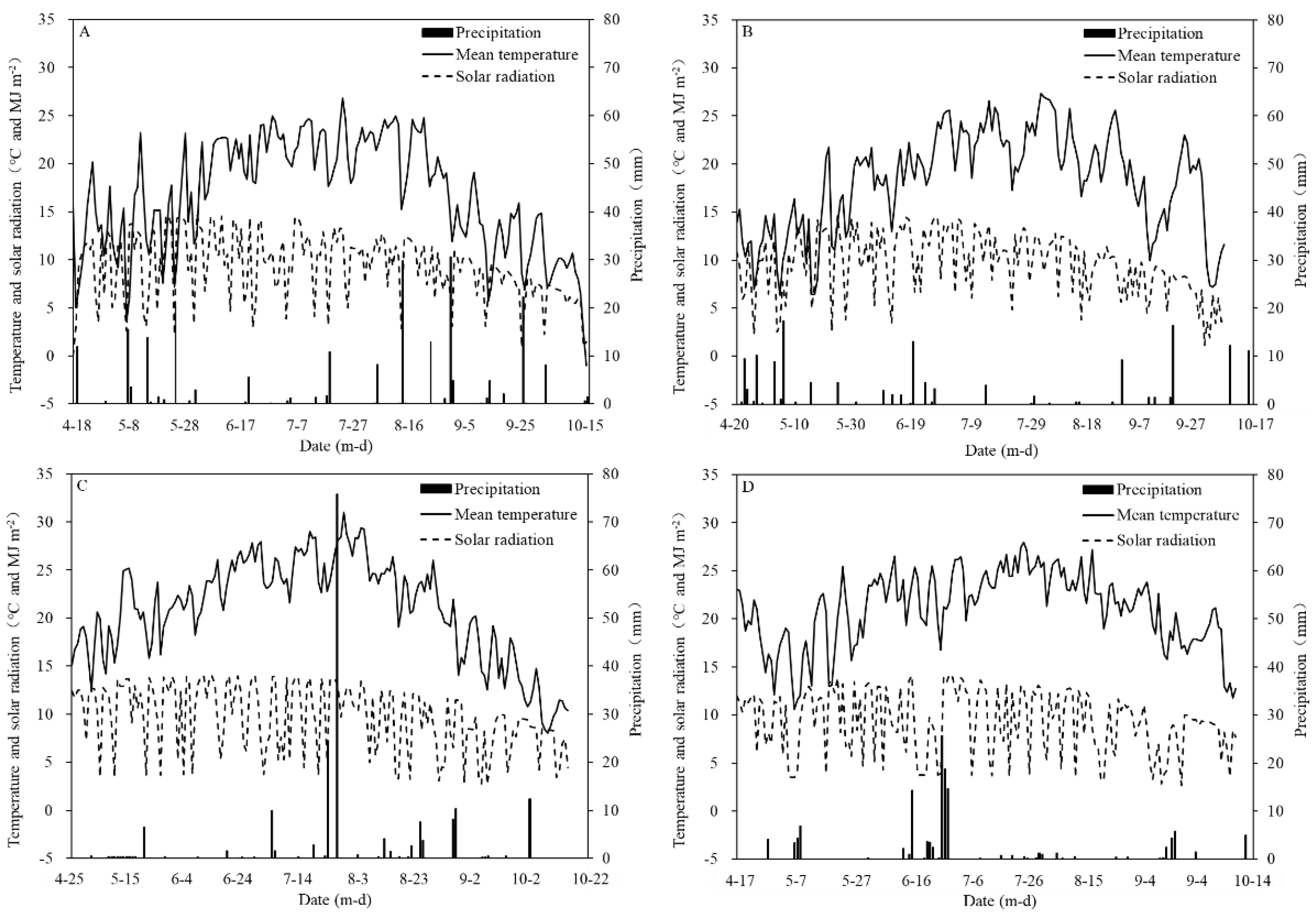
2.1. Site Description and Weather Data
2.2. Experiment Design
2.3. Field Management
2.3.1. Total Intercepted Photosynthetically Active Radiation
2.3.2. Root Sampling
2.4. Statistical Analysis
3. Results
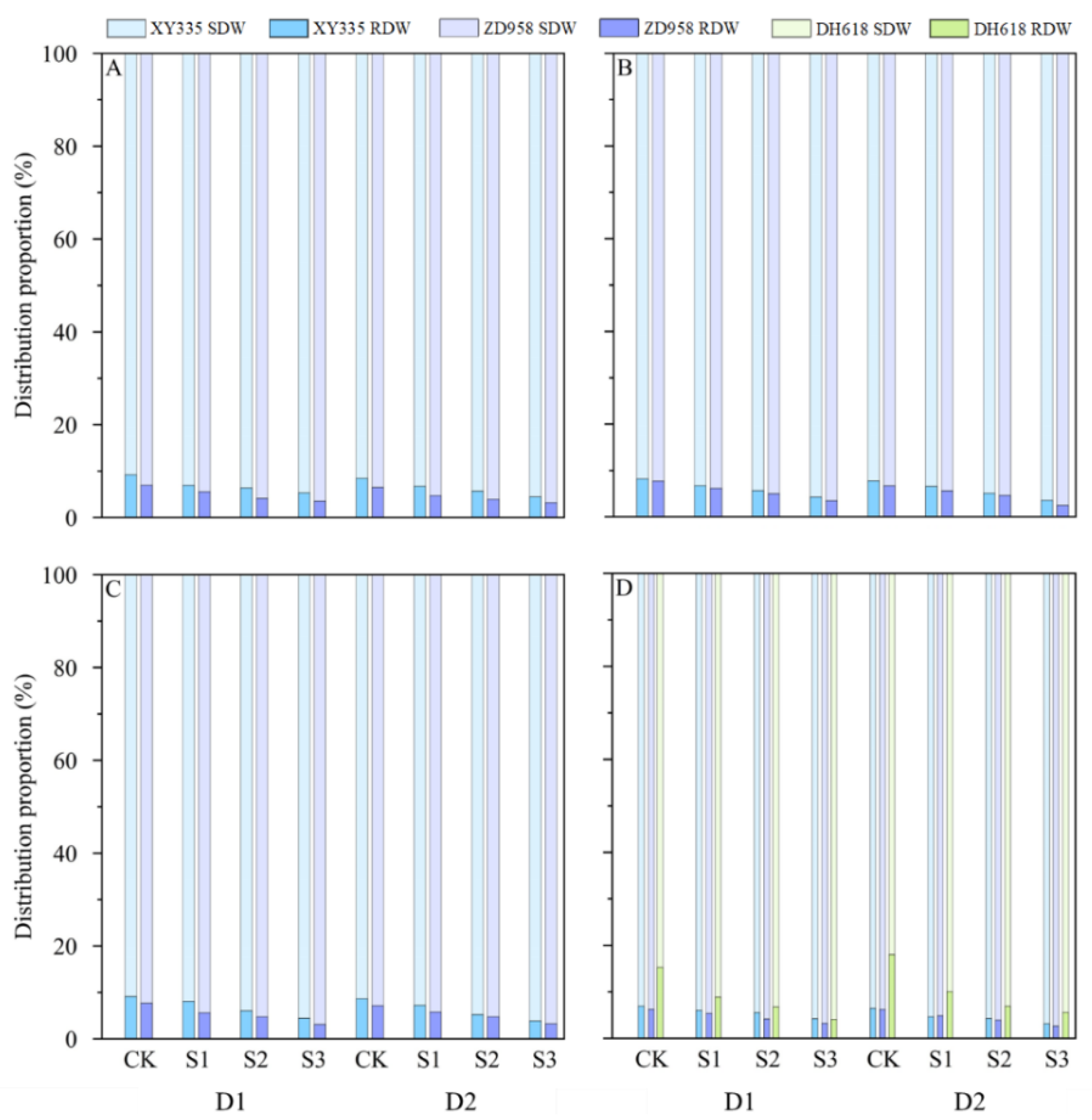
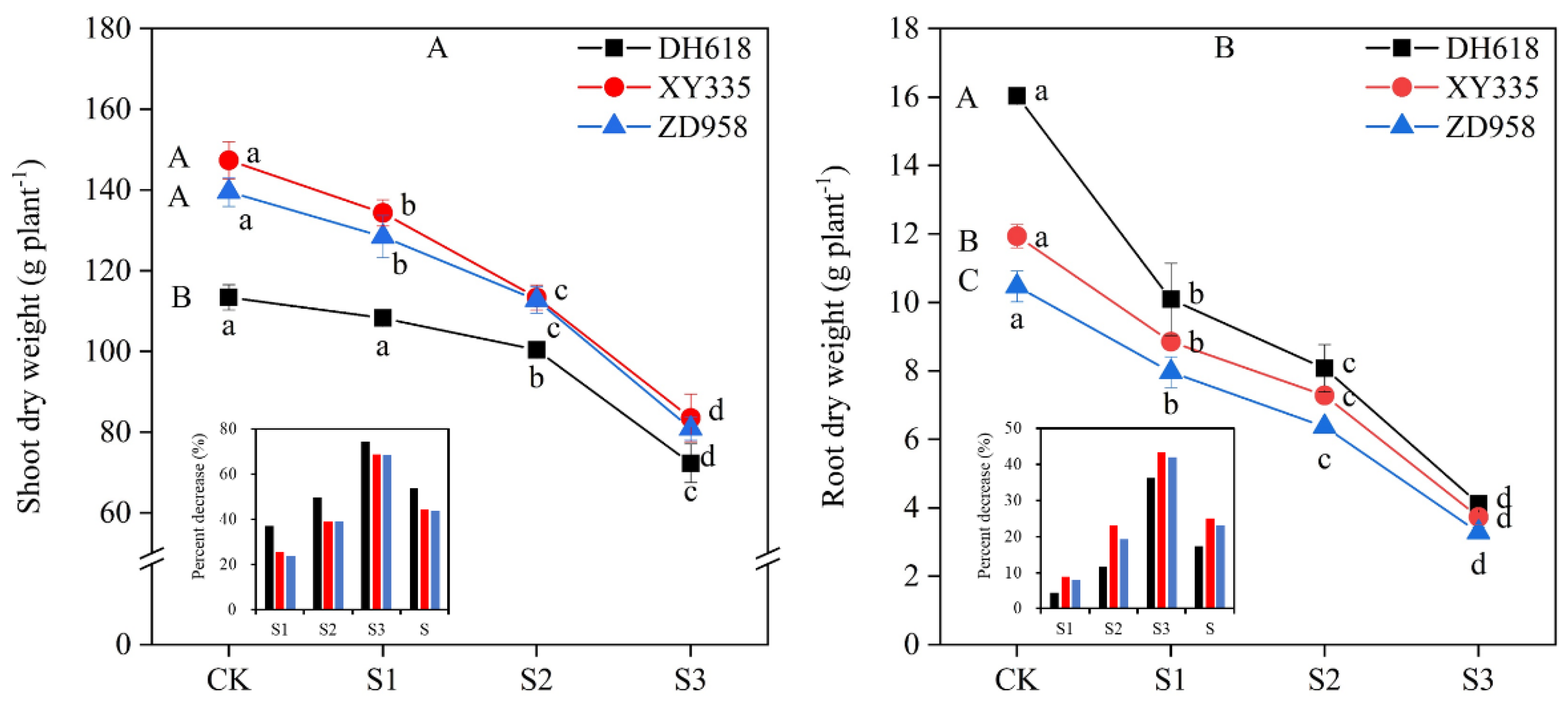
3.1. Effects of Different Shading Levels on Maize Dry Matter Distribution
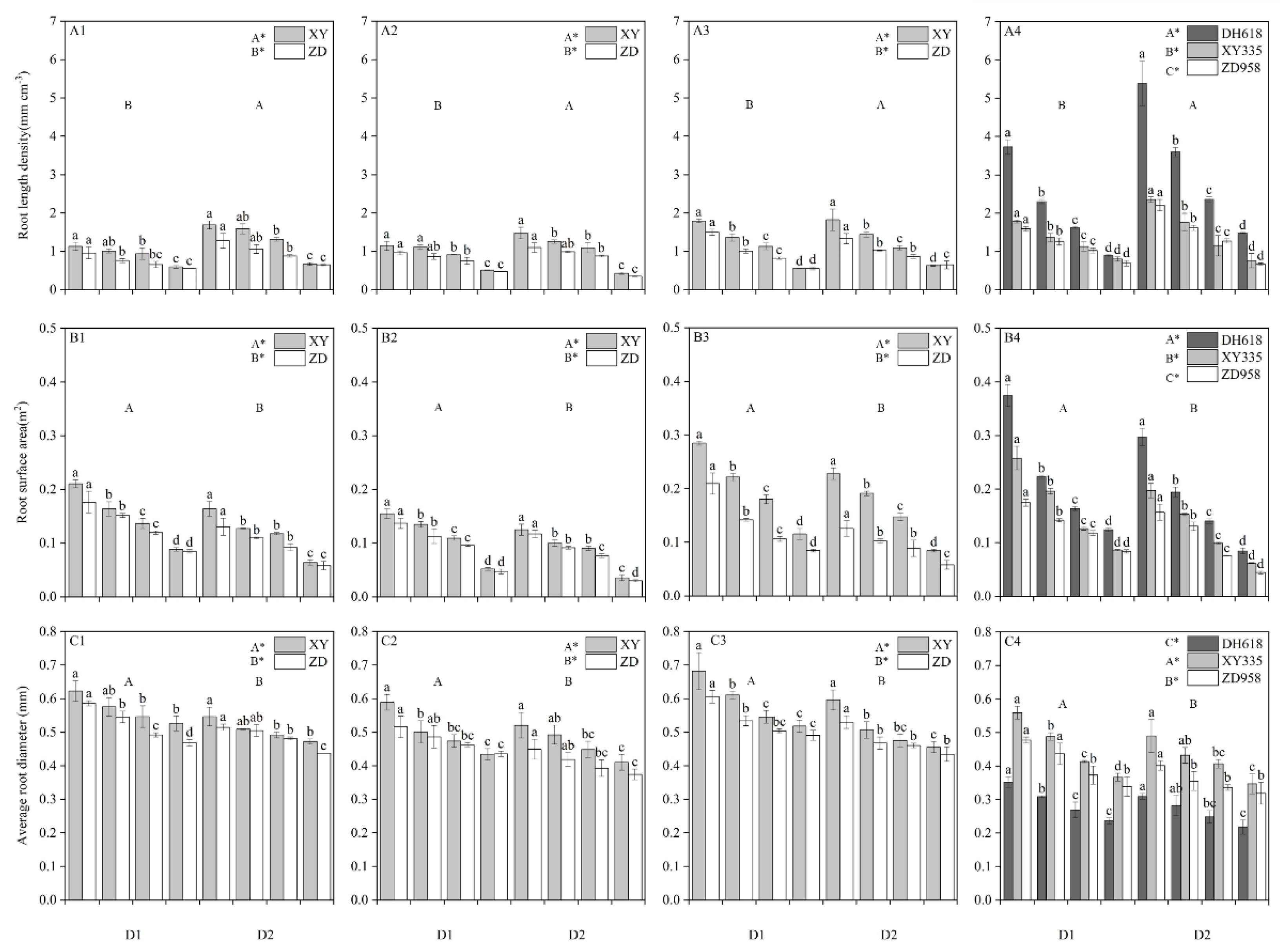
3.2. Effects of Different Shading Levels on Maize Root Morphology
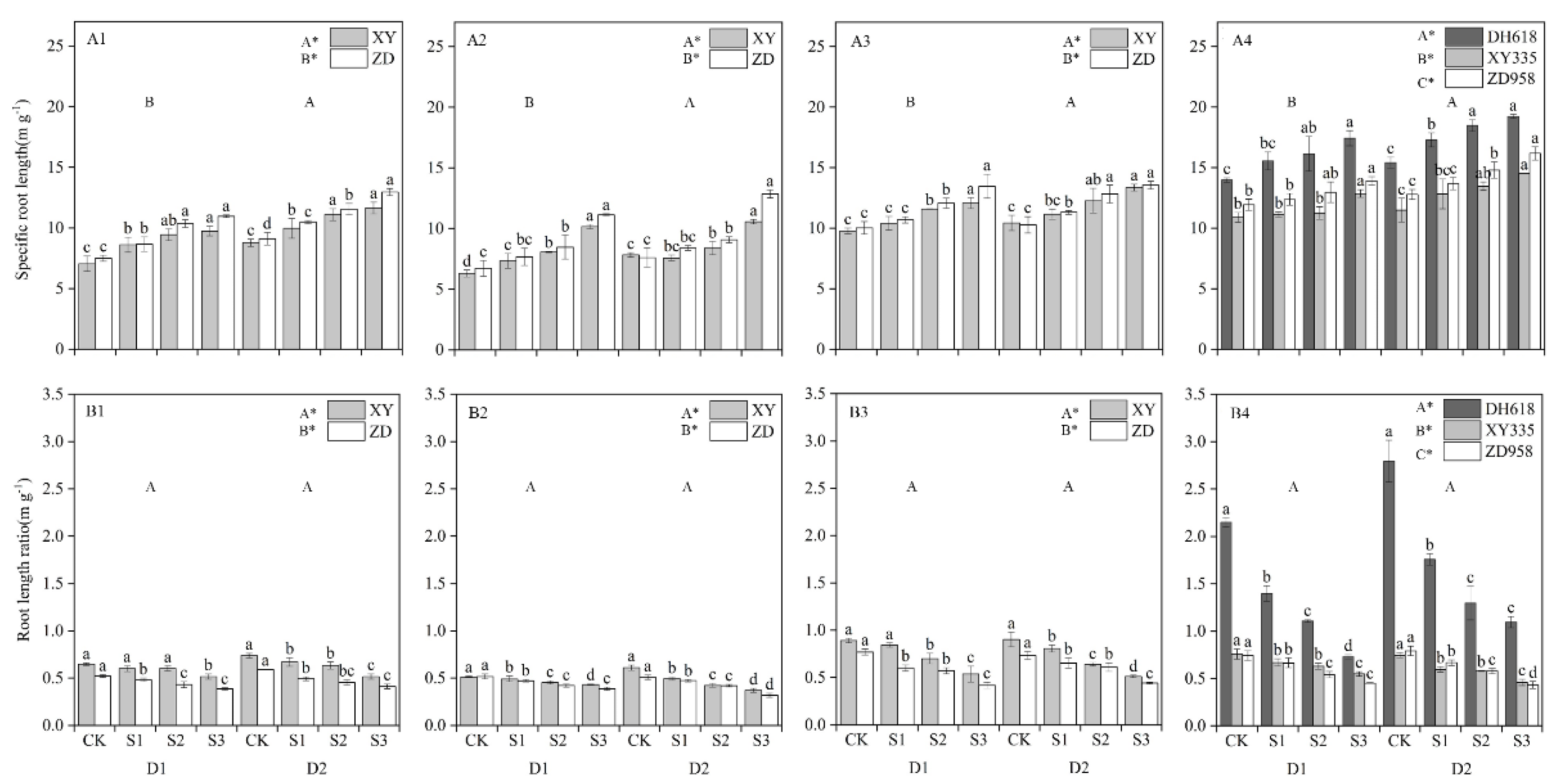
3.3. Effects of Different Shading Levels on the Acquisition Ability of Root
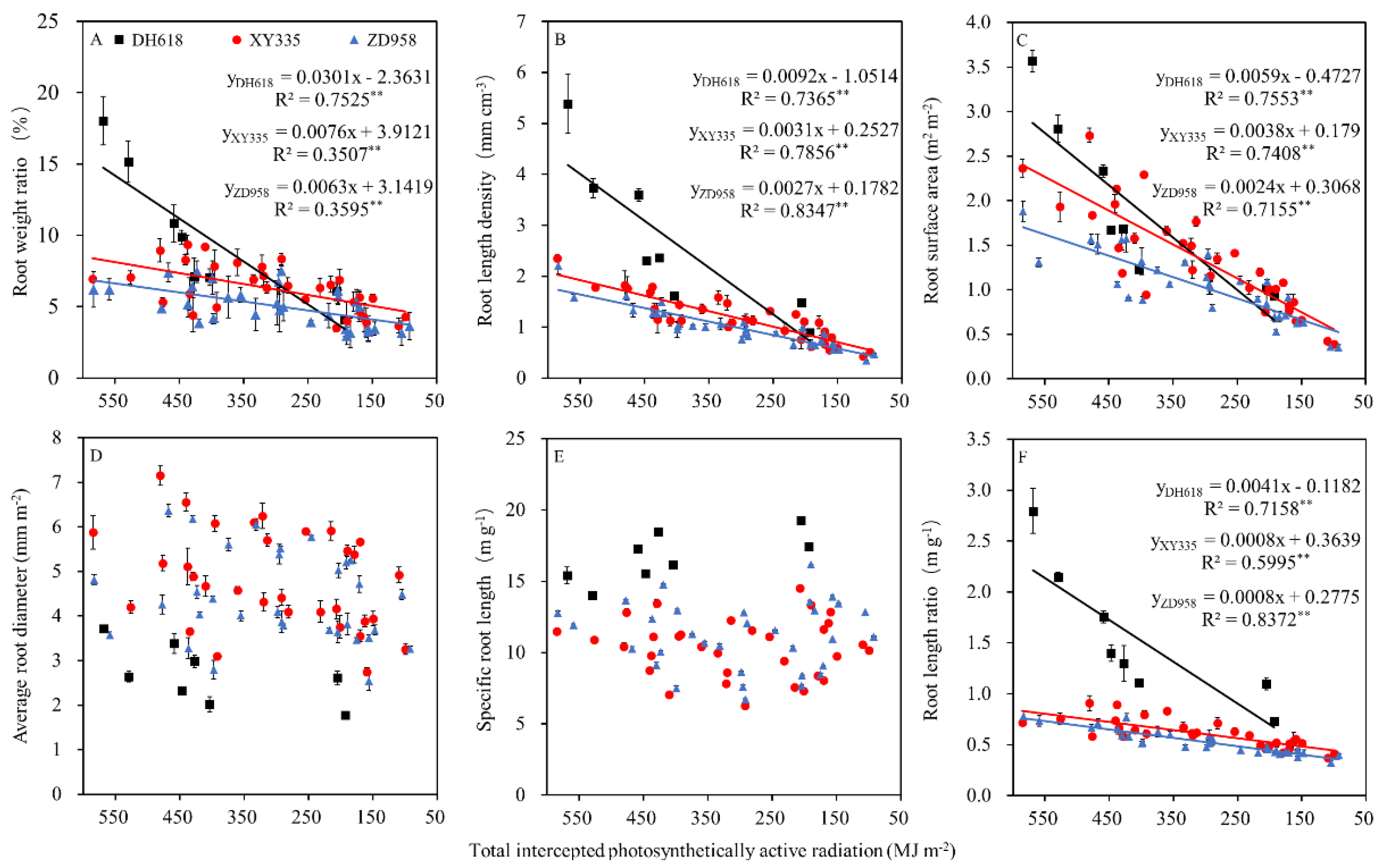
3.4. Quantitative Relationships between TIPAR and Root Morphology
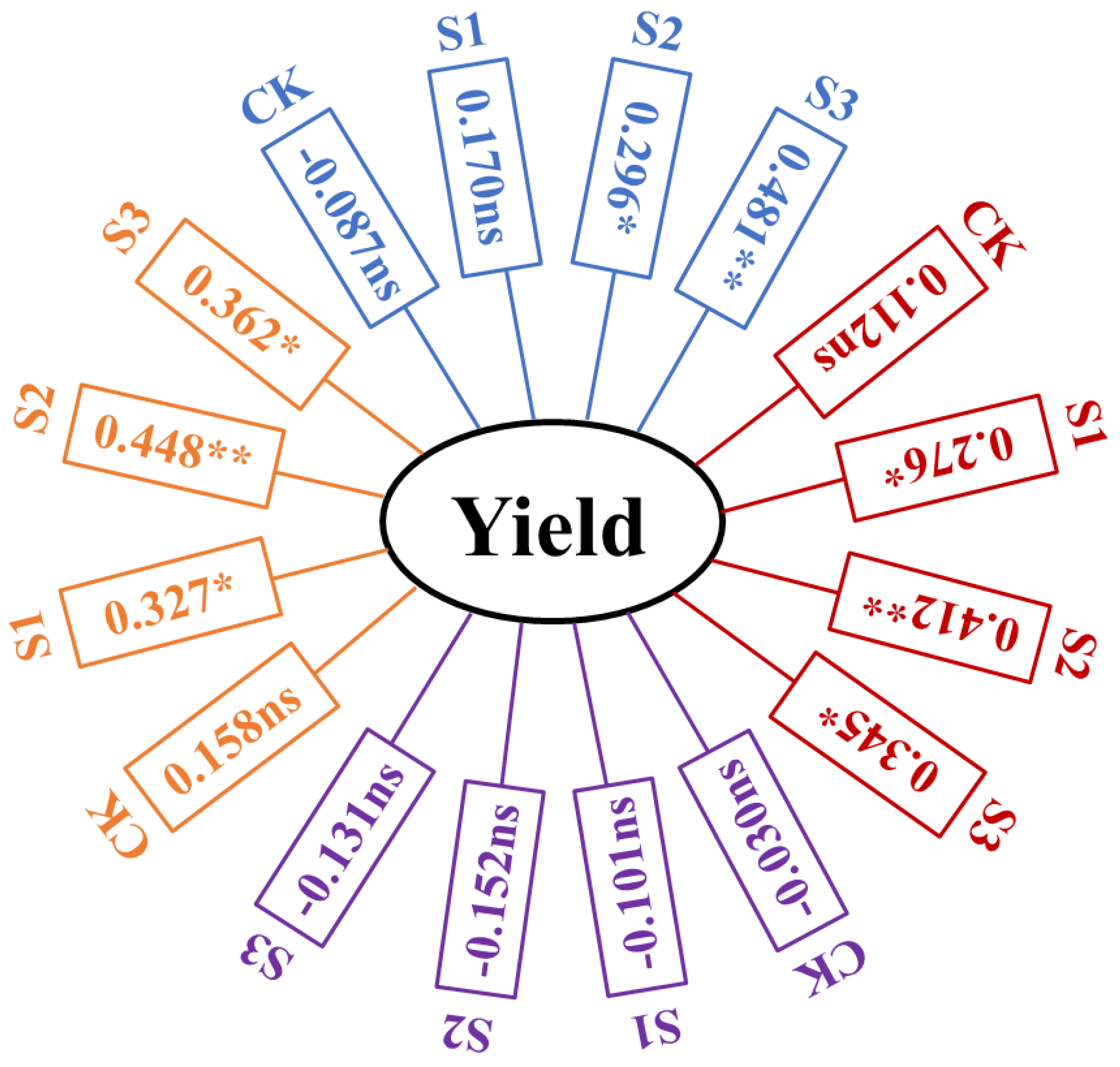
3.5. Effects of Different Root Morphology on Maize Yield
4. Discussion
4.1. Effect of Solar Radiation on Maize Root and Shoot
4.2. Effect of Root Morphology on Yield
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Richards, R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-S.; Guo, X.-X.; Liu, H.-F.; Liu, G.-Z.; Liu, W.-M.; Ming, B.; Xie, R.-Z.; Wang, K.-R.; Hou, P.; Li, S.-K. The effect of solar radiation change on the maize yield gap from the perspectives of dry matter accumulation and distribution. J. Integr. Agric. 2021, 20, 482–493. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, X.X.; Liu, G.Z.; Liu, W.M.; Xue, J.; Ming, B.; Xie, R.Z.; Wang, K.R.; Hou, P.; Li, S.K. Solar radiation effects on dry matter accumulations and transfer in maize. Front. Plant Sci. 2021, 12, 727134. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Liu, Y.E.; Liu, W.M.; Yang, H.S.; Xie, R.Z.; Wang, K.R.; Ming, B.; Liu, G.Z.; Xue, J.; Wang, Y.H.; et al. Quantifying maize grain yield losses caused by climate change based on extensive field data across China. Resour. Conserv. Recy. 2021, 174, 105811. [Google Scholar] [CrossRef]
- Wu, A.; Hammer, G.L.; Doherty, A.; Von Caemmerer, S.; Farquhar, G. Quantifying impacts of enhancing photosynthesis on crop yield. Nat. Plants 2019, 5, 380–388. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, M.A. Prospects of photosynthetic research for increasing agricultural productivity, with emphasis on the tropical C4 Amaranthus and the cassava C3-C4 crops. Photosynthetica 2016, 54, 161–184. [Google Scholar] [CrossRef]
- Jansson, C.; Vogel, J.; Hazen, S.; Brutnell, T.; Mockler, T. Climate-smart crops with enhanced photosynthesis. J. Exp. Bot. 2018, 69, 3801–3809. [Google Scholar] [CrossRef]
- Hu, Q.; Ma, X.; He, H.; Pan, F.; He, Q.; Huang, B.; Pan, X. Warming and Dimming: Interactive Impacts on Potential Summer Maize Yield in North China Plain. Sustainability 2019, 11, 2588. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Liu, B.; Yang, H.; Chen, X. Solar dimming decreased maize yield potential on the North China Plain. Food Energy Secur. 2020, 9, e235. [Google Scholar] [CrossRef]
- Zhao, J.; Kong, X.; He, K.; Xu, H.; Mu, J. Assessment of the radiation effect of aerosols on maize production in China. Sci. Total Environ. 2020, 720, 137567. [Google Scholar] [CrossRef]
- Abakumova, G.M.; Gorbarenko, E.V.; Nezval, E.I.; Shilovtseva, O.A. Fifty years of actinometrical measurements in Moscow. Int. J. Remote Sens. 2008, 29, 2629–2665. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, B.; Dong, S.; Liu, P.; Ren, B.; Zhang, J. Response of Summer Maize Photosynthate Accumulation and Distribution to Shading Stress Assessed by Using 13CO2 Stable Isotope Tracer in the Field. Front. Plant Sci. 2017, 8, 1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.W.; Dong, S.T.; Wang, K.J.; Hu, C.H.; Liu, P. Effects of shading in field on key enzymes involved in starch synthesis of summer maize. Acta Agronica Sinica 2008, 34, 1470–1474. [Google Scholar] [CrossRef]
- Liu, W.D.; Matthijs, T. Physiological mechanisms underlying heterosis for shade tolerance in maize. Crop Sci. 2009, 49, 1817–1826. [Google Scholar] [CrossRef]
- Ren, B.; Cui, H.; Camberato, J.J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Effects of shading on the photosynthetic characteristics and mesophyll cell ultrastructure of summer maize. Die Nat. 2016, 103, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.-Y.; Jin, L.-B.; Li, B.; Dong, S.-T.; Liu, P.; Zhao, B.; Zhang, J.-W. Effects of shading on dry matter accumulation and nutrient absorption of summer maize. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2013, 24, 3099–3105. [Google Scholar]
- Qian, C.-J.; Zhang, W.; Zhong, X.-M.; Li, F.-H.; Shi, Z.-S. Comparative studies on the photosynthetic characteristics of two maize (Zea mays L.) near-isogenic lines differing in their susceptibility to low light intensity. Emir. J. Food Agric. 2017, 1, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.M.; Shi, Z.S.; Li, F.H.; Huang, H.J. Photosynthesis and chlorophyll fluorescence of infertile and fertile stalks of paired near-isogenic lines in maize (Zea mays L.) under shade conditions. Photosynthetica 2014, 52, 597–603. [Google Scholar] [CrossRef]
- Yang, H.; Shi, Y.; Xu, R.; Lu, D.; Lu, W. Effects of shading after pollination on kernel filling and physicochemical quality traits of waxy maize. Crop J. 2016, 4, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Shi, J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Grain development and endogenous hormones in summer maize (Zea mays L.) submitted to different light conditions. Int. J. Biometeorol. 2018, 62, 2131–2138. [Google Scholar] [CrossRef]
- Xue, J.; Gou, L.; Zhao, Y.; Yao, M.; Yao, H.; Tian, J.; Zhang, W. Effects of light intensity within the canopy on maize lodging. Field Crops Res. 2016, 188, 133–141. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, X.X.; Hou, P.; Xue, J.; Liu, G.Z.; Liu, W.M.; Wang, Y.H.; Zhao, R.L.; Ming, B.; Xie, R.Z.; et al. Quantitative effects of solar radiation on maize lodging resistance mechanical properties. Field Crops Res. 2020, 255, 107906. [Google Scholar] [CrossRef]
- Eapen, D.; Barroso, M.L.; Ponce, G.; Campos, M.E.; Cassab, G.I. Hydrotropism: Root growth responses to water. Trends Plant Sci. 2005, 10, 44–50. [Google Scholar] [CrossRef]
- Hardtke, C.S.; Villalobos, D.P. The Brachypodium distachyon Root System: A Tractable Model to Investigate Grass Roots. In Genetics and Genomics of Brachypodium; Plant Genetics and Genomics: Crops and Models; Vogel, J.P., Ed.; Springer: Cham, Switzerland, 2015; Volume 18, pp. 245–258. [Google Scholar]
- Singh, V.; Oosterom, E.J.V.; Jordan, D.R.; Messina, C.D.; Cooper, M.; Hammer, G.L. Morphological and architectural development of root systems in sorghum and maize. Plant Soil 2010, 333, 287–299. [Google Scholar] [CrossRef]
- Takehisa, H.; Sato, Y.; Igarashi, M.; Abiko, T.; Antonio, B.A.; Kamatsuki, K.; Minami, H.; Namiki, N.; Inukai, Y.; Nakazono, M.; et al. Genome-wide transcriptome dissection of the rice root system: Implications for developmental and physiological functions. Plant J. 2011, 69, 126–140. [Google Scholar] [CrossRef]
- Berry, P.; Baker, C.; Hatley, D.; Dong, R.; Wang, X.; Blackburn, G.; Miao, Y.; Sterling, M.; Whyatt, J. Development and application of a model for calculating the risk of stem and root lodging in maize. Field Crops Res. 2020, 262, 108037. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 265, 6155–6166. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef]
- de Dorlodot, S.; Forster, B.; Pages, L.; Price, A.; Tuberosa, R.; Draye, X. Root system architecture: Opportunities and constraints for genetic improvement of crops. Trends Plant Sci. 2007, 12, 474–481. [Google Scholar] [CrossRef]
- Gowda, V.R.; Henry, A.; Yamauchi, A.; Shashidhar, H.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. Field Crops Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Maurel, C.; Simonneau, T.; Sutka, M. The significance of roots as hydraulic rheostats. J. Exp. Bot. 2010, 61, 3191–3198. [Google Scholar] [CrossRef] [Green Version]
- Franco, J.A.; Bañón, S.; Vicente, M.M.J.; Miralles, J.; Martínez-Sánchez, J.J. Review Article: Root development in horticultural plants grown under abiotic stress conditions—A review. J. Hortic. Sci. Biotech. 2011, 86, 543–556. [Google Scholar] [CrossRef]
- Guo, X.; Yang, Y.; Liu, H.; Liu, G.; Liu, W.; Wang, Y.; Zhao, R.; Ming, B.; Xie, R.; Wang, K.; et al. Effects of solar radiation on root and shoot growth of maize and the quantitative relationship between them. Crop Sci. 2020, 61, 1414–1425. [Google Scholar] [CrossRef]
- Hund, A.; Richner, W.; Soldati, A.; Fracheboud, Y.; Stamp, P. Root morphology and photosynthetic performance of maize inbred lines at low temperature. Eur. J. Agron. 2007, 27, 52–61. [Google Scholar] [CrossRef]
- Zou, H.Y.; Zhang, F.C.; Wu, L.F.; Xiang, Y.Z.; Fan, J.L.; Li, Z.J.; Li, S.E. Normalized root length density distribution model for spring maize under different water and fertilizer combination. Trans. Chin. Soc. Agric. Eng. 2018, 34, 133–142. [Google Scholar]
- Tuberosa, R.; Salvi, S.; Sanguineti, M.C.; Maccaferri, M.; Giuliani, S.; Landi, P. Searching for quantitative trait loci controlling root traits in maize: A critical appraisal. Plant Soil 2003, 255, 35–54. [Google Scholar] [CrossRef]
- Mu, X.H.; Chen, F.J.; Wu, Q.P.; Chen, Q.W.; Wang, J.F.; Yuan, L.X.; Mi, G.H. Genetic improvement of root growth in-creases maize yield via enhanced post-silking nitrogen uptake. Eur. J. Agron. 2015, 63, 55–61. [Google Scholar] [CrossRef]
- Zhang, F.L.; Niu, X.K.; Zhang, Y.M.; Xie, R.Z.; Liu, X.; Li, S.K.; Gao, S.J. Studies on the root characteristics of maize varieties of different eras. J. Integr. Agric. 2013, 12, 426–435. [Google Scholar] [CrossRef]
- Shao, H.; Xia, T.T.; Wu, D.L.; Chen, F.J.; Mi, G.H. Root growth and root system architecture of field-grown maize in response to high planting density. Plant Soil 2018, 430, 395–411. [Google Scholar] [CrossRef]
- Liu, G.; Hou, P.; Xie, R.; Ming, B.; Wang, K.; Xu, W.; Liu, W.; Yang, Y.; Li, S. Canopy characteristics of high-yield maize with yield potential of 22.5 Mg ha−1. Field Crops Res. 2017, 213, 221–230. [Google Scholar] [CrossRef]
- Liu, G.; Yang, H.; Xie, R.; Yang, Y.; Liu, W.; Guo, X.; Xue, J.; Ming, B.; Wang, K.; Hou, P.; et al. Genetic gains in maize yield and related traits for high-yielding cultivars released during 1980s to 2010s in China. Field Crops Res. 2021, 270, 108223. [Google Scholar] [CrossRef]
- Vazin, F.; Hassanzadeh, M.; Madani, A.; Nassiri-Mahallati, M.; Nasri, M. Modeling light interception and distribution in mixed canopy of common cocklebur (Xanthium stramarium) in competition with corn. Planta Daninha 2010, 28, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, Y.; Shi, Y.; Song, Y.; Zhang, D.; Buckler, E.; Zhang, Z.; Wang, T.; Li, Y. Genetic Control of the Leaf Angle and Leaf Orientation Value as Revealed by Ultra-High Density Maps in Three Connected Maize Populations. PLoS ONE 2015, 10, e0121624. [Google Scholar] [CrossRef]
- Hou, P.; Liu, Y.; Liu, W.; Liu, G.; Xie, R.; Wang, K.; Ming, B.; Wang, Y.; Zhao, R.; Zhang, W.; et al. How to increase maize production without extra nitrogen input. Resour. Conserv. Recycl. 2020, 160, 104913. [Google Scholar] [CrossRef]
- Purcell, L.C.; Ball, R.A.; Reaper, J.D.; Vories, E.D. Radiation use efficiency and biomass production in soybean at different plant population densities. Crop Sci. 2002, 42, 172–177. [Google Scholar] [CrossRef]
- Mao, L.L.; Zhang, L.Z.; Zhao, X.H.; Liu, S.D.; van der Werf, W.; Zhang, S.P.; Spiertz, H.; Li, Z.H. Crop growth, light utilization and yield of relay intercropped cotton as affected by plant density and a plant growth regulator. Field Crops Res. 2014, 155, 67–76. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, J.; Bai, W.; Zhang, Y.; Zhang, W.H. The response of root traits to precipitation change of herbaceous species in temperate steppes. Funct. Ecol. 2019, 33, 2030–2041. [Google Scholar] [CrossRef]
- Gao, Q.; Li, M.; Pu, L.; Yan, J.Y.; Li, Q.; Xiao, J.L.; Jin, P. Effects of weak light and waterlogging stress at jointing stage on photosynthesis, root growth and yield of spring maize. J. Jiangsu Agric. Sci. 2018, 34, 1276–1286. [Google Scholar]
- Wang, Q.; Zhao, X.Y.; Liu, D.Y.; Yan, Z.H.; Li, H.P.; Dong, P.F.; Li, C.H. Root morphological, physiological traits and yield of maize under waterlogging and low light stress. Sci. Agric. Sin. 2020, 53, 3479–3495. [Google Scholar]
- Shi, D.Y.; Li, Y.H.; Xia, D.J.; Zhang, J.W.; Liu, P.; Zhao, B.; Dong, S.T. Effects of planting density on root characteristics and nitrogen uptake in summer maize. Sci. Agric. Sin. 2017, 50, 2006–2017. [Google Scholar]
- Lambers, H.; Posthumus, F. The Effect of Light Intensity and Relative Humidity on Growth Rate and Root Respiration of Plantago lanceolata and Zea mays. J. Exp. Bot. 1980, 31, 1621–1630. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Grain yield and root characteristics of summer maize (Zea mays L.) under shade stress conditions. J. Agron. Crop Sci. 2017, 203, 562–573. [Google Scholar] [CrossRef]
- Zhang, G.; Shen, D.; Xie, R.; Ming, B.; Hou, P.; Xue, J.; Li, R.; Chen, J.; Wang, K.; Li, S. Optimizing planting density to improve nitrogen use of super high-yield maize. Agron. J. 2020, 112, 4147–4158. [Google Scholar] [CrossRef]
- Hou, P.; Chen, X.P.; Cui, Z.L.; Li, S.Q.; Wang, W.; Ye, Y.L.; Chen, Y.X.; Zhang, F.S. Potential maize yield realization and related driving factors in four typical soils. Chin. J. Eco-Agric. 2012, 20, 874–881. [Google Scholar] [CrossRef]
- Vescio, R.; Abenavoli, M.R.; Sorgonà, A. Single and Combined Abiotic Stress in Maize Root Morphology. Plants 2020, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Sattelmacher, B.; Gerendas, J.; Thoms, K.; Brück, H.; Bagdady, N. Interaction between root growth and mineral nutrition. Environ. Exp. Bot. 1993, 33, 63–73. [Google Scholar] [CrossRef]
- Jackson, R.B.; Sperry, J.S.; Dawson, T.E. Root water uptake and transport: Using physiological processes in global predictions. Trends Plant Sci. 2000, 5, 482–488. [Google Scholar] [CrossRef]
- Fan, J.; McConkey, B.; Wang, H.; Janzen, H. Root distribution by depth for temperate agricultural crops. Field Crops Res. 2016, 189, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Bañoc, D.M.; Yamauchi, A.; Kamoshita, A.; Wade, L.J.; Pardales, J.R.; Banco, D.M. Dry Matter Production and Root System Development of Rice Cultivars under Fluctuating Soil Moisture. Plant Prod. Sci. 2000, 3, 197–207. [Google Scholar] [CrossRef]
- Ogawa, A.; Kawashima, C.; Yamauchi, A. Sugar Accumulation along the Seminal Root Axis, as Affected by Osmotic Stress in Maize: A Possible Physiological Basis for Plastic Lateral Root Development. Plant Prod. Sci. 2005, 8, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, L.; Sun, X.; Wang, X.C.; Chen, Y.L.; Rengel, Z.; Liu, W.G.; Yang, W.Y. Light intensity influence maize ad-aptation to low P stress by altering root morphology. Plant Soil 2020, 447, 183–197. [Google Scholar] [CrossRef]
- Amos, B.; Walters, D.T. Maize root biomass and net rhizodeposited carbon: An analysis of the literature. Soil Sci. Soc. Am. J. 2006, 70, 1489–1503. [Google Scholar] [CrossRef]
- Hébert, Y.; Guingo, E.; Loudet, O. The Response of Root/Shoot Partitioning and Root Morphology to Light Reduction in Maize Genotypes. Crop Sci. 2001, 41, 363–371. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zhu, Y.; Li, D.; Gao, Y. Effects of stage drought and re-watering on photosynthesis, root shoot ratio and water use efficiency of summer maize (Zea mays L.). Appl. Ecol. Environ. Res. 2020, 18, 7643–7653. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, K.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Effects of integrated agronomic practices management on root growth and development of summer maize. Eur. J. Agron. 2017, 84, 140–151. [Google Scholar] [CrossRef]
- Wu, Q.; Pagès, L.; Wu, J. Relationships between root diameter, root length and root branching along lateral roots in adult, field-grown maize. Ann. Bot. 2016, 117, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Zobel, R.W.; Kinraide, T.B.; Baligar, V.C. Fine root diameters can change in response to changes in nutrient concentrations. Plant Soil 2007, 297, 243–254. [Google Scholar] [CrossRef]
- Chen, Y.L.; Wu, Q.P.; Chen, X.C.; Chen, F.J.; Zhang, Y.J.; Li, Q.; Yuan, L.X.; Mi, G.H. Root growth and its response to increasing planting density in different maize hybrids. Plant Nutr. Fertil. Sci. 2012, 18, 52–59. [Google Scholar]
- Ryser, P.; Lambers, H. Root and leaf attributes accounting for the performance of fast- and slow-growing grasses at different nutrient supply. Plant Soil 1995, 170, 251–265. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Chen, Y.; Li, Q.; Chen, F.; Yuan, L.; Mi, G. Changes in root size and distribution in relation to nitrogen accumulation during maize breeding in China. Plant Soil 2013, 374, 121–130. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Costs and benefits of constructing roots of small diameter. J. Plant Nutr. 1992, 15, 763–782. [Google Scholar] [CrossRef]
- Braconnier, S. Maize-coconut intercropping: Effects of shade and root competition on maize growth and yield. Agronomie 1998, 18, 373–382. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, X.; Lin, X.; Sassenrath, G.F.; Dai, S.; Lv, S.; Chen, X.; Chen, F.; Mi, G. Radiation Interception and Use Efficiency Contributes to Higher Yields of Newer Maize Hybrids in Northeast China. Agron. J. 2015, 107, 1473–1480. [Google Scholar] [CrossRef]
- Liu, G.-Z.; Liu, W.-M.; Hou, P.; Ming, B.; Yang, Y.-S.; Guo, X.-X.; Xie, R.-Z.; Wang, K.-R.; Li, S.-K. Reducing maize yield gap by matching plant density and solar radiation. J. Integr. Agric. 2021, 20, 363–370. [Google Scholar] [CrossRef]
- Van Ittersum, M.K.; Cassman, K.G. Yield gap analysis—Rationale, methods and applications—Introduction to the Special Issue. Field Crops Res. 2013, 143, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.S.; Xu, W.J.; Hou, P.; Liu, G.Z.; Liu, W.M.; Wang, Y.H.; Zhao, R.L.; Ming, B.; Xie, R.Z.; Wang, K.R.; et al. Improving maize grain yield by matching maize growth and solar radiation. Sci. Rep.-UK 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hammer, G.L.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paszkiewicz, S.; Cooper, M. Can Changes in Canopy and/or Root System Architecture Explain Historical Maize Yield Trends in the US Corn Belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Xia, H.-Y.; Zhao, J.; Sun, J.-H.; Bao, X.-G.; Christie, P.; Zhang, F.-S.; Li, L. Dynamics of root length and distribution and shoot biomass of maize as affected by intercropping with different companion crops and phosphorus application rates. Field Crops Res. 2013, 150, 52–62. [Google Scholar] [CrossRef]
- Li, S.-X.; Wang, Z.-H.; Malhi, S.; Li, S.-Q.; Gao, Y.-J.; Tian, X.-H. Chapter 7 Nutrient and Water Management Effects on Crop Production, and Nutrient and Water Use Efficiency in Dryland Areas of China. Adv. Agron. 2009, 102, 223–265. [Google Scholar] [CrossRef]
- Zarebanadkouki, M.; Kroener, E.; Kaestner, A.; Carminati, A.M. Visualization of root water uptake: Quantification of deuterated water transport in roots using neutron radiography and numerical modeling. Plant Physiol. 2014, 166, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application. Field Crops Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Base Fertilizers | Topdressing | |||
|---|---|---|---|---|
| N | P2O5 | K2O | N | |
| Kg ha−1 | Kg ha−1 | Kg ha−1 | Kg ha−1 | |
| Qitai | 150 | 225 | 75 | 300 |
| Yinchuan | 75 | 188 | 53 | 300 |
| Root Trait | Functional Characteristics |
|---|---|
| Total root length/surface area (RSA)/root weight ratio (RWR) | The total system size: the size of contact with soil (major determinant of water and nutrient uptake as an entire root system) |
| Root length density (RLD) | Rate of water and nutrient uptake |
| Root diameter | Potential for penetration ability, branching, hydraulic conductivity |
| Specific root length (SRL) | Degree of branching, density of root materials, porosity due to aerenchyma development |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Yang, Y.; Liu, H.; Liu, G.; Liu, W.; Wang, Y.; Zhao, R.; Ming, B.; Xie, R.; Wang, K.; et al. Effects of Solar Radiation on Dry Matter Distribution and Root Morphology of High Yielding Maize Cultivars. Agriculture 2022, 12, 299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020299
Guo X, Yang Y, Liu H, Liu G, Liu W, Wang Y, Zhao R, Ming B, Xie R, Wang K, et al. Effects of Solar Radiation on Dry Matter Distribution and Root Morphology of High Yielding Maize Cultivars. Agriculture. 2022; 12(2):299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020299
Chicago/Turabian StyleGuo, Xiaoxia, Yunshan Yang, Huifang Liu, Guangzhou Liu, Wanmao Liu, Yonghong Wang, Rulang Zhao, Bo Ming, Ruizhi Xie, Keru Wang, and et al. 2022. "Effects of Solar Radiation on Dry Matter Distribution and Root Morphology of High Yielding Maize Cultivars" Agriculture 12, no. 2: 299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020299