Chemical Composition of the Cuticle Membrane of Pitaya Fruits (Hylocereus Polyrhizus)


1
Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
2
Provincial Key Laboratory for Fruits and Vegetables Storage Processingand Quality Safety, Agricultural Product Processing Institute, Hunan Academy of Agricultural Sciences, Changsha 410125, China
*
Author to whom correspondence should be addressed.
Agriculture 2019, 9(12), 250; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9120250
Submission received: 27 October 2019
/
Revised: 15 November 2019
/
Accepted: 25 November 2019
/
Published: 27 November 2019
(This article belongs to the Special Issue Postharvest Physiology and Technology of Fruits and Vegetables)
Abstract
:This study comprehensively analysed the chemical composition of the cuticle in pitaya fruits. The total coverage amount of the waxes versus cutin monomers accumulated at a ratio of 0.6, corresponding to masses per unit of 30.3 μg·cm−2 and 50.8 μg·cm−2, respectively. The predominant wax mixtures were n-alkanes in homologous series of C20–C35, dominated by C31 and C33; as well as triterpenoids with an abundant amount of uvaol, lupenon, β-amyrinon, and β-amyrin. The most prominent cutin compounds were C16- and C18-type monomers, in which 9(10),16-diOH-hexadecanoic acid and 9,10-epoxy-ω-OH-octadecanoic acid predominated, respectively. The average chain length (ACL) of aliphates in pitaya fruit cuticle (30.5) was similar to that estimated in leaf waxes, and higher than that in most of the fruit and petal waxes that have been reported. We propose that the relatively high ACL and wax/cutin ratio might enhance the cuticular barrier properties in pitaya fruit cuticle to withstand drought.
1. Introduction
Pitaya or dragon fruits are the fruits of Hylocereus polyrhizus (Weber) Britton and Rose (Cactaceae), which was originally cultivated in Central American. As a cactus species, the modified stems of H. polyrhizus is involved in photosynthesis as opposed to leaves. The leaves are shaped as spines, minimizing the rapid water loss under high temperatures and dry environments [1]. When mature, flesh colour can be white, crimson, or pale-yellow depending on the cultivars. For its sweet taste, the fruit flesh is also used to flavour or colour juices. The pitaya peel is commonly bright-red, representing approximately 22% of the total fruit mass, which is usually discarded during processing [2]. Recently, several studies paid attention to the importance of the peel related to its significance in delaying the shelf life with treatments, i.e., hot air, different temperatures with plastic bags, and ozone gas [1,3,4]. However, the potential importance of the cuticular membrane of the peel has not yet been comprehensively investigated.
The cuticular membranes covering almost all aerial plant organs, protect the cells from uncontrolled water loss as well as other biotic and abiotic stresses [5]. The plant cuticle is composed of lipid components and minor amounts of phenolics, polysaccharides, and proteins. Therefore, the functional properties of cuticle are largely related to the chemical compositions and the structural arrangement of the cuticular layers. As a succulent and drought-tolerant plant, H. polyrhizus might possess a well-stated cuticular membrane, which add, besides the protection of fruits against rapid water loss, the maintenance of the fruit taste and flavour. The present study aims to analyze comprehensively the chemical composition of cuticular waxes and cutin monomers of pitaya fruit peel. The possible relationships between the diversity of chemical compositions in the cuticle and its barrier properties in pitaya fruits will also be discussed.
2. Materials and Methods
2.1. Plant Materials
Red-coloured fruits of H. polyrhizus ‘Hongshuijing’ at the mature red stage were harvested from an orchard in Conghua, Guangzhou, Guangdong Province, P. R. China (23°30′ N, 113°30′ E). The mature pitaya fruits were large (10–15 cm length) with an attractive colour, and bell shaped; with the detailed BBCH (Biologische Bundesantalt, Bundessortenamt und Chemische Industrie) scale previously described [6]. It is largely planted in South China, especially in Conghua, Guangzhou. The water status of pitaya fruit is important for the quality of fruits. Hence, the fruits were transported to laboratory immediately within 3 h. Fruits with uniformity of colour, shape, and free of blemish and disease were selected and used in further experimental materials.
2.2. Chemicals and Standards
All the reagents used for experiments were of analytical grade. Authentic n-tetracosane (CAS#: 646-31-1) and n-dotriacontane (CAS#: 544-85-4) served as standards, N,O-Bis(trimethylsilyl)trifluoroacetamide (BSTFA, CAS#: 25561-30-2), and Boron trifluoride in methanol (1.3 M, CAS#: 373-57-9) were purchased from Sigma-Aldrich (Shanghai, China). Pyridine (CAS#: 110-86-1) was purchased from Shanghai Aladdin Bio-Chem Technology Co., Ltd. (Shanghai, China). Cellulase (CAS#: 90212-54-8) and pectinase (CAS#: 9032-75-1) were purchased from Beijing Solarbio Science and Technology Co., Ltd. (Beijing, China).
2.3. Isolation of Cuticular Membranes
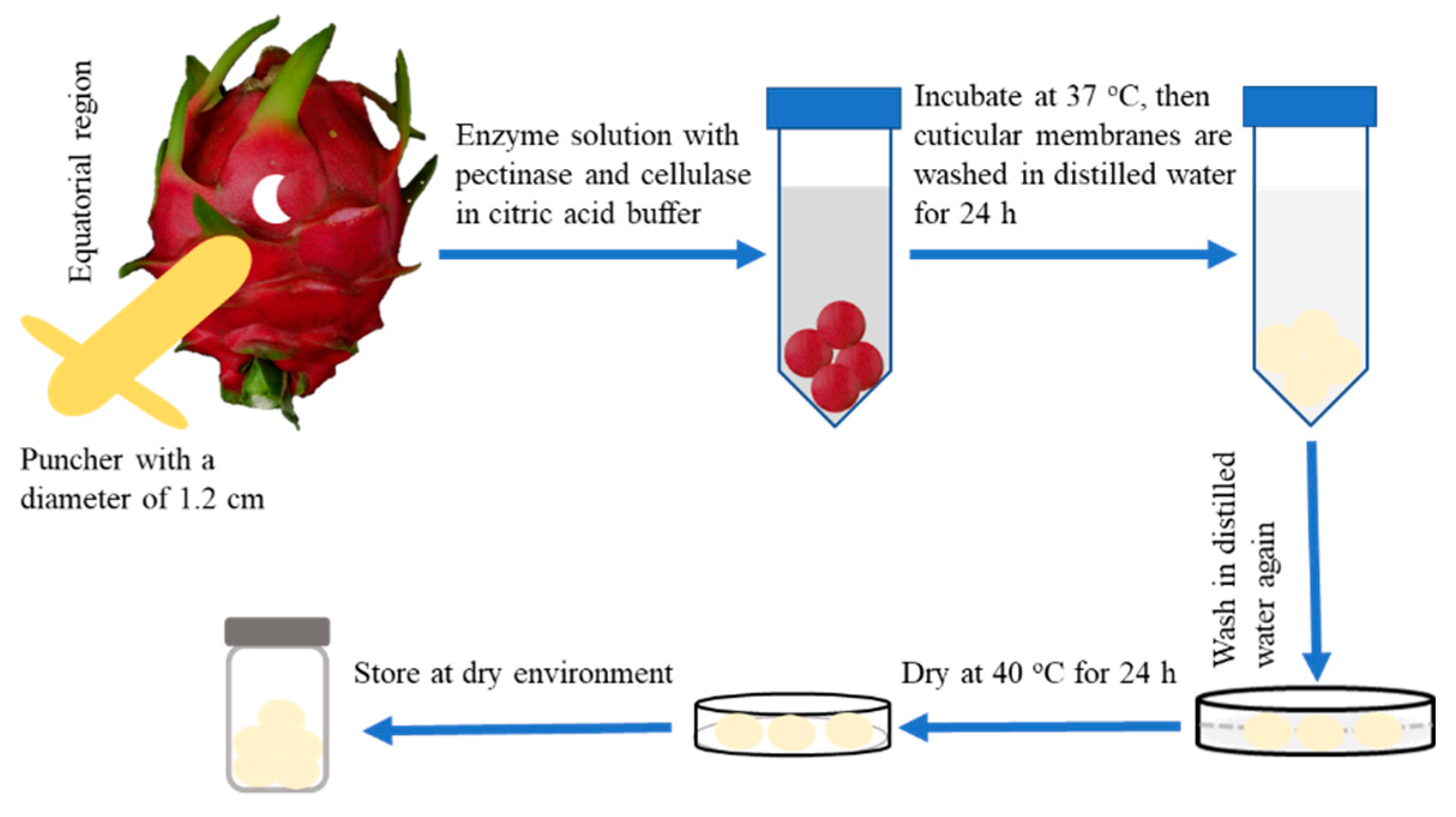
Cuticular membranes (CMs) from pitaya fruits were isolated enzymatically. To isolate the CMs from pitaya fruit, discs were punched from the fresh, non-contaminated and non-treated fruit peels at the middle section of the fruit using a puncher with diameter of 1.2 cm (Runzekang Biology Technology Co., Ltd., Beijing, China). The discs were immediately immersed in an enzyme solution, containing 10 mM citric acid buffer with pectinase (1% w/v), cellulase (1% w/v) and 0.1 mM sodium azide to avoid the growth of microorganisms. After the CMs were completely isolated from tissues at room temperature for 7 day, the CMs were washed with 10 mM sodium tetraborate decahydrate and distilled water, then dried under dry atmosphere (with dry silica gel) for further experiments (Figure 1).
2.4. Cuticular Wax Extraction
To extract the cuticular waxes from the pitaya fruit cuticle, the enzymatically isolated fruit CMs were dipped and extracted three times in 50 °C hot chloroform, each time for 60 s. The two extracts per each disc obtained were combined, and n-tetracosane was added. The extracted mixtures were evaporated under a gentle stream of nitrogen gas until obtaining dry samples for further analysis.
2.5. Cutin Depolymerization for Chemical Analysis
For the cutin analysis, the matrix obtained by wax extraction as described above was subsequently immersed in boron trifluoride dissolved in methanol (1.3 M) and depolymerized at 70 °C for 16 h. After extraction in chloroform, n-dotriacontane was added. Then, a saturated aqueous sodium chloride solution was added. The mixtures were further extracted by chloroform three times. The collected organic phase, which contained cutin monomers was dried over sodium sulphate, and evaporated under a gentle stream of nitrogen gas until dry samples were obtained for further analysis.
2.6. Chemical Analyses: Gas Chromatography and Mass Spectrometry
The above dried extracts were then derivatized with BSTFA in pyridine at 70 °C for 30min. To detect the wax and cutin monomer components, the extracts were analysed using a capillary gas chromatograph with a flame ionization detector (7820A, GC System; Agilent Technologies, Santa Clara, CA, USA), and on-column injection with a capillary column (30 m × 0.32 mm, DB-1 ms, 0.1 μm film; J&W Scientific, Agilent Technologies, Santa Clara, California, USA). To separate the cuticular wax compounds, the samples were injected, and kept at 50 °C for 2 min. Then, the temperature was raised at a rate of 40 °C min−1 to 200 °C, and kept at 200 °C for 2 min, then raised at a rate of 3 °C min−1 to 320 °C and kept at 320 °C for 30 min. For separation of the cutin monomers, samples were injected and kept at 50 °C for 1 min. Then the temperature was raised at a rate of 10 °C min−1 to 150 °C, kept at 150 °C for 2 min, raised at a rate of 3 °C min−1 to 320 °C and kept at 320 °C for 30 min. The sample volume was 10 µL. The carrier gas was hydrogen. The area of the peaks was compared with that of the internal standard to obtain the quantity of cuticular wax and cutin monomer components per unit cuticle area.
The chemical components were analysed using a temperature-controlled capillary gas chromatograph equipped with a mass spectrometric detector (m/z 50–750, MSD 5975; Agilent Technologies, Santa Clara, California, USA) under the same gas chromatographic conditions but with helium as carrier gas. Single compounds were identified based on their electron ionization mass spectra using authentic standards, the Wiley 10th/NIST 2014 mass spectral library (W10N14; John Wiley and Sons) or by interpretation of the spectra according to their retention times and/or by comparison with a mass spectra library established by our own laboratory or with literature data [7,8,9,10].
The component coverage (Cs) was quantified against the amount of internal standard (Mis) by integrating the peak area of the component (As) and the peak area of internal standard (Ais), and dividing by the extracted area (Aea):
The weighted average of carbon chain length (ACL) of aliphatic compounds was calculated from the chain length (Li,) and the molar mass fraction (Mi) of the component (i):
2.7. Statistical Analysis
Statistical analyses were performed using SPSS 17.0 (IBM Corp., Armonk, NY, USA) and SigmaPlot 10.0 (Systat Software, Inc., San Jose, CA, USA). SigmaPlot 10 was used to generate the graphs.
3. Results and Discussion
3.1. Composition of Cutin of Pitaya Fruit
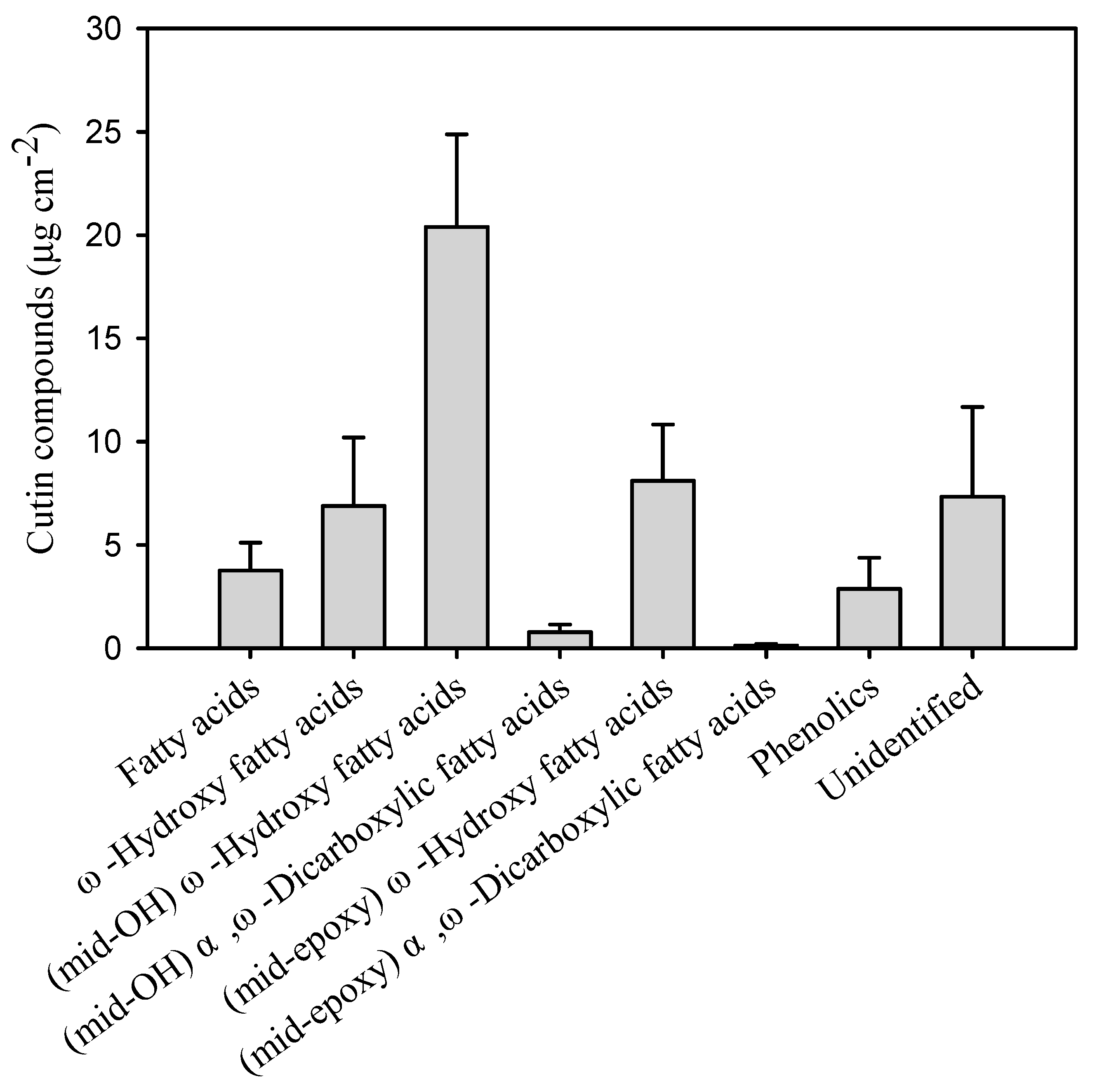
The chemical composition and quantity of the compounds of the cutin matrix involved in the cuticle of pitaya fruits were analysed and detail in this study. The overall coverage of the cutin matrix was 50.8 μg cm−2 (Table 1). The cutin monomers were largely accumulations of fatty acids (7.7% of total cutin) without added groups, ω-hydroxy fatty acids (13.4%), ω-hydroxy fatty acids with mid-chain hydroxy groups (39.8%) and ω-hydroxy fatty acids with mid-chain epoxy groups (15.7%, Figure 1 and Supplementary Table S1). The predominant cutin compounds were 9/10,16-dihydroxy-hexadecanoic acid (37.5% of the total cutin mass), 9,10-epoxy-ω-hydroxy-octadecanoic acid (14.0%) and ω-hydroxy-octadec-9/12-enoic acid (7.9%, Figure 2 and Supplementary Table S1). Small amounts of fatty acids with most even-numbered carbon chain lengths from C15 to C32 (7.7% of the total cutin), α,ω-dicarboxylic fatty acids with mid-chain hydroxy group (1.5%), and α,ω-dicarboxylic fatty acids with mid-chain epoxy group (<1%) were also identified in the fruit cutin matrix (Figure 1 and Supplementary Table S1). The unsaturation degree of the cutin matrix was 14.3%, and the epoxidation degree was 15.9%. Furthermore, the ratio of the carbon chain length of C16 and C18 cutin monomers amounted to 1.7 (Table 1).
In pitaya fruit, the most abundant cutin monomers were of C16 and C18 chain length. The predominant 9/10,ω-dihydroxyhexadecanoic acid was also detected in other fruits such as Prunus avium L. [11], Solanum lycopersicum L. [12], Olea europaea L. [13], and Malus domestica Borkh. [14]. In contrast, the 9,10-epoxy-ω-18-hydroxy-octadecanoic acid detected as the second most abundant monomers in pitaya fruit cuticle (14.0% of total cutin) are commonly found in the leaf cutin matrix of examples such as Olea europaea L., Rhazya stricta and Agave Americana etc., but was scarcely detected in fruit cuticle of olive and apple [9,12,13,15,16].
The cutin matrix, polymerized by mostly oxygenated fatty acids, is proposed to construct the framework of the plant cuticle [17]. However, cuticular waxes rather than cutin polymers may form the crucial barrier against uncontrolled water loss [18,19]. The chain-length distribution of cutin monomers in the matrix was also found to slightly affect the permeability to water and organic or inorganic substances [20]. The abundant accumulation of C18 chains could lead to a slower permeability unlike that of C16 chains. Previous investigations implied that a unique polymeric cutin matrix might result from the various characteristics of carbon chain lengths, the degree of unsaturation, and mid-chain epoxidation in both leaf and fruit cuticle [13]. The ratio of C16/C18 cutin monomers was 1.7, the C18-long chains of 9,10-epoxy-ω-hydroxyoctadecanoic acid was detected, which might add to enhancement of the mechanical strength of the cutin matrix in pitaya fruits. In addition, the substantial content of unsaturated and mid-chain epoxidation cutin monomers might also broaden the network of polymers in the matrix, providing spaces for wax accumulation in the cuticle of pitaya fruits.
3.2. Composition of Cuticular Waxes of Pitaya Fruit
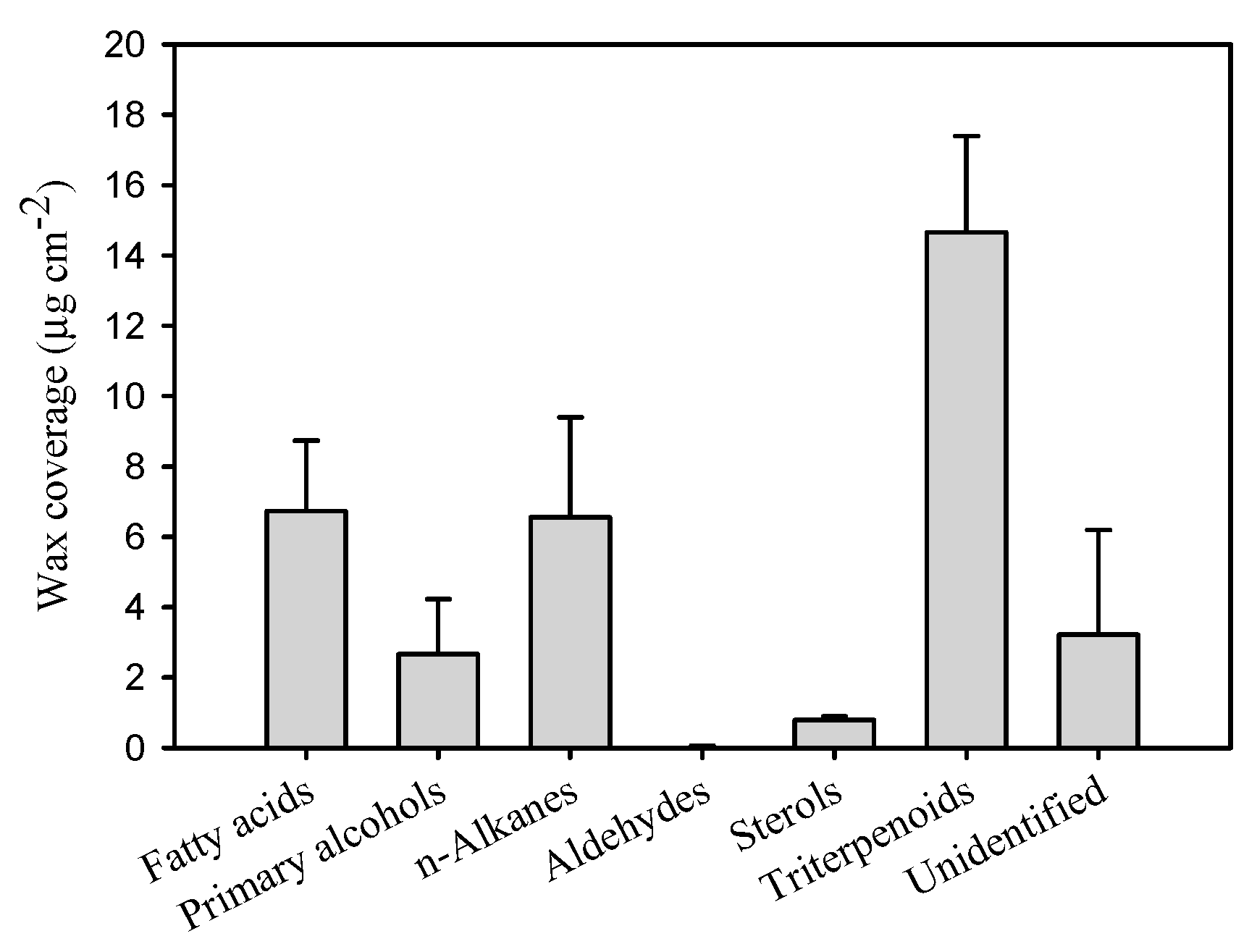
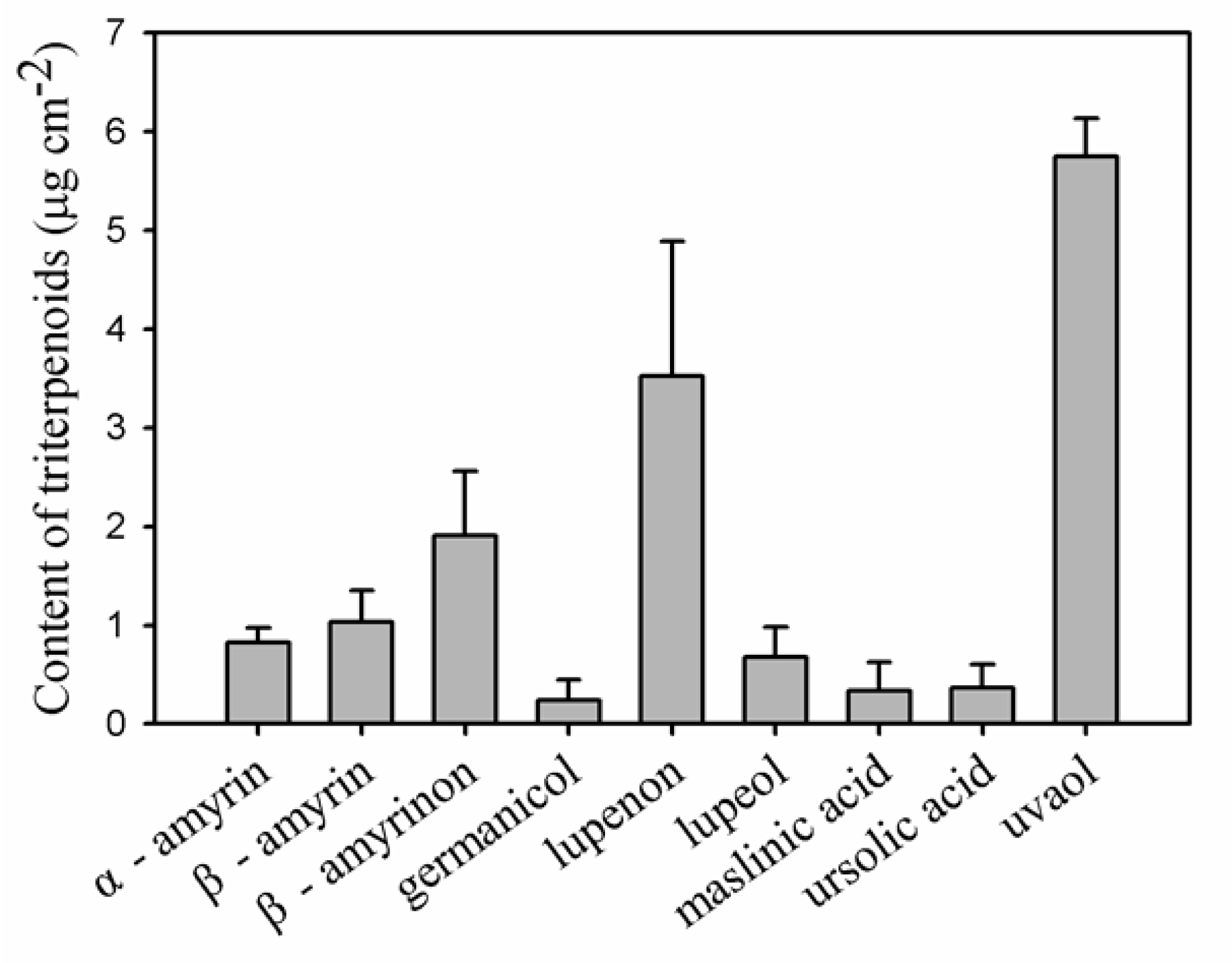
As cuticular waxes are the main compounds resulting in the functional pattern for the barrier properties of cuticle, the chemical composition of the cuticular waxes was thoroughly analysed for pitaya fruit. The total wax coverage was 30.3 μg cm−2 (Table 1), which was similar to the wax coverage on the fruits of Prunus avium L. [11] and Solanum lycopersicum L. [12]. Typical very-long-chain fatty acids (VLCFAs) and their derivatives, including fatty acids, primary alcohols, aldehydes, and n-alkanes, as well as a variety of sterols and pentacyclic triterpenoids were detected in pitaya cuticular wax mixtures (Figure 3 and Supplementary Table S2). In particular, n-alkanes (17.6% of total wax mass) and the variety of pentacyclic triterpenoids (44.8% of total wax mass) were the prominent components. The pentacyclic triterpenoids contained various components with the most abundant being uvaol (18.8% of total wax mass), followed by lupenon (9.8% of total wax mass), β-amyrinon (5.5% of total wax mass), β-amyrin (3.0% of total wax mass), α-amyrin (2.7% of total wax mass), lupeol (1.9% of total wax mass), ursolic acid (1.4% of total wax mass), as well as small amount of maslinic acid and germanicol (Figure 4 and Supplementary Table S2). Relatively small amounts of other VLCFAs such as fatty acids (2.1% of total wax mass, the chain-length larger than 19 carbons, fatty acids in C16 and C18 were excluded), primary alcohols (6.6% of total wax mass), and trace of aldehydes, as well as various cyclic sterols (2.7% of total wax mass) were detected (Figure 3 and Supplementary Table S2).
Additionally, the diversity in the accumulation and amount of VLCFAs (>19 carbon chain-length), i.e., aliphates, versus the cyclic compounds including sterols and pentacyclic triterpenoids, was in a ratio of 0.7 (Table 1), which showed similar amounts accumulating for aliphates and cyclics in the cuticular wax of pitaya fruit cuticle. The n-alkanes and pentacyclic triterpenoids being the major wax components in pitaya fruit cuticle, which were similar to the wax mixtures of tomato fruit cuticle [12], sweet cherry [11], as well as similar to the leaf waxes of most previously studied plant species [13,16,21]. However, this differed from fruit cuticular waxes of many other plant species [13,14].
Simultaneously, a ratio of 0.3 for the accumulated amount of triterpenoids versus cutin monomers, and a ratio of 0.6 for total wax versus total cutin monomers, were found (Table 1). So far, few studies have tried to take the triterpenoids into account as fillers to reinforce the cutin matrix mechanically. The cuticular wax, especially the intracuticular wax has been reported to fix and restrict the strain of cuticle of leaves and fruits [22]. The triterpenoids were detected mainly in intracuticualr waxes [21]. Therefore, triterpenoids might be one of the main wax fractions embedded in the cutin matrix. It has been implied that triterpenoids accumulated in the leaf cuticles of R. stricta might play an important role in protecting the barrier of cuticular layer from thermal stress under very hot and drought environments as found in desert plants [16]. In addition, the triterpenoids were indicated to be nanofillers embedded in the cuticular matrix, which may increase the mechanical strength of the cuticle in Fuyu Persimmon fruit [23]. Likewise, as a member of the cactus family, pitaya usually sustains high temperatures and less water resources. Thus, the relatively high amount of triterpenoids embedding in cuticular layer together with cutin matrix may also help to adapt the thermal stress, maintaining the integrity of cuticular membrane for the pitaya fruit cuticle.
3.3. Chain Length Distribution of Aliphates and their Putative Significance for Barrier Properties
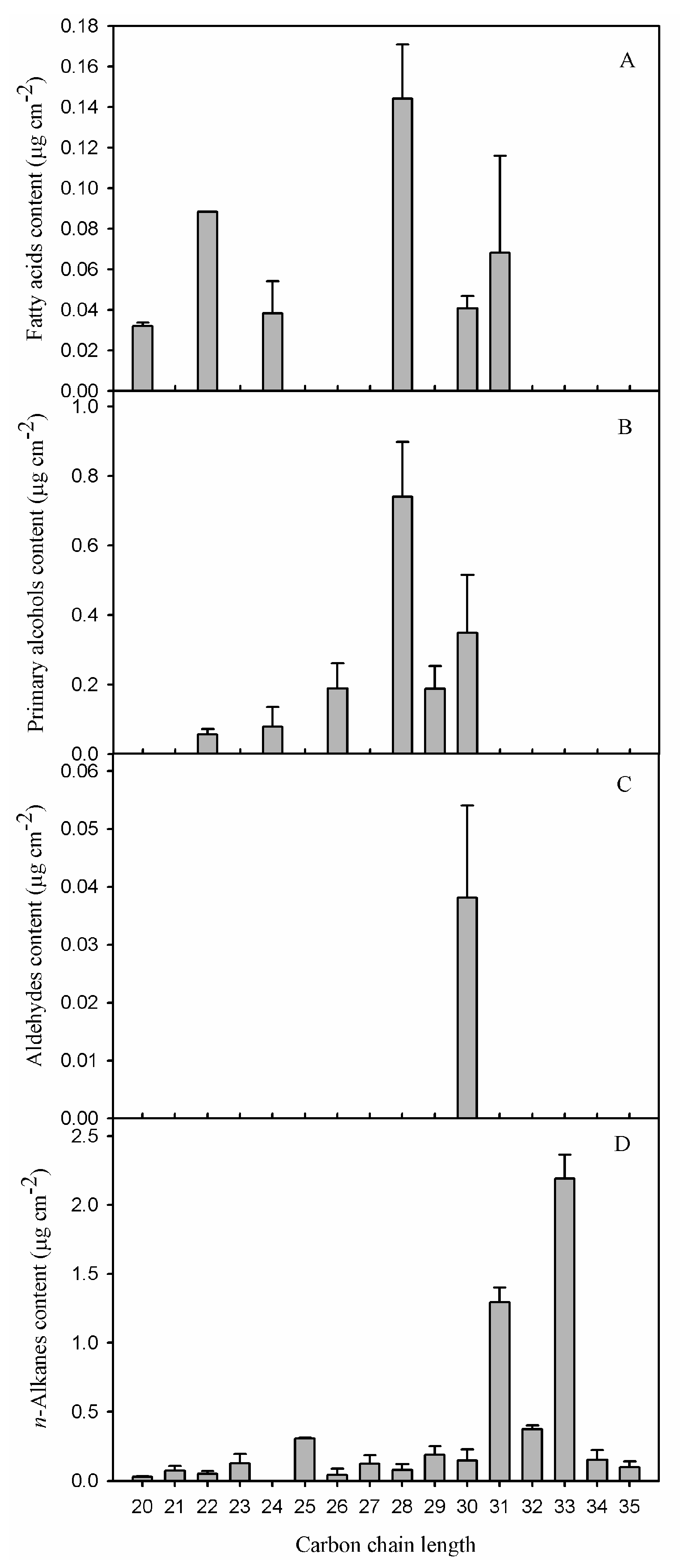
It has been pointed out that the accumulation of total wax or homologous substances could not directly contribute to the barrier properties, but is more likely related to the chain-length distribution of straight chain components based on the comprehensive studies in various petals [24], leaves [5,21] and fruits [25]. The chain-length distribution of VLCFAs differed in each compositional class. Fatty acids and primary alcohols of the pitaya fruit had a similar carbon chain-length distribution, which ranged from C20 to C31 and from C22 to C30, respectively (Figure 5A,B). Similar small amounts of each carbon chain were found for fatty acids, while the primary alcohol fraction predominated by C28 and C30. The aldehydes with trace of C30 were found in pitaya fruit cuticle (Figure 5C). In contrast, the predominant VLCFAs fraction of n-alkanes occurred in a fairly broad range with a continuous homologous of chain-length from C20 to C35. Remarkably, high concentrations of C31 (4.3% of total wax) and C33 (7.3%) were deposited and found to be the most abundant of n-alkanes in pitaya fruit waxes (Figure 5D).
A variable chain-length distribution of aliphatic waxes in pitaya fruit cuticle was comparable to the previously studied fruits, and/or other plant organs such as leaves and petals. For example, the aliphatic pattern of waxes was dominated by C29 n-alkanes in cherry fruit cuticle [11]; by odd-numbered n-alkanes ranging from C29 to C33 in tomato fruit cuticle [12]; by a mixture of C30 aldehydes, fatty acids, and alcohols, as well as C29 n-alkanes in berry fruit cuticle [26]; and by C26 and C28 for both fatty acids and primary alcohols in olive fruit cuticle [13] etc. The predominant aliphates were odd-numbered n-alkanes that ranged from C29 to C33 in most leaf waxes that have been investigated [13,21,27]. In pitaya fruit cuticle, the most abundant n-alkanes were dominated by C31 and C33 (Supplementary Table S2), which were about two carbons longer, when compared to other fruits, and were more similar to that of leaf waxes.
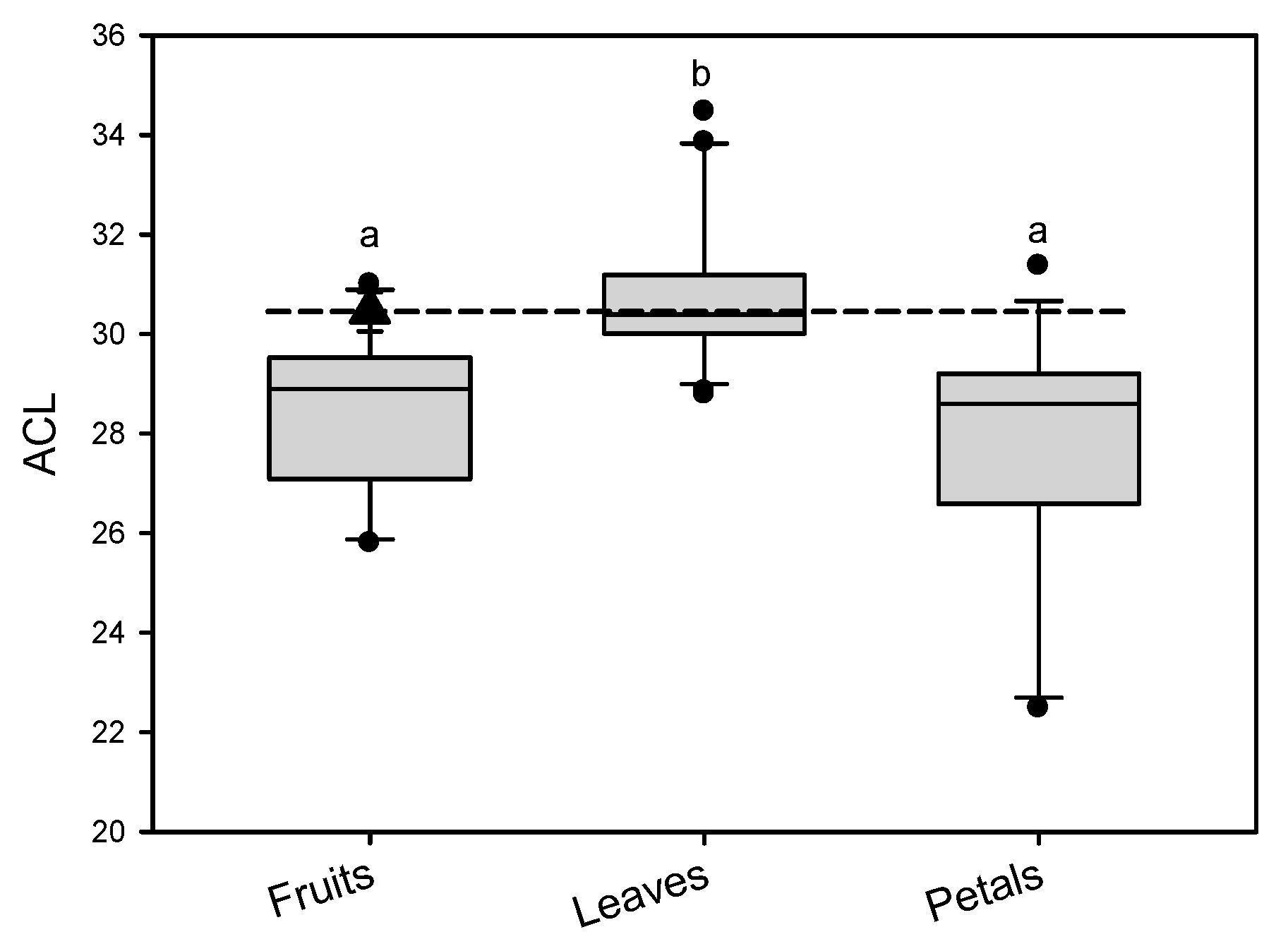
It has long been mentioned that the barrier properties of the cuticle in each plant species and different organs were largely related to the accumulation of VLCFAs, especially their chain-length distributions [28]. Given the wide range distribution of carbon chain lengths and variety of compositions in cuticular waxes, ACL was used as a proxy to indicate the overall wax compositional quality [29]. ACL has recently been noticed as one of the important parameters leading barrier property differences between leaves and fruits, and/or petals [13,24]. In the present study, the weighted ACL of aliphatic compounds, which were calculated based on the molar concentration of VLCFAs, were 30.5 in pitaya fruit cuticle (Table 1). The ACL loading has been reported in the cuticle of a variety of plant species and/or different fruits [11,13,14], leaves [13,21,24], and petals [24,30] reported previously as comparison groups. ACL of aliphates in pitaya fruit cuticle was similar to the median value in leaf waxes, and higher than that in most of the fruit and petal cuticular waxes previously reported (Figure 6). Previous research reported that water permeability of leaves was significantly lower than that of fruits and petals [12,24,25], which was putatively related to the higher ACL loading in the leaf waxes. Therefore, the relatively high ACL value in pitaya fruit might provide an efficient barrier for the cuticular membrane, leading to slow water transpiration and maintaining fruit water status for pitaya. Certainly, this could also be an intrinsic/unique characteristic for pitaya to withstand high temperature and arid stresses.
4. Conclusions
In conclusion, the chemical composition of cuticular waxes and cutin monomers in pitaya fruit peel has been characterized and discussed to link to the eco-physiological adaptation of pitaya fruit cuticle. The cuticular membrane of pitaya fruit contained both abundant VLCFAs dominated by n-alkanes (C31 and C33), and various pentacyclic triterpenoids for the wax pattern, embedded incutin matrix composed of mixture of C16, and C18 chain cutin monomers (Figure 7). Similar amount of waxes and cutin monomers were detected in cuticle of pitaya fruit. ACL of aliphatic compounds of waxes in pitaya fruit cuticle was similar to that in leaves and higher than that in the cuticle of fruits and petals of most previously studied plant species [13,24,25]. On the one hand, the abundant triterpenoids and equivalent of wax coverage and cutin polymer in pitaya fruit cuticle might strengthen the mechanical support/stability and the plasticity of the cuticular membrane, protecting the cuticle from thermal extension for pitaya to adapt to high-temperature stress. On the other hand, the relatively high ACL of VLCFA loads could enhance the tight and highly ordered crystalline zones in cuticular wax layer, leading an efficient transpiration barrier for rapid water loss for pitaya in arid habits. Altogether, the results of wax and cutin in pitaya fruit cuticle add further insights into the relationship between the chemical composition and the eco-physiological context for the plant fruit cuticle.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2077-0472/9/12/250/s1, Table S1: Chemical composition of the cutin matrix in the pitaya fruit peel (µg cm−2). Data are given as means ± standard deviations (n = 5), Table S2: Wax composition of the pitaya fruit cuticle (µg cm−2). Data are given as means ± standard deviations (n = 5).
Author Contributions
Conceptualization, H.H. and Y.J.; methodology, H.H.; formal analysis, H.H.; investigation, H.H.; writing—original draft preparation, H.H.; writing—review and editing, H.H. and Y.J.; project administration, H.H. and Y.J.; funding acquisition, H.H. and Y.J.
Funding
This work was supported by the National Natural Science Foundation of China (31770726), the “Pearl River Talent Plan” Postdoctoral Program of Guangdong Province (2018(02)), China Postdoctoral Science Foundation (2018M643228), and The Open Project of Hunan Key Lab of Fruits and Vegetables Storage, Processing, Quality and Safety (HKLFP201901).
Acknowledgments
We extend sincere thanks and appreciation to Afiya John, an English native speaker, who kindly assisted in improving this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hoa, T.T.; Clark, C.J.; Waddell, B.C.; Woolf, A.B. Postharvest quality of Dragon fruit (Hylocereus undatus) following disinfesting hot air treatments. Postharvest Biol. Technol. 2006, 41, 62–69. [Google Scholar] [CrossRef]
- Jamilah, B.; Shu, C.E.; Kharidah, M.; Dzulkily, M.A.; Noranizan, A. Physico-chemical characteristics of red pitaya (Hylocereus polyrhizus) peel. Int. Food Res. J. 2011, 18, 279–285. [Google Scholar]
- Freitas, S.T.D.; Mitcham, E.J. Quality of pitaya fruit (Hylocereus undatus) as influenced by storage temperature and packaging. Sci. Agric. 2013, 70, 257–262. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, Z.; Zhu, H.; Li, T.; Zhu, X.; Gao, H.; Yun, Z.; Jiang, Y. Comparative volatile compounds and primary metabolites profiling of pitaya fruit peel after ozone treatment. J. Sci. Food Agric. 2018. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef]
- Kishore, K. Phenological growth stages of dragon fruit (Hylocereus undatus) according to the extended BBCH-scale. Sci. Hortic. 2016, 213, 294–302. [Google Scholar] [CrossRef]
- Burnouf-Radosevich, M.; Delfel, N.E.; England, R. Gas chromatography-mass spectrometry of oleanane-and ursane-type triterpenes-application to Chenopodium quinoa triterpenes. Phytochemistry 1985, 24, 2063–2066. [Google Scholar] [CrossRef]
- Franke, R.; Briesen, I.; Wojciechowski, T.; Wojciechowski, T.; Faust, A.; Yephremov, A.; Nawrath, C.; Schreiber, L. Apoplastic polyesters in Arabidopsis surface tissues–a typical suberin and a particular cutin. Phytochemistry 2015, 66, 2643–2658. [Google Scholar]
- Holloway, P. The Chemical Constitution of Plant Cutins. In The Plant Cuticle; Cutler, D.F., Alvin, K.L., Price, C., Eds.; Academic Press: London, UK, 1982; pp. 45–85. [Google Scholar]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of plant cuticular waxes. Biol. Plant Cuticle 2008, 23, 145–181. [Google Scholar]
- Peschel, S.; Franke, R.; Schreiber, L.; Knoche, M. Composition of the cuticle of developing sweet cherry fruit. Phytochemistry 2007, 68, 1017–1025. [Google Scholar] [CrossRef]
- Leide, J.; Hildebrandt, U.; Reussing, K.; Riederer, M.; Vogg, G. The developmental pattern of tomato fruit wax accumulation and its impact on cuticular transpiration barrier properties: Effects of a deficiency in a β-ketoacyl-coenzyme A synthase (LeCER6). Plant Physiol. 2007, 144, 1667–1679. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Burghardt, M.; Schuster, A.-C.; Leide, J.; Lara, I.; Riederer, M. Chemical composition and water permeability of fruit and leaf cuticles of Olea europaea L. J. Agric. Food Chem. 2017, 65, 8790–8797. [Google Scholar] [CrossRef] [PubMed]
- Leide, J.; de Souza, A.X.; Papp, I.; Riederer, M. Specific characteristics of the apple fruit cuticle: Investigation of early and late season cultivars ‘Prima’ and ‘Florina’ (Malus domestica Borkh.). Sci. Hortic. 2018, 229, 137–147. [Google Scholar] [CrossRef]
- Holloway, P.J.; Deas, A.H.B. Epoxyoctadecanoic acids in plant cutins and suberins. Phytochemistry 1973, 12, 1721–1735. [Google Scholar] [CrossRef]
- Schuster, A.-C.; Burghardt, M.; Alfarhan, A.; Bueno, A.; Hedrich, R.; Leide, J.; Thomas, J.; Riederer, M. Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants 2016, 8, pii: plw027. [Google Scholar] [CrossRef]
- Fich, E.A.; Segerson, N.A.; Rose, J.K. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef]
- Riederer, M. Waxes: The transport barriers of plant cuticles. In Waxes: Chemistry, Molecular Biology and Functions; Hamilton, R.J., Ed.; The Oily Press: Dundee, Scotland, 1995; pp. 130–156. [Google Scholar]
- Schönherr, J. Water permeability of isolated cuticular membranes: The effect of cuticular waxes on diffusion of water. Planta 1976, 131, 159–164. [Google Scholar] [CrossRef]
- Schreiber, L.; Schonherr, J. Water and Solute Permeability of Plant Cuticles; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Jetter, R.; Riederer, M. Localization of the transpiration barrier in the epi-and intracuticular waxes of eight plant species: Water transport resistances are associated with fatty acyl rather than alicyclic components. Plant Physiol. 2015, 170, 921–934. [Google Scholar] [CrossRef]
- Khanal, B.P.; Grimm, E.; Finger, S.; Blume, A.; Knoche, M. Intracuticular wax fixes and restricts strain in leaf and fruit cuticles. New Phytol. 2013, 200, 134–143. [Google Scholar] [CrossRef]
- Tsubaki, S.; Sugimura, K.; Teramoto, Y.; Yonemori, K.; Azuma, J. Cuticular Membrane of Fuyu Persimmon Fruit is Strengthened by Triterpenoid Nano-Fillers. PLoS ONE 2013, 8, e75275. [Google Scholar] [CrossRef]
- Cheng, G.; Huang, H.; Zhou, L.; He, S.; Zhang, Y.; Cheng, X.A. Chemical composition and water permeability of the cuticular wax barrier in rose leaf and petal: A comparative investigation. Plant Physiol. Biochem. 2019, 135, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Huang, H. Comparative Investigation of the Chemical Composition and the Water Permeability of Fruit and Leaf Cuticles. Ph.D. Thesis, University of Würzburg, Würzburg, Germany, 2017. [Google Scholar]
- Chu, W.; Gao, H.; Cao, S.; Fang, X.; Chen, H.; Xiao, S. Composition and morphology of cuticular wax in blueberry (Vaccinium spp.) fruits. Food Chem. 2017, 219, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Jenks, M.A.; Tuttle, H.A.; Eigenbrode, S.D.; Feldmann, K.A. Leaf epicuticular waxes of the eceriferum mutants in Arabidopsis. Plant Physiol. 1995, 108, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Riederer, M.; Schneider, G. The effect of the environment on the permeability and composition of Citrus leaf cuticles. Planta 1990, 180, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Poynter, D. Judging the duration of time intervals: A process of remembering segments of experience. Adv. Psychol. 1989, 59, 305–331. [Google Scholar]
- Buschhaus, C.; Hager, D.; Jetter, R. Wax layers on Cosmos bipinnatus petals contribute unequally to total petal water resistance. Plant Physiol. 2015, 167, 80–88. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic of the protocol for isolating the cuticular membranes from pitaya fruit peel.

Figure 2.
Wax compounds in the pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).
Figure 2.
Wax compounds in the pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).

Figure 3.
Chemical compounds of the cuticular waxes in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).
Figure 3.
Chemical compounds of the cuticular waxes in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).

Figure 4.
Content of triterpenoids accumulation in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).
Figure 4.
Content of triterpenoids accumulation in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).

Figure 5.
Chain-length distribution and content of very-long-chain fatty acids and their derivatives in pitaya fruit. (A) fatty acids; (B) primary alcohols; (C) aldehydes; and (D) n-alkanes. Data are given as means ± standard deviation (n = 5).
Figure 5.
Chain-length distribution and content of very-long-chain fatty acids and their derivatives in pitaya fruit. (A) fatty acids; (B) primary alcohols; (C) aldehydes; and (D) n-alkanes. Data are given as means ± standard deviation (n = 5).

Figure 6.
The comparisons of ACL (average chain length) load in the pitaya fruit cuticle to the different fruits, leaves, and petals in different plant species. The dark triangle corresponds to the ACL value of pitaya fruit cuticle. The different lowercase letters on the bars indicate significant differences of ACL load in cuticle of different fruits, leaves and petals (p < 0.05).
Figure 6.
The comparisons of ACL (average chain length) load in the pitaya fruit cuticle to the different fruits, leaves, and petals in different plant species. The dark triangle corresponds to the ACL value of pitaya fruit cuticle. The different lowercase letters on the bars indicate significant differences of ACL load in cuticle of different fruits, leaves and petals (p < 0.05).

Figure 7.
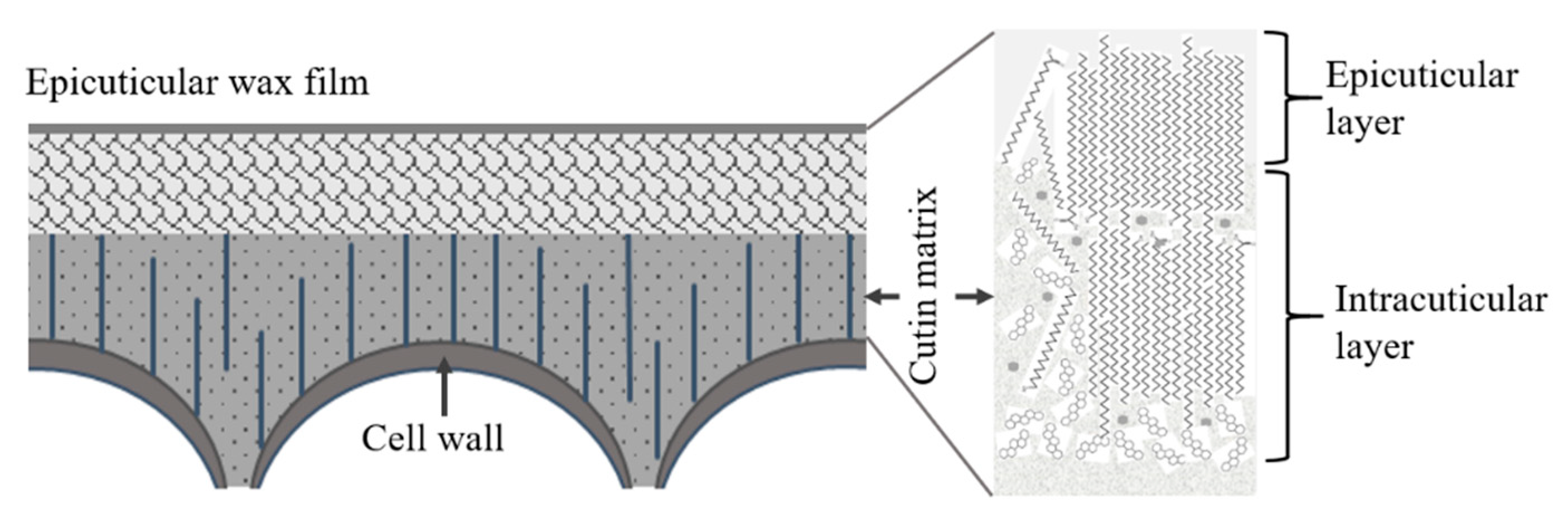
Schematic of the proposed cuticule structure of pitaya fruit (not drawn in scale).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Total coverage of wax and cutin matrix, as well as the content of aliphatics, cyclics, C16 and C18 monomers; Ratio of very long chain aliphatic fraction to cyclic fraction, ratio of C16 to C18 monomers, total wax to cutin; and average chain length (ACL) of the very long chain aliphatic fraction in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).
Table 1.
Total coverage of wax and cutin matrix, as well as the content of aliphatics, cyclics, C16 and C18 monomers; Ratio of very long chain aliphatic fraction to cyclic fraction, ratio of C16 to C18 monomers, total wax to cutin; and average chain length (ACL) of the very long chain aliphatic fraction in pitaya fruit cuticle. Data are given as means ± standard deviation (n = 5).
| Waxes (µg cm−2) | Cutin Monomers (µg cm−2) | Ratios | ACL | ||||
|---|---|---|---|---|---|---|---|
| Total wax | 30.3 ± 1.47 | Total cutin | 50.84 ± 7.4 | Aliphatics/cyclics | 0.7 ± 0.11 | Aliphates | 30.45 ± 0.76 |
| Aliphates (≥C20) | 9.93 ± 0.99 | C16 monomers | 24.15 ± 5.33 | C16/C18 | 1.71 ± 0.48 | ||
| Cyclics | 14.38 ± 1.38 | C18 monomers | 14.92 ± 4.84 | Total wax/cutin | 0.57 ± 0.05 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, H.; Jiang, Y. Chemical Composition of the Cuticle Membrane of Pitaya Fruits (Hylocereus Polyrhizus). Agriculture 2019, 9, 250. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9120250
AMA Style
Huang H, Jiang Y. Chemical Composition of the Cuticle Membrane of Pitaya Fruits (Hylocereus Polyrhizus). Agriculture. 2019; 9(12):250. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9120250
Chicago/Turabian StyleHuang, Hua, and Yueming Jiang. 2019. "Chemical Composition of the Cuticle Membrane of Pitaya Fruits (Hylocereus Polyrhizus)" Agriculture 9, no. 12: 250. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9120250
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.