
Trace Mineral Solubility and Digestibility in the Small Intestine of Piglets Are Affected by Zinc and Fibre Sources

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Experiment In Vivo
Animals and Diets
Animal Performance
Apparent Digestibility
2.1.2. Experiment In Situ
2.1.3. Experiment In Vitro
In Vitro Solubility of Zn Sources
In Vitro Simulated Solubility of TEs
In Vitro Dry Matter Digestibility and pH
2.2. Chemical Analysis
2.3. Statistical Analysis
3. Results
3.1. Experimetn In Vivo
3.1.1. Animal performance
3.1.2. Apparent Digestibility
3.2. Experiment in situ
In Situ Soluble TEs
3.3. Experiment In Vitro
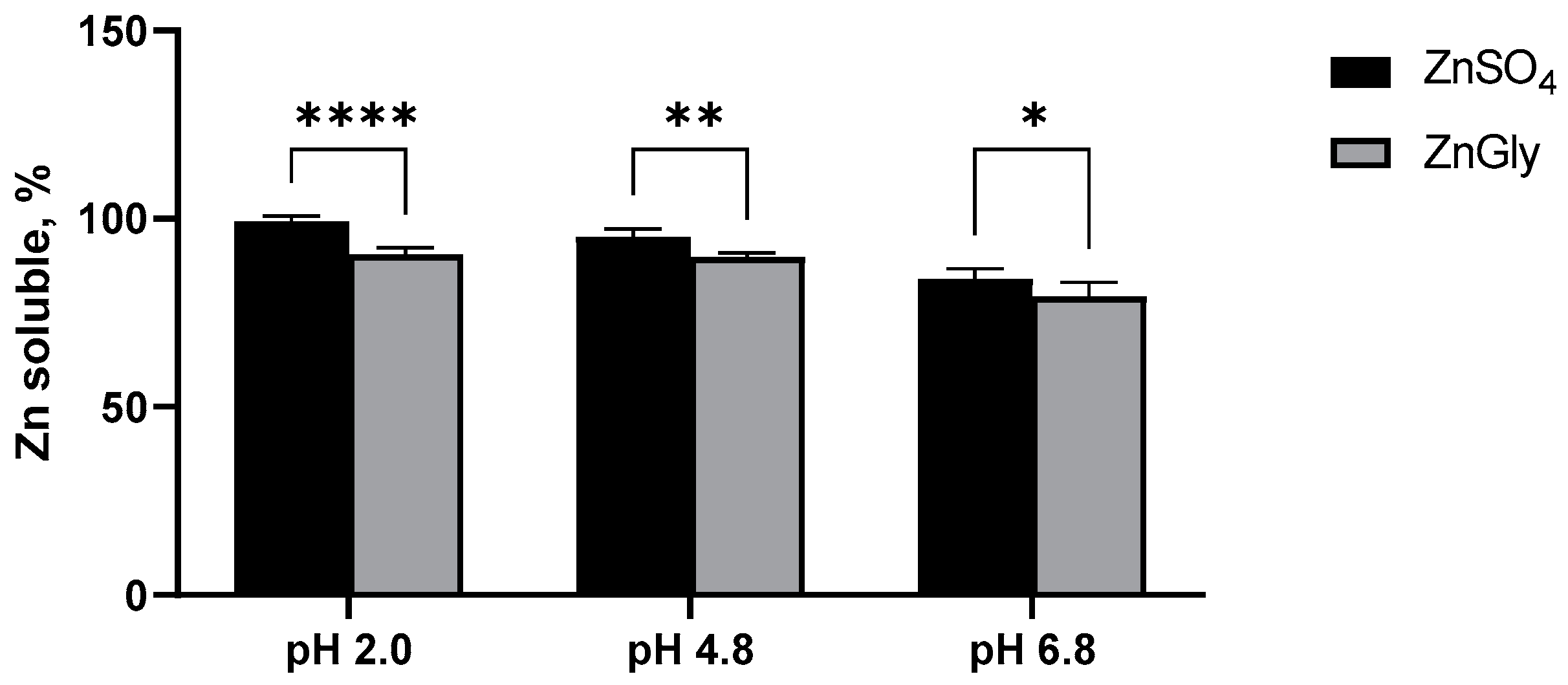
3.3.1. In Vitro Solubility of Zn Sources
3.3.2. In Vitro Simulated Solubility of TEs
3.3.3. In Vitro Dry Matter Digestibility and pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdelrahman, M.M.; Kincaid, R.L.; Elzubeir, E.A. Mineral Nutrition of Livestock, 4th ed.; CABI: Wallingford, UK, 2010; ISBN 978-1845934729. [Google Scholar]
- Fairweather-Tait, S.J. Bioavailability of trace elements. Food Chem. 1992, 43, 213–217. [Google Scholar] [CrossRef]
- Etcheverry, P.; Grusak, M.A.; Fleige, L.E. Application of in vitro bioaccessibility and bioavailability methods for calcium, carotenoids, folate, iron, magnesium, polyphenols, zinc, and vitamins B6, B12, D, and E. Front Physiol. 2012, 3, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandberg, A.-S. Bioavailability of minerals in legumes. Br. J. Nutr. 2002, 88, 281–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef]
- Salgueiro, M.J.; Zubillaga, M.; Lysionek, A.; Sarabia, M.I.; Caro, R.; De Paoli, T.; Hager, A.; Weill, R.; Boccio, J. Zinc as an essential micronutrient: A review. Nutr. Res. 2000, 20, 737–755. [Google Scholar] [CrossRef]
- Walk, C.L.; Wilcock, P.; Magowan, E. Evaluation of the effects of pharmacological zinc oxide and phosphorus source on weaned piglet growth performance, plasma minerals and mineral digestibility. Animal 2015, 9, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Song, J.; You, Z.; Luan, Z.; Li, W. Zinc oxide-montmorillonite hybrid influences diarrhea, intestinal mucosal integrity, and digestive enzyme activity in weaned pigs. Biol. Trace Elem. Res. 2012, 149, 190–196. [Google Scholar] [CrossRef]
- EC. Commission regulation (EC) No 1334/2003 of 25 July 2003 amending the conditions for authorisation of a number of additives in feedingstuffs belonging to the group of trace elements. Off. J. Eur. Union 2003, 1334, L187/11–L187/15. [Google Scholar]
- EC. Commission implementing regulation (EC), No 2016/1095 of 6 July 2016. Off. J. Eur. Union 2016, 10, 182/7–182/27. [Google Scholar]
- Scientific Committee for Animal Nutrition of the European Commission. Opinion of the Scientific Committee for Animal Nutrition on the Use of Zinc in Feedingstuffs; European Commission—Health & Consumer Protection Directorate General: Brussels, Belgium, 2003; Available online: https://ec.europa.eu/food/system/files/2020-12/sci-com_scan-old_report_out120.pdf (accessed on 16 December 2021).
- Wedekind, K.J.; Hortin, A.E.; Baker, D.H. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J. Anim. Sci. 1992, 70, 178–187. [Google Scholar] [CrossRef]
- Cao, J.; Henry, P.R.; Guo, R.; Holwerda, R.A.; Toth, J.P.; Littell, R.C.; Miles, R.D.; Ammerman, C.B. Chemical characteristics and relative bioavailability of supplemental organic zinc sources for poultry and ruminants 1,2. J. Anim. Sci 2000, 78, 2039–2054. [Google Scholar] [CrossRef] [Green Version]
- Lönnerdal, B. Phytic acid-trace element (Zn, Cu, Mn) interactions. Int. J. Food Sci. Technol. 2002, 37, 749–758. [Google Scholar] [CrossRef]
- Windisch, W. Interaction of chemical species with biological regulation of the metabolism of essential trace elements. Fresenius. J. Anal. Chem. 2002, 372, 421–425. [Google Scholar] [CrossRef]
- Lönnerdal, B. Dietary Factors Influencing Zinc Absorption. J. Nutr. 2000, 130, 1378S–1383S. [Google Scholar] [CrossRef]
- Yu, Y.; Lu, L.; Li, S.F.; Zhang, L.Y.; Luo, X.G. Organic zinc absorption by the intestine of broilers in vivo. Br. J. Nutr. 2017, 117, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Yue, M.; Fang, S.L.; Zhuo, Z.; Li, D.D.; Feng, J. Zinc glycine chelate absorption characteristics in sprague dawley rat. J. Anim. Physiol. Anim. Nutr. 2015, 99, 457–464. [Google Scholar] [CrossRef]
- Affonfere, M.; Chadare, F.J.; Fassinou, F.T.K.; Linnemann, A.R.; Duodu, K.G. In-vitro Digestibility Methods and Factors Affecting Minerals Bioavailability: A Review. Food Rev. Int. 2021, 1–29. [Google Scholar] [CrossRef]
- Greger, J.L. Nondigestible carbohydrates and mineral bioavailability. J. Nutr. 1999, 129, 1434S–1435S. [Google Scholar] [CrossRef] [Green Version]
- Solomons, N.W. Dietary sources of zinc and factors affecting its bioavailability. Food Nutr. Bull. 2001, 22, 138–154. [Google Scholar] [CrossRef]
- Knudsen, E.; Sandstrom, B.; Solgaard, P. Zinc, copper and magnesium absorption from a fibre-rich diet. J. Trace Elem. Med. Biol. 1996, 10, 68–76. [Google Scholar] [CrossRef]
- Barszcz, M.; Taciak, M.; Tuśnio, A.; Čobanová, K.; Grešáková, L. The effect of organic and inorganic zinc source, used in combination with potato fiber, on growth, nutrient digestibility and biochemical blood profile in growing pigs. Livest. Sci. 2019, 227, 37–43. [Google Scholar] [CrossRef]
- Kleinman, P.; Sullivan, D.; Wolf, A.; Brandt, R.; Dou, Z.; Elliott, H.; Kovar, J.; Leytem, A.; Maguire, R.; Moore, P.; et al. Selection of a Water-Extractable Phosphorus Test for Manures and Biosolids as an Indicator of Runoff Loss Potential. J. Environ. Qual. 2007, 36, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.F.; Zeringue, L.K. Laboratory Evaluations of Solubility and Structural Integrity of Complexed and Chelated Trace Mineral Supplements. J. Dairy Sci. 1994, 77, 181–189. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J.A. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; revision 2; AOAC: Gaithersburg, MD, USA, 2007; Chapter 4; pp. 1–2. [Google Scholar]
- Gresakova, L.; Venglovska, K.; Cobanova, K. Dietary manganese source does not affect Mn, Zn and Cu tissue deposition and the activity of manganese-containing enzymes in lambs. J. Trace Elem. Med. Biol. 2016, 38, 138–143. [Google Scholar] [CrossRef]
- Kimura, F.T.; Miller, V.L. Chromic Oxide Measurement, Improved Determination of Chromic Oxide in Cow Feed and Feces. J. Agric. Food Chem. 2002, 5, 216. [Google Scholar] [CrossRef]
- Brugger, D.; Windisch, W.M. Environmental responsibilities of livestock feeding using trace mineral supplements. Anim. Nutr. 2015, 1, 113–118. [Google Scholar] [CrossRef]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Windisch, W.; Vikari, A.; Hilz, C. Homeostatic response of Zn metabolism to dietary Zn supplements from sulfate, gluconate, orotate, aspartate or histidine in 65Zn labeled non-growing rats as a model to adult individuals. Trace Elem. Electrolytes 2003, 20, 125–133. [Google Scholar] [CrossRef]
- Schlegel, P.; Windisch, W. Bioavailability of zinc glycinate in comparison with zinc sulphate in the presence of dietary phytate in an animal model with 65Zn labelled rats. J. Anim. Physiol. Anim. Nutr. 2006, 90, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Spears, J.W.; Schlegel, P.; Seal, M.C.; Lloyd, K.E. Bioavailability of zinc from zinc sulfate and different organic zinc sources and their effects on ruminal volatile fatty acid proportions. Livest. Prod. Sci. 2004, 90, 211–217. [Google Scholar] [CrossRef]
- Hambidge, K.M.; Miller, L.V.; Westcott, J.E.; Sheng, X.; Krebs, N.F. Zinc bioavailability and homeostasis. Am. J. Clin. Nutr. 2010, 91, 1478S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, R.J. Gastrointestinal factors influencing zinc absorption and homeostasis. Int. J. Vitam. Nutr. Res. 2010, 80, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.R.; Beiseigel, J.M.; Johnson, L.A.K. Adaptation in human zinc absorption as influenced by dietary zinc and bioavailability. Am. J. Clin. Nutr. 2008, 87, 1336–1345. [Google Scholar] [CrossRef] [Green Version]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [Green Version]
- Nitrayova, S.; Windisch, W.; von Heimendahl, E.; Müller, A.; Bartelt, J. Bioavailability of zinc from different sources in pigs. J. Anim. Sci. 2012, 90 (Suppl. S4), 185–187. [Google Scholar] [CrossRef]
- Kelsay, J.L. Effects of Fiber on Mineral and Vitamin Bioavailability. In Dietary Fiber in Health and Disease; Springer: Boston, MA, USA, 1982; pp. 91–103. [Google Scholar] [CrossRef]
- Torre, M.; Rodriguez, A.R.; Saura-Calixto, F. Interactions of Fe(II), Ca(II) and Fe(III) with high dietary fibre materials: A physicochemical approach. Food Chem. 1995, 54, 23–31. [Google Scholar] [CrossRef]
- Persson, H.; Nyman, M.; Liljeberg, H.; Önning, G.; Frølich, W. Binding of mineral elements by dietary fibre components in cereals—In vitro (III). Food Chem. 1991, 40, 169–183. [Google Scholar] [CrossRef]
- Ward, A.T.; Reichert, R.D. Comparison of the effect of cell wall and hull fiber from canola and soybean on the bioavailability for rats of minerals, protein and lipid. J. Nutr. 1986, 116, 233–241. [Google Scholar] [CrossRef]
- Barczynska, R.; Slizewska, K.; Litwin, M.; Szalecki, M.; Zarski, A.; Kapusniak, J. The effect of dietary fibre preparations from potato starch on the growth and activity of bacterial strains belonging to the phyla Firmicutes, Bacteroidetes, and Actinobacteria. J. Funct. Foods 2015, 19, 661–668. [Google Scholar] [CrossRef]
- Dobbing, J. Dietary Starches and Sugars in Man: A Comparison; Springer Science&Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Morais, M.B.; Feste, A.; Miller, R.G.; Lifschitz, C.H. Effect of Resistant and Digestible Starch on Intestinal Absorption of Calcium, Iron, and Zinc in Infant Pigs. Pediatr. Res. 1996, 39, 872–876. [Google Scholar] [CrossRef] [Green Version]
- Gillooly, M.; Bothwell, T.H.; Torrance, J.D.; MacPhail, A.P.; Derman, D.P.; Bezwoda, W.R.; Mills, W.; Charlton, R.W.; Mayet, F. The effects of organic acids, phytates and polyphenols on the absorption of iron from vegetables. Br. J. Nutr. 1983, 49, 331–342. [Google Scholar] [CrossRef]
- Van Den Berghe, P.V.E.; Klomp, L.W.J. New developments in the regulation of intestinal copper absorption. Nutr. Rev. 2009, 67, 658–672. [Google Scholar] [CrossRef]
- Pang, Y.; Applegate, T.J. Effects of dietary copper supplementation and copper source on digesta pH, calcium, zinc, and copper complex size in the gastrointestinal tract of the broiler chicken. Poult. Sci. 2007, 86, 531–537. [Google Scholar] [CrossRef]
- Drago, S.R. Minerals. In Nutraceutical and Functional Food Components: Effects of Innovative Processing Techniques; Academic Press: Cambridge, MA, USA, 2017; pp. 129–157. [Google Scholar] [CrossRef]
- Boisen, A.N.S.; Eggum, D.B. Critical Evaluation of in Vitro Methods for Estimating Digestibility in Simple-Stomach Animals. Nutr. Res. Rev. 1991, 4, 141–162. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Cai, X.; Chen, X.; Du, H.; Xu, J.; Wang, L.; Sun, G.; Cui, Y. Investigation of bioaccessibility of Cu, Fe, Mn, and Zn in market vegetables in the colon using PBET combined with SHIME. Sci. Rep. 2017, 7, 17578. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.S.; Bailey, K.B.; Gibbs, M.; Ferguson, E.L. A review of phytate, iron, zinc, and calcium concentrations in plant-based complementary foods used in low-income countries and implications for bioavailability. Food Nutr. Bull. 2010, 31, 134–146. [Google Scholar] [CrossRef]
- Lyon, D.B. Studies on the solubility of Ca, Mg, Zn, and Cu in cereal products. Am. J. Clin. Nutr. 1984, 39, 190–195. [Google Scholar] [CrossRef]
- Lönnerdal, B.; Jayawickrama, L.; Lien, E.L. Effect of reducing the phytate content and of partially hydrolyzing the protein in soy formula on zinc and copper absorption and status in infant rhesus monkeys and rat pups. Am. J. Clin. Nutr. 1999, 69, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Egli, I.; Davidsson, L.; Zeder, C.; Walczyk, T.; Hurrell, R. Dephytinization of a Complementary Food Based on Wheat and Soy Increases Zinc, but Not Copper, Apparent Absorption in Adults. J. Nutr. 2004, 134, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Khouzam, R.B.; Pohl, P.; Lobinski, R. Bioaccessibility of essential elements from white cheese, bread, fruit and vegetables. Talanta 2011, 86, 425–428. [Google Scholar] [CrossRef]
- Schümann, K.; Elsenhans, B. The impact of food contaminants on the bioavailability of trace metals. J. Trace Elem. Med. Biol. 2002, 16, 139–144. [Google Scholar] [CrossRef]
- Greiner, R.; Konietzny, U. Phytases: Biochemistry, enzymology and characteristics relevant to animal feed use. Enzym. Farm Anim. Nutr. 2010, 2, 96–128. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Phytate-degrading enzymes in pig nutrition. Livest. Sci. 2008, 113, 99–122. [Google Scholar] [CrossRef]
- Kemme, P.A.; Schlemmer, U.; Mroz, Z.; Jongbloed, A.W. Monitoring the stepwise phytate degradation in the upper gastrointestinal tract of pigs. J. Sci. Food Agric. 2006, 86, 612–622. [Google Scholar] [CrossRef]
- Schlemmer, U.; Jany, K.-D.; Berk, A.; Schulz, E.; Rechkemmer, G. Degradation of phytate in the gut of pigs—Pathway of gastro-intestinal inositol phosphate hydrolysis and enzymes involved. Arch. Tierernahr. 2001, 55, 255–280. [Google Scholar] [CrossRef]
- Leach, G.; Patton, R.S. Analysis techniques for chelated minerals evaluated. Feedstuffs 1997, 69, 13–15. [Google Scholar]
- Donkoh, A.; Moughan, P.J.; Smith, W.C. Comparison of the slaughter method and simple T-piece cannulation of the terminal ileum for determining ileal amino acid digestibility in meat and bone meal for the growing pig. Anim. Feed Sci. Technol. 1994, 49, 43–56. [Google Scholar] [CrossRef]
- Guo, R.; Henry, P.R.; Holwerda, R.A.; Cao, J.; Littell, R.C.; Miles, R.D.; Ammerman, C.B. Chemical characteristics and relative bioavailability of supplemental organic copper sources for poultry. J. Anim. Sci. 2001, 79, 1132–1141. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Ingredients (g/kg) | Diets 1 | Analysed Composition (g/kg) | Diets 1 | ||
|---|---|---|---|---|---|
| C | PF | C | PF | ||
| Barley | 200 | 200 | Dry matter | 889 | 890 |
| Wheat | 450 | 450 | Crude ash | 47 | 49 |
| Soybean meal | 170 | 170 | Crude protein | 182 | 183 |
| Yellow lupin | 60 | 60 | Crude fat | 42 | 41 |
| Rapeseed oil | 25.5 | 25.5 | Crude fibre | 41 | 40 |
| Corn starch | 33 | - | Sugars | 41 | 39 |
| Cellulose | 17 | - | Starch | 378 | 379 |
| Potato fibre | - | 50 | Total phosphorus | 5.1 | 5.08 |
| Mineral-vitamin mix 2 | 4 | 4 | Phytic phosphorus | 1.90 | 1.68 |
| Monocalcium phosphate | 7 | 7 | aNDFom | 108 | 112 |
| Calcium carbonate | 19 | 19 | ADFom | 55 | 56 |
| Sodium chloride | 3 | 3 | Lignin (sa) | 11 | 10 |
| L-Lysine | 6 | 6 | Cellulose | 43 | 47 |
| DL-Methionine | 2 | 2 | Hemicellulose | 53 | 55 |
| L-Threonine | 3 | 3 | Zn (mg/kg) | 142 | 141 |
| L-Tryptophan | 0.5 | 0.5 | Mn (mg/kg) | 99 | 94 |
| Cu (mg/kg) | 125 | 132 | |||
| Fe (mg/kg) | 315 | 300 | |||
| Gross energy (MJ/kg) | 16.8 | 16.6 | |||
| Apparent Digestibility, % | Dietary Treatment 1 | SE | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | ZnGly | PF | PF + ZnGly | Zn | Fibre | Zn × Fibre | ||
| Zinc | Zn digestibility, % | |||||||
| Total tract | 29.08 b | 21.81 a | 30.53 b | 28.56 b | 1.363 | 0.012 | 0.002 | 0.187 |
| Duodenum | 68.47 | 66.03 | 64.92 | 77.06 | 4.758 | 0.351 | 0.441 | 0.142 |
| Jejunum | 60.61 | 54.36 | 58.00 | 57.90 | 4.597 | 0.499 | 0.920 | 0.512 |
| Ileum | 57.31 | 50.23 | 55.73 | 53.95 | 2.991 | 0.156 | 0.724 | 0.387 |
| Copper | Cu digestibility, % | |||||||
| Total tract | 20.05 ab | 12.94 a | 23.77 ab | 26.51 b | 2.965 | 0.470 | 0.009 | 0.114 |
| Duodenum | 61.37 AB | 55.27 AB | 52.04 A | 67.50 B | 4.982 | 0.378 | 0.774 | 0.043 |
| Jejunum | 27.44 | 21.58 | 30.07 | 38.83 | 5.086 | 0.806 | 0.105 | 0.225 |
| Ileum | 31.77 | 24.75 | 33.92 | 35.81 | 7.37 | 0.732 | 0.382 | 0.553 |
| Iron | Fe digestibility, % | |||||||
| Total tract | 14.42 b | 16.54 b | 4.255 a | 6.064 a | 2.226 | 0.391 | 0.0003 | 0.946 |
| Duodenum | 42.97 | 37.35 | 32.37 | 43.26 | 8.453 | 0.426 | 0.759 | 0.342 |
| Jejunum | 29.79 | 28.80 | 23.39 | 21.76 | 6.618 | 0.846 | 0.330 | 0.963 |
| Ileum | 23.40 | 30.70 | 10.86 | 11.19 | 5.278 | 0.480 | 0.008 | 0.519 |
| Manganese | Mn digestibility, % | |||||||
| Total tract | −21.93 B | −29.27 B | −25.59 B | −52.06 A | 4.051 | 0.0001 | 0.0001 | 0.0001 |
| Duodenum | 28.73 | 43.09 | 32.61 | 57.06 | 9.292 | 0.0498 | 0.349 | 0.5934 |
| Jejunum | 16.28 | 10.73 | 10.83 | 20.62 | 10.03 | 0.835 | 0.828 | 0.456 |
| Ileum | 12.85 | 14.46 | 14.22 | 12.55 | 5.210 | 0.995 | 0.959 | 0.757 |
| In Situ Solubility, % | Dietary Treatment 1 | SE | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | ZnGly | PF | PF + ZnGly | Zn | Fibre | Zn × Fibre | ||
| Zinc | Soluble Zn, % | |||||||
| Duodenum | 40.11 AB | 52.42 B | 57.02 B | 19.30 A | 9.349 | 0.1847 | 0.3929 | 0.0121 |
| Jejunum | 31.83 BC | 25.52 AB | 22.18 A | 34.40 C | 2.586 | 0.2605 | 0.8823 | 0.0009 |
| Ileum | 19.30 A | 16.50 A | 31.92 B | 19.20 A | 2.129 | 0.0007 | 0.0008 | 0.0243 |
| Copper | Soluble Cu, % | |||||||
| Duodenum | 50.51 | 54.27 | 49.86 | 43.47 | 3.647 | 0.7216 | 0.1267 | 0.1739 |
| Jejunum | 102.8 | 99.88 | 75.03 | 85.33 | 8.250 | 0.6088 | 0.0052 | 0.3608 |
| Ileum | 82.55 | 83.64 | 78.99 | 75.51 | 3.286 | 0.7186 | 0.0832 | 0.4910 |
| Iron | Soluble Fe, % | |||||||
| Duodenum | 15.35 | 16.25 | 20.39 | 14.28 | 1.955 | 0.1924 | 0.4383 | 0.0827 |
| Jejunum | 48.62 b | 30.48 a | 45.21 ab | 44.25 ab | 4.483 | 0.0389 | 0.2544 | 0.0621 |
| Ileum | 44.94 b | 29.51 a | 63.42 c | 46.25 b | 3.450 | <0.0001 | <0.0001 | 0.8022 |
| Manganese | Soluble Mn, % | |||||||
| Duodenum | 89.88 b | 77.40 ab | 79.00 ab | 44.05 a | 10.80 | 0.0366 | 0.0501 | 0.3072 |
| Jejunum | 39.67 b | 44.77 b | 27.73 a | 39.44 b | 2.775 | 0.0032 | 0.0041 | 0.2397 |
| Ileum | 15.77 A | 15.58 A | 25.65 B | 17.83 A | 1.698 | 0.0009 | 0.0229 | 0.0295 |
| In Vitro Solubility, % | Dietary Treatment 1 | SE | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | ZnGly | PF | PF + ZnGly | Zn | Fibre | Zn × Fibre | ||
| In vitro phase | Soluble Zn, % | |||||||
| Gastric | 104.5 | 109.5 | 108.0 | 114.8 | 6.055 | 0.3376 | 0.4766 | 0.8778 |
| Small intestine | 3.229 | 2.729 | 1.957 | 3.771 | 1.201 | 0.5893 | 0.9250 | 0.3449 |
| Large intestine | 119.1 | 120.1 | 110.5 | 124.0 | 7.032 | 0.3102 | 0.7395 | 0.3804 |
| In vitro phase | Soluble Cu, % | |||||||
| Gastric | 99.37 ab | 110.9 b | 89.99 a | 93.63 a | 3.986 | 0.0665 | 0.0022 | 0.3319 |
| Small intestine | 85.52 | 94.93 | 79.28 | 95.40 | 8.226 | 0.1293 | 0.7280 | 0.6857 |
| Large intestine | 95.14 ab | 112.6 b | 87.99 a | 100.4 ab | 5.668 | 0.0119 | 0.0949 | 0.6549 |
| In vitro phase | Soluble Fe, % | |||||||
| Gastric | 72.24 C | 60.64 A | 66.84 B | 68.34 BC | 1.558 | 0.0031 | 0.4643 | 0.0002 |
| Small intestine | 4.022 | 2.789 | 3.644 | 4.211 | 0.933 | 0.7233 | 0.5796 | 0.3420 |
| Large intestine | 101.4 B | 78.89 A | 80.42 A | 82.05 A | 4.144 | 0.0161 | 0.0381 | 0.0059 |
| In vitro phase | Soluble Mn, % | |||||||
| Gastric | 90.49 a | 100.5 b | 88.30 a | 96.59 ab | 2.386 | 0.0006 | 0.2096 | 0.7189 |
| Small intestine | 11.06 | 12.10 | 9.613 | 12.60 | 1.586 | 0.2143 | 0.7667 | 0.5434 |
| Large intestine | 97.88 | 103.9 | 101.3 | 103.0 | 3.865 | 0.3269 | 0.7445 | 0.5815 |
| In Vitro, In Vivo Assay | Dietary Treatment | SE | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | ZnGly | PF | PF + ZnGly | Zn | Fibre | Zn × Fibre | ||
| In vitro | pH | |||||||
| Gastric phase | 2.005 a | 2.010 a | 2.070 ab | 2.090 b | 0.016 | 0.4669 | 0.0022 | 0.6589 |
| Small intestinal phase | 6.753 | 6.750 | 6.753 | 6.737 | 0.004 | 0.0359 | 0.1232 | 0.1232 |
| Large intestinal phase | 4.773 A | 4.793 B | 4.793 B | 4.773 A | 0.003 | 0.9999 | 0.9999 | 0.0001 |
| In vivo | pH | |||||||
| Duodenum digesta | 4.385 | 4.506 | 4.178 | 4.858 | 0.2749 | 0.1622 | 0.7946 | 0.3224 |
| Jejunum digesta | 5.505 | 5.160 | 5.378 | 5.505 | 0.1401 | 0.4449 | 0.4449 | 0.1078 |
| Ileum digesta | 7.325 | 6.818 | 7.257 | 6.867 | 0.2709 | 0.1135 | 0.9709 | 0.8317 |
| Dry matter (DM) | DM digestibility, % | |||||||
| In vivo assay | 83.17 a | 83.21 a | 84.67 ab | 85.31 b | 0.3841 | 0.3900 | <0.0001 | 0.4502 |
| In vitro assay | 74.78 | 73.93 | 75.35 | 76.25 | 2.404 | 0.9746 | 0.5776 | 0.7392 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokarčíková, K.; Čobanová, K.; Takácsová, M.; Barszcz, M.; Taciak, M.; Tuśnio, A.; Grešaková, Ľ. Trace Mineral Solubility and Digestibility in the Small Intestine of Piglets Are Affected by Zinc and Fibre Sources. Agriculture 2022, 12, 517. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12040517
Tokarčíková K, Čobanová K, Takácsová M, Barszcz M, Taciak M, Tuśnio A, Grešaková Ľ. Trace Mineral Solubility and Digestibility in the Small Intestine of Piglets Are Affected by Zinc and Fibre Sources. Agriculture. 2022; 12(4):517. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12040517
Chicago/Turabian StyleTokarčíková, Katarína, Klaudia Čobanová, Margaréta Takácsová, Marcin Barszcz, Marcin Taciak, Anna Tuśnio, and Ľubomíra Grešaková. 2022. "Trace Mineral Solubility and Digestibility in the Small Intestine of Piglets Are Affected by Zinc and Fibre Sources" Agriculture 12, no. 4: 517. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12040517