
Dynamic Resistant Starch Accumulation in Contrasting Wheat Genotypes Highlights the Lipid Metabolic Pathway Related to Resistant Starch Synthesis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Field Experiments
2.2. Grain TGW, TS and RS Measurement
2.3. RNA Extraction and Sequencing
2.4. Data Filtering, DEG Identification and Functional Annotation
2.5. Weighted Gene Coexpression Network Analysis
2.6. Quantitative Real-Time PCR for RNA-seq Validation
3. Results
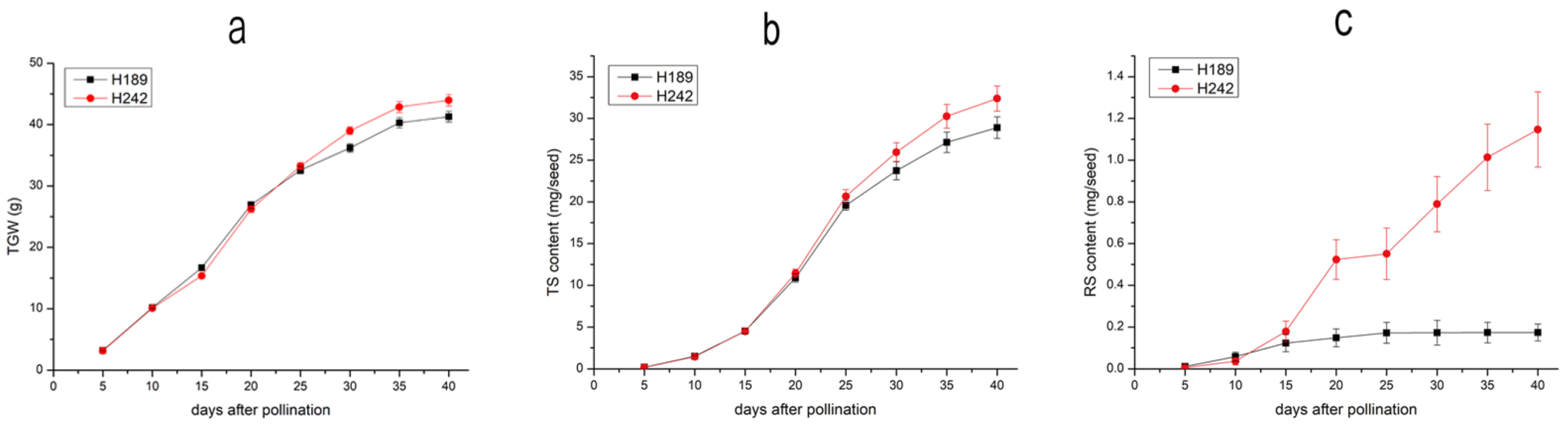
3.1. Grain TGW, TS and RS Contents and Accumulation Rates
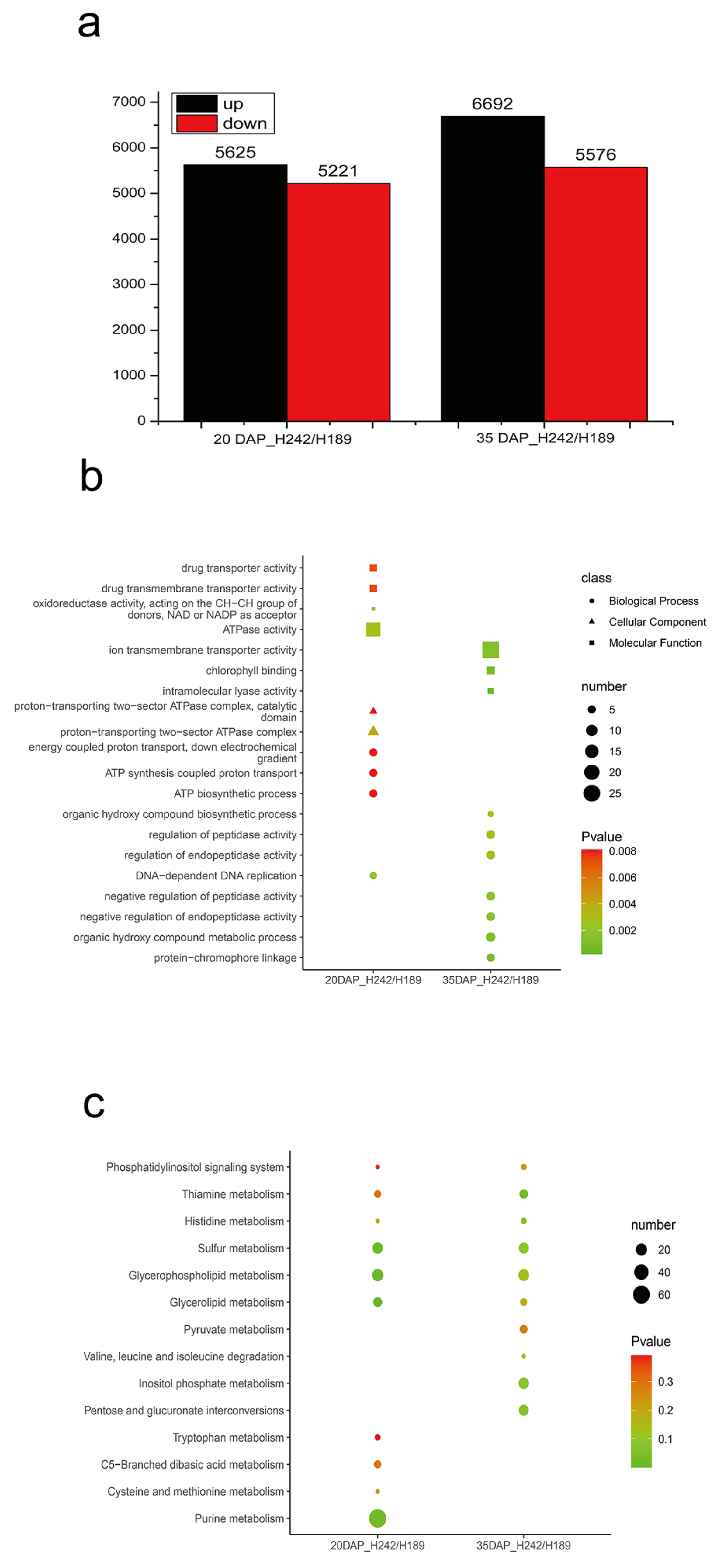
3.2. Differential Gene Expression Analysis
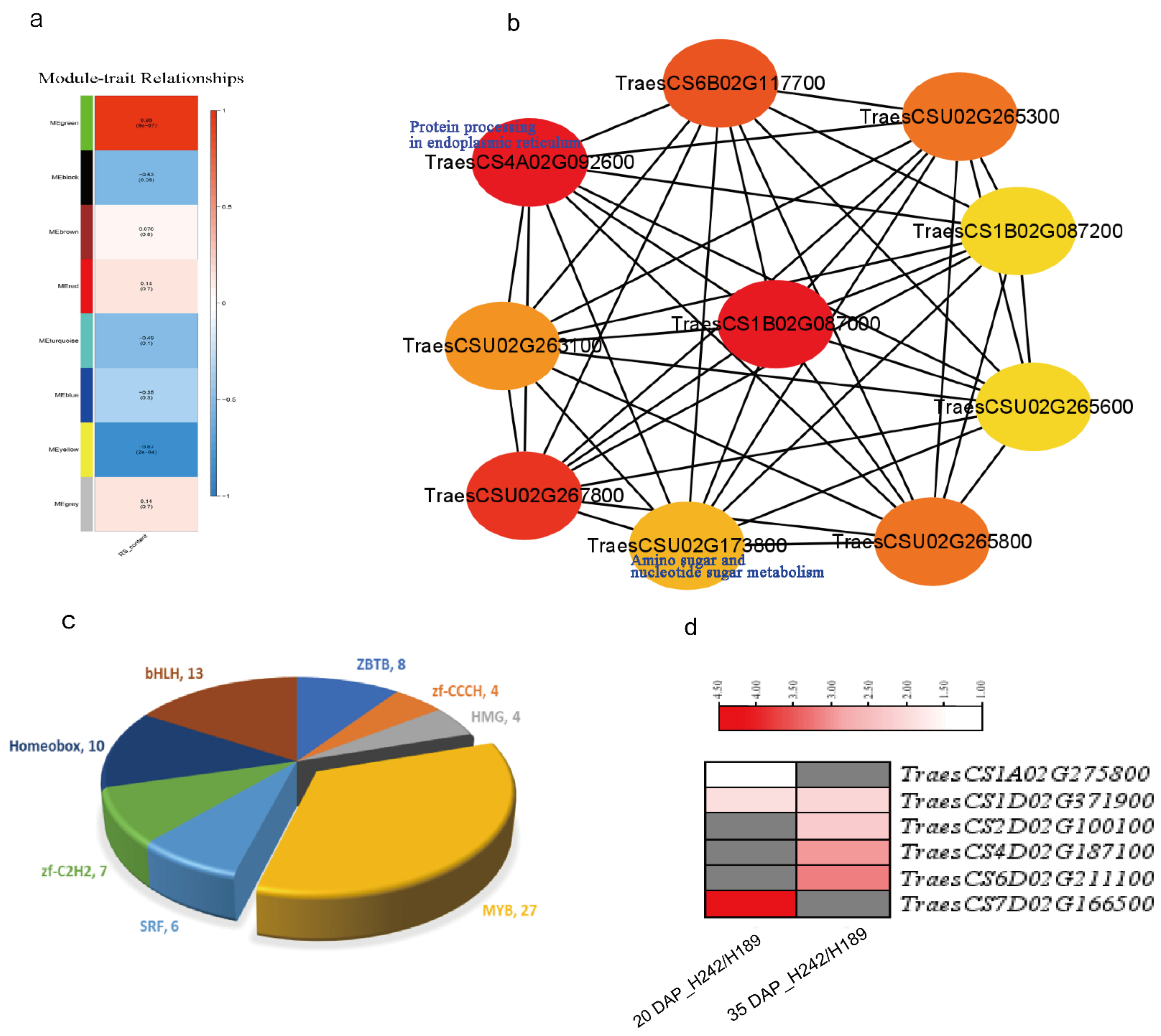
3.3. Pathways and TFs Involved in RS Formation Revealed by WGCNA
3.4. Validation of RNA-Seq Analysis by qRT-PCR
4. Discussion
4.1. Secondary Accumulation of Grain RS
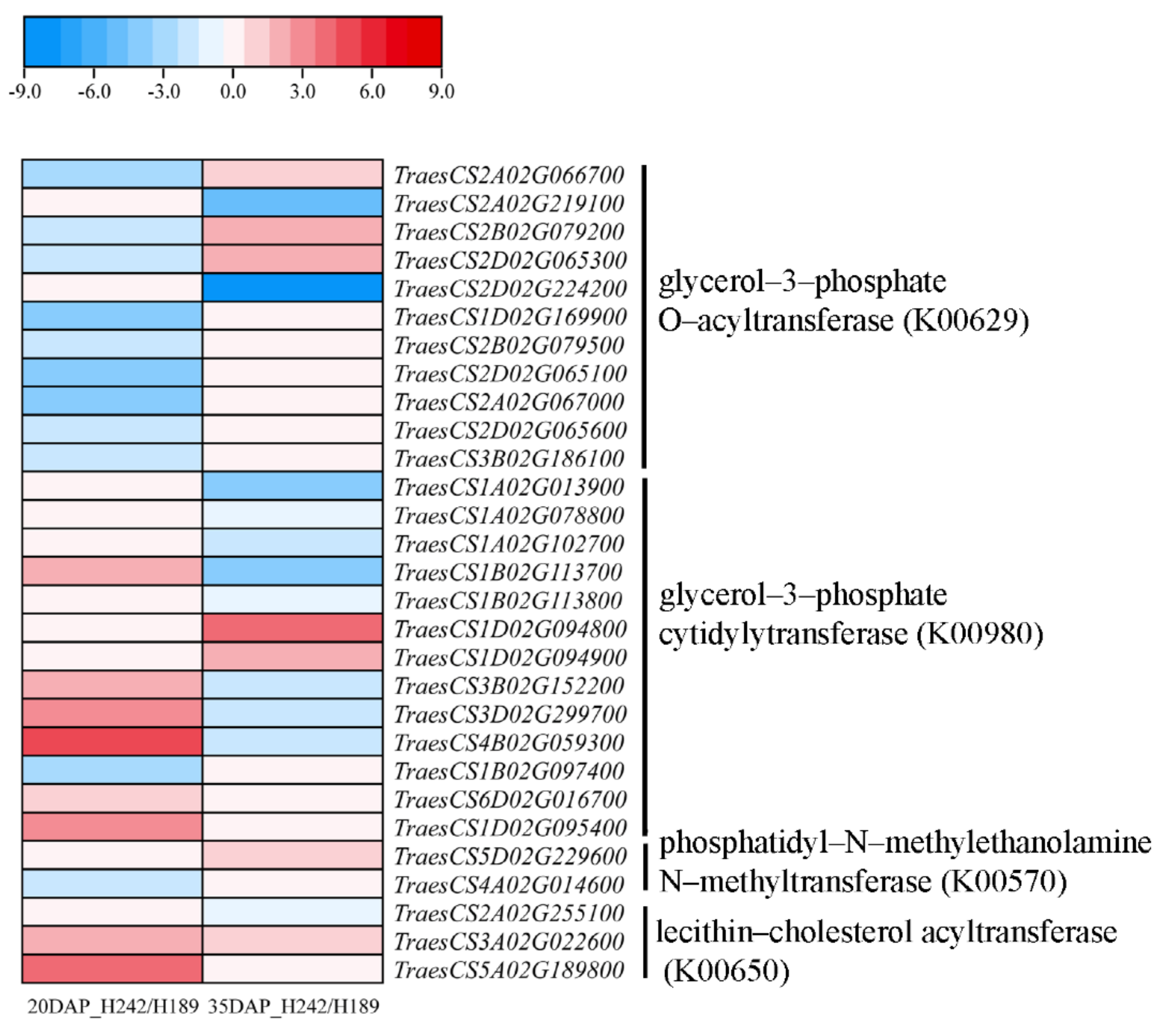
4.2. Lipid Pathways Play a Key Role in RS Formation in High-RS-Content Wheat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Englyst, K.; Goux, A.; Meynier, A.; Quigley, M.; Englyst, H.; Brack, O.; Vinoy, S. Inter-laboratory validation of the starch digestibility method for determination of rapidly digestible and slowly digestible starch. Food Chem. 2018, 245, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Meenu, M.; Xu, B. A critical review on anti-diabetic and anti-obesity effects of dietary resistant starch. Crit. Rev. Food Sci. Nutr. 2019, 59, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.Y.; Rowling, M.J. Resistant starch as a novel dietary strategy to maintain kidney health in diabetes mellitus. Nutr. Rev. 2017, 75, 350–360. [Google Scholar] [CrossRef] [PubMed]
- McRae, M.P. Dietary Fiber Is Beneficial for the Prevention of Cardiovascular Disease: An Umbrella Review of Meta-analyses J. Chiropr. Med. 2017, 16, 289–299. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, P.; Xiao, Z. Resistant starch prevents tumorigenesis of dimethylhydrazine-induced colon tumors via regulation of an ER stress-mediated mitochondrial apoptosis pathway. Int. J. Mol. Med. 2018, 41, 1887–1898. [Google Scholar] [CrossRef]
- Hansson, S.O.; Åman, P.; Becker, W.; Koning, D.-J.; Lagerkvist, C.J.; Larsson, I.; Lehrman, A.; Risérus, U.; Stymne, S. Breeding for public health: A strategy. Trends Food Sci. Technol. 2018, 80, 131–140. [Google Scholar] [CrossRef]
- Gutiérrez, T.J.; Tovar, J. Update of the concept of type 5 resistant starch (RS5): Self-assembled starch V-type complexes. Trends Food Sci. Technol. 2021, 109, 711–724. [Google Scholar] [CrossRef]
- Li, L.; Liu, Z.; Zhang, W.; Xue, B.; Luo, Z. Production and Applications of Amylose-Lipid Complexes as Resistant Starch: Recent Approaches. Starch-Stärke 2021, 73, 2000249. [Google Scholar] [CrossRef]
- Hasjim, J.; Ai, Y.; Jane, J.L. Novel applications of amylose-lipid complex as resistant starch type 5. Resist. Starch 2013, 79–94. [Google Scholar] [CrossRef]
- Okumus, B.N.; Tacer-Caba, Z.; Kahraman, K.; Nilufer-Erdil, D. Resistant starch type V formation in brown lentil (Lens culinaris Medikus) starch with different lipids/fatty acids. Food Chem. 2018, 240, 550–558. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Y.; Zhang, N.; Bao, J.; Wu, D.; Shu, X. The effects of internal endosperm lipids on starch properties: Evidence from rice mutant starches. J. Cereal Sci. 2019, 89, 102804. [Google Scholar] [CrossRef]
- Ye, J.; Hu, X.; Luo, S.; McClements, D.J.; Liang, L.; Liu, C. Effect of endogenous proteins and lipids on starch digestibility in rice flour. Food Res. Int. 2018, 106, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Li, C. Main controllers for improving the resistant starch content in cooked white rice. Food Hydrocoll. 2022, 122, 107083. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M.; et al. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef] [Green Version]
- Miura, S.; Koyama, N.; Crofts, N.; Hosaka, Y.; Abe, M.; Fujita, N. Generation and starch characterization of non-transgenic BEI and BEIIb double mutant rice (Oryza sativa) with ultra-high level of resistant starch. Rice 2021, 14, 1–16. [Google Scholar] [CrossRef]
- Carciofi, M.; Blennow, A.; Jensen, S.L.; Shaik, S.S.; Henriksen, A.; Buléon, A.; Holm, P.B.; Hebelstrup, K.M. Concerted suppression of all starch branching enzyme genes in barley produces amylose-only starch granules. BMC Plant Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hazard, B.; Zhang, X.; Colasuonno, P.; Uauy, C.; Beckles, D.M.; Dubcovsky, J. Induced mutations in the starch branching enzyme II (SBEII) genes increase amylose and resistant starch content in durum wheat. Crop Sci. 2012, 52, 1754–1766. [Google Scholar] [CrossRef]
- Tabassum, J.; Ahmad, S.; Hussain, B.; Mawia, A.M.; Zeb, A.; Ju, L. Applications and potential of genome-editing systems in rice improvement: Current and future perspectives. Agronomy 2021, 11, 1359. [Google Scholar] [CrossRef]
- Ahuja, G.; Jaiswal, S.; Hucl, P.; Chibbar, R.N. Genome-specific granule-bound starch synthase I (GBSSI) influences starch biochemical and functional characteristics in near-isogenic wheat (Triticum aestivum L.) lines. J. Agric. Food Chem. 2013, 61, 12129–12138. [Google Scholar] [CrossRef]
- Biselli, C.; Volante, A.; Desiderio, F.; Tondelli, A.; Gianinetti, A.; Finocchiaro, F.; Taddei, F.; Gazza, L.; Sgrulletta, D.; Cattivelli, L.; et al. GWAS for starch-related parameters in japonica rice (Oryza sativa L.). Plants 2019, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Sun, D.; Du, J.; Pu, X.Y.; Yang, S.M.; Yang, S.M.; Yang, T.; Yang, J.Z. Identification of QTLs for resistant starch and total alkaloid content in brown and polished rice. Genet. Mol. Res. 2016, 15, 15037268. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhou, X.; Xu, F.; He, Q.; Park, Y.J. Genome-wide association study of the resistant starch content in rice grains. Starke 2017, 69, 1600343. [Google Scholar] [CrossRef]
- Johnson, N.; Boatwright, J.L.; Bridges, W.; Thavarajah, P.; Kumar, S.; Shipe, E.; Thavarajah, D. Genome-wide association mapping of lentil (Lens culinaris Medikus) prebiotic carbohydrates toward improved human health and crop stress tolerance. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Backes, G.; Rasmussen, S.K. Genome-wide association study of resistant starch (RS) phenotypes in a barley variety collection. J. Agric. Food Chem. 2012, 60, 10302–10311. [Google Scholar] [CrossRef]
- Pang, H.; Wang, L.; Wang, H.L.; Xu, H.J.; Li, W.H. QTL mapping for kernel starch content in wheat. J. Triticeae Crops. 2014, 34, 1–7. [Google Scholar] [CrossRef]
- Mishra, A. Molecular and genetics basis of amylase resistant starch variation in bread wheat Triticum aestivum L. Doctoral Dissertation, Panjab University, Lahore, Pakistan, 2019. [Google Scholar]
- Shewry, P.R.; Mitchell, R.A.; Tosi, P.; Wan, Y.; Underwood, C.; Lovegrove, A.; Freeman, J.; Toole, G.A.; Mills, E.N.C.; Ward, J.L. An integrated study of grain development of wheat (cv. Hereward). J. Cereal. Sci. 2012, 56, 21–30. [Google Scholar] [CrossRef]
- Shu, X.; Sun, J.; Wu, D. Effects of grain development on formation of resistant starch in rice. Food Chem. 2014, 164, 89–97. [Google Scholar] [CrossRef]
- Yu, Y.; Zhu, D.; Ma, C.; Cao, C.; Wang, Y.; Xu, Y.; Zhang, W.; Yan, Y. Transcriptome analysis reveals key differentially expressed genes involved in wheat grain development. Crop J. 2016, 4, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.H. Plant Transcription Factors: Evolutionary, Structural and Functional Aspects. Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–11. [Google Scholar] [CrossRef]
- Feng, F.; Qi, W.; Lv, Y.; Yan, S.; Xu, L.; Yang, W.; Yuan, Y.; Chen, Y.; Zhao, H.; Song, R. OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism. Plant Cell 2018, 30, 375–396. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Luo, G.; Shen, L.; Yu, K.; Yang, W.; Li, X.; Sun, J.; Zhan, K.; Cui, D.; Liu, D.; et al. TubZIP28, a novel bZIP family transcription factor from Triticum urartu, and TabZIP28, its homologue from Triticum aestivum, enhance starch synthesis in wheat. New Phytol. 2020, 226, 1384–1398. [Google Scholar] [CrossRef]
- Kooiker, M.; Drenth, J.; Glassop, D.; McIntyre, C.L.; Xue, G.P. TaMYB13-1, a R2R3 MYB transcription factor; regulates the fructan synthetic pathway and contributes to enhanced fructan accumulation in bread wheat. J. Exp. Bot. 2013, 64, 3681–3696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Q.; Wang, Y.; Du, J.; Li, H.; Wei, B.; Wang, Y.; Li, Y.; Li, Y.; Yu, G.; Liu, H.; et al. ZmMYB 14 is an important transcription factor involved in the regulation of the activity of the ZmBT1 promoter in starch biosynthesis in maize. FEBS J. 2017, 284, 3079–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parween, S.; Anonuevo, J.J.; Butardo, V.M., Jr.; Misra, G.; Anacleto, R.; Llorente, C.; Kosik, O.; Romero, M.V.; Bandonill, E.H.; Mendioro, M.S.; et al. Balancing the double-edged sword effect of increased resistant starch content and its impact on rice texture: Its genetics and molecular physiological mechanisms. Plant Biotechnol. J. 2020, 18, 1763–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, B.; Akpınar, B.; Alaux, M.; Algharib, A.M.; Sehgal, D.; Ali, Z.; Appels, R.; Aradottir, G.I.; Batley, J.; Bellec, A.; et al. Wheat Genomics and Breeding: Bridging the Gap. AgriRxiv 2021. [Google Scholar] [CrossRef]
- Lal, M.K.; Kumar, A.; Raigond, P.; Dutt, S.; Changan, S.S.; Chourasia, K.N.; Tiwari, R.K.; Kumar, D.; Sharma, S.; Chakrabarti, S.K.; et al. Impact of Starch Storage Condition on Glycemic Index and Resistant Starch of Cooked Potato (Solanum tuberosum) Tubers. Starke 2021, 73, 1900281. [Google Scholar] [CrossRef]
- Xia, J.; Zhu, D.; Wang, R.; Cui, Y.; Yan, Y. Crop resistant starch and genetic improvement: A review of recent advances. Theor. Appl. Genet. 2018, 131, 2495–2511. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Saito, R.; Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Lotia, M.; Pico, A.R.; Bader, G.D.; Ideker, T. A travel guide to Cytoscape plugins. Nat. Methods 2012, 9, 1069–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, W.; Qi, J.; Shi, P.; Yin, Y. Starch accumulation; activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents. J. Food Sci. Technol. 2014, 51, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, X.; Jia, L.; Ye, H.; Li, C.; Wu, D. Slow digestion properties of rice different in resistant starch. J. Agric. Food Chem. 2009, 57, 7552–7559. [Google Scholar] [CrossRef]
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification; structures and tools. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Chao, C.; Cai, J.; Niu, B.; Copeland, L.; Wang, S. Starch–lipid and starch–lipid–protein complexes: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1056–1079. [Google Scholar] [CrossRef] [Green Version]
- Putseys, J.; Lamberts, L.; Delcour, J. Amylose-inclusion complexes: Formation; identity and physico-chemical properties. J. Cereal. Sci. 2010, 51, 238–247. [Google Scholar] [CrossRef]
- Wang, S.; Copeland, L. Molecular disassembly of starch granules during gelatinization and its effect on starch digestibility: A review. Food Funct. 2013, 4, 1564–1580. [Google Scholar] [CrossRef]
- Cai, C.; Tian, Y.; Sun, C.; Jin, Z. Resistant structure of extruded starch: Effects of fatty acids with different chain lengths and degree of unsaturation. Food Chem. 2022, 374, 131510. [Google Scholar] [CrossRef]
- Wang, J.; Singh, S.K.; Geng, S.; Zhang, S.; Yuan, L. Genome-wide analysis of glycerol-3-phosphate O-acyltransferase gene family and functional characterization of two cutin group GPATs in Brassica napus. Planta 2020, 251, 1–16. [Google Scholar] [CrossRef]
- Waschburger, E.; Kulcheski, F.R.; Veto, N.M.; Margis, R.; Margis-Pinheiro, M.; Turchetto-Zolet, A.C. Genome-wide analysis of the Glycerol-3-Phosphate Acyltransferase (GPAT) gene family reveals the evolution and diversification of plant GPATs. Genet. Mol. Biol. 2018, 41, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Ouyang, L.L.; Bao, H.; Liu, J.-G.; Sun, Z.; Zhou, Z.-G. Comparison between two isoforms of glycerol-3-phosphate acyltransferase in microalga Myrmecia incisa: Subcellular localization and role in triacylglycerol synthesis. Algal Res. 2021, 54, 102172. [Google Scholar] [CrossRef]
- Chandrasekaran, U.; Xu, W.; Liu, A. Transcriptome profiling identifies ABA mediated regulatory changes towards storage filling in developing seeds of castor bean (Ricinus communis L.). Cell Biosci. 2014, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- Schrick, K.; Nguyen, D.; Karlowski, W.M.; Mayer, K.F. START lipid/sterol-binding domains are amplified in plants and are predominantly associated with homeodomain transcription factors. Genome Biol. 2004, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.U. Lipid metabolism in plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after Pollination | TGW Accumulation Rate | TS Accumulation Rate | RS Accumulation Rate | |||
|---|---|---|---|---|---|---|
| (g day−1) | (mg seed−1 day−1) | (μg seed−1 day−1) | ||||
| H189 | H242 | H189 | H242 | H189 | H242 | |
| 0–5 | 0.642 a ± 0.005 | 0.631 a ± 0.007 | 0.039 a ± 0.002 | 0.036 a ± 0.002 | 2.3 a ± 0.3 | 2.2 a ± 0.3 |
| 5–10 | 1.390 a ± 0.009 | 1.385 a ± 0.021 | 0.263 a ± 0.010 | 0.256 a ± 0.008 | 9.3 a ± 0.4 | 6.0 b ± 0.3 |
| 10–15 | 1.302 a ± 0.022 | 1.057 b ± 0.009 | 0.598 a ± 0.021 | 0.606 a ± 0.009 | 13.0 b ± 0.4 | 28.4 a ± 3.8 |
| 15–20 | 2.047 b ± 0.027 | 2.171 a ± 0.023 | 1.277 a ± 0.025 | 1.390 a ± 0.040 | 5.0 b ± 0.2 | 69.0 a ± 5.3 |
| 20–25 | 1.132 b ± 0.008 | 1.405 a ± 0.003 | 1.740 a ± 0.056 | 1.844 a ± 0.059 | 4.7 a ± 0.4 | 5.5 a ± 0.4 |
| 25–30 | 0.728 b ± 0.011 | 1.150 a ± 0.007 | 0.830 b ± 0.013 | 1.057 a ± 0.014 | 0.1 b ± 0.0 | 47.7 a ± 1.1 |
| 30–35 | 0.819 a ± 0.016 | 0.772 a ± 0.033 | 0.680 b ± 0.015 | 0.862 a ± 0.034 | 0.1 b ± 0.0 | 44.9 a ± 3.1 |
| 35–40 | 0.199 a ± 0.007 | 0.216 a ± 0.003 | 0.354 b ± 0.009 | 0.425 a ± 0.009 | 0.1 b ± 0.0 | 26.7 a ± 2.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-P.; Cai, H.-Y.; Zhu, J.-H.; Wei, X.; Zhang, S.; Liu, G.; He, Y.-G.; Li, B.; Xu, L.; Jiao, C.-H.; et al. Dynamic Resistant Starch Accumulation in Contrasting Wheat Genotypes Highlights the Lipid Metabolic Pathway Related to Resistant Starch Synthesis. Agriculture 2022, 12, 308. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020308
Wang H-P, Cai H-Y, Zhu J-H, Wei X, Zhang S, Liu G, He Y-G, Li B, Xu L, Jiao C-H, et al. Dynamic Resistant Starch Accumulation in Contrasting Wheat Genotypes Highlights the Lipid Metabolic Pathway Related to Resistant Starch Synthesis. Agriculture. 2022; 12(2):308. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020308
Chicago/Turabian StyleWang, Hong-Pan, Hai-Ya Cai, Jing-Huan Zhu, Xia Wei, Shuo Zhang, Gang Liu, Yong-Gang He, Bo Li, Le Xu, Chun-Hai Jiao, and et al. 2022. "Dynamic Resistant Starch Accumulation in Contrasting Wheat Genotypes Highlights the Lipid Metabolic Pathway Related to Resistant Starch Synthesis" Agriculture 12, no. 2: 308. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020308