
Journey of Trichoderma from Pilot Scale to Mass Production: A Review

1
Department of Plant Pathology, School of Agriculture, Lovely Professional University, Phagwara 144411, Punjab, India
2
Department of Biotechnology, Lovely Professional University, Phagwara 144411, Punjab, India
3
Department of Botany, Guru Teg Bahadur Khalsa College for Women, Dasuya 144205, Punjab, India
4
Department of Botany, Lovely Professional University, Phagwara 144411, Punjab, India
5
Department of Life Science, Yeungnam University, Gyeongsan 38541, Republic of Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agriculture 2023, 13(10), 2022; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13102022
Submission received: 30 August 2023
/
Revised: 11 October 2023
/
Accepted: 13 October 2023
/
Published: 18 October 2023
(This article belongs to the Section Crop Protection, Diseases, Pests and Weeds)
Abstract
:Trichoderma spp. has the ability to inhibit fungal plant pathogens through several mechanisms like the production of hydrolytic enzymes, mycoparasitism, coiling, and antibiosis and is therefore recommended as a potential and native biocontrol agent for effective control of soil-transmitted diseases. Various species of Trichoderma, like T. virens, T. asperellum, T. harzianum, etc., have been explored for their biocontrol activity against phytopathogens. There are different Trichoderma species and strains with respect to plant pathogens. Efforts have been made to develop effective and efficient methods, such as microencapsulation use of different polymers, adjuvants, or carriers, to increase the shelf-life and efficacy of Trichoderma formulations. The crucial aspects for the success of a biocontrol agent include developing and validating formulations, improvement in shelf-life, cost-effectiveness, easy accessibility, improved delivery systems, broad spectrum in action, robust performance (biocontrol), and integrative strategies for sustainable disease management. This review focuses on recent developments in the isolation, identification, preservation, substrates, consortium, quality control, mass production, delivery methods, field performance, registration, and commercialization of Trichoderma formulations for strategic development of next-generation multifunctional biological control formulations.
1. Introduction
Biological control of plant diseases has emerged as a promising area in phytopathology. These methods not only minimize reliance on synthetic pesticides but are also comparatively economical, feasible, robust, and sustainable [1,2]. Among the commercial biofungicides/fungal antagonists, Trichoderma has been broadly acknowledged as a source of potential biocontrol agent, particularly for lowering soil-borne phytopathogens such as Fusarium oxysproum, Rhizoctonia solani [3], Macrophomina phaseolina [4], Sclerotinia rolfsii, and others [5,6,7]. Trichoderma spp. are saprophytic, avirulent, and soil-inhabiting fungi, and they control pathogens with various biocontrol methods such as mycoparasitism [8], retard the pathogen growth by secreting cell-wall-destroying enzymes [9,10], nutrient uptake competition, and rhizospheric competence [2]. Because of its effectiveness against phytopathogens, its market demand is increasing yearly [11].
According to Harman (1991), [12] key points that are required for the production of any biological control system are (1) a potential biocontrol agent, (2) viable propagule with amplified shelf-life, (3) bioprotective delivery mechanisms that may offer the biocontrol agent an advantage to compete against existing microflora, and (4) steady field performance [13]. As mentioned above, the first and most principal step before mass production of a biocontrol agent is the identification of a robust propagule (hyphae, chlamydospores, and/or conidia) [14]. Once a reliable and efficient biocontrol agent is identified, large-scale production, design, and application ideas need to be performed cautiously for the product’s stability during storage and later use. Both liquid and solid formulations are utilized in developing sufficient amounts of viable and active Trichoderma inoculums. Conidia and chlamydospores are the preferred propagules for formulations since they can withstand rigorous treatment procedures, whereas hyphae cannot be used because they are not dehydration-resistant [12,15]. Several reports (Table 1) related to the effectiveness of Trichoderma strains and their formulations have been published periodically [13]; however, reports concerning advancements in mass production, persistent viability, and related field performance of Trichoderma species are fragmentary. Furthermore, the challenges related to the isolation and development methods for elevating the effectiveness and sustainable use of Trichoderma formulations in the field of health security and food also need to be addressed. In this review, we intend to fill this gap and unravel the recent developments regarding isolation, identification, preservation, substrates, consortium, quality control, mass production, delivery approaches, field performance, and registration and commercialization of Trichoderma formulations.
2. Trichoderma Serves as a Biocontrol Agent
Trichoderma was initially reported as a biocontrol agent in the 1930s [18,19]. Since the discovery of mycoparasitic activity of Trichoderma by Weindling against Rhizoctonia solani and other phytopathogens, several other researchers have also obtained positive results with Trichoderma isolates as biocontrol agents of plant pathogens [46]. Trichoderma continues to hold a significant position among commercial biological control agents (BCAs) in a wide range of crop and disease management, either as a single ingredient or in combination with other ingredients [6,45,47,48]. Up till now, more than 80 species of Trichoderma have been reported [13], and among these, T. harzianum, T. virens, and T. viride are frequently deployed biocontrol agents. In India, commercialization of Trichoderma is limited to only two species, namely T. viride and T. harzianum [13]. However, there are ample reports on the effectiveness of T. virens and T. asperellum, but these are still unregistered under the Central Insecticide Board and Registration Committee (CIBRC) in India. This may be due to several hurdles, such as toxicity assessment, environmental effects of microbes and their formulation, and optimization of technology for mass-scale production [49]. Apart from these, some other constraints include multi-location trials for the purpose of proving its safety, followed by registration. In addition, some other challenges are inconsistent field presentation and low shelf-life of formulation, lack of patent protection, preliminary testing, high registration cost, alertness about the beneficial effect of Trichoderma formulations, and training and education shortfalls. Registration also requires good and effective documentation and other confirmations. Moreover, the evaluation process itself (compilation and analysis of data) can be prolonged and costly. In the past, high registration fees were clearly seen as a delay or barrier to BCA market growth, especially for medium and small enterprises that are the chief manufacturers of biocontrol agents. Due to these reasons, there are now diverse products in the market that claim to be plant growth promoters or biofertilizers for managing plant diseases but are not yet registered. The biocontrol activity of Trichoderma spp. is represented in Table 2.
However, their safe use cannot be guaranteed without toxicological and efficacy data [50]. Combined with the competitive pesticide industry, BCA companies are finding it hard to generate enough revenue from product sales to justify the registration cost. Unluckily, this has led to some products being removed from the stores. For instance, T. harzianum T-39 (Trichodex) was launched in Europe and Israel in 1993 for the control of Botrytis fruit rot through biological methods but was detached from the market. It went bankrupt in 2005 due to low sales and the rising cost of registration. Table 3 contains information on gene identification in Trichoderma species.

{kind=link}
{kind=link}
Table 2.
Biocontrol activity of Trichoderma spp.
| Name of Trichoderma Species | Name of Plant Pathogens | Crop Name | Inhibition/Efficacy (%) | Experiment Condition | References |
|---|---|---|---|---|---|
| T. harzianum | Sclerotium rolfsii and Rhizoctonia solani | Ryegrass | 42–47 | Greenhouse and field condition | [51] |
| T. harzianum | Phytophthora cinnamon | Pine | 28.5–37.5 | Greenhouse | [52] |
| T. hamatum and T. harzianum | P. cinnamomi | Avocado | Greenhouse | [53] | |
| T. viride, T. virens T. harzianum, T. pseudokoningii T. koningii | Aspergillus niger, Rhizoctonia solani and Geotrichum candidum | Sapodilla (Manilkara zapota L.) | 54–74 | Laboratory | [54] |
| T. harzianum strain Ths97 | F. solani | Olive trees | 25–50 | - | [55] |
| Trichoderma viride 1433 Mutant strains | Pythium aphanidermatum | Mustard | 85 | Lab and field | [56] |
| Trichoderma spp. | Pythium aphanidermatum | Chilli | 88.00 | Lab and field | [57] |
| Trichoderma harzianumRifai, Trichoderma viride | Pythium aphanidermatum | Tobacco | 100 | Lab and greenhouse | [58] |
| Trichoderma virens Trichoderma harzianum, Trichoderma viride | Phomopsis vexans | Brinjal | 38–49 | Lab | [59] |
| Trichoderma harzianum T22 | Alternaria alternata | Sunflower | 75 | Lab | [60] |
| T. atrobrunneum | Armillaria mellea | Strawberry | 91 | Field | [61] |
| Trichoderma harzianum | Fusarium moniliforme | Maize | 73.33 | Lab | [62] |
| T. viride, Gliocladium virens | Rhizoctonia solani | Rice | 67.94% and 68.62% respectively | Lab | [63] |
Table 3.
Reports on gene identification in Trichoderma species.
| S.No. | Identified Genes | Trichoderma Species | Function | References |
|---|---|---|---|---|
| 1 | Tvsp1 | T. virens | This gene protects cotton seedlings against Rhizoctonia solani. | [64,65] |
| 2 | Tag3 | T. asperellum | Responsible for glucanase production for cell wall degradation. | [66] |
| 3 | TgaA and TgaB | T. virens | Bicontrol efficacy for management of R. solani and Sclerotium rolfsii. | [67] |
| 4 | ThPG1 | T. harzianum | This gene is required for beneficial interaction between Trichoderma harzianum and the host. | [68] |
| 5 | ThPRT2 | T. harzianum | Mycoparasitism activity for Botrytis cinerea. | [69] |
| 6 | tri5 | T. brevicompactum IBT40841 | Production of antifungal activity and trichodermin against fungus causing infection in the human body (Candida spp. and Aspergillus fumigatus). | [70] |
| 7 | erg1 | T. harzianum CECT 2413 | Ergosterol biosynthetic pathway (EBP). | [71] |
| 8 | TvGST | T. virens | This gene provides enhanced tolerance against cadmium stress. | [72] |
| 9 | TrCCD1 | T. reesei | This gene facilitates pigment production and hyphal growth. | [73] |
| 10 | egl1 | T. longibrachiatum | Exhibits antagonistic activity against Pythium ultimum. | [74] |
| 11 | qid74 | T. harzianum | Plant biofertilization and root architecture. | [75] |
| 12 | Taabc2 | T. atroviride | ATP binding cassette transporter plays a major role in cell membranes. | [76] |
| 13 | tac1 | T. virens IMI 304061 | This gene shows mycoparasitism against S. rolfsii and R. solani. | [77] |
| 14 | TrCCD1 | T. reesei | This gene promotes condia formation and elongation of fungal hyphae. | [73] |
| 15 | gluc78 w | T. atroviride | Cell wall disintegartion of Pythium and Rhizoctonia spp. | [78] |
| 16 | SL41 | T. harzinum | Showed mycoparasitic action. | [79] |
| 17 | Taabc2 | T. atroviride | This gene is important for biological control of necrotrophs (B. cinerea and R. solani). | [76] |
| 18 | Monooxygenase | T. hamatum | This gene shows antagonistic activity against Sclerotinia sclerotiourum and S. cepivorum. | [80] |
| 19 | Xl1 | Trichoderma strain Y | This gene is helpful in hemicellulose breakdown. | [81] |
| 20 | eg1, β-1,4 glucanase | T longibrachiatum | This gene shows antagonistic activity against Pythium ultimum in cucumber. | [82] |
| 21 | pacC | T. virens | This gene acts as myctorphy. | [83] |
| 22 | tvhydii1 | T. reesei | Important in mycoparasitism and plant-fungus interaction. | [84] |
| 23 | hmgR | T. koningii | These genes inhibit the pathogen Rhizoctonia solani. | [85] |
| 24 | Tasx1 | T. asperellum | This gene plays a key role in morphological development, mycoparasitism, and antibiosis. | [86] |
| 25 | gpr1 | T. atroviride | This gene is required for the stability of cell walls and hyphal growth. | [87] |
| 26 | TvCyt2 | T. virens | Trichoderma–Arabidopsis interaction | [88] |
| 27 | gluc31 | T. harzianum | Mycoparasitism ability and influence cell wall organization. | [89] |
| 28 | ipa-1 | T. virens | Antibiosis of R. solani. | [90] |
| 29 | TasXyn24.2, TasXyn29.4 | T. asperellum | Induced resistance and promoted growth in seedlings. | [91] |
| 30 | agl1 | T. atroviride | Biological control of plant pathogen. | [92] |
3. Biocontrol Properties of Trichoderma against Phytopathogens
Trichoderma spp. have the ability to produce metabolites, modulate the plant defense responses, and act as a hyperparasite [93,94]. Moreover, Trichoderma strains are effective BCAs because of their high reproductive potential, resilience to harsh environments, efficiency in utilizing nutrients, ability to alter the rhizosphere, aggressiveness against phytopathogenic fungi, and effectiveness in fostering plant growth and defense mechanisms. Due to these characteristics, Trichoderma may be found in all habitats and at high population densities [95]. Trichoderma BCAs can even have a beneficial impact on plants by promoting biofertilization, which increases plant development and enhances plant defense systems [96]. Trichoderma uses indirect and direct methods to control plant pathogens. The power of these mechanisms in the biocontrol method relies upon the type of Trichoderma strain, the antagonized pathogen, including its host, and the ecological situation [96]. The direct mechanism includes mycoparasitism and coiling, whereas the indirect mechanism includes challenges for nutrients and space, systemic acquired resistance, and antibiosis. Among them, mycoparasitism, competition, and antibiosis play a major role in Trichoderma-mediated biological control.
3.1. Mycoparasitism
Parasitism is one of the important mechanisms of fungal antagonist, where one fungus parasitizes (mycoparasite) another fungus (host), and this process is known as mycoparasitism. It has been observed that mycoparasitism involves four sequential steps: chemotaxis, recognition, attachment, and wrapping, as well as penetration of the pathogen cell wall and host digestion [97,98,99]. Trichoderma is widely used as a biofungicide against phytopathogens such as B. cinerea as well as the soil-borne pathogens Rhizoctonia, Sclerotinia, Pythium, and Fusarium spp. [14,100].
Trichoderma species employ several mechanisms to antagonize and mycoparasitize other pathogenic fungi, which includes competing for nutrients [101], releasing antibiotic metabolites, and activities like encircling the host and enlargement of the appressorium-like structure [51,102,103]. The degradation of host tissues containing pathogenic organisms occurs due to the enzymatic breakdown of cell walls facilitated by hydrolytic enzymes (such as chitinase, β-1,3-glucanase, and cellulase) that are synthesized by Trichoderma spp. [104]. In T. atroviride, the nag1 gene coding for N-acetylglucosaminidase has a significant effect on chitinase induction, followed by biocontrol [105]. In the parasite interaction between Trichoderma and R. solani, host-released dispersal factors are responsible for inducing ech42 (encoding endochitinase 42) gene transcription prior to physical touch [30,106]. Lectins present in the host cell wall cause the parasites to cover the hyphae of the host after direct contact [27,100,106].
3.2. Antibiosis
Antibiosis is a phenomenon where one organism is prevented/inhibited by another microorganism through secondary metabolites (SMs). Trichoderma spp. synthesizes SMs (pyrone, heterocyclic compounds, terpenoids, polyketides, etc.) [107] and also produces specific low molecular weight compounds/antibiotics for combatting plant pathogens [108]. Antibiosis has been reported to occur during contact among pathogen, plant, and Trichoderma spp., which triggers Trichoderma to produce antibiotics and SMs to reduce the growth of phytopathogens. More than 180 secondary metabolites have been extracted from Trichoderma spp., showing different classes of chemical compounds [109,110]. On the basis of their biosynthetic origin, these compounds can be classified as peptaiboles, polyketides, and terpenes [111]. Many species of Trichoderma genus are known to synthesize peptidols that are non-proteinogenic amino acids (α-aminoisobutyric acid, a polypeptide antibiotic with a 500 to 2200 Da molecular weight). These compounds are acetylated at the N-terminus and contain an aminoalcohol at the C-terminus [112].
Different types of Trichoderma produce different antibiotics; for example, T. viride produces mucortoxins A and B, mucorin, trichophyton, and mucorin. Similarly, mucorin A and B were isolated from T. mucorin. T. harzianum produces tricholongins BI and BII, while longibrachins and trichokonins were extracted from T. koningii. Atroviridines A-C and neoatroviridines A-D have been obtained from T. atroviride culture. In addition, other antibiotics and fungicidal compounds have been isolated from T. harzianum, T. koningii, T. aureoviride, T. virens, T. hamatum, and T. lignorum [110].
Growth of soil pathogens such as Phytophthora solani, P. middletonii, P. cinnamomi, Bipolaris sorokiniana, and Fusarium oxysporum was adversely affected in the vicinity of Koninginin D [113]. Similarly, viridins obtained from Trichoderma spp. like T. viride, T. koningii, and T. virens inhibit the spore germination of Colletotrichum lini, Botrytis allii, Penicillium expansum, Fusarium caeruleum, Stachybotry satra, and Aspergillus niger [114]. Harzianic acids derived from T. harzianum show antimicrobial activity against Sclerotinia sclerotiorum, R. solani, and Pythium irregulare [115]. It has been found that Trichoderma spp. And Gliocladium suppressed the growth of various soil-borne plant pathogens (Fusarium spp., Macrophomina, Sclerotium rolfsii, and Sclerotinia spp.) [116,117]. Silva et al. (1998) studied the antibiosis mechanism of Trichoderma against Colletotrichum spp. [118].
3.3. Competition
Limitation and competition for nutrient sources can lead to the natural control of fungal pathogens [119]. Trichoderma is a cosmopolitan fungus and is found in all kinds of soils because of its outstanding competitive potential. It can fight with phytopathogens for nutritive sources, such as C, N, and Fe, and also acts as a biological antagonist towards soil-borne pathogens. It is also an aggressive competitor that grows rapidly and quickly colonizes its substrate and controls slow-growing pathogens [120]. Trichoderma is more competitive with other microorganisms due to certain abilities, such as a higher growth rate and enhanced aptitude to mobilize and utilize nutrients from soil/substrate [121,122]. Thus, competition for macro and micronutrients plays a key role in the interaction between Trichoderma-plant pathogen [123] because Trichoderma species compete with bacteria in the rhizosphere of crops for nutrients and sites of infection [124]. Compared to other rhizosphere bacteria, Trichoderma shows a better ability to produce and take up nourishment from soil; therefore, the management of certain disease-causing entities, such as Botrytis cinerea, using Trichoderma through food competition is possible [125]. It is observed that there are four major features of any organism that contribute to its saprophytic ability and inoculum potential: (i) fast germination of fungal propagule, rapid hyphal growth towards nutrients, (ii) production of suitable enzymes for carbon constituents of the host plant, (iii) secretion of growth inhibitor compounds (fungistatic and bacteriostatic), and (iv) tolerance to fungistatic substances produced by competing microorganisms. Antagonistic fungi can compete with the pathogens for food and space by colonizing the normal environment, i.e., plant tissue, rhizosphere, or phyllosphere [126]. It depends on the colonization level of the host plant and acclimatization to the environmental situations in which they are living [127]. In order to successfully compete with other fungal phytopathogens for food and space, Trichoderma should exhibit efficient strategies for colonization of the plant and should be plentiful in an area where competition with other microorganisms occurs [126].
Trichoderma spp. produces iron chelating agents and siderophores, which make iron unobtainable for rhizospheric bacteria, which eventually leads to the extinction of the disease. Thus, Trichoderma acts as a competitor that helps control plant diseases [128]. Apart from this, due to its ability to colonize the rhizosphere and outcompete for nutrients, T. harzianum (T35 Strain) reduces the availability of nutrients and the amount of rhizospheric space available for the fungal wilt agent of watermelon (Fusarium oxysproum f.sp. meloni) to colonize [129]. Srinivasan et al. (1995) demonstrated the importance of competition between siderophore-producing Trichoderma strains and wood decay Basidiomycetes fungi [130].
Mokhtar et al. 2013 studied the interaction between T. harzianum and a few fungal species, such as Alternaria alternata, Fusarium acuminatum, and A. infectoria. The results revealed that lack of nutrients caused death of the pathogenic fungi [131]. It has also been found that Trichoderma can compete with plant pathogens, including Colletotrichum sp., Botrytis sp., and Phytophthora sp., for complex and simple substrates of carbon [132]. In order to successfully compete with other fungal phytopathogens for food and space, Trichoderma should exhibit efficient strategies for colonization of the plant and should be plentiful in an area where competition with other microorganisms occurs [126].
Trichoderma colonization of roots commonly improves nutrient absorption and utilization, crop yield, tolerance to abiotic stressors, and root growth and development [133]. Trichoderma hamatum or Trichoderma koningii can boost crop production up to 300% after addition in the field. In greenhouse experiments, a substantial increase in yield was reported after treating the seedlings with Trichoderma spores [95]. The ability of Trichoderma BCAs to produce metabolites that either prevent spore germination (fungistatic), kill cells (antibiosis), or alter the rhizosphere, for example, making the soil acidic, leads to biocontrol that is unsuitable for pathogen proliferation [96]. Trichoderma strains quickly proliferate when introduced to the soil because they are inherently resistant to a wide range of hazardous substances, including insecticides, fungicides, and herbicides like DDT [95].
3.4. Production of Antibiotics and Other Antifungal Compounds
It has been shown that the Trichoderma species produce a large number of secondary metabolites, about 370 of which are members of several chemical compound classes with potent antagonistic activities [126,134]. Peptaibols and polyketides are the most significant non-volatile and volatile organic compounds (VOCs) produced by the majority of Trichoderma strains [2]. The volatile antibiotic 6-phenyl-pyrone (6PAP), responsible for the distinctive coconut scent and the biological control of F. oxysporum, is produced by the T. viride, T. harzianum, and T. koningii species [135]. In addition, T. harzianum also produces harzianic acid, a tetramic acid that has strong antifungal action as well as the capacity to stimulate plant development and function as a chelator [136].
3.5. Induced Systemic Resistance
Trichoderma can trigger a host plant’s defensive mechanism while preventing harmful pathogens from proliferating and growing, and it can also encourage crops to build self-defense mechanisms to gain local or systemic disease resistance [137]. Two methods are used to achieve Trichoderma-induced plant disease resistance: first, control the elicitors or elicitors that trigger the plant disease resistance response; and second, release oligosaccharides from the cell-wall-degrading enzymes produced by Trichoderma to cause plant resistance [138]. Saravanakumar et al. (2016) found that Trichoderma coated corn seeds dramatically increased the peroxidase (POD) and phenylalanine ammonia lyase (PAL) activity, and the plants were resistant to Curvularia leaf spot of corn [139].
4. Trichoderma Selective Medium
Isolation and quantitative estimation of Trichoderma from the soil is tedious due to fast growth and similar morphology of other existing fungi (competitors) on a routine growth medium. It is isolated from rhizospheric soils using the serial dilution technique. Different types of low-cost and easily procurable culture media have been tested for growth and sporulation in Trichoderma, such as carrot agar (CA), oatmeal agar (OMA), potato-dextrose agar (PDA), Czapek’s Dox medium (CDA), Rose Bengal chloramphenicol agar (RBCA), etc. However, the Trichoderma selective medium (TSM) with low glucose levels is preferred for isolation and quantification of Trichodema because it favors its growth while inhibiting the growth of other soil fungi such as Mucor and Rhizopus spp. [140]. The components of TSM for 1 L media (pH 4) are 0.2 g of magnesium sulfate (MgSO4∙7H2O), 0.9 g of dipotassium hydrogen phosphate K2HPO4, 0.15 g of potassium chloride (KCl), 1.0 g of ammonium nitrate (NH4NO3), 3.0 g of glucose, 0.15 g of Rose Bengal agar (RBA), 20 g of agar-agar, and 0.25 g of chloramphenicol.
5. Methods for Isolation of Trichoderma
There are various techniques for the extraction of Trichoderma spp. However, serial dilution of samples is one of the most reported methods. This method is very simple, economical, and suitable for dealing with various samples. In this method, the soil sample is collected from the rhizosphere zone of the plants, air-dried, and ground into fine powder. Soil suspension is prepared by mixing ten grams of a dry powdered soil sample in 90 mL of sterilized water, followed by shaking it for at least 5–10 min on an orbital shaker to make a homogenous solution. Thereafter, dilutions of samples are prepared from 10−1 to 10−6, and a 100 µL solution is pipetted from the third or fourth diluted test tube and spread evenly on Trichoderma selective medium (TSM) using a sterilized ‘L-shaped’ glass spreader under a laminar air-flow chamber. The inoculated Petri plate is kept in a BOD incubator at 28 ± 1 °C for 4–5 days. After 5 days, each colony that is visible on the plate is considered a single colony-forming unit (cfu). In a report by Gil et al. (2009), rhizosphere soil samples were taken to monitor Trichoderma and Glueomyces populations with different crops and tillage lines. The best medium for isolating Trichoderma and Gliocladium spp. was potato dextrose agar + streptomycin + chloramphenicol and rose bengal, and for Actinomyces, Küster medium + cycloheximide and sodium propionate [141].
In the study conducted by Liu and colleagues in 2020, Trichoderma was isolated from the rhizosphere soil using a serial dilution technique [142]. Subsequently, these isolates were cultured on a Rose Bengal medium, as described by Zhou et al. (2020) [143]. Following incubation for 48 h, Trichoderma colonies were carefully selected and transferred onto a potato dextrose agar (PDA) medium. To ensure the purity of these putative Trichoderma colonies, a single spore isolation technique, as outlined by Dou et al. (2019) [144], was employed. This involved transferring a single spore onto a PDA medium, ultimately resulting in the acquisition of a pure Trichoderma isolate. In a similar report involving the isolation of Trichoderma from rhizosphere soil using a serial dilution technique, the cultures were maintained on PDA at a temperature of 28 °C for 2 to 7 days. Following incubation, specific colonies of Trichoderma were selected and transferred onto fresh PDA medium. Further incubation of these selected colonies was performed at 28 °C in a dark environment [145].
6. Identification
Fungi are morphologically, metabolically, ecologically, and phylogenetically diverse [146]. Hence, exact fungal identification is necessary for all fungal investigations and applications, and it enables us to anticipate the beneficial or harmful properties of fungal spp. and their distribution and establish safety measures. Despite their significance in biological control, identifying fungi is still a difficult task, especially for chemists who are not associated with mycologists [147]. There are various challenges in the identification of Trichoderma species: (i) problems with fungal species identification using morphology criteria; (ii) potential advantages and limitations of ribosomal genes commonly used for identification of fungal pathogens and their ITS region, the official DNA barcode marker for fungal-level identification; (iii) limited knowledge on the use of NCBI-BLAST to search for DNA barcodes; (iv) extensive curated molecular database of fungal sequences used to increase or replace ITS for species-level identification of certain fungal taxa; and (v) method of constructing phylogenetic trees from DNA sequences to facilitate the identification of fungal species [148].
There are some reports of species misidentification using external features; for example, the name T. harzianum was used for a few species [149]. Identification can be obtained using a similarity search tool (TrichoBLAST) and oligonucleotide barcoding (TrichOKEY), both available online at http://www.isth.info/ (accessed on 12 October 2023) [9,150,151]. In addition, phenotypic microarrays can be used to classify new species and allow the analysis of carbon utilization patterns from 96 carbon sources [152]. Chaverri and Samuels (2013) identified endophytic organisms based on habitat preferences and trophic models to understand different groups of radiation types and their potential to be used to develop new biological control strategies [153]. Cellulose acetate electrophoresis can be used to characterize several species with defined isoenzyme structures, indicating that this method can be useful for identifying biochemical differences in some Trichoderma species [154]. Ongoing efforts to characterize the diversity and geographic distribution of T. hypocrea have resulted in extensive documentation of this genus [153,155]. Currently, the International Subcommittee on T. hypocrea provides 104 species properties at the molecular level (http://www.isth.info/biodiversity/index.php, accessed on 12 October 2023).
7. Preservation of Trichoderma Cultures
Different isolation and preservation methods have been tried for the T. viride fungus. The crosslinking isolation method using 0.5% Tween 20 proved to be the best for isolation and statistics [156]. Various methods have been tried to study the long-term storage and recovery of T. viride isolates in the stored conditions. T. viride can live for more than 2 years in dry form/weak form, but drying is a crucial step because if it dries too quickly, the fungal spores lose their vigor/pathogenicity, and it will cause the death of the fungal spores, and becomes infected with other bacteria or viruses. In the case of the H2O preservation method, the culture can be recovered even after 12 months without losing its potency [156,157].
Nowadays, various methods have been optimized to minimize the problems faced by the serial transfer of fungal culture and to extend its life span [158]. Generally, most preservation methods slow down the metabolism of microorganisms by storage at 4 °C and/or mineral oil coating, which restricts the cell’s ability to take up oxygen, slows down the metabolic activity, and lengthens the storage time [159]. There are various methods used for the preservation of Trichoderma spp., such as oil drop cover, direct freezing of culture, water preservation, and spore suspensions, which are easy and lucrative methods [158]. The main objective of preserving Trichoderma culture for the long term is to ensure the viability, physiological, morphological, and genetic integrity (Table 3) of the cultures over time, and safe storage is further required to validate the reliability of strains. The American Type Culture Collection uses a variety of preservation techniques, including soil or oil preservation [160,161], water agar slants, cryopreservation or low temperatures (−20 and −70 °C) [162,163], etc. The cellulose filter paper was used for the preservation of T. harzinum and was found to be simple, less costly, less time-consuming, and requires less refrigeration for the storage of the Trichoderma isolates. Stocco et al. (2010) and Tucci et al. (2011) tested different methods of preservation, including distilled water, mineral oil, and silica gel for long-term storage of Trichoderma [164,165].
8. Estimation of Colony Forming Units (cfu)
Serial dilution, pour plate, and spread plate are standard techniques for CFU estimation of microbes (fungi, bacteria, etc.) [125]. The spread-plate method offers several benefits over the pour-plate method and includes increased flexibility in handling, reducing detrimental effects on temperature-sensitive microbes, preventing aerobic organisms from being trapped in the agar medium, and counting of colony forming units can be performed easily on the surface of the plate [166,167]. Microbial culture is spread on an agar-gelled medium through an L-shaped glass, steel, or plastic spreader. However, the use of a spreader during the standard spread plating method might cause substantial harm to microbial inoculum, which can have an impact on colony-forming units on the surface of the medium [168]. There are alternative approaches that do not involve the use of an inoculum-spreader, known as spotting and tilt spreading (SATS) [168].
9. How to Find a New Bio Control Agent?
Two main techniques are frequently used to find new biocontrol agents. The first strategy entails plant-based or in silico indirect screening of microbial libraries for biocontrol abilities. According to the second method [169,170], organisms are isolated from the environment where the product application is anticipated to take place and then immediately tested for activity. Additionally, the development of NGS (next-generation sequencing) techniques has facilitated a quick shift to a direct functional approach in studies on biological pesticides. Utilizing a variety of bacterial and fungal biofertilizer strains in conjunction with each other can improve plant growth promotion [171,172]. Similar to this, microbes with bio-control abilities can work synergistically with other bio-control agents to exacerbate their resistance to plant diseases [173].
10. Use of Carriers
There are ample reports on the manufacture of Trichoderma products for marketing using common synthetic media such as cellulose, glucose, molasses, and soluble starch [174]. However, one of the primary causes of restricted utilization is the high price of these raw materials for the marketable manufacturing of biocontrol agents. Many studies have deployed inexpensive substrates like composted pith of coir and coffee waste [175,176,177], poultry manure and coffee waste [178], neem cake, degraded coffee pulp, cow dung, and coir pith [179], sorghum grain floor, sawdust, wheat bran, groundnut shell, molasses, farm produced waste, extract of biogas plant, dried cow dung, neem cake, talc, mushroom compost, fly ash, peat soil [180], fruit, vegetable, and crop wastes, poultry manure and FYM [181], rotten wheat grains, sugarcane baggase, fruit juice waste, and vegetable and fruit waste, for mass production of Trichoderma species [182]. Various inexpensive and readily available local organic media such as neem cake, coconut husk, and decayed pulp of coffee have been recommended for Trichoderma production on a large scale [179]. Mustafa et al. (2009) identified organic fertilizers such as neem cake, vermiculite, and mushroom composts as excellent carriers for Trichoderma mass multiplication [183]. Usage of neem cake enriched with a selected isolate of Trichoderma talcum powder is suggested for the management of coconut basal stem rot and areca nut and coconut stem bleeding. Over the last few years, due to the lack of high-quality neem cakes in the market and very expensive commercial formulations, farmers need potential biological control agents with pocket-friendly formulations for effective disease control. The production of acceptable, easy-to-prepare, and lucrative formulations has a key role in the accomplishment of biocontrol programs and agricultural sustainability.
11. Pilot Experiments
The formulation consists of an active ingredient in the form of a microbe or microbe-derived products in an appropriate carrier material (sterile or non-sterile) with additives that aid in the stability and protection of the microbial cells during storage, transit, and the target location. Choosing the right formulation is extremely important because it has the potential to make microorganisms successful or fail [184].
Formulation of the bioagents has a significant function in determining the role of application with high performance under field conditions. The majority of the formulation procedure involves combining bioagents with an appropriate carrier and/or additives that ensure ease of distribution, prolonged storage, and low cost. Trichoderma is frequently produced on a large scale using solid fermentation. A variety of low-cost cereal grains, including sorghum, millets, and ragi, can be used [185]. The grains are sterilized, hydrated, and inoculated with Trichoderma for 10 to 15 days. Trichoderma covers the grains with dark green spores, and these grains can be ground into a fine powder and applied to the seed surface, or they can be applied directly to the soil/ enriched FYM before the field application. This method is appropriate for small-scale production in cottage industries or at the level of the individual farmer. However, the drawbacks include lengthy production time and contaminant-prone products.
As aforementioned, formulations are of many types; they include dry products (such as granules, dust, and wettable powders), liquid products (such as emulsions, oil, and water; usually contain one but sometimes two strains of active ingredient), and microencapsulation. The efficacy of microbial inoculants largely depends on the type of formulation and the delivery technology that extends the shelf life for at least a few months, and in all cases, the PGP/antagonistic activity is retained [186]. The production cost also has to be considered and kept to a minimum while developing a microbial formulation. A good formulation should be easy to handle and apply so that it is delivered at the target site, protects the PGP microbes, and enhances its activity from harmful environmental factors under field conditions. A detailed review of different types of formulations, additives used, and PGP/antagonistic microbes used on various crops was reported by Nakkeeran et al. (2005) [187] and Bashan et al. (2014) [184]. It is understood that the major role of a formulation is to provide a more suitable micro-environment that prevents the rapid decline of an introduced PGP microbe in the soil. BCAs can be formulated in many forms, although commercial biocontrol formulations conventionally come only in very few variations: powder, granules, or liquid forms [188].
The formulation of BCAs is a multistep process that begins immediately after the selection of the most promising BCA for the intended effect on a target pathos stem, and it consists of combining viable microorganisms with carriers and adjuvants to produce high-quality preparations of stable shelf-life and proven efficacy. From a technical point of view, this requires in-depth knowledge of the ecology and biology of the BCA, the pathosystem, the environment, and the application niche, along with considerations of the plant growth stage, inoculation techniques, and types of irrigation systems involved in the agri-food system [189].
12. Types of Formulations
As mentioned, several formulations (Figure 1) have been tested for pilot production of Trichoderma (Table 4). These can be differentiated based on their efficacy in enhancing the stability of Trichoderma cultures.
12.1. Liquid Formulations
Liquid formulations for biological control agents (BCAs) can vary in composition, but they typically involve microbial cultures or suspensions mixed with water, mineral or organic oils, polymers, or a combination of these components. Suspension concentrates, in the context of microbial inoculants, are typically made by introducing solid active ingredients (which can be either free or immobilized microbial cells) into water or an aqueous solution using traditional methods [186]. Liquid formulations for biopesticides are quite common. For instance, P. fluorescens was formulated in coconut water supplemented with glycerol or polyvinylpyrrolidone (PVP) as a suspension concentrate [190]. In a study conducted by Mbarga and colleagues in 2014, they formulated an oil dispersion that included soybean oil, an emulsifying-dispersing agent, a structural agent, and glucose, and this formulation contained conidia of T. asperellum [191]. In another research reported by Peeran et al. in 2014, water-in-oil emulsions of Ps. fluorescens were created using a mixture of water, glycerin, polyethylene glycol, Tween 20, and either coconut, rice bran, or castor oil [192]. Herrera et al. (2020) developed liquid formulations of T. asperellum TV190 using emulsified mineral or vegetable oils; this formulation showed greater viability of 37–43% (mineral) and 56–63% (vegetable) compared to the control (8–12%) [193]. A similar investigation was conducted by Patil et al. 2021 to check the efficacy of the liquid formulation of T. asperellum against the Fusarium wilt of chickpea [194]. The oil-based formulation was chosen because it maintains the organism in a physiologically dormant state. Three different oils, paraffin oil, soybean oil, and groundnut oil, were used as carrier material for the oil-based liquid formulation of T. asperellum. The population density of T. asperellum found in paraffin oil was 28.67 × 108 CFU/mL at 30 days, and it decreased to 18.00 × 108 CFU/mL after 180 days.
For the production of liquid formulations, low-cost growth media like sugarcane molasses and brewer’s yeast have been used [15]. The benefit of this media is that it increases the production and quality of the biomass by permitting regulation of pH, temperature, and nutrients (among other biological factors), hence reducing contamination [195].
12.2. Solid Formulations
Another method to produce Trichoderma inoculum is the solid formulation or fermentation. Agricultural wastes, for example, wheat straw, sugarcane bagasse, sawdust, corn cob meal, and rice bran, are used alone or as a combined substrate or food base for the multiplication of Trichoderma. To provide a stable product with an extended shelf life, both liquid and solid formulations must be dried [196]. Such formulation is cost-effective for small-scale production, but it is immense since it requires a large area for preparation, inoculation, storage, drying, and milling. However, spray drying (cost-effective) is the technique for the large-scale production of microorganisms, including dried particles [197].
Solid fermentation is often used in the research laboratory in the mass production of Trichoderma spp. The fungus is grown in Petri plates, after which spores and other propagules are harvested and processed. The bulk production of biocontrol fungus employing a variety of low-cost agricultural wastes and bioproducts is another way of solid fermentation. Several low-cost cereal grains, including wheat, bajra, and sorghum, are used as a substrate. The grains are soaked in freshwater for 12 h, and extra water is removed the next day. The soaked grains are placed in heat-resistant polypropylene bags and are sterilized in an autoclave. The grains are then inoculated with Trichoderma culture and incubated for 10 to 15 days. Trichoderma covers the grains with a layer of dark green spores. These grains can either be utilized whole for supplementing FYM for soil treatments or finely ground to treat seeds [198].
12.3. Talc Based Formulation
Trichoderma viride formulations using talc as a base material were developed in India at the Agricultural University of Tamil Nadu for application on seeds and soil and seedlings [199]. The process involves culturing Trichoderma in a liquid medium, blending it with talcum powder in a 1:2 ratio, followed by drying it under shade until 8% moisture content. These talc-based formulations of Trichoderma can be stored for a period of 3 to 4 months. In India, it has gained popularity for its effectiveness in controlling various soil-borne diseases by treatment of seeds of different crops at a dosing range of 4 to 5 g/kg seed. Numerous non-government companies in India produce huge volumes of talc-based formulations for farmers. The annual requirement of Trichoderma in India is nearly 5000 metric tonnes, and this quantity would be sufficient to cover approximately 50% of the country’s agricultural land [185].
Pradhan et al. (2022) prepared a talc-based formulation of T. viride and tested it against Fusarium wilt disease in chickpeas. The biocontrol potential of talc-based formulation was found effective in terms of seed and soil application and for reducing the incidence of wilt in chickpeas [200]. Sundaramoorthy and Balabaskar (2013) also developed the talc-based formulation of T. harzinaum to evaluate their biocontrol efficacy against wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici under greenhouse conditions [201]. The application of Trichoderma showed a minimum incidence (15.33%) of wilt disease.
12.4. Vermiculite–Wheat Bran-Based Formulation
For the preparation of Trichoderma vermiculite–wheat bran-based formulation, the fungus was cultivated in a molasses–yeast medium for 240 h. Wheat bran (3.3 g) and vermiculite (100 g) were sterilized for three days at 70 °C in an oven. This substrate was mixed with 14 mL of liquid culture and 17.5 mL of 0.05N HCl and allowed to dry in the shade before packaging [202].
12.5. Oil-Based Formulations
It is a type of liquid formulation where Trichoderma is mixed in an oil carrier. The process involves growing Trichoderma in a broth medium, followed by mixing it with an appropriate oil carrier. The oil carrier can be mineral oil, vegetable oil, coconut oil, or any other oil that is compatible with Trichoderma and provides stability to the formulations. Moreover, it offers several advantages: oil carrier acts as a protective barrier against unfavorable conditions such as UV radiation and high temperature, which can reduce the efficacy and viability of Trichoderma formulations. Additionally, oil-based formulations can enhance the adherence capacity on the surface of seeds, plant leaves, etc. They also facilitate the slow release of Trichoderma in the applied area and increase the activity against the plant pathogens. Bacteria are removed with the help of surfactants in non-aqueous solvents such as gasoline distillates (diesel oil and mineral oil) and vegetable oil (peanut, etc.). This can create a stable emulsion when combined with water. When diluted in water, emulsifiable concentrates rapidly create a homogenous emulsion because they need a high concentration of oil-soluble emulsifying component. Trichoderma is presently being utilized as an antibiotic. The oil-based mixture should be accepted for foliar application in dry weather and must have a long storage life because the propagules covered with oil can remain alive on the crop plants, even in adverse conditions. Batta (2007) prepared an emulsion formulation of T. harzianum, which considerably reduced the postharvest fruit rot caused by Botrytis cinerea. The Project Directorate of Biological Control (PDBC) located in India has successfully developed a modified emulsion formulation of T. harzianum from local sources, extending its shelf life to 8 months. This innovative formulation has proven to be highly effective in protecting peanuts from soil-borne diseases [203].
12.6. Sodium Alginate Encapsulation of Trichoderma
An alginate-based formulation of Trichoderma refers to Trichoderma incorporated into an alginate matrix, which is a natural polymer derived from seaweed. Trichoderma spores are dissolved in a 1:4 ratio with a solution of 0.6 percent sodium alginate, which is then allowed to drip into a beaker containing 1.5 percent CaCl2 for 30 min. The uniformly sized, 3 mm CaCl2 beads are squeezed through a sterile muslin cloth and allowed to aseptically air dry. The Trichoderma beads with sodium alginate coating are placed in plastic bottles with sterile distilled water and maintained at room temperature [204].
12.7. Press Mud-Based Formulation
Press mud is an organic by-product generated from the sugar industry, which can serve as a potential substrate for the cultivation of Trichoderma [205]. To carry out the process, a 9-day-old culture of Trichoderma cultivated in potato broth medium is blended with 120 kg of mud and kept moist. The mixture is covered with burlap to allow airflow and trap moisture in the shade. The seed culture for the next growth gets ready in 25 days. Before the field application, it is mixed with 8 tons of mud and kept at a low temperature for 8 days.
12.8. Coffee Husk-Based Formulation
Sawant and Sawant (1996) used coffee husk as a substrate for the mass production of Trichoderma, which is a by-product of the coffee industry in Karnataka [178]. This formulation has been widely utilized in Karnataka and Kerala in treating Phytophthora foot rot of black pepper. Rosane et al. (2008) used several cost-effective solid substrates (rice, corn, and wheat bran) for the cultivation of Trichoderma on a large scale and found that wheat bran is one of the best substrates for the successful production of Trichoderma spores [206].
12.9. Banana Waste-Based Formulations
Banana waste is also a unique material for the preparation of Trichoderma-based solid formulations. Balasubramanian et al. (2008) developed a method for the cultivation of Trichoderma using banana waste as a substrate. This method involved urea rock phosphate, banana waste, and a consortium of fungal and bacterial biocontrol agents (Bacillus polymixa, P. sajorcaju, and T. viride) for the preparation of Trichoderma formulation [207]. A well is prepared from banana skins, pseudostems, and cores that are cut from 5 to 8 cm long. Five layers of various components are arranged in a prepared pit. One ton of banana waste, 5 kg of urea, 125 kg of rock phosphate, and 1 L of B. polymixa, P. sajorcaju, and T. viride are all present in each layer. These wastes decompose for 45 days and are cultured to be used later in the field.
12.10. Dry Flowable Formulation
Dry flowable/water-dispersible granule formulations are safe for the environment and farmers [208]. Due to their granular structure, such preparations do not easily absorb moisture from the air. Encapsulation is an initial stage in the production of dry flowable formulations, as it is vital for enhancing the survival and efficacy of the formulation type.
As mentioned, encapsulation techniques have been used to establish encapsulated trichoderma. This can be achieved using a dry method [209] or modified alginate microbeads [210]. Another approach involves the development of a spray-dried flowable system based on polyacrylic acid, citric acid, and sodium bicarbonate, with polyvinyl alcohol as a binder and lecithin as a wetting agent [211].
12.11. Sawdust-Based Formulation
The application of sawdust as a carrier material is widely suggested as it is easily procurable and possesses a natural ability to retain a high level of organic matter and water-holding capacity compared to other carrier materials [212,213,214]. Arora et al. (2008) tested several substrates, and sawdust was found to be the most effective carrier for microbial growth [213].
A one-kilogram mixture consisting of sawdust, soil, and molasses is filled in heat-resistant polybags. These polybags are tightly sealed and sterilized at 15 kg/cm2 pressure at 121 °C for 15 min. For T. harzianum, 10 mg chloramphenicol/kg material is added. Subsequently, the bags containing 1 kg autoclaved mixture were inoculated with a homogenized pure liquid culture of the biocontrol agents (5 mL/bag) using a sterilized needle and syringe. A small hole was made in the polybag to inoculate the culture and then resealed with cello tape to avoid contamination. Inoculated bags are placed in a BOD incubator at 28 ± 2 °C for 10–15 days. Throughout the incubation period, the bags are gently shaken to ensure that the biocontrol agent colonizes the material uniformly [215].
Table 4.
Different substrates and their efficacy in Trichoderma biomass production.
| S.No. | Substrate | Composition/Components | Efficacy | References |
|---|---|---|---|---|
| 1 | Cereals (wheat, moong, maize) | Grains (200 g + sugar (1%) + Trichoderma harzianum | 12.96% | [216] |
| 2 | Vermicompost fortified with Trichoderma | Vermicompost + cereals + pulses | Reduction of 10.01%incidence of wilt in chili | [216] |
| 3 | Wheat seeds-based formulation | Grinded grain + sugar solution (1%) + Trichoderma harzianum | 38 × 107 cfu/g | [217] |
| 4 | Cow dung enriched formulation | Decomposed cow dung + Trichoderma formulation | 37.5 × 107 cfu/g | [217] |
| 5 | Talc-based formulation | Talcum powder + CMC + Trichoderma culture | 37 × 107 cfu/g | [217] |
| 6 | Sorghum grain | Partial crushed grain + sugar (1%) solution + distilled water | 6.1 × 104 cfu/g | [218] |
| 7 | Wood pellets-based formulation | Beech, fir, and chestnut + conidial suspensions of T. atroviride + distilled water + soy flour | Growth increase by ten-fold | [219] |
| 8 | Vermiculite–wheat bran-based formulation | Vermiculite (100 g) + wheat bran (33 g) + fermented Trichoderma biomass (20 g) + 0.05N HCl (175 mL) | - | [202] |
| 9 | Vermiculite bentonite-based formulation | Oat (20 g) + bentonite (50 mL) + vermiculite + T. harzianum + water (60 mL) | Maintained cfu after 8 weeks | [220] |
| 10 | Coffee husk-based formulation | Coffee fruit skin decomposed with cow dung + poultry manure + T. harzianum suspension | 9 × 1011 to 3 × 1012 cfu/g substrate | [178] |
| 11 | Oil-based formulation | Glycerol (1%), PVP (1%), Tween 20 (1%) as an emulsifying agent, ZnSO4 (0.5%) to increase the shelf-life, coconut oil, and distilled water | More than 180 days | [221] |
| 12 | Pesta granules-based formulation | Wheat flour (100 g) + fermenter biomass + sterile water (52 mL) | Viable for a long time | [222] |
| 13 | Banana waste-based | Banana waste (chopped 5–6 cm length) + rock phosphate | Six months | [207,223] |
| 14 | Trichoderma sodium alginate encapsulation | Trichoderma suspension + sodium alginate (0.6%) solution + CaCl2 (1.5%) | Viable for more than six years at room temperature | [163] |
| 15 | Wheat flour- kaolin | Wheat flour (80 gm) + kalolin (20 gm) + fermenter biomass (52 mL) | Few months | [224] |
| 16 | T2- liquid formulation (NIPHM medium) | Liquid formulation (NIPHM medium) Trichoderma filtrate (250 mL) + water (750 mL) + glycerol (3%) | - | [225] |
| 17 | Trichoderma-based compost activator | Fifty grams of soil + rice straw (5 g) + Trichoderma biomass (500 mg) | 5.3 × 1010 cfu/g | [226] |
| 18 | Graphite and silica-based formulation | B. subtilis and T. harzianum + 5% graphite 80 mesh + 1% silica NPs | 35–54% efficacy (in vitro) | [227] |
| 19 | Chitosan-PEG based formulation | Chitosan-PEG + T. harzianum spores + glacial acetic acid (0.1%) | More than six months | [228] |
| 20 | Rice powder-based formulation | Sterilized rice powder + dextrose + talc powder + Trichoderma viride | cfu/g 10 × 109 up to six months at room temperature | [229] |
| 21 | Dextrin-based formulation | T. harzianum filtrate (500 g) + Paraffin oil (500 mL) + CMC (0.2%) + chitosan (0.1%) | Efficacy 26.10%; 4.33 × 107 cfu/g) for six months | [230] |
| 22 | Oil-based liquid formulation | T. asperellum + paraffin oil | 28.67 × 108 cfu/mL for 30 days | [231] |
| 23 | Glycerol based formulation | Molasses yeast extract (MYE) medium+ glycerol (3%) (V/V) +T. harzianum + Talc powder | Extended the shelf-life for 7 to 12 months | [232] |
| 24 | Paste formulation of T. harzianum | Starch (10%) + copper sulphate (20 ppm) + T. harzianum | 11.6 × 1010 cfu/g for 120 days | [233] |
13. Exemplary Properties of Trichoderma Formulation
For the development of a successful formulation of Trichoderma, a product should have the following attributes [234]: (i) formulation should have high rhizosphere competence with other existing microorganisms; (ii) formulation should possess highly competitive saprophytic ability; (iii) formulation should have the potential for disease management and also promotes plant growth; (iv) formulation should be easy to multiply as mass scale and safe to humans and the environment; and (v) formulation should be excellent in terms of action, compatible with other biocontrol agents/biofertilizers and tolerant to stresses such as desiccation, heat, and UV rays (Figure 2).
14. Quality Control Parameters
Before the recommendation of the final product (Trichoderma formulation) certain quality parameters are mandatory for successful application. They are: (i) colony forming units (CFUs) of a fresh product should be a minimum of 2 × 106 cfu/g or ml; (ii) shelf life of the formulation should be a minimum of 4–6 months; (iii) the size of talcum powder should be 500 microns, for the development of a formulation; (iv) the final product should be packed in standard white polythene bags; (v) in the final product, moisture should be less than 20%; (vi) additional contamination should not be more than 1 × 104/mL or g; and (vii) in the final product, the presence of other pathogens such as Salmonella, Shigella, and Vibrio should be eliminated.
The quality control parameters of any biopesticide established by the Central Insecticides Board (CIB) have certain lacuna with regard to the assessment of a potential biocontrol agent. It is expected that the Department of Agriculture (government) may appoint a central body for inspection of quality assurance, not only with regard to the viability of the propagules in the formulation but also with regard to their efficacy against plant pathogens. The authorized body must regularly assess the quality of the products in terms of these parameters to ensure that the farmers have access to higher-quality products.
15. Methods of Application of Trichoderma
Success in controlling plant diseases depends on the distribution method and establishment of Trichoderma in the active site. There are many ways to use Trichoderma, including seed biopriming, seedling root soaking, foliar spray, soil furrow application, soil soaking application, foliar application, and cloth wrapping.
15.1. Seed Treatment
Seed treatment with Trichoderma spp. is one of the precise, simple, and profitable approaches to delivering biocontrol agents for controlling soil and seed-borne pathogens [235]. In this method, a small amount of Trichoderma inoculum is applied uniformly on the outer surface of the seed just a few hours before the sowing. This method has been considered promising for effective inoculation of biocontrol agents on crop seeds of different sizes [236,237,238,239]. After coating the seed with the inoculum of the antagonist, spores germinate on the surface of the seed and inhabit the roots and rhizosphere of germinated plantlets [240,241]. However, the effectiveness of Trichoderma seed treatment relies on its biocontrol properties and the capacity to multiply quickly in the rhizospheric zone of the seedling. Papavizas (1985) suggested that if the biocontrol inoculum multiplies around the site of application, it probably suppresses the pathogens, causing seed-rot and seedling diseases [15]. Lamichhane et al. (2022) revealed that seed treatment with biological products drastically enhanced the percentage of seedling emergence (91 ± 5%), seed germination (7 ± 6%), disease control (55 ± 1%), plant biomass (53 ± 5%), and crop harvest (21 ± 2%), in comparison to non-treated seeds [242]. The biological potency of T. viride talc-based formulation (1% w/w) was found effective against FOC (Fusarium oxysproum f.sp ciceri), which caused wilt of chickpea and also improved the physiological parameters of the crop [200]. In a study by Couto et al. (2021), wheat seeds were primed with two distinct strains of Trichoderma, T. harzianum, and T. asperellum (2 × 1012 CFU 100 kg−1 seed). Couto et al. (2021) revealed that antagonist boosts the percentage of seedling emergence, root and shoot lengths, and shoot dry matter under greenhouse conditions [243]. In another study, three antagonists, namely, T. koningii, T. viride, T. asperelum, and T. reseii, were used as bio slurry for seed treatment of peanut crops. All these three antagonists were found to be effective in various physiological parameters of plants, such as vegetative growth, percentage germination, production, and shoot and root mass of the crop [244]. Thus, multiple studies have demonstrated that Trichoderma seed treatment can improve seedling growth and yield in various cereals and vegetables, such as tomato, cucumber, and maize [245,246].
15.2. Seed Biopriming
Biopriming involves the treatment of seed with bioproducts that involves inoculation of seed with beneficial microbes followed by hydration [247]. In this process, the seed is treated with formulations of Trichoderma (6–10 gm/kg seed) and then incubated under moist and warm conditions prior to sowing for the emergence of radicals. During this process, germinated Trichoderma spores form a protective cover surrounding the seed’s coat and defend the seeds from adverse conditions, as well as lead to rapid and uniform germination and high establishment of the crop (good harvest yield and quality) [248,249]. Another study also evaluated the efficacy of T. asperellum BHUT8 biopriming on the growth promotion of seedlings. The finding showed that the treated seeds grew faster than the untreated seeds. The treated plant exhibited a considerable increase in various parameters such as length, dry weight, fresh weight (root and shoot), and leaf count, as compared to the control [250]. Rice seeds bioprimed with Trichoderma strain exhibited high seed germination, total chlorophyll content, and crop harvest [251]. Degani and Dor (2021) studied the effect of two strains of T. longibrachiatum and T. asperelloides on the maize seeds and observed that primed seeds showed several positive effects, including improved yield, significant enhancement in growth parameters and quality of crop as well as effective control of Magnaporthiopsis maydis, transmitted through soil [252]. In another study, strains of T. hebeiensis and T. erinaceum were used for priming of rice seeds, and it was found to reduce the time for germination, improve the seedling vigor, leaf chlorophyll content, and the yield by controlling four major rice pathogens R. solani, S. oryzae, S. rolfsii, and S. delphinii [253].
15.3. Root Dipping
It is well established that biocontrol agents, especially Trichoderma spore or slurry, reduce disease severity and enhance seedling growth in various cereals and vegetables such as rice, brinjal, chili, tomato, and capsicum [254]. Trichoderma can be applied to plant roots by dipping them in a Trichoderma spore suspension/slurry prior to transplanting, and this has resulted in successful control of diseases in different crops such as rice, chili, cucumber, and tomato [255,256].
15.4. Foliar Spraying/Wound Dressing
The phyllosphere, or the surface of the leaves, experiences fluctuations in humidity, temperature, dew, wind, rain, and radiation, which can affect the water potential of microbes; hence, it is crucial to consider the microclimate and environmental factors when using biocontrol agents for foliar diseases. These factors can also vary between different leaves or parts of a plant, which can affect the concentration of nutrients in the plant and ultimately impact the survival and efficacy of biocontrol agents. Studies have shown that foliar spray of Trichoderma viride can reduce linseed blight in controlled conditions [257,258]. Additionally, a mixed formulation of two different species of Trichoderma, such as T. harzianum, T. virens, and T. hamatum, T. viride, has been observed to be impactful with reasonable shelf life for foliar sprays [259]. Therefore, when selecting biocontrol agents for foliar diseases, it is important to consider their tolerance to environmental factors and their ability to survive and thrive under different microclimates [260].
The ability of Trichoderma consortia to establish in the phyllosphere of blue Passiflora has been demonstrated. Through foliar application of a combination of two Trichoderma species, namely T. asperellum and T. harzianum, a synergistic effect was observed. This combination has led to notable enhancements in plant physiological aspects, including increased leaf size and quantity, augmented chloroplast count, and heightened plant antioxidant activity [261]. Trichoderma can be sprayed on plants to suppress foliar pathogens and improve plant health. Studies have shown that foliar spraying with Trichoderma can ameliorate growth and yield in several plants, including cucumber, pepper, and tomato [262,263,264]. In another study, foliar spray of Trichoderma has been shown to facilitate growth and development in plants as well as resistance to diseases [265]. The effect of Trichoderma asperellum foliar spray on tomato plant growth, quantity, and quality was investigated. The results demonstrated that foliar application appreciably enhanced the plant height, leaf area, fruit weight, and total soluble solids in tomatoes [266].
15.5. Seed Material Treatment
For seed material treatment, 8–10 g of Trichoderma formulation is suspended in one liter of water, and the suspension is kept for 30 min before the treatment of seed material (before sowing and transplanting) in sugarcane setts, turmeric ginger rhizomes, banana suckers, and potato tubers. The disease-suppressing ability of different strains of Trichoderma for the control of ‘Panama disease’ in bananas (caused by F. oxysporum f. sp. cubense) was also studied [267]. The results showed that Trichoderma spp. considerably reduced disease occurrence and severity in banana plants. The biocontrol capacity of Trichoderma spp. against dry rot of potato tubers during storage was also studied. The application of Trichoderma spp. to potato tubers before storage significantly retarded the incidence of dry rot [268,269].
15.6. Soil Application
Soil application with Trichoderma is a sustainable and effective strategy for improving soil health, promoting plant growth, and suppressing soil-borne plant pathogens. The specific amount of Trichoderma formulation, either talcum-based (1–2 kg) or liquid (500–1000 mL), can be added to 25–50 kg FYM. The mixture should be mixed and covered with sugarcane leaves/ jute bag/paddy straw and placed for a few weeks in the shade to allow Trichoderma multiplication. Moisture should be maintained for better multiplication, and the mixture should be mixed at an interval of 3–4 days before spreading in the field. It should be applied to the field at least 15 days prior to sowing of the crop. The well-decomposed FYM can be applied in either in-furrow or pots at sowing/transplanting time. This quantity is sufficient for one acre of land. This method of soil application ensures that Trichoderma is close to the plant roots, where it can have the most beneficial impact. Jeyalakshmi et al. (2013) revealed that soil application of T. viride (2.5 kg/ha) is successful for disease control and significantly enhances crop yield [270]. To assess the bioefficacy of the biocontrol agent, T. harzianum was applied to soil as suspension (50 mL/L) to control root-rot pathogen affecting grapevine in a protected environment. This treatment declined 80% of infection of root-rot disease in grapevine infected by Macrophomina phaseolina [271]. The impact of T. harzianum on soil health and maize growth was noted. The finding showed that Trichoderma harzianum treatment considerably improved soil microbial diversity and increased the abundance of beneficial microorganisms [272,273], for example, nitrogen-fixing and phosphate-solubilizing bacteria (PSB). Trichoderma-treated soil also had more soil organic matter content and improved growth and yield of the crop, indicating improved soil function and overall plant performance. Wang et al. (2022) also examined the impact of Trichoderma harzianum on soil health as well as an antagonistic effect against Fusarium wilt in cucumber planting and showed that the Trichoderma treatment significantly retard the risk of Fusarium wilt of cucumber, and improved soil microbial diversity and activity [274]. The Trichoderma-treated soil also had higher soil enzyme activity and increased abundance of beneficial microorganisms, indicating enhanced soil health and decreased risk of soil-borne plant pathogens.
The synergism of Trichoderma and mycorrhiza consortia (T. harzianum and arbuscular mycorrhizae) were tested for soil health, colonization of root, and maize growth promotion in zinc-deficient soils. The finding showed that the combined application of both beneficial fungi (T. harzianum and arbuscular mycorrhizae) drastically improved soil health by increasing organic matter, enzyme activity, and microbial diversity in treated soil. Trichoderma-treated soil also had higher root colonization and improved maize growth and yield, indicating improved soil health and plant performance [275].
15.7. Cutting/Seedling Root Dip Application
Trichoderma application is a sustainable and environmentally friendly method that promotes plant growth and inhibits the growth of phytopathogens. In this application, 20–25 g of Trichoderma sp. (powder form) or 5–10 mL (liquid form) are mixed in 1 L of water for 30 min. The roots and cuttings of seedlings are immersed in the prepared suspension for 1/2 h and then immediately transplanted. This treatment can effectively control soil-borne diseases. A study investigated the impact of T. harzianum on growth parameters of olive cuttings. The results showed that T. harzianum deployment considerably enhanced the rooting percentage, root length, and shoot length of the cuttings. The Trichoderma-treated cuttings also had higher chlorophyll content and lower incidence of fungal pathogens, indicating improved plant health [276].
15.8. Nursery Bed Treatment
Nursery bed treatment with Trichoderma is an effective method to promote healthy seedlings by suppressing soil-borne pathogens and improving plant growth. In this method, 500-g Trichoderma-talcum-based formulation is mixed with 10–20 kg completely decomposed farm yard manure/vermicast. It should be applied in the evening time, and this quantity is sufficient for a one-acre area. Before its application, appropriate moisture is required for its better multiplication. A study confirmed that Trichoderma-enriched compost used as a nursery bed amendment considerably enhanced the yield and growth of tomato seedlings [277]. Trichoderma-enriched compost also enhanced the soil fertility and increased the soil enzyme activity, indicating improved soil health. In another study, the effect of nursery bed soil amendment with Trichoderma was evaluated on the yield and quality of seedlings in different crops such as brinjal, tomato, and chili. The findings demonstrated that the nursery bed treated with Trichoderma greatly enhanced seedling development and decreased the prevalence of soil-borne pathogens. The Trichoderma treatment also increased soil enzyme activity and nutrient availability, indicating improved soil health [278]. The biocontrol effect of nursery bed treatment with Trichoderma on the tomato seedlings was investigated. The results depicted that the Trichoderma treatment significantly improved seedling growth and increased the nutrient content of the seedlings [279]. The seedling also had higher chlorophyll content and reduced the incidence of damping-off disease, indicating improved plant health.
15.9. Soil Drenching
In this method, 1–2 kg Trichoderma formulations are suspended in 200 liters of fresh water and directly applied onto the soil around the base of the plant. This method introduces Trichoderma into the soil, which protects the crop from infection caused by damping off soil-borne pathogens. During the application, optimum soil moisture is maintained in the field. Studies have shown that soil drenching with Trichoderma can improve plant growth and yield in various crops, including maize, tomato, and pepper [280,281].
16. Other Applications of Trichoderma
The versatility of Trichoderma makes it possible to be widely used in various industries. This fungus provides the opportunity for their use in bioremediation, biodegradation of agricultural waste, wine-making, brewery industries, food additives, textile industry, paper, and pulp industry, and the production of bioethanol from farm waste. Trichoderma produces various lytic enzymes that can be used in the animal feed, alcohol, and brewing industries. Cellulase, hemicellulase, and pectinase produced by Trichoderma can be used to partially hydrolyze food walls to improve the nutritional value and digestion of food. This increases animal weight and milk production [282]. In another study, it has been reported that the paper industry may use Trichoderma for the modification of fiber properties and to reduce the amount of lignin in pulp [282]. Kaczmarek and Jedryczka (2011) have reported the potential of Trichoderma to control the pathogen Leptosphaeria maculans and L. biglobosa, which cause stem canker of Brassica [283].
Recently, it has been found that Trichoderma spp. have the ability to metabolize many kinds of pesticides and thus can facilitate bioremediation [284]. Vázquez et al. (2015) revealed that T. harzianum strain successfully detoxifies metsulfuron methyl, which is a type of sulfonylurea herbicide [285]. Trichoderma spp. are also effective natural decomposers that speed up the breakdown of organic materials. According to Amira et al. (2011), Trichoderma spp. considerably accelerate the decomposition of food and oil industry waste [286]. Combined application of T. harzianum and Phanerochaete chrysosporium in olive pomace compost could successfully break down its cellulose and lignin [287].
17. Major Challenges and Future Strategies on Biopesticides
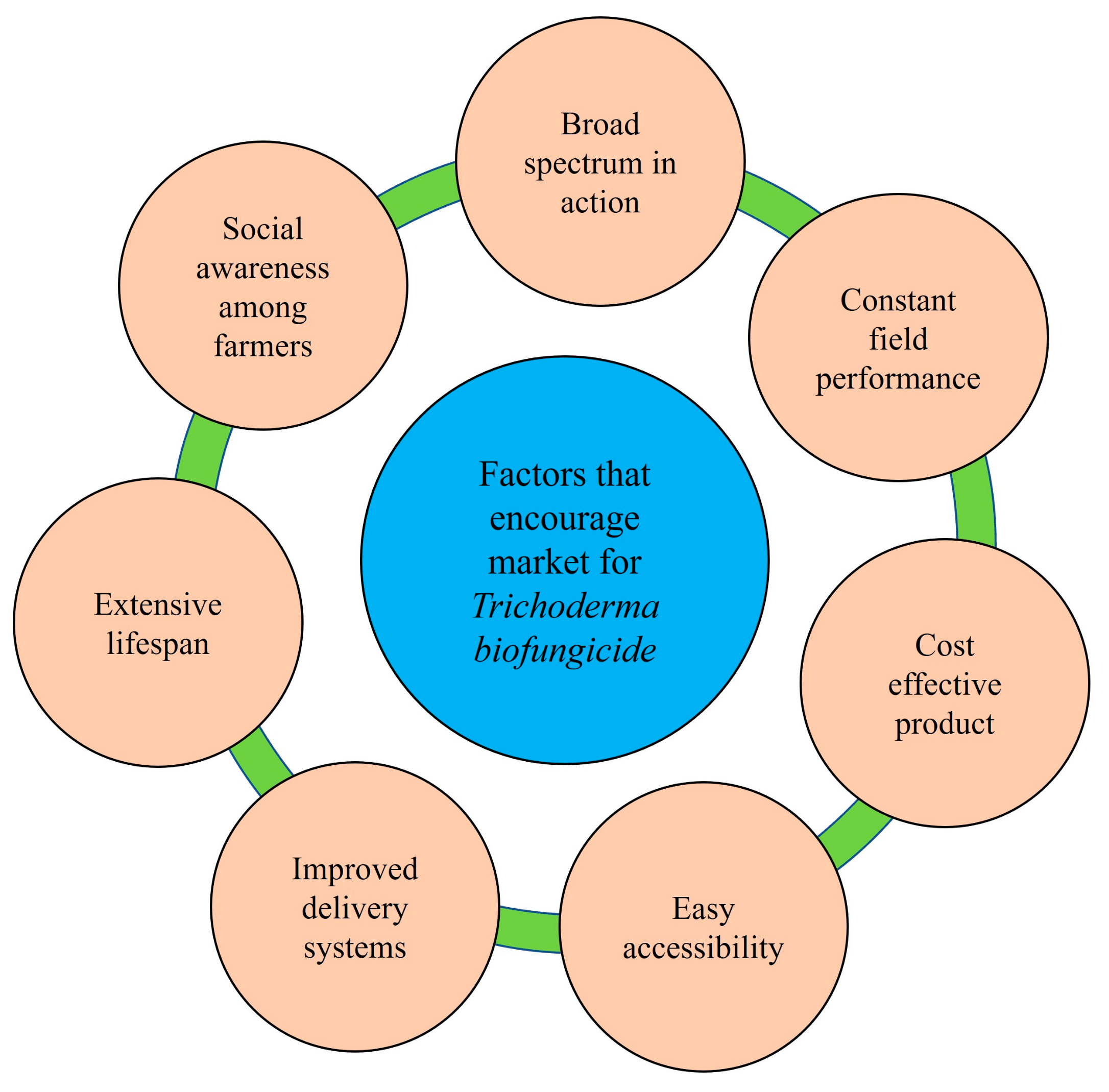
Biofungicides are recognized as safe because they are target-specific, minimize the risk of ecological pollution, and eliminate the problems of resistance development in pathogens. The desire for safe, disease-free food is driving the popularity of biopesticides. Despite these advantages, the market for biopesticides is in its infancy, particularly in India. These challenges and limitations necessitate better strategies and technical solutions that eliminate the flaws of commercial biopesticides [288]. Biopesticide incorporation in the mainstream of crop production requires a better understanding of mechanisms to enhance their delivery systems against targeted plant pathogens, cost-effective product, consistent field performance, longer shelf-life, easily accessible, social awareness among farmers, and trouble-free registration and regulation policies [289,290] (Figure 2).
18. Constraints in the Development of Trichoderma-Based Biopesticide
There are certain constraints in the development of Trichoderma-based biopesticide: (i) lack of an appropriate screening protocol for selecting promising Trichoderma candidates, (ii) inadequate knowledge regarding the Trichoderma ecology and other phytopathogens pathogens, (iii) fermentation technological advancement, coupled with the implementation of large-scale production methods, (iv) inconsistent performance and a short storage life, (v) inadequate patent protection, (vi) registration costs are exorbitant, (vii) training, awareness, and education deficiencies, (viii) an absence of a comprehensive multidisciplinary approach, (ix) technological constraints, and (x) introduction and size of market [291]. Moreover, most of the serious obstacles against the development and commercialization of Trichoderma-based biopesticides is their poor shelf-life [292].
19. Future Prospects of Trichoderma Application
Trichoderma spp. is utilized as a biopesticide for the management of diseases in various crops. Its main role is not only limited to disease suppression and enhancing plant growth, but this fungus can also be helpful in the decomposition of agricultural residue or remediation of soil toxicity. Every day, increasing awareness of ecology, environment protection, sustainable economy, and interest in substitute sources of energy create a possibility of biofuel production from agricultural waste by the addition of CWDs like cellulases, hemicellulases, ligninolytic, etc., which are produced by Trichoderma reesei and this is known as second-generation biofuels. Therefore, Trichoderma can be used in various ways: it can be used in controlling plant diseases, in plant growth promotion, enrichment of the compost, as well as for providing an ecofriendly environment for farming systems. However, several new strains of Trichoderma have been isolated, but their genome sequencing and characterization have to be performed for further implementation. Furthermore, the application of “omics” technology should be introduced for accurate identification of new or existing species of Trichoderma. This technique can also be helpful for the detection of bioactive metabolites at genomic and transcriptomic levels [293]. Apart from this, further studies need to be conducted so as to understand how these biological products influence the environmental microbiome processes. Several reports are present which suggest the role of Trichoderma in the remediation of organic solvents [294], heavy metals [295], organo-phosphate insecticide [296], crude fossil oil, benzopyrene and naphthalene [297], Phenanthrene and pyrene [298], cyanide [299], etc., from the fresh water and soil. T. viride and T. asperellum can remove arsenic by in vitro biovolatilization, and T. viride may be successful for in vitro bioremediation of Cd and Pb. Trichoderma has been shown to support the uptake and translocation of nitrate and various toxic ions in the rhizosphere, thereby promoting the assimilation of metals and plant extracts [300]. In a study, T. atroviride has been reported to enhance the uptake and translocation of Cd, Zn, and Ni in mustard crops, while T. harzianum promoted the growth of Salix fragilis in soil contaminated with heavy metals [301]. T. harzianum can degrade the harmful compound potassium cyanide, promoting root growth and branching in Pteris fern, which is known as an arsenic hyper-accumulatory fern [302]. In addition, Trichoderma is also capable of biodegrading toxic organic pollutants such as cyanide, nitrate, phenols, polycyclic aromatic hydrocarbons (PAH), dichlorodiphenyltrichloroethane (DDT), and other artificial dyes, endosulfan, pentachlorophenol [2,303]. The immobilized biomass of Trichoderma BSCR02 in calcium alginate beads facilitates the biosorption of Cr(VI) [304]. Therefore, Trichoderma spp. can be utilized as a low-priced treatment for wastewater and heavy metal-contaminated water. However, it is clear that future bioremediation research will need to combine different approaches, including genetic, molecular, biochemical, ecological, and technological development. For better utilization of beneficial microorganisms, particularly new strains of Trichoderma, it is crucial to foster a deeper understanding of their biology. Genetic manipulation, mutagenesis, and fusion of protoplast in Trichoderma spp. shall open up new vistas for better performance and fixing the associated problems. Cultivation of Trichoderma to treat infected sites could materialize in the near future, as it can be used as an inexpensive biocontrol agent in many parts of the world.
Next-generation bioproducts also create a new option for sustainable agriculture in solving industry and university research problems. For example, identifying novel Trichoderma strains can help or promote the production of bioproducts or the use of bioenergy. Increased awareness about eco-friendly farming, where the outcomes should speak to the needs and concerns of farmers and industry partners. Farmers can be ready to implement this approach when there are short-term economic benefits and profits in agriculture [305]. Any biological plan must closely adhere to the aims of integrated disease management (IDM), such as ecofriendly, long viability, and social acceptance. These aims address the needs and challenges of the farmers, suppliers, and customers. Numerous beneficial compounds have already been isolated from Trichoderma spp., which is helpful to control plant diseases. Still, these findings could not be converted into active ingredients at the commercial level. Experts from different fields should be involved in the development of any biopesticide. For example, for the commercialization of a potential biopesticide, an ideal team should be involved in the execution, and the researcher should know the status of the market, the chemistry of the product, analysis, and understanding of plant biology/pathology, microbes, pests, etc. There are several publications on novel features of Trichoderma strains. With this large collection, there is a need for additional high-quality genomes and depictions of the many parts of Trichoderma. Overall, this inclusive roadmap can provide us with direction for the upcoming production of bioproducts from Trichoderma. Simultaneously, future research should be centralized on field application of encapsulated formulation of Trichoderma, screening, identification of Trichoderma spp. for control of foliar diseases, longer shelf-life, consistency of field performance, risk assessment, which is one of the greatest obstacles in the registration process of Trichoderma formulation as well as optimization of the cost of formulation and their marketing.
In a nutshell, future studies should concentrate on the improvement and utilization of Trichoderma as an antagonist for disease control in crops. It is formulated for long storage life, performance on a farm, and suitability for unfavorable conditions. The production process in the state can be increased by the cooperation of various industries. Good practices include large-scale agricultural biocontrol technology demonstration, good management, fast registration, identification of suitable organisms for different soils and environments (temperature/low humidity/salt water), and combination of two or more agents to improve performance.
20. Conclusions
Globally, the use of chemical pesticides is severely constrained by rising environmental concerns and the legislation that results from them, necessitating the urgent development of safer and more ecologically friendly solutions. Microbial BCAs have become a potential method for helping plants resist infections. It is now generally acknowledged that formulations are crucial in assisting microbiological BCAs to travel the arduous route between laboratory testing and field application.
Microbial BCAs have traditionally been manufactured as liquids, wettable powders, and granules for use as foliar sprays, soil drenches, or seed treatments. However, combining sophisticated technology from other fields with knowledge of biocontrol formulations (such as spray drying and encapsulation) may result in the creation of more effective and reliable formulations. Technically speaking, this calls for a profound comprehension of the interactions that take place between the BCA, formulation components, and the environment, as well as a comprehension of practical applicability. Recommendations on efficacy evaluation and field testing should be regularly updated while developing sophisticated methodologies for the development and deployment of biocontrol formulations to help provide proof of efficacy and facilitate the commercialization of biopesticides. Commercialization of biocontrol products is steadily growing worldwide, but in the absence of effective regulations, penetration of biological products in the agricultural market is still slow, especially in the European Union, where the registration process is complex and time-consuming. Existing regulations of biopesticides vary from country to country, and requirements for registration are often similar to those of chemical substances. The global harmonization of specific biopesticide regulations would undoubtedly overcome these limitations and encourage the commercialization of microbial-based biopesticides worldwide. To date, the major steps towards this harmonization have been taken by various global agencies such as the Organization for Economic and Cooperative Development, International Organization for Biological Control, and EPPO, but success is limited.
Given that climate change may alter patterns of pest and disease introduction, unification of policymaking procedures is even more essential [306]. It has also been projected that if global warming keeps up, a number of phytopathogens may move to other regions where they would come into contact with new hosts. Similar to animals, plants may become less resistant to disease attacks as a result of climate change [306].
The containment of some pests and illnesses may not be possible in worst-case scenarios when chemicals have been outlawed and/or plants are more sensitive in the absence of efficient microbial-based biopesticides. It is also important to keep in mind that environmental conditions affect, no matter how effective BCAs are. The improvement of BCA manufacturing, formulation, and delivery should be the main focus of future research. In addition, simpler, universal rules and demonstration trials with high-quality products are essential to advancing the commercialization of biopesticides.
Trichoderma spp. is an important biocontrol agent as it can increase crop production through various activities such as plant growth promotion and enhanced nutrient uptake capacity by inhibiting pathogens. Nonetheless, fast research in this field needs to be carefully done to ensure its safe commercialization. Effective production methods are also essential to improve the commercialization of Trichoderma. The search for less expensive and alternative substrates, as well as ideal operation parameters to boost conidia production, is also needed. Thus, it is anticipated that consistent research towards highly effective biocontrol formulations for enhanced agricultural production will pay off, displace agricultural pesticides, and ensure sustainable agricultural productivity.
Author Contributions
Conceptualization, V.K., BK. and M.S.; writing—original draft preparation, V.K., B.K. and P.T.; writing—review and editing, V.K., B.K., P.T., D.Y. and M.S.; supervision, B.K and M.S.; funding acquisition, B.K. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the 2023 Yeungnam University Research Grants.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological Control of Plant Diseases: An Evolutionary and Eco-Economic Consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Larralde-Corona, C.P.; Santiago-Mena, M.R.; Sifuentes-Rincón, A.M.; Rodríguez-Luna, I.C.; Rodríguez-Pérez, M.A.; Shirai, K.; Narváez-Zapata, J.A. Biocontrol potential and polyphasic characterization of novel native Trichoderma strains against Macrophomina phaseolina isolated from sorghum and common bean. Appl. Microbiol. Biotechnol. 2008, 80, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Bueno, V.H.P.; Parra, J.R.P.; Bettiol, W.; Lenteren, J.v. Biological control in Brazil. In Biological Control in Latin America and the Caribbean: Its Rich History and Bright Future; CABI: Wallingford, UK, 2020; pp. 78–107. [Google Scholar]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, S.; Safaie, N.; Shahbazi, S.; Shams-Bakhsh, M.; Askari, H. The Role of Cell Wall Degrading Enzymes in Antagonistic Traits of Trichoderma virens Against Rhizoctonia solani. Iran. J. Biotechnol. 2020, 18, e2333. [Google Scholar]
- Guzmán-Guzmán, P.; Kumar, A.; de Los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.D.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma species: Our best fungal allies in the biocontrol of plant diseases—A review. Plants 2023, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Monte, E.; Bettiol, W.; Hermosa, R. Trichoderma e seus mecanismos de ação para o controle de doenças de plantas. In Trichoderma: Usos na Agricultura; Meyer, M.C., Mazaro, S.M., da Silva, J.C., Eds.; Embrapa Soja: Brasília, Brazil, 2019; pp. 181–200. [Google Scholar]
- De Rezende, L.C.; de Andrade Carvalho, A.L.; Costa, L.B.; de Almeida Halfeld-Vieira, B.; Silva, L.G.; Pinto, Z.V.; Morandi, M.A.B.; de Medeiros, F.H.V.; Mascarin, G.M.; Bettiol, W. Optimizing mass production of Trichoderma asperelloides by submerged liquid fermentation and its antagonism against Sclerotinia sclerotiorum. World J. Microbiol. Biotechnol. 2020, 36, 113. [Google Scholar] [CrossRef]
- Harman, G.; Jin, X.; Stasz, T.; Peruzzotti, G.; Leopold, A.; Taylor, A. Production of conidial biomass of Trichoderma harzianum for biological control. Biol. Control 1991, 1, 23–28. [Google Scholar] [CrossRef]
- Mawar, R.; Manjunatha, B.; Kumar, S. Commercialization, diffusion and adoption of bioformulations for sustainable disease management in indian arid agriculture: Prospects and challenges. Circ. Econ. Sust. 2021, 1, 1367–1385. [Google Scholar] [CrossRef] [PubMed]
- Howell, C. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Papavizas, G. Trichoderma and Gliocladium: Biology, ecology, and potential for biocontrol. Annu. Rev. Phytopathol. 1985, 23, 23–54. [Google Scholar] [CrossRef]
- Persoon, C.H. Disposita methodical fungorum. Romers. Neues. Mag. Bot. 1794, 1, 81–128. [Google Scholar]
- Tulasne, L.-R.; Tulasne, C. Selecta Fungorum Carpologia, 2; Imperial: Paris, France, 1863. [Google Scholar]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Weindling, R. Studies on a lethal principle effective in the parasitic action of Trichoderma lignorum on Rhizoctonia solani and other soil fungi. Phytopathology 1934, 24, 1153–1179. [Google Scholar]
- Gutter, Y. Effect of light in sporulation of Trichoderma viride. Bull. Res. Council Israel Sect. D 1957, 5, 273–286. [Google Scholar]
- Wells, H.D. Efficacy of Trichoderma harzianum as a biocontrol for Sclerotium rolfsii. Phytopathology 1972, 62, 442. [Google Scholar] [CrossRef]
- Montenecourt, B.S.; Eveleigh, D.E. Selective screening methods for the isolation of high yielding cellulase mutants of Trichoderma reesei. Adv. Chem. 1979, 181, 289–301. [Google Scholar]
- Shoemaker, S.; Schweickart, V.; Ladner, M.; Gelfand, D.; Kwok, S.; Myambo, K.; Innis, M. Molecular cloning of exo–cellobiohydrolase I derived from Trichoderma reesei strain L27. Biotechnology 1983, 1, 691–696. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Chang, Y.-C.; Baker, R. Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Penttilä, M.; Nevalainen, H.; Rättö, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Harkki, A.; Uusitalo, J.; Bailey, M.; Penttilä, M.; Knowles, J.K. A novel fungal expression system: Secretion of active calf chymosin from the filamentous fungus Trichoderma reesei. Biotechnology 1989, 7, 596–603. [Google Scholar] [CrossRef]
- Inbar, J.; Chet, I. Biomimics of fungal cell-cell recognition by use of lectin-coated nylon fibers. J. Bacteriol. 1992, 174, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Geremia, R.A.; Goldman, G.H.; Jacobs, D.; Ardrtes, W.; Vila, S.B.; Van Montagu, M.; Herrera-Estrella, A. Molecular characterization of the proteinase-encoding gene, prb1, related to mycoparasitism by Trichoderma harzianum. Mol. Microbiol. 1993, 8, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Bigirimana, J.; De Meyer, G.; Poppe, J.; Elad, Y.; Höfte, M. Induction of systemic resistance on bean (Phaseolus vulgaris) by Trichoderma harzianum. Meded. Fac. Landbouwkd. Toegepaste Biol. Wet. Univ. Gent. 1997, 62, 1001–1007. [Google Scholar]
- Cortes, C.; Gutierrez, A.; Olmedo, V.; Inbar, J.; Chet, I.; Herrera-Estrella, A. The expression of genes involved in parasitism by Trichoderma harzianum is triggered by a diffusible factor. Mol. Gen. Genet. 1998, 260, 218–225. [Google Scholar] [CrossRef]
- Yedidia, I.; Benhamou, N.; Chet, I. Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Appl. Environ. Microbiol. 1999, 65, 1061–1070. [Google Scholar] [CrossRef]
- Rocha-Ramírez, V.; Omero, C.; Chet, I.; Horwitz, B.A.; Herrera-Estrella, A. Trichoderma atroviride G-protein α-subunit gene tga1 is involved in mycoparasitic coiling and conidiation. Eukaryot. Cell 2002, 1, 594–605. [Google Scholar] [CrossRef]
- Mendoza-Mendoza, A.; Pozo, M.J.; Grzegorski, D.; Martínez, P.; García, J.M.; Olmedo-Monfil, V.; Cortés, C.; Kenerley, C.; Herrera-Estrella, A. Enhanced biocontrol activity of Trichoderma through inactivation of a mitogen-activated protein kinase. Proc. Natl. Acad. Sci. USA 2003, 100, 15965–15970. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Latha, J.; Hadar, R.; Horwitz, B.A. TmkA, a mitogen-activated protein kinase of Trichoderma virens, is involved in biocontrol properties and repression of conidiation in the dark. Eukaryot. Cell 2003, 2, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Casas-Flores, S.; Rios-Momberg, M.; Bibbins, M.; Ponce-Noyola, P.; Herrera-Estrella, A. BLR-1 and BLR-2, key regulatory elements of photoconidiation and mycelial growth in Trichoderma atroviride. Microbiology 2004, 150, 3561–3569. [Google Scholar] [CrossRef] [PubMed]
- Viterbo, A.; Harel, M.; Horwitz, B.A.; Chet, I.; Mukherjee, P.K. Trichoderma mitogen-activated protein kinase signaling is involved in induction of plant systemic resistance. Appl. Environ. Microbiol. 2005, 71, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Djonović, S.; Pozo, M.J.; Dangott, L.J.; Howell, C.R.; Kenerley, C.M. Sm1, a proteinaceous elicitor secreted by the biocontrol fungus Trichoderma virens induces plant defense responses and systemic resistance. Mol. Plant Microbe Interact. 2006, 19, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Stricker, A.R.; Grosstessner-Hain, K.; Würleitner, E.; Mach, R.L. Xyr1 (xylanase regulator 1) regulates both the hydrolytic enzyme system and D-xylose metabolism in Hypocrea jecorina. Eukaryot. Cell 2006, 5, 2128–2137. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Sicher, R.C.; Kim, M.S.; Kim, S.-H.; Strem, M.D.; Melnick, R.L.; Bailey, B.A. The beneficial endophyte Trichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in Theobroma cacao. J. Exp. Bot. 2009, 60, 3279–3295. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kenerley, C.M. Regulation of morphogenesis and biocontrol properties in Trichoderma virens by a VELVET protein, Vel1. Appl. Environ. Microbiol. 2010, 76, 2345–2352. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Herrera-Estrella, A.; Seidl-Seiboth, V.; Martinez, D.A.; Druzhinina, I.S.; Thon, M.; Zeilinger, S.; Casas-Flores, S.; Horwitz, B.A.; Mukherjee, P.K. Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol. 2011, 12, R40. [Google Scholar] [CrossRef]
- Schuster, A.; Bruno, K.S.; Collett, J.R.; Baker, S.E.; Seiboth, B.; Kubicek, C.P.; Schmoll, M. A versatile toolkit for high throughput functional genomics with Trichoderma reesei. Biotechnol. Biofuels 2012, 5, 1–10. [Google Scholar] [CrossRef]
- Zhang, G.-Z.; Yang, H.-T.; Zhang, X.-J.; Zhou, F.-Y.; Wu, X.-Q.; Xie, X.-Y.; Zhao, X.-Y.; Zhou, H.-Z. Five new species of Trichoderma from moist soils in China. MycoKeys 2022, 87, 133. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases—What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Sarrocco, S. Biological Disease Control by Beneficial (Micro)Organisms: Selected Breakthroughs in the Past 50 Years. Phytopathology 2023, 113, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, S.; Omann, M. Trichoderma biocontrol: Signal transduction pathways involved in host sensing and mycoparasitism. Gene Regul. Syst. Bio. 2007, 1, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.J.; Bruckart, W.L.; Coulson, J.R.; Goettel, M.S.; Humber, R.A.; Lumsden, R.D.; Maddox, J.V.; McManus, M.L.; Moore, L.; Meyer, S.F. Safety of microorganisms intended for pest and plant disease control: A framework for scientific evaluation. Biol. Control 1996, 7, 333–351. [Google Scholar] [CrossRef]
- Elad, Y.; Chet, I.; Boyle, P.; Henis, Y. Parasitism of Trichoderma spp. on Rhizoctonia solani and Sclerotium rolfsii-scanning electron microscopy and fluorescence microscopy. Phytopathology 1983, 73, 85–88. [Google Scholar] [CrossRef]
- Kelley, W.D. Interactions of Phytophthora cinnamomi and Trichoderma spp. in relation to propagule production in soil cultures at 26 °C. Can. J. Microbiol. 1977, 23, 288–294. [Google Scholar] [CrossRef]
- McLeod, A.; Labuschagne, N.; Kotzé, J. Evaluation of Trichoderma for biological control of avocado root rot in bark medium artificially infested with Phytophthora cinnamomi. S. Afr. Avocado Grow. Assoc. Yearb. 1995, 18, 32–37. [Google Scholar]
- Bhale, U.; Wagh, P.; Rajkonda, J. Antagonistic confrontation of Trichoderma spp. against fruit rot pathogens on Sapodilla (Manilkara zapota L.). J. Yeast Fungal Res. 2013, 4, 5–11. [Google Scholar]
- Ben Amira, M.; Lopez, D.; Triki Mohamed, A.; Khouaja, A.; Chaar, H.; Fumanal, B.; Gousset-Dupont, A.; Bonhomme, L.; Label, P.; Goupil, P.; et al. Beneficial effect of Trichoderma harzianum strain Ths97 in biocontrolling Fusarium solani causal agent of root rot disease in olive trees. Biol. Control 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Khare, A.; Singh, B.; Upadhyay, R. Biological control of Pythium aphanidermatum causing damping-off of mustard by mutants of Trichoderma viride 1433. J. Agric. Technol. 2010, 6, 231–243. [Google Scholar]
- Muthukumar, A.; Eswaran, A.; Sanjeevkumas, K. Exploitation of Trichoderma species on the growth of Pythium aphanidermatum in chilli. Braz. J. Microbiol. 2011, 42, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Fajola, A.; Alasoadura, S. Antagonistic effects of Trichoderma harzianum on Pythium aphanidermatum causing the damping-off disease of tobacco in Nigeria. Mycopathologia 1975, 57, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Thesiya, M.; Rakholiya, K.; Lokesh, R.; Shekhda, M. Comparative Efficacy of Fungicides and Biocontrol Agents against Stem Blight and Fruit Rot Disease of Brinjal under Pot Culture Conditions. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 2020. [Google Scholar]
- Arzanlou, M.; Khodaei, S.; Narmani, A.; Babai-Ahari, A.; Azar, A.M. Inhibitory effect of Trichoderma isolates on growth of Alternaria alternata, the causal agent of leaf spot disease on sunflower, under laboratory conditions. Arch. Phytopathol. Plant Prot. 2014, 47, 1592–1599. [Google Scholar] [CrossRef]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Hossain, K. Trichoderma spp.: A biocontrol agent for sustainable management of plant diseases. Pak. J. Bot. 2014, 46, 1489–1493. [Google Scholar]
- Bhandari, C.; Karuna, V. Screening of different isolates of Trichoderma harzianum and Pseudomonas fluorescens against Fusarium moniliforme infecting maize. Pantnagar J. Res. 2013, 11, 243–247. [Google Scholar]
- Lenka, S.; Mishra, S.; Mohanty, S.; Das, K.; Medhi, B. Bio-control of rice sheath blight through antagonists. ORYZA-Int. J. Rice 2012, 49, 68–69. [Google Scholar]
- Abbas, A.; Mubeen, M.; Zheng, H.; Sohail, M.A.; Shakeel, Q.; Solanki, M.K.; Iftikhar, Y.; Sharma, S.; Kashyap, B.K.; Hussain, S.; et al. Trichoderma spp. Genes Involved in the Biocontrol Activity Against Rhizoctonia solani. Front. Microbiol. 2022, 13, 884469. [Google Scholar] [CrossRef] [PubMed]
- Emani, C.; Garcia, J.M.; Lopata-Finch, E.; Pozo, M.J.; Uribe, P.; Kim, D.-J.; Sunilkumar, G.; Cook, D.R.; Kenerley, C.M.; Rathore, K.S. Enhanced fungal resistance in transgenic cotton expressing an endochitinase gene from Trichoderma virens. Plant Biotechnol. J. 2003, 1, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Marcello, C.M.; Steindorff, A.S.; da Silva, S.P.; do Nascimento Silva, R.; Bataus, L.A.M.; Ulhoa, C.J. Expression analysis of the exo-β-1, 3-glucanase from the mycoparasitic fungus Trichoderma asperellum. Microbiol. Res. 2010, 165, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Latha, J.; Hadar, R.; Horwitz, B.A. Role of two G-protein alpha subunits, TgaA and TgaB, in the antagonism of plant pathogens by Trichoderma virens. Appl. Environ. Microbiol. 2004, 70, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Morán-Diez, E.; Hermosa, R.; Ambrosino, P.; Cardoza, R.E.; Gutiérrez, S.; Lorito, M.; Monte, E. The ThPG1 endopolygalacturonase is required for the Trichoderma harzianum–plant beneficial interaction. Mol. Plant Microbe Interact. 2009, 22, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.; Cardoza, R.; Hauser, M.; Hermosa, R.; Rey, M.; Llobell, A.; Becker, J.; Gutiérrez, S.; Monte, E. ThPTR2, a di/tri-peptide transporter gene from Trichoderma harzianum. Fungal Genet. Biol. 2006, 43, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Tijerino, A.; Cardoza, R.E.; Moraga, J.; Malmierca, M.G.; Vicente, F.; Aleu, J.; Collado, I.G.; Gutiérrez, S.; Monte, E.; Hermosa, R. Overexpression of the trichodiene synthase gene tri5 increases trichodermin production and antimicrobial activity in Trichoderma brevicompactum. Fungal Genet. Biol. 2011, 48, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Cardoza, R.E.; Malmierca, M.G.; Gutiérrez, S. Overexpression of erg1 gene in Trichoderma harzianum CECT 2413: Effect on the induction of tomato defence-related genes. J. Appl. Microbiol. 2014, 117, 812–823. [Google Scholar] [CrossRef]
- Dixit, P.; Mukherjee, P.K.; Ramachandran, V.; Eapen, S. Glutathione transferase from Trichoderma virens enhances cadmium tolerance without enhancing its accumulation in transgenic Nicotiana tabacum. PLoS ONE 2011, 6, e16360. [Google Scholar] [CrossRef]
- Zhong, Y.H.; Wang, T.H.; Wang, X.L.; Zhang, G.T.; Yu, H.N. Identification and characterization of a novel gene, TrCCD1, and its possible function in hyphal growth and conidiospore development of Trichoderma reesei. Fungal Genet. Biol. 2009, 46, 255–263. [Google Scholar] [CrossRef]
- Migheli, Q. Biodiversity of the Genus Trichoderma and Identification of Marker Genes Involved in the Antagonism between Trichoderma spp. and Plant Pathogenic Fungi. Ph.D. Thesis, University of Sassari, Sassari, Italy, 2008. [Google Scholar]
- Samolski, I.; Rincon, A.M.; Pinzon, L.M.; Viterbo, A.; Monte, E. The qid74 gene from Trichoderma harzianum has a role in root architecture and plant biofertilization. Microbiology 2012, 158, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a new biocontrol gene in Trichoderma atroviride: The role of an ABC transporter membrane pump in the interaction with different plant-pathogenic fungi. Mol. Plant Microbe Interact. 2009, 22, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Mukherjee, P.K.; Kale, S.P. cAMP signalling is involved in growth, germination, mycoparasitism and secondary metabolism in Trichoderma virens. Microbiology 2007, 153, 1734–1742. [Google Scholar] [CrossRef] [PubMed]
- Donzelli, B.G.G.; Lorito, M.; Scala, F.; Harman, G. Cloning, sequence and structure of a gene encoding an antifungal glucan 1, 3-β-glucosidase from Trichoderma atroviride (T. harzianum). Gene 2001, 277, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Q.; Song, J. A new serine protease gene from Trichoderma harzianum is expressed in Saccharomyces cerevisiae. Appl. Biochem. Microbiol. 2009, 45, 22–26. [Google Scholar] [CrossRef]
- Carpenter, M.A.; Ridgway, H.J.; Stringer, A.M.; Hay, A.J.; Stewart, A. Characterisation of a Trichoderma hamatum monooxygenase gene involved in antagonistic activity against fungal plant pathogens. Curr. Genet. 2008, 53, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Kim, B.-G.; Lee, C.; Hur, H.-G.; Ahn, J.-H. Purification, characterization, and cDNA cloning of xylanase from fungus Trichoderma strain SY. J. Microbiol. Biotechnol. 2002, 12, 890–894. [Google Scholar]
- Migheli, Q.; González-Candelas, L.; Dealessi, L.; Camponogara, A.; Ramón-Vidal, D. Transformants of Trichoderma longibrachiatum overexpressing the β-1, 4-endoglucanase gene egl1 show enhanced biocontrol of Pythium ultimum on cucumber. Phytopathology 1998, 88, 673–677. [Google Scholar] [CrossRef]
- Trushina, N.; Levin, M.; Mukherjee, P.K.; Horwitz, B.A. PacC and pH–dependent transcriptome of the mycotrophic fungus Trichoderma virens. BMC Genom. 2013, 14, 138. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef]
- Gajera, H.; Hirpara, D.G.; Katakpara, Z.A.; Patel, S.; Golakiya, B. Molecular evolution and phylogenetic analysis of biocontrol genes acquired from SCoT polymorphism of mycoparasitic Trichoderma koningii inhibiting phytopathogen Rhizoctonia solani Kuhn. Infect. Genet. Evol. 2016, 45, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Yang, P. The gene task1 is involved in morphological development, mycoparasitism and antibiosis of Trichoderma asperellum. Biocontrol. Sci. Technol. 2017, 27, 620–635. [Google Scholar] [CrossRef]
- Atanasova, L.; Gruber, S.; Lichius, A.; Radebner, T.; Abendstein, L.; Münsterkötter, M.; Stralis-Pavese, N.; Łabaj, P.P.; Kreil, D.P.; Zeilinger, S. The Gpr1-regulated Sur7 family protein Sfp2 is required for hyphal growth and cell wall stability in the mycoparasite Trichoderma atroviride. Sci. Rep. 2018, 8, 12064. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valdespino, C.A.; Porras-Troncoso, M.D.; Corrales-Escobosa, A.R.; Wrobel, K.; Martínez-Hernández, P.; Olmedo-Monfil, V. Functional characterization of TvCyt2, a member of the p450 monooxygenases from Trichoderma virens relevant during the association with plants and mycoparasitism. Mol. Plant Microbe Interact. 2018, 31, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Suriani Ribeiro, M.; Graciano de Paula, R.; Raquel Voltan, A.; de Castro, R.G.; Carraro, C.B.; José de Assis, L.; Stecca Steindorff, A.; Goldman, G.H.; Silva, R.N.; Ulhoa, C.J. Endo-β-1, 3-glucanase (GH16 family) from Trichoderma harzianum participates in cell wall biogenesis but is not essential for antagonism against plant pathogens. Biomolecules 2019, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Rivera, M.; Hernández-Oñate, M.Á.; Dautt-Castro, M.; Gallardo-Negrete, J.d.J.; Rebolledo-Prudencio, O.G.; Uresti-Rivera, E.E.; Arenas-Huertero, C.; Herrera-Estrella, A.; Casas-Flores, S. IPA-1 a putative chromatin remodeler/helicase-related protein of Trichoderma virens plays important roles in antibiosis against Rhizoctonia solani and induction of Arabidopsis systemic disease resistance. Mol. Plant Microbe Interact. 2020, 33, 808–824. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum xylanases promote growth and induce resistance in poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar] [CrossRef]
- Dubey, M.; Jensen, D.F.; Karlsson, M. Functional characterization of the AGL1 aegerolysin in the mycoparasitic fungus Trichoderma atroviride reveals a role in conidiation and antagonism. Mol. Genet. Genom. 2021, 296, 131–140. [Google Scholar] [CrossRef]
- Harman, G.E. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Manzar, N.; Kashyap, A.S.; Goutam, R.S.; Rajawat, M.V.S.; Sharma, P.K.; Sharma, S.K.; Singh, H.V. Trichoderma: Advent of versatile biocontrol agent, its secrets and insights into mechanism of biocontrol potential. Sustainability 2022, 14, 12786. [Google Scholar] [CrossRef]
- Chet, I.; Inbar, J.; Hadar, I. Fungal antagonists and mycoparasites. In The Mycota IV: Environmental and Microbial Relationships; Springer: Berlin/Heidelberg, Germany, 1997; pp. 165–184. [Google Scholar]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Chet, I.; Benhamou, N.; Haran, S. Trichoderma and Gliocladium. In Mycoparasitism and Lytic Enzymes; Taylor and Francis: London, UK, 1998; pp. 153–172. [Google Scholar]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Mendoza-Mendoza, A.; Zeilinger, S.; Horwitz, B.A. Mycoparasitism as a mechanism of Trichoderma-mediated suppression of plant diseases. Fungal Biol. Rev. 2022, 39, 15–33. [Google Scholar] [CrossRef]
- Barak, R.; Chet, I. Determination, by fluorescein diacetate staining, of fungal viability during mycoparasitism. Soil Biol. Biochem. 1986, 18, 315–319. [Google Scholar] [CrossRef]
- Elad, Y.; Chet, I. Possible role of competition for nutrients in biocontrol of Pythium damping-off by bacteria. Phytopathology 1987, 77, 190–195. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc. 1971, 57, 25–39. [Google Scholar] [CrossRef]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef]
- Hjeljord, L.; Tronsmo, A.; Harman, G.; Kubicek, C. Trichoderma and gliocladium. Biological control: An overview. In Trichoderma & Gliocladium: Enzymes, Biological Control and Commercial Applications; Harman, G.E., Kubice, C.P., Eds.; CRC Press: Boca Raton, FL, USA, 1998; Volume 2, pp. 131–151. [Google Scholar]
- Brunner, K.; Peterbauer, C.K.; Mach, R.L.; Lorito, M.; Zeilinger, S.; Kubicek, C.P. The Nag1 N-acetylglucosaminidase of Trichoderma atroviride is essential for chitinase induction by chitin and of major relevance to biocontrol. Curr. Genet. 2003, 43, 289–295. [Google Scholar]
- Zeilinger, S.; Galhaup, C.; Payer, K.; Woo, S.L.; Mach, R.L.; Fekete, C.; Lorito, M.; Kubicek, C.P. Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol. 1999, 26, 131–140. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Buensanteai, N.; Moran-Diez, M.E.; Druzhinina, I.S.; Kenerley, C.M. Functional analysis of non-ribosomal peptide synthetases (NRPSs) in Trichoderma virens reveals a polyketide synthase (PKS)/NRPS hybrid enzyme involved in the induced systemic resistance response in maize. Microbiology 2012, 158, 155–165. [Google Scholar] [CrossRef]
- Gajera, H.; Domadiya, R.; Patel, S.; Kapopara, M.; Golakiya, B. Molecular mechanism of Trichoderma as bio-control agents against phytopathogen system–a review. Curr. Res. Microbiol. Biotechnol. 2013, 1, 133–142. [Google Scholar]
- Masi, M.; Nocera, P.; Reveglia, P.; Cimmino, A.; Evidente, A. Fungal metabolites antagonists towards plant pests and human pathogens: Structure-activity relationship studies. Molecules 2018, 23, 834. [Google Scholar] [CrossRef] [PubMed]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Hu, M.; Li, Q.-L.; Yang, Y.-B.; Liu, K.; Miao, C.-P.; Zhao, L.-X.; Ding, Z.-T. Koninginins RS from the endophytic fungus Trichoderma koningiopsis. Nat. Prod. Res. 2017, 31, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Ramachander Turaga, V. Peptaibols: Antimicrobial peptides from fungi. In Bioactive Natural Products in Drug Discovery; Springer: Singapore, 2020; pp. 713–730. [Google Scholar]
- Dunlop, R.W.; Simon, A.; Sivasithamparam, K.; Ghisalberti, E.L. An Antibiotic from Trichoderma Koningii Active Against Soilborne Plant Pathogens. J. Nat. Prod. 1989, 52, 67–74. [Google Scholar] [CrossRef]
- Singh, S.; Dureja, P.; Tanwar, R.; Singh, A. Production and antifungal activity of secondary metabolites of Trichoderma virens. Pestic. Res. J. 2005, 17, 26–29. [Google Scholar]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Napolitano, M.; Lombardi, N.; Lorito, M.; Woo, S.L. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef] [PubMed]
- Ragab, M.M.; Abada, K.; Abd-El-Moneim, M.L.; Abo-Shosha, Y.Z. Effect of different mixtures of some bioagents and Rhizobium phaseoli on bean damping-off under field condition. Int. J. Sci. Eng. Res. 2015, 6, 1009–1106. [Google Scholar]
- Chen, J.-L.; Sun, S.-Z.; Miao, C.-P.; Wu, K.; Chen, Y.-W.; Xu, L.-H.; Guan, H.-L.; Zhao, L.-X. Endophytic Trichoderma gamsii YIM PH30019: A promising biocontrol agent with hyperosmolar, mycoparasitism, and antagonistic activities of induced volatile organic compounds on root-rot pathogenic fungi of Panax notoginseng. J. Ginseng Res. 2016, 40, 315–324. [Google Scholar] [CrossRef]
- De Sousa Rocha, J.d.R.; de Oliveira, N.T. In vitro antagonistic potential of Trichoderma spp. against Colletotrichum gloeosporoides agent of anthracnose in the passion fruit (passiflora). Boletín Micológico 1998, 13, 103–110. [Google Scholar] [CrossRef]
- Mahmood, A.; Kataoka, R. Potential of biopriming in enhancing crop productivity and stress tolerance. In Advances in Seed Priming; Springer: Berlin/Heidelberg, Germany, 2018; pp. 127–145. [Google Scholar]
- Da Costa, A.C.; de Miranda, R.F.; Costa, F.A.; Ulhoa, C.J. Potential of Trichoderma piluliferum as a biocontrol agent of Colletotrichum musae in banana fruits. Biocatal. Agric. Biotechnol. 2021, 34, 102028. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [PubMed]
- Jaroszuk-Ściseł, J.; Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyśkiewicz, K.; Pawlik, A.; Janusz, G. Phytohormones (auxin, gibberellin) and ACC deaminase in vitro synthesized by the mycoparasitic Trichoderma DEMTkZ3A0 strain and changes in the level of auxin and plant resistance markers in wheat seedlings inoculated with this strain conidia. Int. J. Mol. Sci. 2019, 20, 4923. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Ahluwalia, V.; Kumar, J.; Rana, V.S.; Sati, O.P.; Walia, S. Comparative evaluation of two Trichoderma harzianum strains for major secondary metabolite production and antifungal activity. Nat. Prod. Res. 2015, 29, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, M.; Omidvari, M.; Abbaszadeh-Dahaji, P.; Omidvar, R.; Kariman, K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Oskiera, M.; Szczech, M.; Bartoszewski, G. Molecular identification of Trichoderma strains collected to develop plant growth-promoting and biocontrol agents. J. Hortic. Res. 2015, 23, 75–86. [Google Scholar] [CrossRef]
- Saber, W.I.; Ghoneem, K.M.; Rashad, Y.M.; Al-Askar, A.A. Trichoderma harzianum WKY1: An indole acetic acid producer for growth improvement and anthracnose disease control in sorghum. Biocontrol Sci. Technol. 2017, 27, 654–676. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A.; Tyśkiewicz, R.; Jaroszuk-Ściseł, J. Endophytic bacteria potentially promote plant growth by synthesizing different metabolites and their phenotypic/physiological profiles in the Biolog GEN III MicroPlateTM Test. Int. J. Mol. Sci. 2019, 20, 5283. [Google Scholar] [CrossRef]
- Srinivasan, U.; Highley, T.; Bruce, A. In The role of siderophore production in the biological control of wood decay fungi by Trichoderma spp. In Proceedings of the 9th International Biodeterioration and Biodegradation Symposium; Institution of Chemical Engineers: Rugby, UK, 1995; pp. 226–231. [Google Scholar]
- Mokhtar, H.; Aid, D. Contribution in isolation and identification of some pathogenic fungi from wheat seeds, and evaluation of antagonistic capability of Trichoderma harzianum against those isolated fungi in vitro. Agric. Biol. J. N. Am. 2013, 4, 145–154. [Google Scholar] [CrossRef]
- Oszust, K.; Cybulska, J.; Frąc, M. How do Trichoderma genus fungi win a nutritional competition battle against soft fruit pathogens? A report on niche overlap nutritional potentiates. Int. J. Mol. Sci. 2020, 21, 4235. [Google Scholar] [CrossRef] [PubMed]
- Boominathan, K.; Reddy, C.; Arora, D.; Bharat, R.; Mukerji, K.; Knudsen, G. Handbook of Applied Mycology: Fungal Biotechnology 4; Marcel Dekker, Inc.: New York, NY, USA, 1992. [Google Scholar]
- Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and characterization of antibacterial siderophores secreted by endophytic fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. [Google Scholar] [CrossRef] [PubMed]
- Blaszczyk, L.; Siwulski, M.; Sobieralski, K.; Lisiecka, J.; Jedryczka, M. Trichoderma spp.—Application and prospects for use in organic farming and industry. J. Plant Prot. Res. 2014, 54, 309–317. [Google Scholar] [CrossRef]
- Vinale, F.; Flematti, G.; Sivasithamparam, K.; Lorito, M.; Marra, R.; Skelton, B.W.; Ghisalberti, E.L. Harzianic acid, an antifungal and plant growth promoting metabolite from Trichoderma harzianum. J. Nat. Prod. 2009, 72, 2032–2035. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Keswani, C.; Singh, S.P.; Sansinenea, E.; Hoat, T.X. Trichoderma spp. mediated induction of systemic defense response in brinjal against Sclerotinia sclerotiorum. Curr. Res. Microb. Sci. 2021, 2, 100051. [Google Scholar]
- Gomes, E.V.; Costa, M.d.N.; de Paula, R.G.; Ricci de Azevedo, R.; da Silva, F.L.; Noronha, E.F.; José Ulhoa, C.; Neves Monteiro, V.; Elena Cardoza, R.; Gutiérrez, S. The Cerato-Platanin protein Epl-1 from Trichoderma harzianum is involved in mycoparasitism, plant resistance induction and self cell wall protection. Sci. Rep. 2015, 5, 17998. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum interacts with roots and triggers induced systemic resistance to foliar disease in maize. Sci. Rep. 2016, 6, 35543. [Google Scholar] [CrossRef]
- Elad, Y.; Chet, I.; Henis, Y. A selective medium for improving quantitative isolation of Trichoderma spp. from soil. Phytoparasitica 1981, 9, 59–67. [Google Scholar] [CrossRef]
- Gil, S.V.; Pastor, S.; March, G. Quantitative isolation of biocontrol agents Trichoderma spp., Gliocladium spp. and actinomycetes from soil with culture media. Microbiol. Res. 2009, 164, 196–205. [Google Scholar]
- Liu, B.; Ji, S.; Zhang, H.; Wang, Y.; Liu, Z. Isolation of Trichoderma in the rhizosphere soil of Syringa oblata from Harbin and their biocontrol and growth promotion function. Microbiol. Res. 2020, 235, 126445. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Guo, R.; Ji, S.; Fan, H.; Wang, J.; Wang, Y.; Liu, Z. Isolation of Trichoderma from forestry model base and the antifungal properties of isolate TpsT17 toward Fusarium oxysporum. Microbiol. Res. 2020, 231, 126371. [Google Scholar] [CrossRef] [PubMed]
- Dou, K.; Gao, J.; Zhang, C.; Yang, H.; Jiang, X.; Li, J.; Li, Y.; Wang, W.; Xian, H.; Li, S. Trichoderma biodiversity in major ecological systems of China. J. Microbiol. 2019, 57, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, Z.; Zhang, Y.; Wang, Y.; Liu, Z. Biocontrol and growth-promoting effect of Trichoderma asperellum TaspHu1 isolate from Juglans mandshurica rhizosphere soil. Microbiol. Res. 2021, 242, 126596. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Ismaiel, A.; Bon, M.-C.; De Respinis, S.; Petrini, O. Trichoderma asperellum sensu lato consists of two cryptic species. Mycologia 2010, 102, 944–966. [Google Scholar] [CrossRef] [PubMed]
- Gezgin, Y.; Güralp, G.; Barlas, A.B.; Eltem, R. Morphological and molecular identification of trichoderma isolates used as biocontrol agents by DNA barcoding. Eur. J. Biol. 2023, 82, 59–69. [Google Scholar] [CrossRef]
- Kullnig-Gradinger, C.M.; Szakacs, G.; Kubicek, C.P. Phylogeny and evolution of the genus Trichoderma: A multigene approach. Mycol. Res. 2002, 106, 757–767. [Google Scholar] [CrossRef]
- Kopchinskiy, A.; Komoń, M.; Kubicek, C.P.; Druzhinina, I.S. TrichoBLAST: A multilocus database for Trichoderma and Hypocrea identifications. Mycol. Res. 2005, 109, 658–660. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Kopchinskiy, A.G.; Komoń, M.; Bissett, J.; Szakacs, G.; Kubicek, C.P. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea. Fungal Genet. Biol. 2005, 42, 813–828. [Google Scholar] [CrossRef]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J. Evolution of habitat preference and nutrition mode in a cosmopolitan fungal genus with evidence of interkingdom host jumps and major shifts in ecology. Evolution 2013, 67, 2823–2837. [Google Scholar] [CrossRef] [PubMed]
- Marik, T.; Urbán, P.; Tyagi, C.; Szekeres, A.; Leitgeb, B.; Vágvölgyi, M.; Manczinger, L.; Druzhinina, I.S.; Vágvölgyi, C.; Kredics, L. Diversity profile and dynamics of peptaibols produced by green mould Trichoderma species in interactions with their hosts Agaricus bisporus and Pleurotus ostreatus. Chem. Biodivers. 2017, 14, e1700033. [Google Scholar] [CrossRef]
- Samuels, G.J.; Ismaiel, A.; Mulaw, T.B.; Szakacs, G.; Druzhinina, I.S.; Kubicek, C.P.; Jaklitsch, W.M. The Longibrachiatum Clade of Trichoderma: A revision with new species. Fungal Divers. 2012, 55, 77–108. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Ashfaq, M.; Malik, A.; Khan, K.; Mathew, P. Isolation, preservation and revival of Trichoderma viride in culture media. J. Entomol. Zool. Stud. 2017, 5, 1640–1646. [Google Scholar]
- Elad, Y.; Chet, I.; Katan, J. Trichoderma harzianum: A biocontrol agent effective against Sclerotium rolfsii and Rhizoctonia solani. Phytopathology 1980, 70, 119–121. [Google Scholar] [CrossRef]
- Sikandar, S.; Ujor, V.C.; Ezeji, T.C.; Rossington, J.L.; Michel, F.C.; McMahan, C.M.; Ali, N.; Cornish, K. Thermomyces lanuginosus STm: A source of thermostable hydrolytic enzymes for novel application in extraction of high-quality natural rubber from Taraxacum kok-saghyz (Rubber dandelion). Ind. Crops Prod. 2017, 103, 161–168. [Google Scholar] [CrossRef]
- López Lastra, C.C.; Hajek, A.E.; Humber, R.A. Comparing methods of preservation for cultures of entomopathogenic fungi. Can. J. Bot. 2002, 80, 1126–1130. [Google Scholar] [CrossRef]
- Bakerspigel, A. Soil as a storage medium for fungi. Mycologia 1953, 45, 596–604. [Google Scholar] [CrossRef]
- Buell, C.B.; Weston, W.H. Application of the mineral oil conservation method to maintaining collections of fungous cultures. Am. J. Bot. 1947, 34, 555–561. [Google Scholar] [CrossRef]
- Elliott, M.L. Survival, Growth and Pathogenicity of Gaeumannomyces graminis var. graminis with Different Methods of Long-Term Storage. Mycologia 2005, 97, 901–907. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Yassin, M.A.; Moslem, M.A.; Bahkali, A.H.; de Wit, P.J.; McKenzie, E.H.; Stephenson, S.L.; Cai, L.; Hyde, K.D. Culture collections, the new herbaria for fungal pathogens. Fungal Divers. 2010, 45, 21–32. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Stoccho, M.; Mónaco, C.; Cordo, C. A comparison of preservation methods for Trichoderma harzianum cultures. Rev. Iberoam. Micol. 2010, 27, 213. [Google Scholar] [CrossRef] [PubMed]
- Messer, J.W.; Rice, E.; Johnson, C. Total viable counts. Spread plate technique. Encycl. Food Microbiol. 2000, 3, 2159–2160. [Google Scholar]
- Hoben, H.; Somasegaran, P. Comparison of the pour, spread, and drop plate methods for enumeration of Rhizobium spp. in inoculants made from presterilized peat. Appl. Environ. Microbiol. 1982, 44, 1246–1247. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Sekhar, A.C.; Upreti, R.; Mujawar, M.M.; Pasha, S.S. Optimization of single plate-serial dilution spotting (SP-SDS) with sample anchoring as an assured method for bacterial and yeast cfu enumeration and single colony isolation from diverse samples. Biotechnol. Rep. 2015, 8, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jørgensen, H.J.; Latz, M.A.; Manzotti, A.; Ntana, F.; Rojas, E.C.; Jensen, B. Searching for novel fungal biological control agents for plant disease control among endophytes. Endophytes Grow. World 2019, 31, 25. [Google Scholar]
- Vafadar, F.; Amooaghaie, R.; Otroshy, M. Effects of plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungus on plant growth, stevioside, NPK, and chlorophyll content of Stevia rebaudiana. J. Plant Interact. 2014, 9, 128–136. [Google Scholar] [CrossRef]
- Pohjanen, J.; Koskimäki, J.J.; Sutela, S.; Ardanov, P.; Suorsa, M.; Niemi, K.; Sarjala, T.; Häggman, H.; Pirttilä, A.M. Interaction with ectomycorrhizal fungi and endophytic Methylobacterium affects nutrient uptake and growth of pine seedlings in vitro. Tree Physiol. 2014, 34, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Bora, P.S.L.; Baruah, K.P.A.; Talukdar, R.B.K.; Kataky, L.; Phukan, A. Review on bacterial blight of rice caused by Xanthomonas oryzae pv. oryzae: Different management approaches and role of Pseudomonas fluorescens as a potential biocontrol agent. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 982–1005. [Google Scholar] [CrossRef]
- Gupta, R.; Saxena, R.; Goel, S. Photoinduced sporulation in Trichoderma harzianum–an experimental approach to primary events. World J. Microbiol. Biotechnol. 1997, 13, 249–250. [Google Scholar] [CrossRef]
- Van Gerrewey, T.; Ameloot, N.; Navarrete, O.; Vandecruys, M.; Perneel, M.; Boon, N.; Geelen, D. Microbial activity in peat-reduced plant growing media: Identifying influential growing medium constituents and physicochemical properties using fractional factorial design of experiments. J. Clean. Prod. 2020, 256, 120323. [Google Scholar] [CrossRef]
- Dattatraya Saratale, G.; Bhosale, R.; Shobana, S.; Banu, J.R.; Pugazhendhi, A.; Mahmoud, E.; Sirohi, R.; Kant Bhatia, S.; Atabani, A.E.; Mulone, V.; et al. A review on valorization of spent coffee grounds (SCG) towards biopolymers and biocatalysts production. Bioresour. Technol. 2020, 314, 123800. [Google Scholar] [CrossRef] [PubMed]
- Rukmani, S.; Mariappan, V. Influence of organic amendments with Trichoderma viride on the control of root rot of blackgram. Plant Dis. Res. 1990, 5, 244. [Google Scholar]
- Sawant, I.S.; Sawant, S. A simple method for achieving high cfu of Trichoderma harzianum on organic wastes for field applications. Indian Phytopathol. 1996, 49, 185–187. [Google Scholar]
- Saju, K.; Anandaraj, M.; Sarma, Y. On farm production of Trichoderma harzianum using organic matter. Indian Phytopathol. 2002, 55, 277–281. [Google Scholar]
- Sangle, U.; Bambawale, O. Evaluation of substrates for mass multiplication of Trichoderma spp. Indian J. Plant Prot. 2005, 33, 298. [Google Scholar]
- Simon, S. Agro-based waste products as a substrate for mass production of Trichoderma spp. J. Agric. Sci. 2011, 3, 168. [Google Scholar]
- Babu, K.N.; Pallavi, P. Isolation, identification and mass multiplication of Trichoderma an important bio-control agent. Int. J. Pharm. Life Sci. 2013, 4, 2320–2323. [Google Scholar]
- Mustafa, A.; Khan, M.A.; Inam-ul-Haq, M.; Khan, S.; Pervez, M.A. Mass multiplication of Trichoderma spp. on organic substrate and their effect in management of seed borne fungi. Pak. J. Phytopathol. 2009, 21, 108–114. [Google Scholar]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.; Hernandez, J.-P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Jeyarajan, R. Prospects of Indigenous Mass Production and Formulation of Trichoderma, Current status of biological control of plant diseases using antagonistic organisms in India. In Proceedings of the Group Meeting on Antagonistic Organisms in Plant Disease Management Held at Project Directorate of Biological Control, Bangalore, India, 10–11 July 2003; Project Directorate of Biological Control, Indian Council of Agricultural Research: Bangalore, India, 2006; pp. 74–80. [Google Scholar]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Srinivas, V. Formulations of plant growth-promoting microbes for field applications. Microb. Inoculants Sustain. Agric. Product. Funct. Appl. 2016, 2, 239–251. [Google Scholar]
- Nakkeeran, S.; Fernando, W.D.; Siddiqui, Z.A. Plant growth promoting rhizobacteria formulations and its scope in commercialization for the management of pests and diseases. In PGPR: Biocontrol and Biofertilization; Springer: Dordrecht, The Netherlands, 2006; pp. 257–296. [Google Scholar]
- Keswani, C.; Bisen, K.; Singh, V.; Sarma, B.K.; Singh, H.B. Formulation technology of biocontrol agents: Present status and future prospects. In Bioformulations: For Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 35–52. [Google Scholar]
- Bejarano, A.; Puopolo, G. Bioformulation of microbial biocontrol agents for a sustainable agriculture. In How Research Can Stimulate the Development of Commercial Biological Control against Plant Diseases; Springer: Berlin/Heidelberg, Germany, 2020; pp. 275–293. [Google Scholar]
- Anith, K.; Vaishakhi, A.; Viswanathan, A.; Varkey, S.; Aswini, S. Population dynamics and efficiency of coconut water based liquid formulation of Pseudomonas fluorescens AMB-8. J. Trop. Agric. 2017, 54, 184. [Google Scholar]
- Mbarga, J.B.; Begoude, B.; Ambang, Z.; Meboma, M.; Kuaté, J.; Schiffers, B.; Ewbank, W.; Dedieu, L.; Ten Hoopen, G.M. A new oil-based formulation of Trichoderma asperellum for the biological control of cacao black pod disease caused by Phytophthora megakarya. Biol. Control 2014, 77, 15–22. [Google Scholar] [CrossRef]
- Peeran, M.F.; Krishnan, N.; Thangamani, P.R.; Gandhi, K.; Thiruvengadam, R.; Kuppusamy, P. Development and evaluation of water-in-oil formulation of Pseudomonas fluorescens (FP7) against Colletotrichum musae incitant of anthracnose disease in banana. Eur. J. Plant Pathol. 2014, 138, 167–180. [Google Scholar] [CrossRef]
- Herrera, W.; Valbuena, O.; Pavone-Maniscalco, D. Formulation of Trichoderma asperellum TV190 for biological control of Rhizoctonia solani on corn seedlings. Egypt J. Biol. Pest Control 2020, 30, 1–8. [Google Scholar] [CrossRef]
- Patil, S.S.; Guldegar, D.D.; Potdukhe, S.R.; Khobragade, H.M. Investigation of Liquid Formulation of Trichoderma asperellum against Fusarium Wilt of Chickpea. Biol. Forum Int. J. 2021, 13, 571–576. [Google Scholar]
- Whipps, J.M. Developments in the biological control of soil-borne plant pathogens. Adv. Bot. Res. 1997, 26, 1–134. [Google Scholar]
- Jin, X.; Hayes, C.K.; Harman, G.E. Principles in the development of biological control systems employing Trichoderma species against soil-borne plant pathogenic fungi. In Frontiers in Industrial Mycology; Springer: Berlin/Heidelberg, Germany, 1992; pp. 174–195. [Google Scholar]
- Morgan, C.A.; Herman, N.; White, P.; Vesey, G. Preservation of micro-organisms by drying; a review. J. Microbiol. Methods 2006, 66, 183–193. [Google Scholar] [CrossRef]
- Cumagun, C.J.R. Advances in formulation of Trichoderma for biocontrol. In Biotechnology and Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; pp. 527–531. [Google Scholar]
- Jeyarajan, R.; Ramakrishnan, G.; Dinakaran, D.; Sridar, R. Development of products Trichoderma viride and Bacillus subtilis for biocontrol of root rot diseases. In Biotechnology in India; Dwivedi, B.K., Ed.; Bioved Research Society: Allahabad, India, 1994; pp. 25–36. [Google Scholar]
- Pradhan, P.C.; Mukhopadhyay, A.; Kumar, R.; Kundu, A.; Patanjali, N.; Dutta, A.; Kamil, D.; Bag, T.K.; Aggarwal, R.; Bharadwaj, C. Performance appraisal of Trichoderma viride based novel tablet and powder formulations for management of Fusarium wilt disease in chickpea. Front. Plant Sci. 2022, 13, 990392. [Google Scholar] [CrossRef] [PubMed]
- Sundaramoorthy, S.; Balabaskar, P. Biocontrol efficacy of Trichoderma spp. against wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. J. Appl. Biol. Biotechnol. 2013, 1, 036–040. [Google Scholar]
- Lewis, J. Formulation and delivery systems of biocontrol agents with emphasis on fungi. In The Rhizosphere and Plant Growth, Proceedings of the Beltsville Symposia in Agricultural Research, Beltsville, MD, USA, 8–11 May 1989; Springer: Berlin/Heidelberg, Germany, 1991; pp. 279–287. [Google Scholar]
- Batta, Y. Control of postharvest diseases of fruit with an invert emulsion formulation of Trichoderma harzianum Rifai. Postharvest Biol. Technol. 2007, 43, 143–150. [Google Scholar] [CrossRef]
- Bhai, R. Preservation and long-term storage of Trichoderma spp. by sodium alginate encapsulation. J. Plant Crops 2020, 48, 36–44. [Google Scholar]
- Byakodi, A.; Babu, B.S. Experimental studies on effects of selected species of fungi on aerobic composting of sugar pressmud. Environ. Res. Eng. Manag. 2021, 77, 74–85. [Google Scholar] [CrossRef]
- Cavalcante, R.S.; Lima, H.L.S.; Pinto, G.A.S.; Gava, C.A.T.; Rodrigues, S. Effect of Moisture on Trichoderma Conidia Production on Corn and Wheat Bran by Solid State Fermentation. Food Bioproc. Tech. 2008, 1, 100–104. [Google Scholar] [CrossRef]
- Balasubramanian, C.; Udaysoorian, P.; Prabhu, C.; Kumar, G. Enriched compost for yield and quality enhancement in sugarcane. J. Ecobiol. 2008, 22, 173–176. [Google Scholar]
- Knowles, A. Recent developments of safer formulations of agrochemicals. Environmentalist 2008, 28, 35–44. [Google Scholar] [CrossRef]
- Jin, X.; Custis, D. Microencapsulating aerial conidia of Trichoderma harzianum through spray drying at elevated temperatures. Biol. Control 2011, 56, 202–208. [Google Scholar] [CrossRef]
- Dos Santos, G.; Locatelli, G.; Coêlho, D.; Botelho, P.; De Amorim, M.; De Vasconcelos, T.; Bueno, L. Factorial design, preparation and characterization of new beads formed from alginate, polyphosphate and glycerol gelling solution for microorganism microencapsulation. J. Solgel Sci. Technol. 2015, 75, 345–352. [Google Scholar] [CrossRef]
- Oancea, F.; Raut, I.; Şesan, T.E.; Cornea, P.C. Dry Flowable Formulation of Biostimulants Trichoderma Strains. Agric. Agric. Sci. Procedia 2016, 10, 494–502. [Google Scholar] [CrossRef]
- Arora, N.K.; Kumar, V.; Maheshwari, D.K. Constraints, development and future of the inoculants with special reference to rhizobial inoculants. In Innovative Approaches in Microbiology; Maheshwari, D.K., Dubey, R.C., Eds.; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2001; pp. 241–245. [Google Scholar]
- Arora, N.; Khare, E.; Naraian, R.; Maheshwari, D. Sawdust as a superior carrier for production of multipurpose bioinoculant using plant growth promoting rhizobial and pseudomonad strains and their impact on productivity of Trifolium repense. Curr. Sci. 2008, 95, 90–94. [Google Scholar]
- Moses, K. Assessment of sawdust as carrier material for fungal inoculum intended for faster composting. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 608–613. [Google Scholar]
- Khan, M.R.; Majid, S.; Mohidin, F.A.; Khan, N. A new bioprocess to produce low cost powder formulations of biocontrol bacteria and fungi to control fusarial wilt and root-knot nematode of pulses. Biol. Control 2011, 59, 130–140. [Google Scholar] [CrossRef]
- Nabi, S.U.; Raja, W.H.; Sharma, A.; Malik, G. Evaluation of Different Substrates for Development of Trichoderma harzianum Based Stock Cultures and their Utilization in Management of Chilli Wilt Disease. Chem. Sci. Rev. Lett. 2017, 6, 2229–2235. [Google Scholar]
- Mohiddin, F.; Bashir, I.; Padder, S.A.; Hamid, B. Evaluation of different substrates for mass multiplication of Trichoderma species. J. Pharmacogn. Phytochem. 2017, 6, 563–569. [Google Scholar]
- Ananda, N.; Vadlani, P.V.; Prasad, P.V.V. Evaluation of drought and heat stressed grain sorghum (Sorghum bicolor) for ethanol production. Ind. Crops Prod. 2011, 33, 779–782. [Google Scholar] [CrossRef]
- Chammem, H.; Nesler, A.; Pertot, I. Wood pellets as carriers of conidia of Trichoderma atroviride SC1 for soil application. Fungal Biol. 2021, 125, 989–998. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Performance of a Trichoderma harzianum bentonite–vermiculite formulation against Fusarium wilt in seedling nursery melon plants. HortScience 2009, 44, 2025–2027. [Google Scholar] [CrossRef]
- Sreenayana, B.; Nakkeeran, S. Effect of Oil Based Formulation of Trichoderma spp. on Growth Parameters of Cucumber Seedlings. Int. J. Curr. Microbiol. App. Sci 2019, 8, 200–209. [Google Scholar] [CrossRef]
- Connick, W.J.; Boyette, C.D.; McAlpine, J.R. Formulation of mycoherbicides using a pasta-like process. Biol. Control 1991, 1, 281–287. [Google Scholar] [CrossRef]
- Thangavelu, R.; Palaniswami, A.; Velazhahan, R. Mass production of Trichoderma harzianum for managing fusarium wilt of banana. Agric. Ecosyst. Environ. 2004, 103, 259–263. [Google Scholar] [CrossRef]
- Ramanujam, B.; Prasad, R.; Sriram, S.; Rangeswaran, R. Mass production, formulation, quality control and delivery of Trichoderma for plant disease management. J. Plant Prot. Sci. 2010, 2, 1–8. [Google Scholar]
- Komala, G.; Madhavi, G.B.; Nath, R. Shelf life studies of different formulations of Trichoderma harzianum. Plant Cell Biotechnol. Mol. Biol. 2019, 20, 1100–1105. [Google Scholar]
- Organo, N.D.; Granada, S.M.J.M.; Pineda, H.G.S.; Sandro, J.M.; Nguyen, V.H.; Gummert, M. Assessing the potential of a Trichoderma-based compost activator to hasten the decomposition of incorporated rice straw. Sci. Rep. 2022, 12, 448. [Google Scholar] [CrossRef] [PubMed]
- Marcelo, L.R.; de Gois, J.S.; da Silva, A.A.; Cesar, D.V. Synthesis of iron-based magnetic nanocomposites and applications in adsorption processes for water treatment: A review. Environ. Chem. Lett. 2021, 19, 1229–1274. [Google Scholar] [CrossRef]
- Prasad, R.D.; Poorna Chandrika, K.S.; Desai, S.; Greeshma, K.; Vijaykumar, S. Development, Production, and Storage of Trichoderma Formulations for Agricultural Applications. In Advances in Trichoderma Biology for Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 371–385. [Google Scholar]
- Rini, C.; Ramya, J.; Jayakumar, G.; Shajan, V. Low cost carrier material for mass production of Trichoderma inoculants. J. Trop. Agric. 2018, 56, 63–68. [Google Scholar]
- Rai, D.; Tewari, A. Shelf life studies of different formulations based on Trichoderma harzianum (Th14). Ann. Biol. Res. 2016, 7, 1–5. [Google Scholar]
- Patil, P.D.; Patil, S.P.; Kelkar, R.K.; Patil, N.P.; Pise, P.V.; Nadar, S.S. Enzyme-assisted supercritical fluid extraction: An integral approach to extract bioactive compounds. Trends Food Sci. Technol. 2021, 116, 357–369. [Google Scholar] [CrossRef]
- Sriram, S.; Roopa, K.P.; Savitha, M.J. Extended shelf-life of liquid fermentation derived talc formulations of Trichoderma harzianum with the addition of glycerol in the production medium. Crop Prot. 2011, 30, 1334–1339. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Parthasarathy, S.; Kamalakannan, A.; Kuppusamy, K.; Gopalakrishnan, C. A novel method to develop paste formulation of Trichoderma harzianum. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3286–3293. [Google Scholar]
- Jeyarajan, R.; Nakkeeran, S. Exploitation of microorganisms and viruses as biocontrol agents for crop disease management. In Biocontrol Potential and its Exploitation in Sustainable Agriculture: Crop Diseases, Weeds, and Nematodes; Springer: Berlin/Heidelberg, Germany, 2000; pp. 95–116. [Google Scholar]
- O’Callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef] [PubMed]
- Rocha, I.; Ma, Y.; Souza-Alonso, P.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed coating: A tool for delivering beneficial microbes to agricultural crops. Front. Plant Sci. 2019, 10, 1357. [Google Scholar] [CrossRef] [PubMed]
- Rocha, I.; Ma, Y.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Growth and nutrition of cowpea (Vigna unguiculata) under water deficit as influenced by microbial inoculation via seed coating. J. Agron. Crop Sci. 2019, 205, 447–459. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.N.; Shrestha, S.M.; Mukherjee, P.K. Biological seed treatment for control of soil-borne plant pathogens. FAO Plant Prot. Bull. 1992, 40, 21–30. [Google Scholar]
- Jogani, V.; John, P. Evaluation of different application methods of Trichoderma harzianum (Rifai) against Fusarium wilt of tomato. Crop Res. 2014, 48, 76–79. [Google Scholar]
- Tewari, A.; Mukhopadhyay, A. Testing of different formulations of Gliocladium virens against chickpea wilt-complex. Indian Phytopathol. 2001, 54, 67–71. [Google Scholar]
- Paravar, A.; Piri, R.; Balouchi, H.; Ma, Y. Microbial seed coating: An attractive tool for sustainable agriculture. Biotechnol. Rep. 2023, 37, e00781. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Corrales, D.C.; Soltani, E. Biological seed treatments promote crop establishment and yield: A global meta-analysis. Agron. Sustain. Dev. 2022, 42, 45. [Google Scholar] [CrossRef]
- Couto, A.P.S.; Pereira, A.E.; Abati, J.; Fontanela, M.L.C.; Dias-Arieira, C.R.; Krohn, N.G. Seed treatment with Trichoderma and chemicals to improve physiological and sanitary quality of wheat cultivars. Rev. Caatinga 2021, 34, 813–823. [Google Scholar] [CrossRef]
- Sudana, I.M.; Raka, I.G.N. Seeds Treatment Using Trichoderma spp. Formulated in Biosluryy and Vermicompost to Induct the Resistance of the Peanut (Arachis hypogaea L.) Diseases. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 1253–1265. [Google Scholar] [CrossRef]
- Hossain, S.; Akter, F. Effects of Trichoderma-enriched biofertilizer and farmyard manure on the growth and yield of brinjal (Solanum melongena L.). Dhaka Univ. J. Biol. Sci. 2020, 29, 1–8. [Google Scholar] [CrossRef]
- Kim, K.; Heo, Y.M.; Jang, S.; Lee, H.; Kwon, S.-L.; Park, M.S.; Lim, Y.W.; Kim, J.-J. Diversity of Trichoderma spp. in marine environments and their biological potential for sustainable industrial applications. Sustainability 2020, 12, 4327. [Google Scholar] [CrossRef]
- Sarkar, D.; Singh, S.; Parihar, M.; Rakshit, A. Seed bio-priming with microbial inoculants: A tailored approach towards improved crop performance, nutritional security, and agricultural sustainability for smallholder farmers. Curr. Res. Environ. Sustain. 2021, 3, 100093. [Google Scholar] [CrossRef]
- Swain, H.; Adak, T.; Mukherjee, A.K.; Mukherjee, P.K.; Bhattacharyya, P.; Behera, S.; Bagchi, T.B.; Patro, R.; Khandual, A.; Bag, M. Novel Trichoderma strains isolated from tree barks as potential biocontrol agents and biofertilizers for direct seeded rice. Microbiol. Res. 2018, 214, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Rajput, R.S.; Singh, P.; Singh, J.; Vaishnav, A.; Ray, S.; Singh, H. Trichoderma mediated seed biopriming augments antioxidant and phenylpropanoid activities in tomato plant against Sclerotium rolfsii. J. Pharmacogn. Phytochem. 2019, 8, 2641–2647. [Google Scholar]
- Singh, V.; Upadhyay, R.S.; Sarma, B.; Singh, H. Seed bio-priming with Trichoderma asperellum effectively modulate plant growth promotion in pea. Int. J. Agric. Environ. Biotechnol. 2016, 9, 361–365. [Google Scholar] [CrossRef]
- Swain, H.; Adak, T.; Mukherjee, A.K.; Sarangi, S.; Samal, P.; Khandual, A.; Jena, R.; Bhattacharyya, P.; Naik, S.K.; Mehetre, S.T. Seed biopriming with Trichoderma strains isolated from tree bark improves plant growth, antioxidative defense system in rice and enhance straw degradation capacity. Front. Microbiol. 2021, 12, 633881. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S. Trichoderma biological control to protect sensitive maize hybrids against late wilt disease in the field. J. Fungi 2021, 7, 315. [Google Scholar] [CrossRef]
- Jambhulkar, P.P.; Raja, M.; Singh, B.; Katoch, S.; Kumar, S.; Sharma, P. Potential native Trichoderma strains against Fusarium verticillioides causing post flowering stalk rot in winter maize. Crop Prot. 2022, 152, 105838. [Google Scholar] [CrossRef]
- Singh, U.S.; Zaidi, N.W. Current status of formulation and delivery of fungal and bacterial antagonists for disease management in India. In Microbial Biopesticide Formulations and Application; Project Directorate of Biological Control: Bangalore, India, 2002; p. 269. [Google Scholar]
- Yang, Z.; Zhi, P.; Chang, C. Priming seeds for the future: Plant immune memory and application in crop protection. Front. Plant Sci. 2022, 13, 961840. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma as biological control agent: Scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef]
- Panwar, V.; Aggarwal, A.; Singh, G.; Verma, A.; Sharma, I.; Saharan, M. Efficacy of foliar spray of Trichoderma isolates against Fusarium graminearum causing head blight of wheat. J. Wheat Res. 2014, 6, 59–63. [Google Scholar]
- Karthikeyan, M.; Radhika, K.; Bhaskaran, R.; Mathiyazhagan, S.; Sandosskumar, R.; Velazhahan, R.; Alice, D. Biological control of onion leaf blight disease by bulb and foliar application of powder formulation of antagonist mixture. Arch. Phytopathol. Plant Prot. 2008, 41, 407–417. [Google Scholar] [CrossRef]
- Gade, R.; Wardhe, S.; Armarker, S. Shelf life study of Trichoderma spp. in different carrier materials. J. Maharashtra Agric. Univ. 2009, 34, 181–182. [Google Scholar]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Şesan, T.E.; Oancea, A.O.; Ştefan, L.M.; Mănoiu, V.S.; Ghiurea, M.; Răut, I.; Constantinescu-Aruxandei, D.; Toma, A.; Savin, S.; Bira, A.F.; et al. Effects of Foliar Treatment with a Trichoderma Plant Biostimulant Consortium on Passiflora caerulea L. Yield and Quality. Microorganisms 2020, 8, 123. [Google Scholar] [CrossRef]
- Uddin, A.; Hussain, M.; Rahman, S.; Ahmad, H.; Roni, M. Effect of Trichoderma concentrations on growth and yield of tomato. Bangladesh Res. Publ. J. 2015, 11, 228–232. [Google Scholar]
- Dursun, A.; Ekinci, M.; Dönmez, M.F. Effects of foliar application of plant growth promoting bacterium on chemical contents, yield and growth of tomato (Lycopersicon esculentum L.) and cucumber (Cucumis sativus L.). Pak. J. Bot. 2010, 42, 3349–3356. [Google Scholar]
- Syam, N.; Hidrawati; Sabahannur, S.; Nurdin, A. Effects of Trichoderma and Foliar Fertilizer on the Vegetative Growth of Black Pepper (Piper nigrum L.) Seedlings. Int. J. Agron. 2021, 2021, 9953239. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 2023, 14, 1160551. [Google Scholar] [CrossRef] [PubMed]
- Sehim, A.E.; Hewedy, O.A.; Altammar, K.A.; Alhumaidi, M.S.; Abd Elghaffar, R.Y. Trichoderma asperellum empowers tomato plants and suppresses Fusarium oxysporum through priming responses. Front. Microbiol. 2023, 14, 1140378. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.; Kay, W.; Kilaru, S.; Schuster, M.; Gurr, S.J.; Steinberg, G. Multi-site fungicides suppress banana Panama disease, caused by Fusarium oxysporum f. sp. cubense Tropical Race 4. PLoS Pathog. 2022, 18, e1010860. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Liu, Q.; Yang, Z. Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. J. Fungi 2023, 9, 843. [Google Scholar] [CrossRef] [PubMed]
- Jeyalakshmi, C.; Rettinassababady, C.; Nema, S. Integrated management of sesame diseases. J. Biopestic. 2013, 6, 68. [Google Scholar]
- El-Mohamedy, R.S.; Ziedan, E.; Abdalla, A. Biological soil treatment with Trichoderma harzianum to control root rot disease of grapevine (Vitis vinifera L.) in newly reclaimed lands in Nobaria province. Arch. Phytopathol. Plant Prot. 2010, 43, 73–87. [Google Scholar] [CrossRef]
- Liu, L.; Xu, Y.; Cao, H.; Fan, Y.; Du, K.; Bu, X.; Gao, D. Effects of Trichoderma harzianum biofertilizer on growth, yield, and quality of Bupleurum chinense. Plant Direct 2022, 6, e461. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhao, Q.; Li, W.; Gao, L.; Liu, G. Strain improvement of Trichoderma harzianum for enhanced biocontrol capacity: Strategies and prospects. Front. Microbiol. 2023, 14, 1146210. [Google Scholar] [CrossRef]
- Wang, R.; Liu, C.; Jiang, X.; Tan, Z.; Li, H.; Xu, S.; Zhang, S.; Shang, Q.; Deising, H.B.; Behrens, S.E.; et al. The Newly Identified Trichoderma harzianum Partitivirus (ThPV2) Does Not Diminish Spore Production and Biocontrol Activity of Its Host. Viruses 2022, 14, 1532. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Kalia, A.; Lore, J.S.; Kaur, A.; Yadav, I.; Sharma, P.; Sandhu, J.S. Trichoderma sp. endochitinase and β-1,3-glucanase impede Rhizoctonia solani growth independently, and their combined use does not enhance impediment. Plant Pathol. 2021, 70, 1388–1396. [Google Scholar] [CrossRef]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Kumhar, K.C.; Kumar, K.; Arora, I.; Bhatia, A.K.; Batra, V.K.; Raj, H. Management of tomato damping-off disease in the nursery using of Trichoderma asperellum. Int. J. Econ. Plants 2022, 9, 145–151. [Google Scholar] [CrossRef]
- Yu, C.; Jiang, X.; Xu, H.; Ding, G. Trichoderma longibrachiatum Inoculation Improves Drought Resistance and Growth of Pinus massoniana Seedlings through Regulating Physiological Responses and Soil Microbial Community. J. Fungi 2023, 9, 694. [Google Scholar] [CrossRef]
- Pascual, J.A.; Ceglie, F.; Tuzel, Y.; Koller, M.; Koren, A.; Hitchings, R.; Tittarelli, F. Organic substrate for transplant production in organic nurseries. A review. Agron. Sustain. Dev. 2018, 38, 1–23. [Google Scholar] [CrossRef]
- Van Jaarsveld, W.J.; Halleen, F.; Bester, M.C.; Pierron, R.J.; Stempien, E.; Mostert, L. Investigation of Trichoderma species colonization of nursery grapevines for improved management of black foot disease. Pest Manag. Sci. 2021, 77, 397–405. [Google Scholar] [CrossRef]
- Lanzuise, S.; Manganiello, G.; Guastaferro, V.M.; Vincenzo, C.; Vitaglione, P.; Ferracane, R.; Vecchi, A.; Vinale, F.; Kamau, S.; Lorito, M.; et al. Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality. Agronomy 2022, 12, 275. [Google Scholar] [CrossRef]
- Galante, Y.; De Conti, A.; Monteverdi, R. Trichoderma and Gliocladium. In Trichoderma & Gliocladium: Enzymes, Biological Control and Commercial Applications; Taylor & Francis: Abingdon, UK, 1998. [Google Scholar]
- Kaczmarek, J.; Jedryczka, M. Characterization of two coexisting pathogen populations of Leptosphaeria spp., the cause of stem canker of brassicas. Acta Agrobot. 2011, 64, 2. [Google Scholar] [CrossRef]
- Zafra, G.; Moreno-Montaño, A.; Absalón, Á.E.; Cortés-Espinosa, D.V. Degradation of polycyclic aromatic hydrocarbons in soil by a tolerant strain of Trichoderma asperellum. Environ. Sci. Pollut. Res. 2015, 22, 1034–1042. [Google Scholar] [CrossRef]
- Vázquez, M.B.; Barrera, V.; Bianchinotti, V. Molecular identification of three isolates of Trichoderma harzianum isolated from agricultural soils in Argentina, and their abilities to detoxify in vitro metsulfuron methyl. Botany 2015, 93, 793–800. [Google Scholar] [CrossRef]
- Amira, R.D.; Roshanida, A.; Rosli, M.; Zahrah, M.S.F.; Anuar, J.M.; Adha, C.N. Bioconversion of empty fruit bunches (EFB) and palm oil mill effluent (POME) into compost using Trichoderma virens. Afr. J. Biotechnol. 2011, 10, 18775–18780. [Google Scholar]
- Haddadin, M.S.; Haddadin, J.; Arabiyat, O.I.; Hattar, B. Biological conversion of olive pomace into compost by using Trichoderma harzianum and Phanerochaete chrysosporium. Bioresour. Technol. 2009, 100, 4773–4782. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Singh, R.P. Current challenges, constraints and future strategies for development of successful market for biopesticides. Clim. Chang. Environ. Sustain. 2016, 4, 129–136. [Google Scholar] [CrossRef]
- Mishra, J.; Tewari, S.; Singh, S. Biopesticides: Where we stand? In Plant Microbes Symbiosis; Arora, N.K., Ed.; Springer: New Delhi, India, 2015. [Google Scholar]
- Chakraborty, N.; Mitra, R.; Pal, S.; Ganguly, R.; Acharya, K.; Minkina, T.; Sarkar, A.; Keswani, C. Biopesticide Consumption in India: Insights into the Current Trends. Agriculture 2023, 13, 557. [Google Scholar] [CrossRef]
- Topolovec-Pintarić, S. Trichoderma: Invisible partner for visible impact on agriculture. In Trichoderma—The Most Widely used Fungicide; IntechOpen: London, UK, 2019; p. 15. [Google Scholar]
- Kumar, S.; Thakur, M.; Rani, A. Trichoderma: Mass production, formulation, quality control, delivery and its scope in commercialization in India for the management of plant diseases. Afr. J. Agric. Res. 2014, 9, 3838–3852. [Google Scholar]
- Köberl, M.; Ramadan, E.M.; Roßmann, B.; Staver, C.; Fürnkranz, M.; Lukesch, B.; Grube, M.; Berg, G. Using ecological knowledge and molecular tools to develop effective and safe biocontrol strategies. In Pesticides in the Modern World—Pests Control and Pesticides Exposure and Toxicity Assessment; IntechOpen: London, UK, 2011; pp. 3–26. [Google Scholar]
- Oros, G.; Naár, Z.; Cserháti, T. Growth response of Trichoderma species to organic solvents. Mol. Inform. 2011, 30, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Hajieghrari, B. Effect of some metal-containing compounds and fertilizers on mycoparasite Trichoderma species mycelia growth response. Afr. J. Biotechnol. 2010, 9, 4025–4033. [Google Scholar]
- Niti, C.; Sunita, S.; Kamlesh, K.; Rakesh, K. Bioremediation: An emerging technology for remediation of pesticides. Res. J. Chem. Environ. 2013, 17, 4. [Google Scholar]
- Argumedo-Delira, R.; Alarcón, A.; Ferrera-Cerrato, R.; Almaraz, J.J.; Peña-Cabriales, J.J. Tolerance and growth of 11 Trichoderma strains to crude oil, naphthalene, phenanthrene and benzo [a] pyrene. J. Environ. Manage. 2012, 95, S291–S299. [Google Scholar] [CrossRef]
- Matsubara, M.; Lynch, J.; De Leij, F. A simple screening procedure for selecting fungi with potential for use in the bioremediation of contaminated land. Enzyme Microb. Technol. 2006, 39, 1365–1372. [Google Scholar] [CrossRef]
- Ezzi, M.I.; Lynch, J.M. Biodegradation of cyanide by Trichoderma spp. and Fusarium spp. Enzyme Microb. Technol. 2005, 36, 849–854. [Google Scholar] [CrossRef]
- Cao, L.; Jiang, M.; Zeng, Z.; Du, A.; Tan, H.; Liu, Y. Trichoderma atroviride F6 improves phytoextraction efficiency of mustard (Brassica juncea (L.) Coss. var. foliosa Bailey) in Cd, Ni contaminated soils. Chemosphere 2008, 71, 1769–1773. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.; De-Leij, F.A.; Lynch, J. Trichoderma harzianum Rifai 1295-22 mediates growth promotion of crack willow (Salix fragilis) saplings in both clean and metal-contaminated soil. Microb. Ecol. 2007, 54, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M.; Moffat, A.J. Bioremediation–prospects for the future application of innovative applied biological research. Ann. Appl. Biol. 2005, 146, 217–221. [Google Scholar] [CrossRef]
- Huang, J.; Pang, Y.; Zhang, F.; Huang, Q.; Zhang, M.; Tang, S.; Fu, H.; Li, P. Suppression of Fusarium wilt of banana by combining acid soil ameliorant with biofertilizer made from Bacillus velezensis H-6. Eur. J. Plant Pathol. 2019, 154, 585–596. [Google Scholar] [CrossRef]
- Smily, J.R.M.B.; Sumithra, P.A. Optimization of chromium biosorption by fungal adsorbent, Trichoderma sp. BSCR02 and its desorption studies. HAYATI J. Biosci. 2017, 24, 65–71. [Google Scholar] [CrossRef]
- Moreno-Ruiz, D.; Lichius, A.; Turrà, D.; Di Pietro, A.; Zeilinger, S. Chemotropism assays for plant symbiosis and mycoparasitism related compound screening in Trichoderma atroviride. Front. Microbiol. 2020, 11, 601251. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Banga, S.S. Combating Climate Change: An Agricultural Perspective; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
Figure 1.
Procedure of Trichoderma formulation. (a) Rhizospheric soil. (b) Selective medium for isolation. (c) Pure culture. (d) Liquid fermentation. (e) Soil fermentation. (f) Talcum powder. (g) Mixing of Trichoderma with Talcum powder. (h) Packaging. (i) Mixing in FYM. (j) Seed priming. (k) Seedling treatment. (l) Pot application.
Figure 1.
Procedure of Trichoderma formulation. (a) Rhizospheric soil. (b) Selective medium for isolation. (c) Pure culture. (d) Liquid fermentation. (e) Soil fermentation. (f) Talcum powder. (g) Mixing of Trichoderma with Talcum powder. (h) Packaging. (i) Mixing in FYM. (j) Seed priming. (k) Seedling treatment. (l) Pot application.

Figure 2.
Pictorial representation of factors that encourage a market for the Trichoderma bio fungicide.
Figure 2.
Pictorial representation of factors that encourage a market for the Trichoderma bio fungicide.

Table 1.
Timeline of historical events in Trichoderma research.
| Decades | Year | Landmarks in Trichoderma Research | References |
|---|---|---|---|
| 1700s | 1794 | In this year, the name Trichoderma was introduced. | [16] |
| 1800s | 1865 | The sexual stage of Trichoderma viride (Hypocrea rufa) was reported. | [17] |
| 1900s | 1932 | First evidence that Trichoderma lignorum (Hypocrea virens) has mycoparasitic and biocontrol abilities. | [18] |
| 1934 | The invention of the first anti-microbial compound (e.g., gliotoxin). | [19] | |
| 1957 | Discovery of the effect of light on T. viride; syn. T. reesei. | [20] | |
| 1972 | Demonstration of biocontrol activity of T. harzianum against Sclerotium rolfsii in field conditions. | [21] | |
| 1979 | RUT C30, a carbon catabolite mutant of T. reesei, was isolated using ultraviolet (UV) mutagenesis. | [22] | |
| 1983 | Cloning of the first Trichoderma species (e.g., T. reesei). | [23] | |
| 1984 | The first international workshop was held on Trichoderma. | - | |
| 1985 | Papavizas wrote the first review on biology, ecology, and potential for biocontrol by Trichoderma. | [15] | |
| 1986 | Report on the expression of growth promotion in the root. | [24] | |
| 1987 | Successful transformation of T. reesei. | [25] | |
| 1989 | First registration of commercial formulation (e.g., Binab T). | [26] | |
| 1992 | Evidence of cloning of lectin-coated fibers by Trichoderma species. | [27] | |
| 1993 | prb1 gene cloning related to mycoparasitism and induced by cell wall. | [28] | |
| 1997 | Expression of enhanced plant immunity (ISR) by application of Trichoderma spp. | [29] | |
| 1998 | Identification of factors that induce genes for mycoparasitism. | [30] | |
| 1999 | Trichoderma internal colonization of plant roots demonstrated. | [31] | |
| 2000s | 2002 | Role of signaling pathways and G protein in mechanism of biocontrol and conidia formation. | [32] |
| 2003 | Role of mitogen-activated protein kinase (MAPK), which negatively regulates conidiation in T. virens. | [33,34] | |
| 2004 | Identification of photoreceptors (Brl1 and Brl2) in T. atroviride. | [35] | |
| 2005 | Function of Trichoderma MAPK (mitogen-activated protein kinase) in Induce systemic resistance (ISR). | [36] | |
| 2006 | Purification of the first true elicitor protein (Sm1/EplI) of Hypocreaatro viridis on glucose. | [37] | |
| 2006 | Cellulases and xylanases regulators identified in T. reesei (XYR1). | [38] | |
| 2008 | First genome sequenced of Trichoderma species (e.g., T. reesei). | [39] | |
| 2009 | Endophytic Trichoderma for stress tolerance in plants. | [40] | |
| 2010 | Discovery of VELVET protein (Vel1) as a key regulator in biocontrol agents (e.g., T. virens) and Trichoderma spp. peptide pheromone precursor genes have been identified. | [41] | |
| 2011 | Genome comparison of three species of Trichoderma. | [42] | |
| 2012 | Next-generation sequencing and high-throughput procedures were used for the sequencing of Trichoderma spp. (T. asperellum, T. harzianum, T. citrinoviride, T. longibrachiatum) genome. | [43] | |
| 2022 | Five new Trichoderma species were reported. | [44] | |
| 2014–2022 | 144 Trichoderma strains were registered in 40 countries. | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kumar, V.; Koul, B.; Taak, P.; Yadav, D.; Song, M. Journey of Trichoderma from Pilot Scale to Mass Production: A Review. Agriculture 2023, 13, 2022. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13102022
AMA Style
Kumar V, Koul B, Taak P, Yadav D, Song M. Journey of Trichoderma from Pilot Scale to Mass Production: A Review. Agriculture. 2023; 13(10):2022. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13102022
Chicago/Turabian StyleKumar, Vipul, Bhupendra Koul, Pooja Taak, Dhananjay Yadav, and Minseok Song. 2023. "Journey of Trichoderma from Pilot Scale to Mass Production: A Review" Agriculture 13, no. 10: 2022. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13102022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.