
Applying Silver Nanoparticles to Enhance Metabolite Accumulation and Biodiesel Production in New Algal Resources
 ,
,  ,
, 
Abstract
:1. Introduction
2. Material and Methodology
2.1. Sampling and Identification of Algae
2.2. Response Surface Methodology (RSM) Design
2.3. Quantification of Silver Uptake Using Atomic Absorption Spectrophotometry
2.4. Algal Growth
2.5. Biochemical Profiling
2.6. Transesterification and FAME Analysis via GC-MS
2.7. Fuel Properties Determined from the Fatty Acid Profile Generated from GCMS
2.8. Statistical Analyses
3. Results and Discussion
3.1. Molecular Identification of Algae
3.2. Bioremediation of Silver Nanoparticles by Algal Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor 1 | Factor 2 | Response 1 | Response 2 | Response 3 | Response 4 | ||
|---|---|---|---|---|---|---|---|
| Std | Run | A: Silver Nanoparticle | B: Days | Oedogonium sp. | Ulothrix sp. | Cladophora sp. | Spirogyra sp. |
| mg/L | mg/g | mg/g | mg/g | mg/g | |||
| 5 | 1 | 0.05 | 5 | 0.00105 | 0.00114 | 0.00111 | 0.00109 |
| 13 | 2 | 0.02 | 3 | 0.00053 | 0.00062 | 0.00059 | 0.00057 |
| 2 | 3 | 0.02 | 5 | 0.00072 | 0.00081 | 0.00078 | 0.00076 |
| 9 | 4 | 0.02 | 7 | 0.00094 | 0.00102 | 0.00099 | 0.00097 |
| 11 | 5 | 0.05 | 5 | 0.00103 | 0.00114 | 0.00111 | 0.00109 |
| 6 | 6 | 0.08 | 7 | 0.00137 | 0.00149 | 0.00144 | 0.00142 |
| 3 | 7 | 0.02 | 5 | 0.00072 | 0.00081 | 0.00078 | 0.00076 |
| 12 | 8 | 0.05 | 5 | 0.00105 | 0.00114 | 0.00111 | 0.00109 |
| 7 | 9 | 0.08 | 3 | 0.00093 | 0.00105 | 0.00101 | 0.00099 |
| 10 | 10 | 0.05 | 8 | 0.00122 | 0.00135 | 0.00131 | 0.00129 |
| 8 | 11 | 0.05 | 3 | 0.00074 | 0.00085 | 0.00081 | 0.00078 |
| 1 | 12 | 0.05 | 5 | 0.00105 | 0.00114 | 0.00111 | 0.00109 |
| 4 | 13 | 0.05 | 5 | 0.00105 | 0.00114 | 0.00111 | 0.00109 |
3.3. Effect of Silver Nanoparticles on Algal Growth
| Std | Run | Factor 1 | Factor 2 | Algal Strain | Response 1 | Response 2 | Response 3 | Response 4 | Response 5 |
|---|---|---|---|---|---|---|---|---|---|
| A: Silver Nanoparticle (mg/L) | B: Days | Growth (g FW) | Carbohydrates (mg/g) | Proteins (mg/g) | Chlorophyll (µg/gfw) | Lipids (%) | |||
| 8 | 1 | 0 | 1 | Oedogonium sp. | 5 | 0.15 | 0.019 | 40 | 46 |
| Ulothrix sp. | 5 | 0.14 | 0.020 | 34 | 50 | ||||
| Cladophora sp. | 5 | 0.17 | 0.022 | 38 | 48 | ||||
| Spirogyra sp. | 5 | 0.2 | 0.017 | 35 | 44 | ||||
| 9 | 2 | 0.02 | 3 | Oedogonium sp. | 5.3 | 0.16 | 0.021 | 39 | 48 |
| Ulothrix sp. | 6 | 0.17 | 0.022 | 33 | 51 | ||||
| Cladophora sp. | 6.2 | 0.20 | 0.025 | 37 | 50 | ||||
| Spirogyra sp. | 5.1 | 0.24 | 0.020 | 35 | 45 | ||||
| 1 | 3 | 0.02 | 5 | Oedogonium sp. | 3.3 | 0.18 | 0.023 | 34 | 49 |
| Ulothrix sp. | 4.4 | 0.16 | 0.024 | 27 | 53 | ||||
| Cladophora sp. | 3.6 | 0.2 | 0.026 | 32 | 51 | ||||
| Spirogyra sp. | 3.3 | 0.23 | 0.021 | 29 | 47 | ||||
| 7 | 4 | 0.02 | 7 | Oedogonium sp. | 2.7 | 0.21 | 0.026 | 29 | 53 |
| Ulothrix sp. | 3.5 | 0.19 | 0.027 | 23 | 56 | ||||
| Cladophora sp. | 2.7 | 0.23 | 0.028 | 27 | 54 | ||||
| Spirogyra sp. | 2.4 | 0.26 | 0.024 | 25 | 51 | ||||
| 4 | 5 | 0.05 | 5 | Oedogonium sp. | 3.0 | 0.45 | 0.031 | 32 | 67 |
| Ulothrix sp. | 4 | 0.41 | 0.032 | 25 | 71 | ||||
| Cladophora sp. | 3.2 | 0.45 | 0.034 | 30 | 69 | ||||
| Spirogyra sp. | 2.9 | 0.48 | 0.029 | 27 | 65 | ||||
| 11 | 6 | 0.08 | 7 | Oedogonium sp. | 1.8 | 0.49 | 0.038 | 19 | 74 |
| Ulothrix sp. | 2.3 | 0.47 | 0.039 | 13 | 78 | ||||
| Cladophora sp. | 1.5 | 0.51 | 0.042 | 17 | 76 | ||||
| Spirogyra sp. | 1.3 | 0.54 | 0.036 | 15 | 72 | ||||
| 12 | 7 | 0.02 | 5 | Oedogonium sp. | 3.3 | 0.18 | 0.023 | 34 | 52 |
| Ulothrix sp. | 4.2 | 0.16 | 0.024 | 27 | 56 | ||||
| Cladophora sp. | 3.4 | 0.2 | 0.026 | 32 | 54 | ||||
| Spirogyra sp. | 3.3 | 0.23 | 0.021 | 29 | 49 | ||||
| 13 | 8 | 0.05 | 5 | Oedogonium sp. | 3 | 0.43 | 0.031 | 32 | 67 |
| Ulothrix sp. | 4 | 0.41 | 0.032 | 25 | 71 | ||||
| Cladophora sp. | 3.2 | 0.45 | 0.034 | 30 | 69 | ||||
| Spirogyra sp. | 2.9 | 0.48 | 0.029 | 27 | 65 | ||||
| 10 | 9 | 0.08 | 3 | Oedogonium sp. | 3.8 | 0.47 | 0.036 | 21 | 61 |
| Ulothrix sp. | 4.8 | 0.45 | 0.037 | 15 | 65 | ||||
| Cladophora sp. | 4 | 0.49 | 0.039 | 19 | 63 | ||||
| Spirogyra sp. | 3.7 | 0.51 | 0.034 | 17 | 59 | ||||
| 6 | 10 | 0.05 | 8 | Oedogonium sp. | 2.2 | 0.45 | 0.034 | 28 | 68 |
| Ulothrix sp. | 3.2 | 0.43 | 0.035 | 21 | 72 | ||||
| Cladophora sp. | 2.4 | 0.47 | 0.037 | 26 | 70 | ||||
| Spirogyra sp. | 2.1 | 0.49 | 0.032 | 23 | 66 | ||||
| 3 | 11 | 0.05 | 3 | Oedogonium sp. | 4.3 | 0.4 | 0.029 | 33 | 64 |
| Ulothrix sp. | 5.5 | 0.38 | 0.030 | 27 | 68 | ||||
| Cladophora sp. | 4.7 | 0.42 | 0.033 | 31 | 66 | ||||
| Spirogyra sp. | 4.4 | 0.45 | 0.027 | 29 | 62 | ||||
| 5 | 12 | 0.05 | 5 | Oedogonium sp. | 3 | 0.43 | 0.031 | 32 | 67 |
| Ulothrix sp. | 4 | 0.41 | 0.032 | 25 | 71 | ||||
| Cladophora sp. | 3.2 | 0.45 | 0.034 | 30 | 69 | ||||
| Spirogyra sp. | 2.9 | 0.48 | 0.029 | 27 | 65 | ||||
| 2 | 13 | 0.05 | 5 | Oedogonium sp. | 3 | 0.43 | 0.031 | 32 | 67 |
| Ulothrix sp. | 4 | 0.41 | 0.032 | 25 | 71 | ||||
| Cladophora sp. | 3.2 | 0.45 | 0.034 | 30 | 69 | ||||
| Spirogyra sp. | 2.9 | 0.48 | 0.029 | 27 | 65 |
3.4. The Effect of Silver Nanoparticles on the Carbohydrates
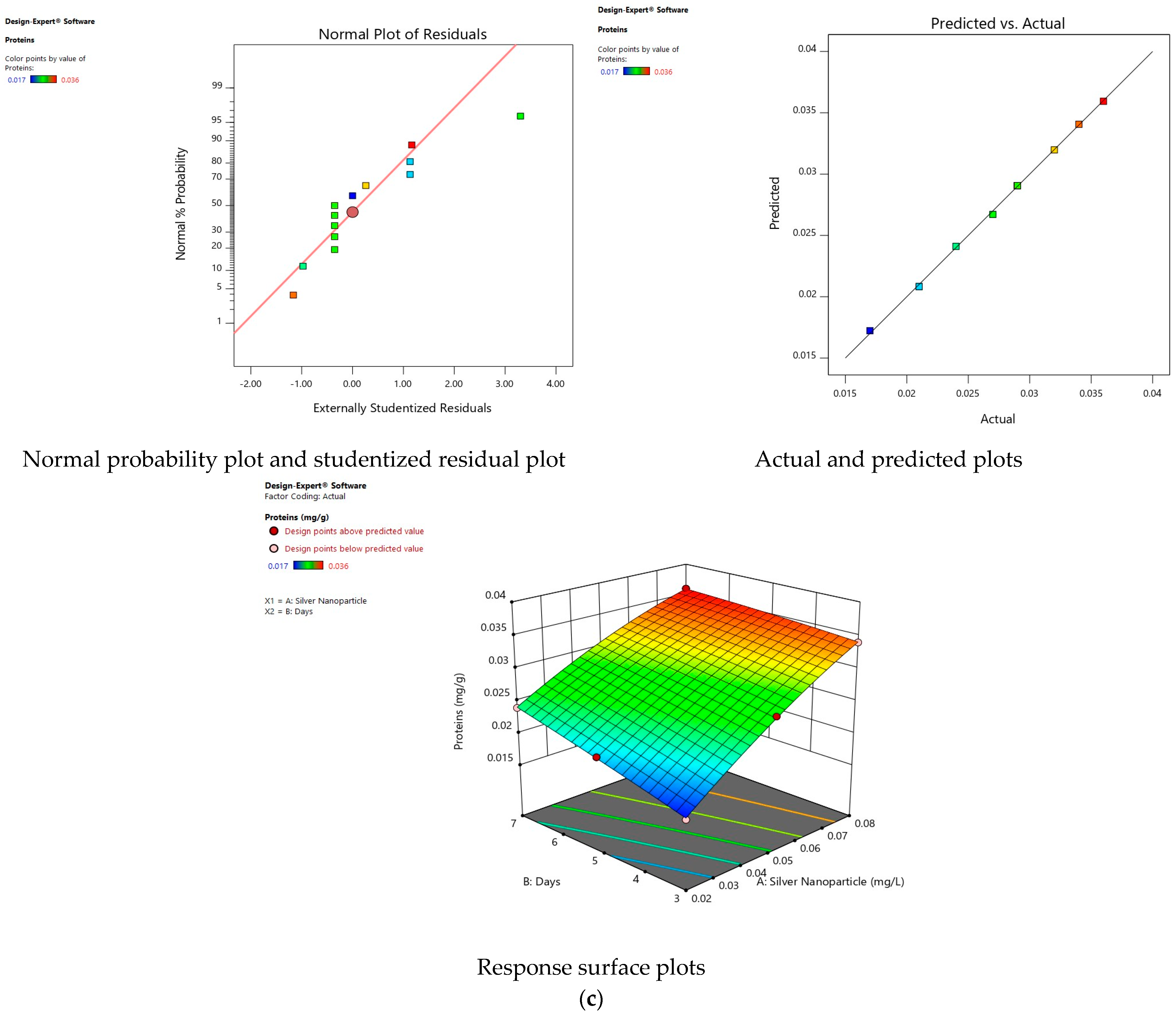
3.5. Effect of Silver Nanoparticles on Proteins
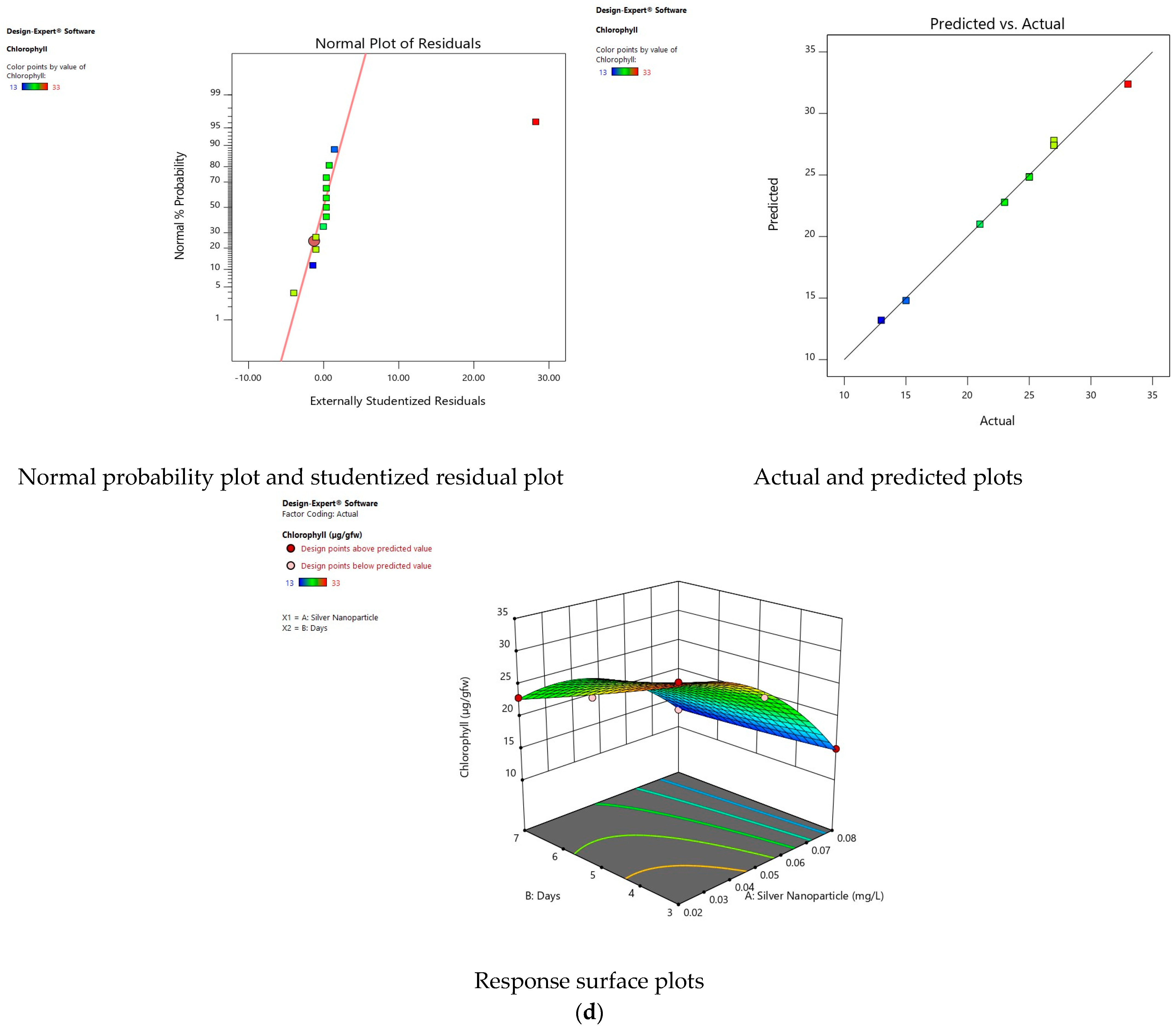
3.6. Effect of Silver Nanoparticles on Chlorophyll
3.7. Effect of Silver Nanoparticles on Lipids
3.8. Analysis of Variance (ANOVA)
3.9. Fit Statistic
3.10. Equations in Terms of the Coded Equation
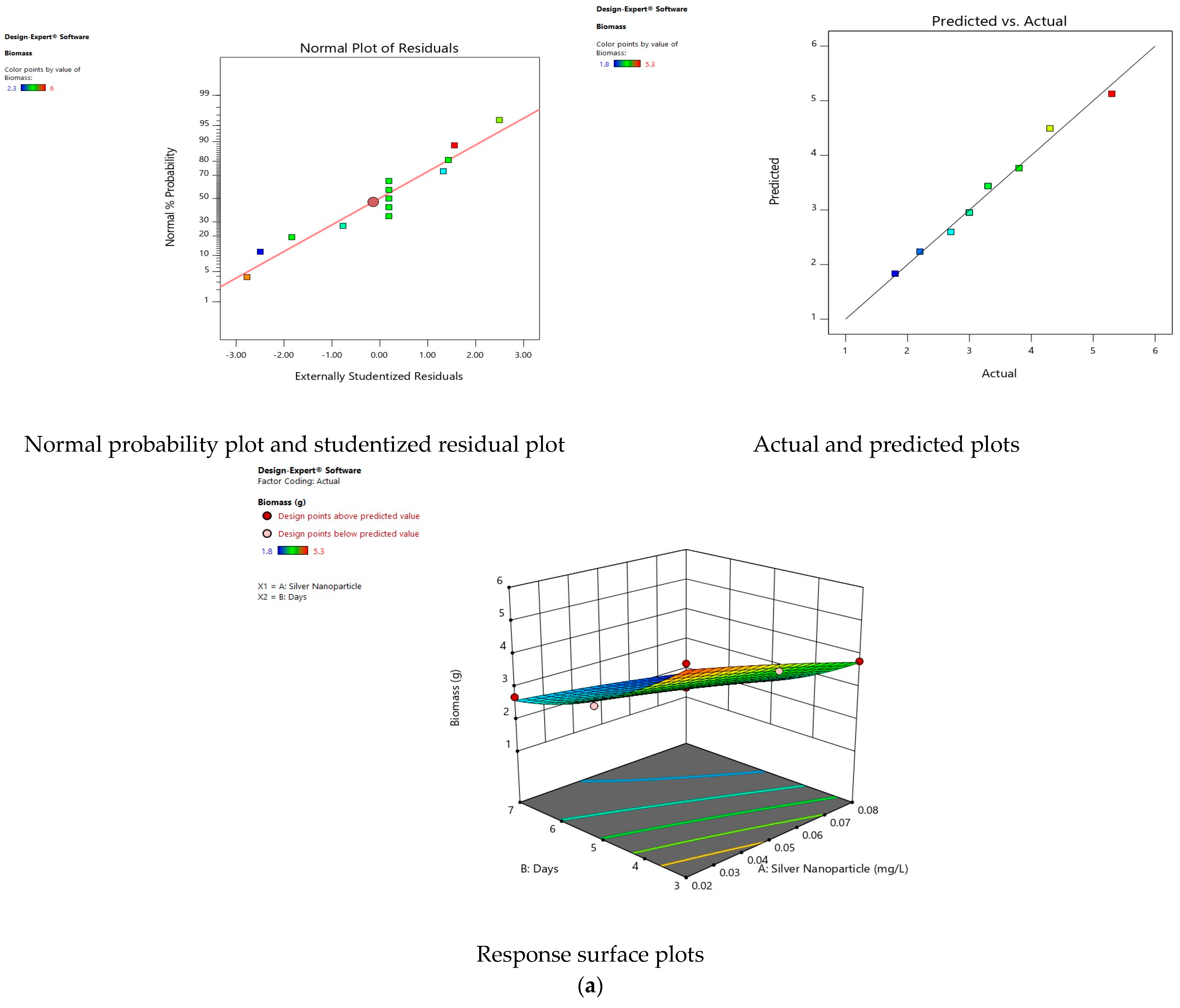
3.11. Validity of Models
3.12. Interaction of Factors on Response
3.13. Fatty Acid Profile via GCMS
3.14. Algal Fuel Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bologna, M.; Aquino, G. Deforestation and World Population Sustainability: A Quantitative Analysis. Sci. Rep. 2020, 10, 7631. [Google Scholar] [CrossRef] [PubMed]
- Hasnain, M.; Munir, N.; Siddiqui, Z.S.; Ali, F.; El-Keblawy, A.; Abideen, Z. Integral Approach for the Evaluation of Sugar Cane Bio-Waste Molasses and Effects on Algal Lipids and Biodiesel Production. Waste Biomass Valorization 2022, 1–20. [Google Scholar] [CrossRef]
- Hasnain, M.; Abideen, Z.; Naz, S.; Roessner, U.; Munir, N. Biodiesel Production from New Algal Sources Using Response Surface Methodology and Microwave Application. Biomass Convers. Biorefinery 2021. [Google Scholar] [CrossRef]
- Abideen, Z.; Waqif, H.; Munir, N.; El-Keblawy, A.; Hasnain, M.; Radicetti, E.; Mancinelli, R.; Nielsen, B.L.; Haider, G. Algal-Mediated Nanoparticles, Phycochar, and Biofertilizers for Mitigating Abiotic Stresses in Plants: A Review. Agronomy 2022, 12, 1788. [Google Scholar] [CrossRef]
- Munir, N.; Hasnain, M.; Waqif, H.; Adetuyi, B.O.; Egbuna, C.; Olisah, M.C.; Chikwendu, C.J.; Uche, C.Z.; C Patrick-Iwuanyanwu, K.; El Sayed, A.M.A. Gelling Agents, Micro and Nanogels in Food System Applications. In Application of Nanotechnology in Food Science, Processing and Packaging; Springer: Berlin/Heidelberg, Germany, 2022; pp. 153–167. [Google Scholar]
- Munir, N.; Hasnain, M.; Hanif, M.; Waqif, H.; Sharif, N. Fungal Organisms: A Check for Harmful Algal Blooms. In Freshwater Mycology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 91–115. [Google Scholar]
- Qidwai, A.; Kumar, R.; Dikshit, A. Green Synthesis of Silver Nanoparticles by Seed of Phoenix sylvestris L. and Their Role in the Management of Cosmetics Embarrassment. Green Chem. Lett. Rev. 2018, 11, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Cheng, J.; Yi, J. Impact of Silver Nanoparticles on Marine Diatom Skeletonema costatum. J. Appl. Toxicol. 2016, 36, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M.; Bedaiwy, M.Y.; Osman, M.E.; Ismail, M.M. Influence of Molasses on Growth, Biochemical Composition and Ethanol Production of the Green Algae Chlorella vulgaris and Scenedesmus obliquus. J. Agric. Eng. Biotechnol. 2014, 2, 20. [Google Scholar] [CrossRef]
- Kumar, D.; Abdul Rub, M.; Akram, M.; Kabir-ud-Din. Interaction of Chromium (III) Complex of Glycylphenylalanine with Ninhydrin in Aqueous and Cetyltrimethylammonium Bromide (CTAB) Micellar Media. Tenside Surfactants Deterg. 2014, 51, 157–163. [Google Scholar] [CrossRef]
- Isaac, R.A.; Kerber, J.D. Atomic Absorption and Flame Photometry: Techniques and Uses in Soil, Plant, and Water Analysis. In Instrumental Methods for Analysis of Soils and Plant Tissue; Wiley: Hoboken, NJ, USA, 1971; pp. 17–37. [Google Scholar]
- Hasnain, M.; Abideen, Z.; Hashmi, S.; Naz, S.; Munir, N. Assessing the Potential of Nutrient Deficiency for Enhancement of Biodiesel Production in Algal Resources. Biofuels 2022, 1–34. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4-3. [Google Scholar] [CrossRef]
- Munir, N.; Sharif, N.; Naz, S.; Saleem, F.; Manzoor, F. Harvesting and Processing of Microalgae Biomass Fractions for Biodiesel Production (a Review). Sci. Technol. Dev. 2016, 32, 235–243. [Google Scholar]
- Munir, N.; Hasnain, M.; Sarwar, Z.; Ali, F.; Hessini, K.; Abideen, Z. Changes in Environmental Conditions Are Critical Factors for Optimum Biomass, Lipid Pattern and Biodiesel Production in Algal Biomass. Biologia 2022, 77, 3099–3124. [Google Scholar] [CrossRef]
- Hasnain, M.; Abideen, Z.; Anthony Dias, D.; Naz, S.; Munir, N. Utilization of Saline Water Enhances Lipid Accumulation in Green Microalgae for the Sustainable Production of Biodiesel. BioEnergy Res. 2022, 1–14. [Google Scholar] [CrossRef]
- Borah, D.; Das, N.; Das, N.; Bhattacharjee, A.; Sarmah, P.; Ghosh, K.; Chandel, M.; Rout, J.; Pandey, P.; Ghosh, N.N. Alga-mediated Facile Green Synthesis of Silver Nanoparticles: Photophysical, Catalytic and Antibacterial Activity. Appl. Organomet. Chem. 2020, 34, e5597. [Google Scholar] [CrossRef]
- Fatima, R.; Priya, M.; Indurthi, L.; Radhakrishnan, V.; Sudhakaran, R. Biosynthesis of Silver Nanoparticles Using Red Algae Portieria hornemannii and Its Antibacterial Activity against Fish Pathogens. Microb. Pathog. 2020, 138, 103780. [Google Scholar] [CrossRef]
- Alsammarraie, F.K.; Wang, W.; Zhou, P.; Mustapha, A.; Lin, M. Green Synthesis of Silver Nanoparticles Using Turmeric Extracts and Investigation of Their Antibacterial Activities. Colloids Surf. B Biointerfaces 2018, 171, 398–405. [Google Scholar] [CrossRef]
- Fazelian, N.; Movafeghi, A.; Yousefzadi, M.; Rahimzadeh, M.; Zarei, M. Impact of Silver Nanoparticles on the Growth, Fatty Acid Profile, and Antioxidative Response of Nannochloropsis oculata. Acta Physiol. Plant. 2020, 42, 126. [Google Scholar] [CrossRef]
- Ghazaei, F.; Shariati, M. Effects of Titanium Nanoparticles on the Photosynthesis, Respiration, and Physiological Parameters in Dunaliella salina and Dunaliella tertiolecta. Protoplasma 2020, 257, 75–88. [Google Scholar] [CrossRef]
- He, M.; Yan, Y.; Pei, F.; Wu, M.; Gebreluel, T.; Zou, S.; Wang, C. Improvement on Lipid Production by Scenedesmus obliquus Triggered by Low Dose Exposure to Nanoparticles. Sci. Rep. 2017, 7, 15526. [Google Scholar] [CrossRef] [Green Version]
- Saied, E.; Eid, A.M.; Hassan, S.E.-D.; Salem, S.S.; Radwan, A.A.; Halawa, M.; Saleh, F.M.; Saad, H.A.; Saied, E.M.; Fouda, A. The Catalytic Activity of Biosynthesized Magnesium Oxide Nanoparticles (MgO-NPs) for Inhibiting the Growth of Pathogenic Microbes, Tanning Effluent Treatment, and Chromium Ion Removal. Catalysts 2021, 11, 821. [Google Scholar] [CrossRef]
- Shanab, S.M.M.; Partila, A.M.; Ali, H.E.A.; Abdullah, M.A. Impact of Gamma-Irradiated Silver Nanoparticles Biosynthesized from Pseudomonas Aeruginosa on Growth, Lipid, and Carbohydrates of Chlorella vulgaris and Dictyochloropsis splendida. J. Radiat. Res. Appl. Sci. 2020, 14, 70–81. [Google Scholar] [CrossRef]
- Razack, S.A.; Duraiarasan, S.; Mani, V. Biosynthesis of Silver Nanoparticle and Its Application in Cell Wall Disruption to Release Carbohydrate and Lipid from C. vulgaris for Biofuel Production. Biotechnol. Rep. 2016, 11, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Wang, Y.; Huang, W.; Xu, J.; Wang, Z.; Xu, J.; Yuan, Z. Enhanced Accumulation of Carbohydrate and Starch in Chlorella zofingiensis Induced by Nitrogen Starvation. Appl. Biochem. Biotechnol. 2014, 174, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qiu, Y.; He, L.; Luo, K.; Wang, Z. Effect of Iron and Phosphorus on the Microalgae Growth in Co-Culture. Arch. Microbiol. 2020, 203, 733–740. [Google Scholar] [CrossRef]
- Romero, N.; Visentini, F.F.; Márquez, V.E.; Santiago, L.G.; Castro, G.R.; Gagneten, A.M. Physiological and Morphological Responses of Green Microalgae Chlorella vulgaris to Silver Nanoparticles. Environ. Res. 2020, 189, 109857. [Google Scholar] [CrossRef]
- Deng, X.-Y.; Cheng, J.; Hu, X.-L.; Wang, L.; Li, D.; Gao, K. Biological Effects of TiO2 and CeO2 Nanoparticles on the Growth, Photosynthetic Activity, and Cellular Components of a Marine Diatom Phaeodactylum tricornutum. Sci. Total Environ. 2017, 575, 87–96. [Google Scholar] [CrossRef]
- Marchello, A.E.; Barreto, D.M.; Lombardi, A.T. Effects of Titanium Dioxide Nanoparticles in Different Metabolic Pathways in the Freshwater Microalga Chlorella Sorokiniana (Trebouxiophyceae). Water Air Soil Pollut. 2018, 229, 48. [Google Scholar] [CrossRef]
- Johari, S.A.; Sarkheil, M.; Tayemeh, M.B.; Veisi, S. Influence of Salinity on the Toxicity of Silver Nanoparticles (AgNPs) and Silver Nitrate (AgNO3) in Halophilic Microalgae, Dunaliella salina. Chemosphere 2018, 209, 156–162. [Google Scholar] [CrossRef]
- Peng, X.; Palma, S.; Fisher, N.S.; Wong, S.S. Effect of Morphology of ZnO Nanostructures on Their Toxicity to Marine Algae. Aquat. Toxicol. 2011, 102, 186–196. [Google Scholar] [CrossRef]
- Da Costa, C.H.; Perreault, F.; Oukarroum, A.; Melegari, S.P.; Popovic, R.; Matias, W.G. Effect of Chromium Oxide (III) Nanoparticles on the Production of Reactive Oxygen Species and Photosystem II Activity in the Green Alga Chlamydomonas reinhardtii. Sci. Total Environ. 2016, 565, 951–960. [Google Scholar] [CrossRef]
- Pirzadah, T.B.; Malik, B.; Salam, S.T.; Ahmad Dar, P.; Rashid, S. Impact of Heavy Metal Stress on Plants and the Role of Various Defense Elements. Iran. J. Plant Physiol. 2019, 9, 2883–2900. [Google Scholar]
- Kaliamurthi, S.; Selvaraj, G.; Cakmak, Z.E.; Korkmaz, A.D.; Cakmak, T. The Relationship between Chlorella sp. and Zinc Oxide Nanoparticles: Changes in Biochemical, Oxygen Evolution, and Lipid Production Ability. Process Biochem. 2019, 85, 43–50. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; Hussein, M.H.; Shaaban-Dessuuki, S.A.; Dalal, S.R. Production, Extraction and Characterization of Chlorella vulgaris Soluble Polysaccharides and Their Applications in AgNPs Biosynthesis and Biostimulation of Plant Growth. Sci. Rep. 2020, 10, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shariati, F.; Ayatallahzadeh Shirazi, M. Effect of SiO2 Nanoparticles on Chlorophyll, Carotenoid and Growth of Green Micro-Algae Dunaliella salina. Nanomedicine Res. J. 2019, 4, 164–175. [Google Scholar]
- Sandhya, S.V.; Vijayan, K.K. Biogenesis of Silver Nanoparticles by Marine Bacteria Labrenzia sp. Mab 26 Associated with Isochrysis galbana. Curr. Sci. 2020, 119, 1830–1833. [Google Scholar] [CrossRef]
- Al-Khazali, Z.K.M.; Alghanmi, H.A. Influence of Different Concentrations of Nano-Magnesium Oxide on the Growth of Coelastrella terrestris. J. Phys. Conf. Ser. 2019, 1234, 12070. [Google Scholar] [CrossRef]
- Serif, M.; Dubois, G.; Finoux, A.-L.; Teste, M.-A.; Jallet, D.; Daboussi, F. One-Step Generation of Multiple Gene Knock-Outs in the Diatom Phaeodactylum tricornutum by DNA-Free Genome Editing. Nat. Commun. 2018, 9, 3924. [Google Scholar] [CrossRef] [Green Version]
- Patil, P.D.; Gude, V.G.; Mannarswamy, A.; Deng, S.; Cooke, P.; Munson-McGee, S.; Rhodes, I.; Lammers, P.; Nirmalakhandan, N. Optimization of Direct Conversion of Wet Algae to Biodiesel under Supercritical Methanol Conditions. Bioresour. Technol. 2011, 102, 118–122. [Google Scholar] [CrossRef]
- Hussain, J.; Wang, X.; Sousa, L.; Ali, R.; Rittmann, B.E.; Liao, W. Using Non-Metric Multi-Dimensional Scaling Analysis and Multi-Objective Optimization to Evaluate Green Algae for Production of Proteins, Carbohydrates, Lipids, and Simultaneously Fix Carbon Dioxide. Biomass Bioenergy 2020, 141, 105711. [Google Scholar] [CrossRef]
- Yin, Z.; Zhu, L.; Li, S.; Hu, T.; Chu, R.; Mo, F.; Hu, D.; Liu, C.; Li, B. A Comprehensive Review on Cultivation and Harvesting of Microalgae for Biodiesel Production: Environmental Pollution Control and Future Directions. Bioresour. Technol. 2020, 301, 122804. [Google Scholar] [CrossRef]
- Hauser-Davis, R.A.; Lavradas, R.T.; Monteiro, F.; Rocha, R.C.C.; Bastos, F.F.; Araújo, G.F.; Júnior, S.F.S.; Bordon, I.C.; Correia, F.V.; Saggioro, E.M. Biochemical Metal Accumulation Effects and Metalloprotein Metal Detoxification in Environmentally Exposed Tropical Perna Perna Mussels. Ecotoxicol. Environ. Saf. 2021, 208, 111589. [Google Scholar] [CrossRef]
- Ren, H.-Y.; Dai, Y.-Q.; Kong, F.; Xing, D.; Zhao, L.; Ren, N.-Q.; Ma, J.; Liu, B.-F. Enhanced Microalgal Growth and Lipid Accumulation by Addition of Different Nanoparticles under Xenon Lamp Illumination. Bioresour. Technol. 2020, 297, 122409. [Google Scholar] [CrossRef] [PubMed]
- Kadar, E.; Rooks, P.; Lakey, C.; White, D.A. The Effect of Engineered Iron Nanoparticles on Growth and Metabolic Status of Marine Microalgae Cultures. Sci. Total Environ. 2012, 439, 8–17. [Google Scholar] [CrossRef]
- Rocha, A.A.; Wilde, C.; Hu, Z.; Nepotchatykh, O.; Nazarenko, Y.; Ariya, P.A. Development of a Hybrid Photo-Bioreactor and Nanoparticle Adsorbent System for the Removal of CO2, and Selected Organic and Metal Co-Pollutants. J. Environ. Sci. 2017, 57, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Katare, A.K.; Singh, B.; Shukla, P.; Gupta, S.; Singh, B.; Yalamanchili, K.; Kulshrestha, N.; Bhanwaria, R.; Sharma, A.K.; Sharma, S. Rapid Determination and Optimisation of Berberine from Himalayan Berberis lycium by Soxhlet Apparatus Using CCD-RSM and Its Quality Control as a Potential Candidate for COVID-19. Nat. Prod. Res. 2020, 36, 868–873. [Google Scholar] [CrossRef]
- Aydın, M.; Uslu, S.; Çelik, M.B. Performance and Emission Prediction of a Compression Ignition Engine Fueled with Biodiesel-Diesel Blends: A Combined Application of ANN and RSM Based Optimization. Fuel 2020, 269, 117472. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Zabed, H.M.; Qi, X.; El-Shenody, R.A. Enhancing Biomass and Lipid Productivity of a Green Microalga Parachlorella kessleri for Biodiesel Production Using Rapid Mutation of Atmospheric and Room Temperature Plasma. Biotechnol. Biofuels Bioprod. 2022, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Basheer, S.; Liu, F.; Elshobary, M.; Zhang, C.; Qian, J.; Xu, L.; Arslan, M.; Cui, F.; Zan, X. Bacterial Intervention on the Growth, Nutrient Removal and Lipid Production of Filamentous oleaginous Microalgae Tribonema sp. Algal Res. 2020, 52, 102088. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Abo-Shady, A.M.; Khairy, H.M.; Essa, D.; Zabed, H.M.; Qi, X.; Abomohra, A.E.-F. Influence of Nutrient Supplementation and Starvation Conditions on the Biomass and Lipid Productivities of Micractinium reisseri Grown in Wastewater for Biodiesel Production. J. Environ. Manage. 2019, 250, 109529. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, J.; Pei, H.; Pan, J.; Jiang, L.; Hou, Q.; Han, F. Cultivation of Microalgae Using Anaerobically Digested Effluent from Kitchen Waste as a Nutrient Source for Biodiesel Production. Renew. Energy 2018, 115, 276–287. [Google Scholar] [CrossRef]
- El Shafay, S.M.; Gaber, A.; Alsanie, W.F.; Elshobary, M.E. Influence of Nutrient Manipulation on Growth and Biochemical Constituent in Anabaena variabilis and Nostoc muscorum to Enhance Biodiesel Production. Sustainability 2021, 13, 9081. [Google Scholar] [CrossRef]





















| Algae | Variable | R2 | Adjusted R2 | Adeq Precision | Std. Dev. | Mean | C.V. % |
|---|---|---|---|---|---|---|---|
| Oedogonium sp. | Growth | 0.9862 | 0.9763 | 35.5425 | 0.1364 | 3.21 | 4.25 |
| Carbohydrates | 0.9980 | 0.9965 | 67.2033 | 0.0076 | 0.3615 | 2.09 | |
| Proteins | 0.9993 | 0.9989 | 151.171 | 0.0002 | 0.0295 | 0.61 | |
| Chlorophyll | 0.9925 | 0.9872 | 45.9952 | 0.6074 | 30.54 | 1.99 | |
| Lipids | 0.9811 | 0.9676 | 24.4970 | 1.57 | 61.69 | 2.55 | |
| Ulothrix sp. | Growth | 0.9956 | 0.9924 | 64.6544 | 0.0819 | 4.15 | 1.97 |
| Carbohydrates | 0.9974 | 0.9955 | 58.3222 | 0.0085 | 0.3423 | 2.50 | |
| Proteins | 0.9993 | 0.9989 | 0.0002 | 0.0305 | 151.17 | 0.59 | |
| Chlorophyll | 0.9950 | 0.9914 | 58.4744 | 0.4830 | 23.92 | 2.02 | |
| Lipids | 0.9842 | 0.9728 | 26.4956 | 1.45 | 65.62 | 2.21 | |
| Cladophora sp. | Growth | 0.9743 | 0.9560 | 26.7868 | 0.2364 | 3.42 | 6.90 |
| Carbohydrates | 0.9980 | 0.9965 | 67.2033 | 0.0076 | 0.3815 | 1.98 | |
| Proteins | 0.9946 | 0.9907 | 51.6865 | 0.0005 | 0.0325 | 1.67 | |
| Chlorophyll | 0.9925 | 0.9872 | 45.9952 | 0.6074 | 28.54 | 2.13 | |
| Lipids | 0.9842 | 0.9728 | 26.4956 | 1.45 | 63.62 | 2.28 | |
| Spirogyra sp. | Growth | 0.9966 | 0.9942 | 74.2051 | 0.0730 | 3.08 | 2.37 |
| Carbohydrates | 0.9996 | 0.9993 | 146.2298 | 0.0034 | 0.4085 | 0.84 | |
| Proteins | 0.9993 | 0.9989 | 151.1711 | 0.0002 | 0.0275 | 0.6632 | |
| Chlorophyll | 0.9950 | 0.9914 | 58.4744 | 0.4830 | 25.92 | 1.86 | |
| Lipids | 0.9833 | 0.9713 | 25.8908 | 1.49 | 59.62 | 2.51 |
| Fatty Acids | Oedogonium sp. | Ulothrix sp. | Cladophora sp. | Spirogyra sp. | ||||
|---|---|---|---|---|---|---|---|---|
| Control | AgNPs Stress | Control | AgNPs Stress | Control | AgNPs Stress | Control | AgNPs Stress | |
| Myristoleic acid (14:1) | 0.18 | 0.15 | - | - | 0.34 | 0.31 | 0.18 | 0.17 |
| Myristic acid (14:0) | 0.36 | 0.35 | 0.17 | 0.15 | 0.16 | 0.14 | 0.09 | 0.07 |
| Hexadecadienoic (16:2) | 0.48 | - | - | - | - | - | 2.26 | - |
| Palmitoleic acid (16:1) | 2.97 | 3.10 | - | - | - | - | 0.32 | 0.35 |
| Palmatic acid (16:0) | 1.16 | 2.5 | 0.26 | 1.01 | 0.44 | 0.50 | 0.11 | 0.21 |
| Linolenic acid (18:3) | 0.46 | 0.21 | - | - | - | 0.68 | 0.31 | |
| Linoleic acid (18:2) | 0.37 | 0.39 | 0.50 | 0.48 | 2.07 | 2.05 | 1.09 | 1.01 |
| Oleic acid (18:1) | 3.14 | 3.21 | 0.45 | 0.50 | - | - | 0.39 | 0.45 |
| Stearic acid (18:0) | 1.33 | 1.38 | - | - | - | - | 0.10 | 0.19 |
| Gondoic acid (20:1) | - | - | - | - | - | - | 0.19 | 0.21 |
| Arachidic acid (20:0) | - | - | - | - | - | - | 0.19 | 0.23 |
| Erucic acid (22:1) | 0.41 | - | - | - | 0.14 | - | - | - |
| Caprylate (8:0) | - | - | 0.35 | 0.35 | 0.36 | 0.36 | - | - |
| Caprate (10:0) | - | - | 0.63 | 0.63 | 0.12 | 0.12 | - | - |
| Laurate (12:0) | - | - | 0.23 | 0.23 | 0.61 | 0.60 | - | - |
| Methyl myristate (14:0) | 4.54 | 4.12 | 3.35 | 1.90 | 3.35 | 2.13 | 2.54 | 2.10 |
| Methyl palmitoleate (16:1) | 3.15 | 2.92 | 5.12 | 3.01 | 3.24 | 3.01 | 6.10 | 5.91 |
| Methyl palmitate (16:0) | 46.4 | 44.2 | 24.6 | 21.0 | 34.3 | 32.1 | 49.9 | 48.0 |
| Linolenate (18:3) | 2.33 | 2.01 | 2.64 | 1.81 | 7.60 | 6.10 | 1.19 | 1.10 |
| Linoleate (18:2) | 2.52 | 1.91 | 9.12 | 6.21 | 5.41 | 4.51 | 0.58 | 0.23 |
| Methyl oleate (18:1) | 28.1 | 26.01 | 44.5 | 37.1 | 42.2 | 33.1 | 29.9 | 28.0 |
| Methyl stearate (18:0) | 2.12 | 1.92 | 3.83 | 2.21 | - | - | 5.43 | 4.23 |
| Arachidate (21:0) | - | - | 2.43 | 2.21 | - | - | 1.17 | 1.01 |
| Erucate (22:1) | - | - | 2.23 | 1.72 | - | - | 0.58 | 0.43 |
| Behenic acid (22:0) | - | 0.09 | - | 0.07 | - | 0.10 | - | 0.14 |
| Algae | IV (g I2100/g fat) | SV (mg KOH/g) | CN | LCSF | CFPP (°C) | HHVi (MJ/kg) | Ρ (g cm−3) | ʋ (mm2/s) | Y at 110 °C (hour) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Biodiesel Standard EN 14214 | ≤120 | - | ≥51 | - | ≤5/≤−20 | - | 0.86–0.9 | 3.5–5.0 | ≥ 6 | |
| Biodiesel Standard ASTM D6751-02 | - | - | ≥47 | - | - | - | - | 1.9–6.0 | - | |
| Oedogonium sp. | Control | 110 | 202 | 48.6 | 0.77 | −14.03 | 38.4 | 0.87 | 3.6 | 114 |
| AgNPs Stress | 102 | 191 | 53 | 0.94 | −13.52 | 39.5 | 0.88 | 3.6 | 199 | |
| Ulothrix sp. | Control | 114 | 197 | 48.5 | 0.026 | −16.39 | 39.3 | 0.88 | 3.7 | 238 |
| AgNPs Stress | 89 | 158 | 61 | 0.101 | −16.15 | 40 | 0.88 | 3.8 | 248 | |
| Cladophora sp. | Control | 122 | 199 | 47 | 0.04 | −16.47 | 36 | 0.86 | 3.7 | 59 |
| AgNPs Stress | 103 | 170 | 56 | 0.05 | −16.31 | 39 | 0.88 | 3.8 | 61 | |
| Spirogyra sp. | Control | 109 | 206 | 49 | 0.25 | −15.68 | 37 | 0.86 | 3.5 | 82 |
| AgNPs Stress | 104 | 189 | 52 | 0.34 | −15.38 | 38 | 0.87 | 3.6 | 84 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasnain, M.; Munir, N.; Abideen, Z.; Dias, D.A.; Aslam, F.; Mancinelli, R. Applying Silver Nanoparticles to Enhance Metabolite Accumulation and Biodiesel Production in New Algal Resources. Agriculture 2023, 13, 73. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010073
Hasnain M, Munir N, Abideen Z, Dias DA, Aslam F, Mancinelli R. Applying Silver Nanoparticles to Enhance Metabolite Accumulation and Biodiesel Production in New Algal Resources. Agriculture. 2023; 13(1):73. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010073
Chicago/Turabian StyleHasnain, Maria, Neelma Munir, Zainul Abideen, Daniel Anthony Dias, Farheen Aslam, and Roberto Mancinelli. 2023. "Applying Silver Nanoparticles to Enhance Metabolite Accumulation and Biodiesel Production in New Algal Resources" Agriculture 13, no. 1: 73. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13010073