
The Effects of Platelet-Rich Fibrin in the Behavior of Mineralizing Cells Related to Bone Tissue Regeneration—A Scoping Review of In Vitro Evidence
 ,
,  and
and
Abstract
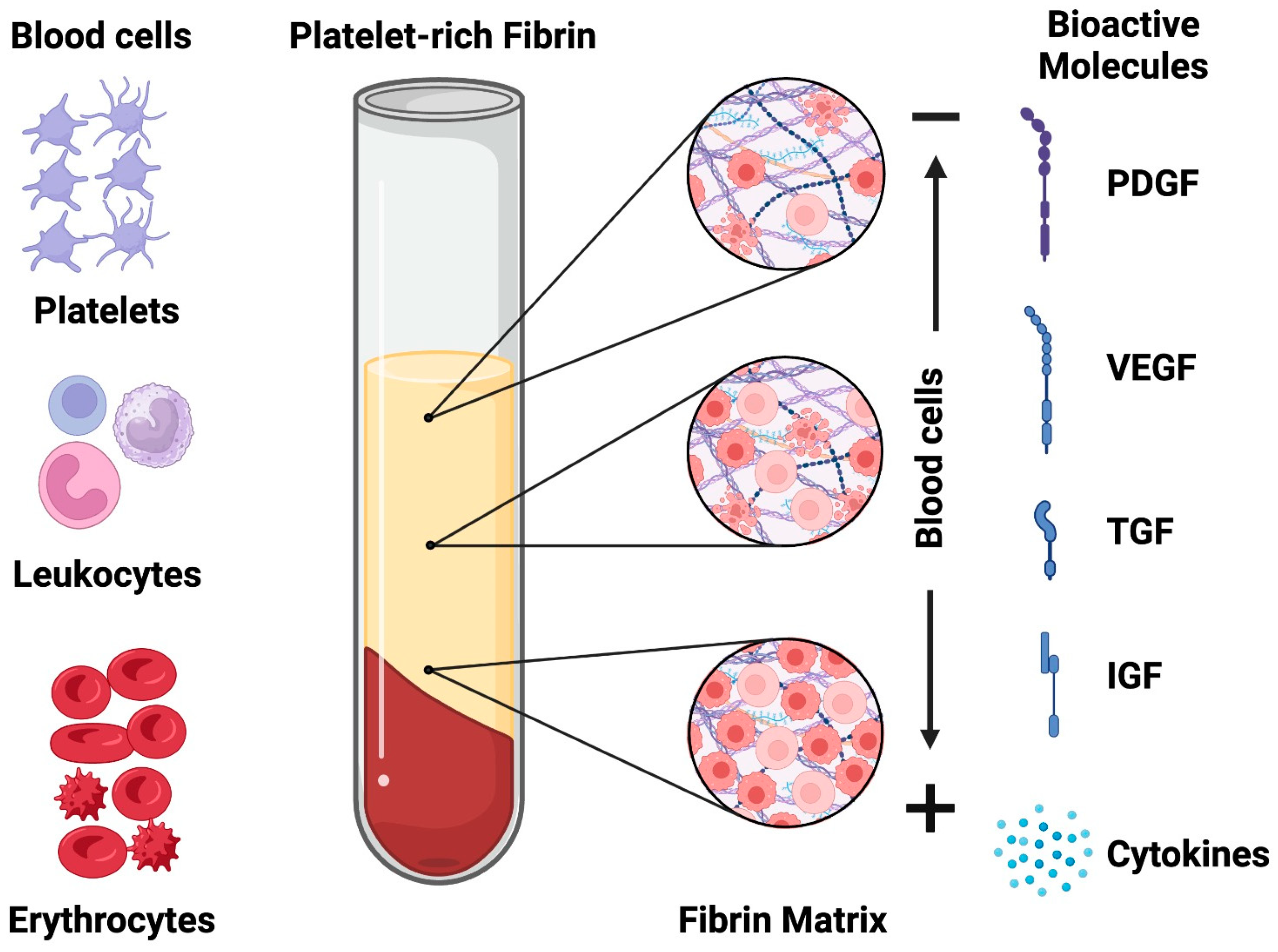
:1. Introduction
2. Materials and Methods
2.1. Protocol and Registry
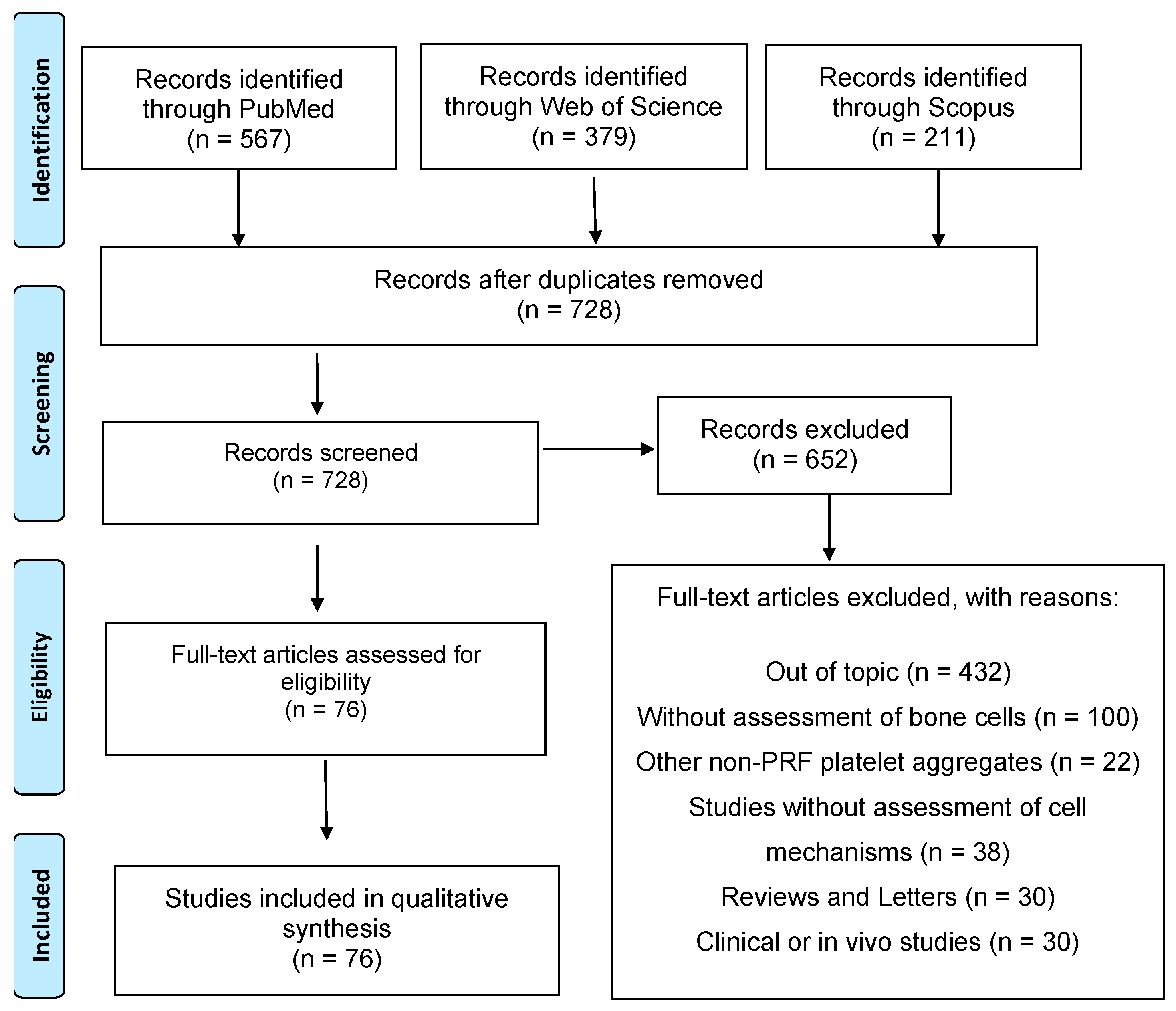
2.2. Information Sources and Search Strategy
2.3. Study Selection
2.4. Critical Appraisal
2.5. Data Extraction
3. Results
4. Discussion
4.1. Protocols for PRF Production and Preservation
4.2. Association of PRF and Other Materials/Compounds
4.3. Cell-Type Related Effects of PRF
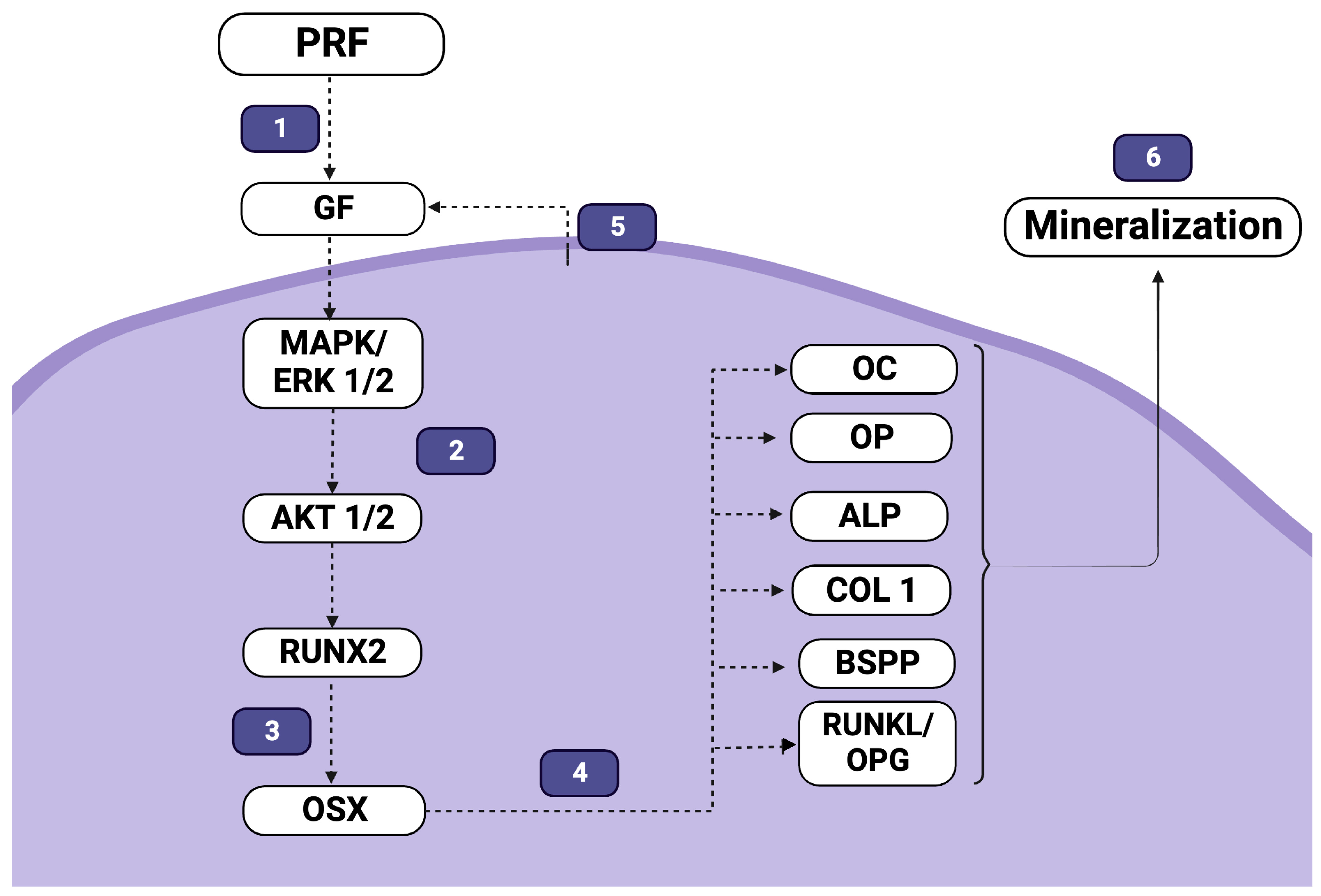
4.4. The Molecular Effects of PRF on Differentiation and Mineralization
5. Final Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: Current concepts and future directions. BMC Med. 2011, 9, 66. [Google Scholar] [CrossRef]
- Miron, R.J.; Zucchelli, G.; Pikos, M.A.; Salama, M.; Lee, S.; Guillemette, V.; Fujioka-Kobayashi, M.; Bishara, M.; Zhang, Y.; Wang, H.L.; et al. Use of platelet-rich fibrin in regenerative dentistry: A systematic review. Clin. Oral Investig. 2017, 21, 1913–1927. [Google Scholar] [CrossRef] [PubMed]
- Choukroun, J.; Diss, A.; Simonpieri, A.; Girard, M.-O.; Schoeffler, C.; Dohan, S.L.; Dohan, A.J.; Mouhyi, J.; Dohan, D.M. Platelet-rich fibrin (PRF): A second-generation platelet concentrate. Part IV: Clinical effects on tissue healing. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, e56–e60. [Google Scholar] [CrossRef] [PubMed]
- Choukroun, J.; Diss, A.; Simonpieri, A.; Girard, M.-O.; Schoeffler, C.; Dohan, S.L.; Dohan, A.J.; Mouhyi, J.; Dohan, D.M. Platelet-rich fibrin (PRF): A second-generation platelet concentrate. Part V: Histologic evaluations of PRF effects on bone allograft maturation in sinus lift. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, 299–303. [Google Scholar] [CrossRef]
- Dohan, D.M.; Choukroun, J.; Diss, A.; Dohan, S.L.; Dohan, A.J.; Mouhyi, J.; Gogly, B. Platelet-rich fibrin (PRF): A second-generation platelet concentrate. Part III: Leucocyte activation: A new feature for platelet concentrates? Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, e51–e55. [Google Scholar] [CrossRef]
- Dohan, D.M.; Choukroun, J.; Diss, A.; Dohan, S.L.; Dohan, A.J.; Mouhyi, J.; Gogly, B. Platelet-rich fibrin (PRF): A second-generation platelet concentrate. Part I: Technological concepts and evolution. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, e37–e44. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Yeh, Y.Y.; Chen, C.H.; Manga, Y.B.; Jheng, P.R.; Lu, C.X.; Chuang, E.Y. Effectiveness of treating segmental bone defects with a synergistic co-delivery approach with platelet-rich fibrin and tricalcium phosphate. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 129, 112364. [Google Scholar] [CrossRef] [PubMed]
- Borie, E.; Oliví, D.G.; Orsi, I.A.; Garlet, K.; Weber, B.; Beltrán, V.; Fuentes, R. Platelet-rich fibrin application in dentistry: A literature review. Int. J. Clin. Exp. Med. 2015, 8, 7922. [Google Scholar]
- Patel, G.K.; Gaekwad, S.S.; Gujjari, S.K.; SC, V.K. Platelet-Rich Fibrin in Regeneration of Intrabony Defects: A Randomized Controlled Trial. J. Periodontol. 2017, 88, 1192–1199. [Google Scholar] [CrossRef]
- Shivashankar, V.Y.; Johns, D.A.; Vidyanath, S.; Sam, G. Combination of platelet rich fibrin, hydroxyapatite and PRF membrane in the management of large inflammatory periapical lesion. J. Conserv. Dent. JCD 2013, 16, 261–264. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; You, X.; Xing, W.; Zhang, Z.; Zou, W. Paracrine and endocrine actions of bone—The functions of secretory proteins from osteoblasts, osteocytes, and osteoclasts. Bone Res. 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, E.S.; Mourão, C.; Leite, P.E.C.; Granjeiro, J.M.; Calasans-Maia, M.D.; Alves, G.G. The in vitro release of cytokines and growth factors from fibrin membranes produced through horizontal centrifugation. J. Biomed. Mater. Res. Part A 2018, 106, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.L.; Chen, Z.H.; Teng, Y.Y.; Liu, S.Y.; Jia, Y.; Zhang, K.W.; Sun, Z.L.; Wu, J.J.; Yuan, Z.D.; Feng, Y.; et al. The Smad Dependent TGF-β and BMP Signaling Pathway in Bone Remodeling and Therapies. Front. Mol. Biosci. 2021, 8, 593310. [Google Scholar] [CrossRef]
- McGowan, J.; Straus, S.; Moher, D.; Langlois, E.V.; O’Brien, K.K.; Horsley, T.; Aldcroft, A.; Zarin, W.; Garitty, C.M.; Hempel, S.; et al. Reporting scoping reviews-PRISMA ScR extension. J. Clin. Epidemiol. 2020, 123, 177–179. [Google Scholar] [CrossRef]
- Schneider, K.; Schwarz, M.; Burkholder, I.; Kopp-Schneider, A.; Edler, L.; Kinsner-Ovaskainen, A.; Hartung, T.; Hoffmann, S. “ToxRTool”, a new tool to assess the reliability of toxicological data. Toxicol. Lett. 2009, 189, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Al-Maawi, S.; Dohle, E.; Lim, J.; Weigl, P.; Teoh, S.H.; Sader, R.; Ghanaati, S. Biologization of Pcl-mesh using platelet rich fibrin (Prf) enhances its regenerative potential in vitro. Int. J. Mol. Sci. 2021, 22, 2159. [Google Scholar] [CrossRef]
- Al-Maawi, S.; Dohle, E.; Kretschmer, W.; Rutkowski, J.; Sader, R.; Ghanaati, S. A standardized g-force allows the preparation of similar platelet-rich fibrin qualities regardless of rotor angle. Tissue Eng. Part A 2022, 28, 353–365. [Google Scholar] [CrossRef]
- Bagio, D.A.; Julianto, I.; Margono, A.; Suprastiwi, E. Increased VEGF-A Expression of Human Dental Pulp Stem Cells (hDPSCs) Cultured with Advanced Platelet Rich Fibrin (A-PRF). Open Dent. J. 2021, 15. [Google Scholar] [CrossRef]
- Banyatworakul, P.; Osathanon, T.; Chumprasert, S.; Pavasant, P.; Pirarat, N. Responses of canine periodontal ligament cells to bubaline blood derived platelet rich fibrin in vitro. Sci. Rep. 2021, 11, 11409. [Google Scholar] [CrossRef]
- Bi, J.; Liu, Y.; Liu, X.-M.; Lei, S.; Chen, X. Platelet-rich Fibrin Improves the Osteo-/Odontogenic Differentiation of Stem Cells from Apical Papilla via the Extracellular Signal–regulated Protein Kinase Signaling Pathway. J. Endod. 2020, 46, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Blatt, S.; Thiem, D.G.; Kyyak, S.; Pabst, A.; Al-Nawas, B.; Kämmerer, P.W. Possible implications for improved osteogenesis? The combination of platelet-rich fibrin with different bone substitute materials. Front. Bioeng. Biotechnol. 2021, 9, 640053. [Google Scholar] [CrossRef]
- Chang, I.C.; Tsai, C.H.; Chang, Y.C. Platelet-rich fibrin modulates the expression of extracellular signal-regulated protein kinase and osteoprotegerin in human osteoblasts. J. Biomed. Mater. Res. Part A 2010, 95, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Zhao, Y.-H.; Zhao, Y.-J.; Liu, N.-X.; Lv, X.; Li, Q.; Chen, F.-M.; Zhang, M. Potential dental pulp revascularization and odonto-/osteogenic capacity of a novel transplant combined with dental pulp stem cells and platelet-rich fibrin. Cell Tissue Res. 2015, 361, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Feng, F.; Shi, F.; Huang, J.; Zhang, S.; Quan, Y.; Tu, T.; Liu, Y.; Wang, J.; Zhao, Y. Distinctive roles of Wnt signaling in chondrogenic differentiation of BMSCs under coupling of pressure and platelet-rich fibrin. Tissue Eng. Regen. Med. 2022, 19, 823–837. [Google Scholar] [CrossRef]
- Chi, H.; Song, X.; Song, C.; Zhao, W.; Chen, G.; Jiang, A.; Wang, X.; Yu, T.; Zheng, L.; Yan, J. Chitosan-gelatin scaffolds incorporating decellularized platelet-rich fibrin promote bone regeneration. ACS Biomater. Sci. Eng. 2019, 5, 5305–5315. [Google Scholar] [CrossRef]
- Clipet, F.; Tricot, S.; Alno, N.; Massot, M.; Solhi, H.; Cathelineau, G.; Perez, F.; De Mello, G.; Pellen-Mussi, P. In vitro effects of Choukroun’s platelet-rich fibrin conditioned medium on 3 different cell lines implicated in dental implantology. Implant. Dent. 2012, 21, 51–56. [Google Scholar] [CrossRef]
- Ehrenfest, D.M.D.; Doglioli, P.; de Peppo, G.M.; Del Corso, M.; Charrier, J.-B. Choukroun’s platelet-rich fibrin (PRF) stimulates in vitro proliferation and differentiation of human oral bone mesenchymal stem cell in a dose-dependent way. Arch. Oral Biol. 2010, 55, 185–194. [Google Scholar] [CrossRef]
- Dohle, E.; El Bagdadi, K.; Sader, R.; Choukroun, J.; James Kirkpatrick, C.; Ghanaati, S. Platelet-rich fibrin-based matrices to improve angiogenesis in an in vitro co-culture model for bone tissue engineering. J. Tissue Eng. Regen. Med. 2018, 12, 598–610. [Google Scholar] [CrossRef]
- Douglas, T.E.; Gassling, V.; Declercq, H.A.; Purcz, N.; Pamula, E.; Haugen, H.J.; Chasan, S.; de Mulder, E.L.; Jansen, J.A.; Leeuwenburgh, S.C. Enzymatically induced mineralization of platelet-rich fibrin. J. Biomed. Mater. Res. Part A 2012, 100, 1335–1346. [Google Scholar] [CrossRef]
- Duan, X.; Lin, Z.; Lin, X.; Wang, Z.; Wu, Y.; Ji, M.; Lu, W.; Wang, X.; Zhang, D. Study of platelet-rich fibrin combined with rat periodontal ligament stem cells in periodontal tissue regeneration. J. Cell. Mol. Med. 2018, 22, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfest, D.M.D.; Diss, A.; Odin, G.; Doglioli, P.; Hippolyte, M.-P.; Charrier, J.-B. In vitro effects of Choukroun’s PRF (platelet-rich fibrin) on human gingival fibroblasts, dermal prekeratinocytes, preadipocytes, and maxillofacial osteoblasts in primary cultures. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 108, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilnejad, A.; Ardakani, M.T.; Shokri, M.; Khou, N.H.; Kamani, M. Comparative evaluation of the effect of two platelet concentrates (a-PRF and L-PRF) on the cellular activity of pre-osteoblastic MG-63 cell line: An in vitro study. J. Dent. 2023, 24, 235. [Google Scholar]
- Fernández-Medina, T.; Vaquette, C.; Ivanovski, S. Systematic comparison of the effect of four clinical-grade platelet rich hemoderivatives on osteoblast behaviour. Int. J. Mol. Sci. 2019, 20, 6243. [Google Scholar] [CrossRef] [PubMed]
- Gassling, V.L.; Açil, Y.; Springer, I.N.; Hubert, N.; Wiltfang, J. Platelet-rich plasma and platelet-rich fibrin in human cell culture. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 108, 48–55. [Google Scholar] [CrossRef]
- Gassling, V.; Douglas, T.; Warnke, P.H.; Açil, Y.; Wiltfang, J.; Becker, S.T. Platelet-rich fibrin membranes as scaffolds for periosteal tissue engineering. Clin. Oral Implant. Res. 2010, 21, 543–549. [Google Scholar] [CrossRef]
- Gassling, V.; Douglas, T.E.; Purcz, N.; Schaubroeck, D.; Balcaen, L.; Bliznuk, V.; Declercq, H.A.; Vanhaecke, F.; Dubruel, P. Magnesium-enhanced enzymatically mineralized platelet-rich fibrin for bone regeneration applications. Biomed. Mater. 2013, 8, 055001. [Google Scholar] [CrossRef]
- Gassling, V.; Hedderich, J.; Açil, Y.; Purcz, N.; Wiltfang, J.; Douglas, T. Comparison of platelet rich fibrin and collagen as osteoblast-seeded scaffolds for bone tissue engineering applications. Clin. Oral Implant. Res. 2013, 24, 320–328. [Google Scholar] [CrossRef]
- Girija, K.; Kavitha, M. Comparative evaluation of platelet-rich fibrin, platelet-rich fibrin+ 50 wt% nanohydroxyapatite, platelet-rich fibrin+ 50 wt% dentin chips on odontoblastic differentiation-An in vitro study-part 2. J. Conserv. Dent. JCD 2020, 23, 354. [Google Scholar] [CrossRef]
- He, L.; Lin, Y.; Hu, X.; Zhang, Y.; Wu, H. A comparative study of platelet-rich fibrin (PRF) and platelet-rich plasma (PRP) on the effect of proliferation and differentiation of rat osteoblasts in vitro. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 108, 707–713. [Google Scholar] [CrossRef]
- Hong, S.; Chen, W.; Jiang, B. A Comparative Evaluation of Concentrated Growth Factor and Platelet-rich Fibrin on the Proliferation, Migration, and Differentiation of Human Stem Cells of the Apical Papilla. J. Endod. 2018, 44, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.M.; Yang, S.F.; Zhao, J.H.; Chang, Y.C. Platelet-rich fibrin increases proliferation and differentiation of human dental pulp cells. J. Endod. 2010, 36, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Irastorza, I.; Luzuriaga, J.; Martinez-Conde, R.; Ibarretxe, G.; Unda, F. Adhesion, integration and osteogenesis of human dental pulp stem cells on biomimetic implant surfaces combined with plasma derived products. Eur. Cells Mater. 2019, 38, 201–214. [Google Scholar] [CrossRef]
- Isobe, K.; Suzuki, M.; Watanabe, T.; Kitamura, Y.; Suzuki, T.; Kawabata, H.; Nakamura, M.; Okudera, T.; Okudera, H.; Uematsu, K.; et al. Platelet-rich fibrin prepared from stored whole-blood samples. Int. J. Implant Dent. 2017, 3, 6. [Google Scholar] [CrossRef]
- Ji, B.; Sheng, L.; Chen, G.; Guo, S.; Xie, L.; Yang, B.; Guo, W.; Tian, W. The combination use of platelet-rich fibrin and treated dentin matrix for tooth root regeneration by cell homing. Tissue Eng. Part A 2015, 21, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Jeon, S.H.; Park, J.Y.; Chung, J.H.; Choung, Y.H.; Choung, H.W.; Kim, E.S.; Choung, P.H. Platelet-rich fibrin is a Bioscaffold and reservoir of growth factors for tissue regeneration. Tissue Eng. Part A 2011, 17, 349–359. [Google Scholar] [CrossRef]
- Kardos, D.; Hornyák, I.; Simon, M.; Hinsenkamp, A.; Marschall, B.; Várdai, R.; Kállay-Menyhárd, A.; Pinke, B.; Mészáros, L.; Kuten, O.; et al. Biological and Mechanical Properties of Platelet-Rich Fibrin Membranes after Thermal Manipulation and Preparation in a Single-Syringe Closed System. Int. J. Mol. Sci. 2018, 19, 3433. [Google Scholar] [CrossRef]
- Kim, J.; Ha, Y.; Kang, N.H. Effects of Growth Factors From Platelet-Rich Fibrin on the Bone Regeneration. J. Craniofacial Surg. 2017, 28, 860–865. [Google Scholar] [CrossRef]
- Kim, J.H.; Woo, S.M.; Choi, N.K.; Kim, W.J.; Kim, S.M.; Jung, J.Y. Effect of Platelet-rich Fibrin on Odontoblastic Differentiation in Human Dental Pulp Cells Exposed to Lipopolysaccharide. J. Endod. 2017, 43, 433–438. [Google Scholar] [CrossRef]
- Kosmidis, K.; Ehsan, K.; Pitzurra, L.; Loos, B.; Jansen, I. An in vitro study into three different PRF preparations for osteogenesis potential. J. Periodontal Res. 2023, 58, 483–492. [Google Scholar] [CrossRef]
- Koyanagi, M.; Fujioka-Kobayashi, M.; Yoneyama, Y.; Inada, R.; Satomi, T. Regenerative Potential of Solid Bone Marrow Aspirate Concentrate Compared with Platelet-Rich Fibrin. Tissue Eng. Part A 2022, 28, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Kyyak, S.; Blatt, S.; Pabst, A.; Thiem, D.; Al-Nawas, B.; Kämmerer, P.W. Combination of an allogenic and a xenogenic bone substitute material with injectable platelet-rich fibrin—A comparative in vitro study. J. Biomater. Appl. 2020, 35, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Kyyak, S.; Blatt, S.; Schiegnitz, E.; Heimes, D.; Staedt, H.; Thiem, D.G.E.; Sagheb, K.; Al-Nawas, B.; Kämmerer, P.W. Activation of Human Osteoblasts via Different Bovine Bone Substitute Materials With and Without Injectable Platelet Rich Fibrin in vitro. Front. Bioeng. Biotechnol. 2021, 9, 599224. [Google Scholar] [CrossRef]
- Li, Q.; Pan, S.; Dangaria, S.J.; Gopinathan, G.; Kolokythas, A.; Chu, S.; Geng, Y.; Zhou, Y.; Luan, X. Platelet-rich fibrin promotes periodontal regeneration and enhances alveolar bone augmentation. BioMed Res. Int. 2013, 2013, 638043. [Google Scholar] [CrossRef]
- Li, Q.; Reed, D.A.; Min, L.; Gopinathan, G.; Li, S.; Dangaria, S.J.; Li, L.; Geng, Y.; Galang, M.T.; Gajendrareddy, P.; et al. Lyophilized platelet-rich fibrin (PRF) promotes craniofacial bone regeneration through Runx2. Int. J. Mol. Sci. 2014, 15, 8509–8525. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, H.; Zhang, Z.; Yan, Z.; Lv, H.; Zhang, Y.; Wu, B. Platelet-rich fibrin exudate promotes the proliferation and osteogenic differentiation of human periodontal ligament cells in vitro. Mol. Med. Rep. 2018, 18, 4477–4485. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, J.; Wu, J.; Du, X.; Jing, W.; Liu, L. Roles of PRF and IGF-1 in promoting alveolar osteoblast growth and proliferation and molecular mechanism. Int. J. Clin. Exp. Pathol. 2018, 11, 3294–3301. [Google Scholar]
- Liang, Z.; Huang, D.; Nong, W.; Mo, J.; Zhu, D.; Wang, M.; Chen, M.; Wei, C.; Li, H. Advanced-platelet-rich fibrin extract promotes adipogenic and osteogenic differentiation of human adipose-derived stem cells in a dose-dependent manner in vitro. Tissue Cell 2021, 71, 101506. [Google Scholar] [CrossRef]
- Liu, Z.; Jin, H.; Xie, Q.; Jiang, Z.; Guo, S.; Li, Y.; Zhang, B. Controlled Release Strategies for the Combination of Fresh and Lyophilized Platelet-Rich Fibrin on Bone Tissue Regeneration. BioMed Res. Int. 2019, 2019, 4923767. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yin, M.; Li, Y.; Wang, J.; Da, J.; Liu, Z.; Zhang, K.; Liu, L.; Zhang, W.; Wang, P. Genipin modified lyophilized platelet-rich fibrin scaffold for sustained release of growth factors to promote bone regeneration. Front. Physiol. 2022, 13, 1007692. [Google Scholar] [CrossRef]
- Lo Monaco, M.; Gervois, P.; Beaumont, J.; Clegg, P.; Bronckaers, A.; Vandeweerd, J.M.; Lambrichts, I. Therapeutic Potential of Dental Pulp Stem Cells and Leukocyte- and Platelet-Rich Fibrin for Osteoarthritis. Cells 2020, 9, 980. [Google Scholar] [CrossRef]
- Marchetti, E.; Mancini, L.; Bernardi, S.; Bianchi, S.; Cristiano, L.; Torge, D.; Marzo, G.; Macchiarelli, G. Evaluation of Different Autologous Platelet Concentrate Biomaterials: Morphological and Biological Comparisons and Considerations. Materials 2020, 13, 2282. [Google Scholar] [CrossRef]
- Moradian, H.; Rafiee, A.; Ayatollahi, M. Design and Fabrication of a Novel Transplant Combined with Human Bone Marrow Mesenchymal Stem Cells and Platelet-rich Fibrin: New Horizons for Periodontal Tissue Regeneration after Dental Trauma. Iran. J. Pharm. Res. 2017, 16, 1370–1378. [Google Scholar]
- Nguyen, M.; Nguyen, T.T.; Tran, H.L.B.; Tran, D.N.; Ngo, L.T.Q.; Huynh, N.C. Effects of advanced platelet-rich fibrin combined with xenogenic bone on human periodontal ligament stem cells. Clin. Exp. Dent. Res. 2022, 8, 875–882. [Google Scholar] [CrossRef]
- Nie, J.; Zhang, S.; Wu, P.; Liu, Y.; Su, Y. Electrospinning With Lyophilized Platelet-Rich Fibrin Has the Potential to Enhance the Proliferation and Osteogenesis of MC3T3-E1 Cells. Front. Bioeng. Biotechnol. 2020, 8, 595579. [Google Scholar] [CrossRef]
- Nugraha, A.P.; Narmada, I.B.; Ernawati, D.S.; Dinaryanti, A.; Hendrianto, E.; Ihsan, I.S.; Riawan, W.; Rantam, F.A. Osteogenic potential of gingival stromal progenitor cells cultured in platelet rich fibrin is predicted by core-binding factor subunit-α1/Sox9 expression ratio (in vitro). F1000Research 2018, 7, 1134. [Google Scholar] [CrossRef]
- Nugraha, A.P.; Narmada, I.B.; Ernawati, D.S.; Dinaryanti, A.; Hendrianto, E.; Ihsan, I.S.; Riawan, W.; Rantam, F.A. In vitro bone sialoprotein-I expression in combined gingival stromal progenitor cells and platelet rich fibrin during osteogenic differentiation. Trop. J. Pharm. Res. 2018, 17, 2341–2345. [Google Scholar] [CrossRef]
- Nugraha, A.P.; Narmada, I.B.; Ernawati, D.S.; Dinaryanti, A.; Hendrianto, E.; Riawan, W.; Rantam, F.A. Bone alkaline phosphatase and osteocalcin expression of rat’s Gingival mesenchymal stem cells cultured in platelet-rich fibrin for bone remodeling (in vitro study). Eur. J. Dent. 2018, 12, 566–573. [Google Scholar] [CrossRef]
- Nugraha, A.; Narmada, I.B.; Ernawatı, D.S.; Dınaryantı, A.; Hendrianto, E.; Ihsan, I.; Riawan, W.; Rantam, F. The Aggrecan Expression Post Platelet Rich Fibrin Administration in Gingival Medicinal Signaling Cells in Wistar Rats (Rattus novergicus) During the Early Osteogenic Differentiation (In Vitro). Kafkas Univ. Vet. Fak. Derg. 2019, 25, 421–425. [Google Scholar]
- Rastegar, A.; Mahmoodi, M.; Mirjalili, M.; Nasirizadeh, N. Platelet-rich fibrin-loaded PCL/chitosan core-shell fibers scaffold for enhanced osteogenic differentiation of mesenchymal stem cells. Carbohydr. Polym. 2021, 269, 118351. [Google Scholar] [CrossRef]
- Shah, R.; Thomas, R.; Gowda, T.M.; Baron, T.K.A.; Vemanaradhya, G.G.; Bhagat, S. In Vitro Evaluation of Osteoblast Response to the Effect of Injectable Platelet-rich Fibrin Coating on Titanium Disks. J. Contemp. Dent. Pract. 2021, 22, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lin, K.; He, S.; Wang, C.; Zhang, S.; Li, D.; Wang, J.; Cao, T.; Bi, L.; Pei, G. Nano-biphasic calcium phosphate/polyvinyl alcohol composites with enhanced bioactivity for bone repair via low-temperature three-dimensional printing and loading with platelet-rich fibrin. Int. J. Nanomed. 2018, 13, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Steller, D.; Herbst, N.; Pries, R.; Juhl, D.; Hakim, S.G. Positive impact of Platelet-rich plasma and Platelet-rich fibrin on viability, migration and proliferation of osteoblasts and fibroblasts treated with zoledronic acid. Sci. Rep. 2019, 9, 8310. [Google Scholar] [CrossRef]
- Steller, D.; Herbst, N.; Pries, R.; Juhl, D.; Klinger, M.; Hakim, S.G. Impacts of platelet-rich fibrin and platelet-rich plasma on primary osteoblast adhesion onto titanium implants in a bisphosphonate in vitro model. J. Oral Pathol. Med. 2019, 48, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Zhang, H.; Yao, J.; Yang, L.; Zhang, X.; Li, L.; Wang, J.; Li, M.; Liu, Z. 3D printing of ‘green’thermo-sensitive chitosan-hydroxyapatite bone scaffold based on lyophilized platelet-rich fibrin. Biomed. Mater. 2023, 18, 025022. [Google Scholar] [CrossRef] [PubMed]
- Thanasrisuebwong, P.; Kiattavorncharoen, S.; Surarit, R.; Phruksaniyom, C.; Ruangsawasdi, N. Red and Yellow Injectable Platelet-Rich Fibrin Demonstrated Differential Effects on Periodontal Ligament Stem Cell Proliferation, Migration, and Osteogenic Differentiation. Int. J. Mol. Sci. 2020, 21, 5153. [Google Scholar] [CrossRef] [PubMed]
- Verboket, R.; Herrera-Vizcaíno, C.; Thorwart, K.; Booms, P.; Bellen, M.; Al-Maawi, S.; Sader, R.; Marzi, I.; Henrich, D.; Ghanaati, S. Influence of concentration and preparation of platelet rich fibrin on human bone marrow mononuclear cells (in vitro). Platelets 2019, 30, 861–870. [Google Scholar] [CrossRef]
- Wang, Z.; Weng, Y.; Lu, S.; Zong, C.; Qiu, J.; Liu, Y.; Liu, B. Osteoblastic mesenchymal stem cell sheet combined with Choukroun platelet-rich fibrin induces bone formation at an ectopic site. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 103, 1204–1216. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Choukroun, J.; Ghanaati, S.; Miron, R.J. Effects of an injectable platelet-rich fibrin on osteoblast behavior and bone tissue formation in comparison to platelet-rich plasma. Platelets 2018, 29, 48–55. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.; Liu, Y.; Yu, J.; Sun, X.; Wang, L.; Zhou, Y. Effects of platelet-rich fibrin on osteogenic differentiation of Schneiderian membrane derived mesenchymal stem cells and bone formation in maxillary sinus. Cell Commun. Signal. 2022, 20, 88. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; He, X.; Li, S.; Pan, H.; Yin, L. Injectable platelet-rich fibrin positively regulates osteogenic differentiation of stem cells from implant hole via the ERK1/2 pathway. Platelets 2023, 34, 2159020. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.C.; Wang, C.Y.; Jang, J.S.; Lee, C.H.; Wu, J.L. Large-Pore Platelet-Rich Fibrin with a Mg Ring to Allow MC3T3-E1 Preosteoblast Migration and to Improve Osteogenic Ability for Bone Defect Repair. Int. J. Mol. Sci. 2021, 22, 4022. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.M.; Kim, W.J.; Lim, H.S.; Choi, N.K.; Kim, S.H.; Kim, S.M.; Jung, J.Y. Combination of Mineral Trioxide Aggregate and Platelet-rich Fibrin Promotes the Odontoblastic Differentiation and Mineralization of Human Dental Pulp Cells via BMP/Smad Signaling Pathway. J. Endod. 2016, 42, 82–88. [Google Scholar] [CrossRef]
- Wu, C.L.; Lee, S.S.; Tsai, C.H.; Lu, K.H.; Zhao, J.H.; Chang, Y.C. Platelet-rich fibrin increases cell attachment, proliferation and collagen-related protein expression of human osteoblasts. Aust. Dent. J. 2012, 57, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, W.; Lu, J.; Hao, Y.; Lv, C.; Cao, C.; Zou, D. Platelet-rich fibrin as a scaffold in combination with either deciduous or permanent dental pulp cells for bone tissue engineering. Int. J. Clin. Exp. Med. 2016, 9, 15177–15184. [Google Scholar]
- Yu, S.; Bd, Y.T.; Bd, Y.W.; Bd, M.F.; BMed, S.L.; BMed, G.T.; BMed, Z.Y.; Miron, R.J.; Zhang, Y.; Yang, Z.; et al. Early tissue and healing responses after maxillary sinus augmentation using horizontal platelet rich fibrin bone blocks. BMC Oral Health 2023, 23, 589. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, Y.; Xue, Y.; Shi, J.; Zhang, X.; Liu, Y.; Midgley, A.C.; Wang, S. Multifunctional Triple-Layered Composite Scaffolds Combining Platelet-Rich Fibrin Promote Bone Regeneration. ACS Biomater. Sci. Eng. 2019, 5, 6691–6702. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Lin, X.; Liu, X. Platelet-rich Fibrin Promotes the Proliferation and Osteo-/odontoblastic Differentiation of Human Dental Pulp Stem Cells. Curr. Stem Cell Res. Ther. 2023, 18, 560–567. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Zhang, M.; Liu, N.X.; Lv, X.; Zhang, J.; Chen, F.M.; Chen, Y.J. The combined use of cell sheet fragments of periodontal ligament stem cells and platelet-rich fibrin granules for avulsed tooth reimplantation. Biomaterials 2013, 34, 5506–5520. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, L.; Qin, J.; Sun, X.; Yang, T.; Ni, Y.; Zhou, Y. New Biodegradable Implant Material Containing Hydrogel with Growth Factors of Lyophilized PRF in Combination with an nHA/PLGA Scaffold. J. Hard Tissue Biol. 2015, 24, 54–60. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, X.; Zhao, Q.; Chai, J.; Zhang, Y. Liquid platelet-rich fibrin promotes the regenerative potential of human periodontal ligament cells. Oral Dis. 2020, 26, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Dohle, E.; Kolbe, M.; Kirkpatrick, C.J. Outgrowth endothelial cells: Sources, characteristics and potential applications in tissue engineering and regenerative medicine. In Bioreactor Systems for Tissue Engineering II; Advances in Biochemical Engineering / Biotechnology; Springer: Berlin/Heidelberg, Germany, 2010; Volume 123, pp. 201–217. [Google Scholar] [CrossRef]
- Niu, B.; Li, B.; Gu, Y.; Shen, X.; Liu, Y.; Chen, L. In vitro evaluation of electrospun silk fibroin/nano-hydroxyapatite/BMP-2 scaffolds for bone regeneration. J. Biomater. Sci. Polym. Ed. 2017, 28, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, K.; Kottuppallil, G.; Sekar, V. Comparative evaluation of radiopacity and cytotoxicity of platelet-rich fibrin, platelet-rich fibrin + 50wt% nano-hydroxyapatite, platelet-rich fibrin + 50wt% dentin chips: An in vitro study. J. Conserv. Dent. JCD 2019, 22, 28–33. [Google Scholar]
- Chen, G.; Deng, C.; Li, Y.-P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Polymeri, A.; Giannobile, W.V.; Kaigler, D. Bone Marrow Stromal Stem Cells in Tissue Engineering and Regenerative Medicine. Horm. Metab. Res. 2016, 48, 700–713. [Google Scholar] [CrossRef]
- Canceill, T.; Jourdan, G.; Kémoun, P.; Guissard, C.; Monsef, Y.A.; Bourdens, M.; Chaput, B.; Cavalie, S.; Casteilla, L.; Planat-Bénard, V.; et al. Characterization and Safety Profile of a New Combined Advanced Therapeutic Medical Product Platelet Lysate-Based Fibrin Hydrogel for Mesenchymal Stromal Cell Local Delivery in Regenerative Medicine. Int. J. Mol. Sci. 2023, 24, 2206. [Google Scholar] [CrossRef]
- Zanetti, A.S.; Sabliov, C.; Gimble, J.M.; Hayes, D.J. Human adipose-derived stem cells and three-dimensional scaffold constructs: A review of the biomaterials and models currently used for bone regeneration. J. Biomed. Mater. Res. Part B Appl. Biomater. 2013, 101, 187–199. [Google Scholar] [CrossRef]
- Lai, F.; Kakudo, N.; Morimoto, N.; Taketani, S.; Hara, T.; Ogawa, T.; Kusumoto, K. Platelet-rich plasma enhances the proliferation of human adipose stem cells through multiple signaling pathways. Stem Cell Res. Ther. 2018, 9, 107. [Google Scholar] [CrossRef]
- Lei, F.; Li, M.; Lin, T.; Zhou, H.; Wang, F.; Su, X. Treatment of inflammatory bone loss in periodontitis by stem cell-derived exosomes. Acta Biomater. 2022, 141, 333–343. [Google Scholar] [CrossRef]
- Cho, Y.S.; Hwang, K.G.; Jun, S.H.; Tallarico, M.; Kwon, A.M.; Park, C.J. Radiologic comparative analysis between saline and platelet-rich fibrin filling after hydraulic transcrestal sinus lifting without adjunctive bone graft: A randomized controlled trial. Clin. Oral Implant. Res. 2020, 31, 1087–1093. [Google Scholar] [CrossRef]
- Chang, Y.C.; Zhao, J.H. Effects of platelet-rich fibrin on human periodontal ligament fibroblasts and application for periodontal infrabony defects. Aust. Dent. J. 2011, 56, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Lee, J.K.; Han, J.; Lee, B.; Kang, S.; Hwang, K.T.; Park, Y.S.; Kim, T.H. Identification and characterization of human bone-derived cells. Biochem. Biophys. Res. Commun. 2018, 495, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, M.; Wang, S.; Xiao, Z.; Xiong, Y.; Wang, G. Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation. Front. Cell Dev. Biol. 2020, 8, 601224. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.M.; Li, W.T.; Xia, X.M.; Wang, F.; MacDougall, M.; Chen, S. Dentine sialophosphoprotein signal in dentineogenesis and dentine regeneration. Eur. Cells Mater. 2021, 42, 43–62. [Google Scholar] [CrossRef]
- Wang, Z.; Han, L.; Sun, T.; Wang, W.; Li, X.; Wu, B. Preparation and effect of lyophilized platelet-rich fibrin on the osteogenic potential of bone marrow mesenchymal stem cells in vitro and in vivo. Heliyon 2019, 5, e02739. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nakayamada, S.; Okada, Y. Osteoblasts and osteoclasts in bone remodeling and inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 325–328. [Google Scholar] [CrossRef]
- Lourenço, E.S.; Alves, G.G.; de Lima Barbosa, R.; Spiegel, C.N.; de Mello-Machado, R.C.; Al-Maawi, S.; Ghanaati, S.; de Almeida Barros Mourão, C.F. Effects of rotor angle and time after centrifugation on the biological in vitro properties of platelet rich fibrin membranes. J. Biomed. Mater. Res. Part B Appl. Biomater. 2021, 109, 60–68. [Google Scholar] [CrossRef]
- Jann, J.; Gascon, S.; Roux, S.; Faucheux, N. Influence of the TGF-β Superfamily on Osteoclasts/Osteoblasts Balance in Physiological and Pathological Bone Conditions. Int. J. Mol. Sci. 2020, 21, 7597. [Google Scholar] [CrossRef]
- Ghanaati, S.; Herrera-Vizcaino, C.; Al-Maawi, S.; Lorenz, J.; Miron, R.J.; Nelson, K.; Schwarz, F.; Choukroun, J.; Sader, R. Fifteen years of platelet rich fibrin in dentistry and oromaxillofacial surgery: How high is the level of scientific evidence? J. Oral Implantol. 2018, 44, 471–492. [Google Scholar] [CrossRef]
- Reis, N.T.d.A.; João Lucas Carvalho, P.; Paranhos, L.R.; Bernardino, I.d.M.; Moura, C.C.G.; Irie, M.S.; Soares, P.B.F. Use of platelet-rich fibrin for bone repair: A systematic review and meta-analysis of preclinical studies. Braz. Oral Res. 2022, 36. [Google Scholar] [CrossRef]
- Farshidfar, N.; Jafarpour, D.; Firoozi, P.; Sahmeddini, S.; Hamedani, S.; de Souza, R.F.; Tayebi, L. The application of injectable platelet-rich fibrin in regenerative dentistry: A systematic scoping review of In vitro and In vivo studies. Jpn. Dent. Sci. Rev. 2022, 58, 89–123. [Google Scholar] [CrossRef] [PubMed]
- Strauss, F.-J.; Nasirzade, J.; Kargarpoor, Z.; Stähli, A.; Gruber, R. Effect of platelet-rich fibrin on cell proliferation, migration, differentiation, inflammation, and osteoclastogenesis: A systematic review of in vitro studies. Clin. Oral Investig. 2020, 24, 569–584. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| DATABASE | Search Key |
|---|---|
| PubMed (https://pubmed.ncbi.nlm.nih.gov/, accessed on 14 August 2023) | (PRF OR “platelet rich fibrin” OR L-PRF OR i-PRF OR “Sticky bone” OR “concentrated growth factors” OR CGF) AND (Bone OR osteoblast* OR MSC OR “mesenchymal stem cell” OR “bone marrow” OR “Bone and bones” [mh] OR “bone cell” OR preosteoblast* OR Skeleton) AND (“in vitro” OR In Vitro Techniques” [mh] OR “Cell Lineage” [mh] OR “Cells, Cultured” [mh]). |
| Web of Science (https://www.webofscience.com/, accessed on 14 August 2023) | ((PRF OR “platelet rich fibrin” OR L-PRF OR i-PRF OR “Sticky bone” OR “concentrated growth factors” OR CGF) AND (Bone OR osteoblast OR MSC OR “mesenchymal stem cell” OR “bone marrow” OR “Bone and bones” OR “bone cell” OR preosteoblast OR PDL OR “periodontal ligament OR mineralization) AND (“in vitro” OR “In Vitro Techniques” OR “Cell Lineage” OR “Cells, Cultured”)) Refined by DOCTYPE: (ARTICLE). |
| Scopus (https://0-www-scopus-com.brum.beds.ac.uk/search/form.uri?display=basic, accessed on 14 August 2023) | TITLE-ABS-KEY (prf OR “platelet rich fibrin” OR l-prf OR i-prf OR “Sticky bone” OR “concentrated growth factors” OR cgf) AND TITLE-ABS-KEY (bone OR osteoblast OR MSC OR “mesenchymal stem cell” OR “bone marrow” OR “Bone and bones” OR “bone cell” OR preosteoblast OR pdl OR “periodontal ligament OR mineralization) AND TITLE-ABS-KEY ((“in vitro” OR “In Vitro Techniques” OR “Cell Lineage” OR “Cell Culture”) AND (LIMIT TO (DOCTYPE, “ar”))). |
| Publication | Group I: Test Substance Identification (4) | Group II: Test System Characterization (3) | Group III: Study Design Description (6) | Group IV: Study Results Documentation (3) | Group V: Plausibility of Study Design and Data (2) | Total (18) |
|---|---|---|---|---|---|---|
| Al-Maawi et al., 2021 [17] | 4 | 3 | 6 | 3 | 2 | 18 |
| Al-Maawi et al., 2022 [18] | 4 | 3 | 6 | 3 | 2 | 18 |
| Bagio et al., 2021 [19] | 3 | 3 | 6 | 3 | 2 | 17 |
| Banyatworakul et al., 2021 [20] | 4 | 3 | 6 | 3 | 2 | 18 |
| Bi et al., 2020 [21] | 4 | 3 | 6 | 3 | 2 | 18 |
| Blatt et al., 2021 [22] | 4 | 3 | 5 | 3 | 2 | 17 |
| Chang, Tsai, and Chang, 2010 [23] | 4 | 2 | 5 | 3 | 2 | 16 |
| Chen et al., 2015 [24] | 4 | 3 | 6 | 3 | 2 | 18 |
| Cheng et al., 2022 [25] | 4 | 3 | 5 | 3 | 2 | 17 |
| Chi et al., 2019 [26] | 4 | 3 | 5 | 3 | 2 | 17 |
| Clipet et al., 2012 [27] | 4 | 3 | 6 | 3 | 2 | 18 |
| Dohan Ehrenfest et al., 2010 [28] | 4 | 1 | 5 | 3 | 2 | 15 |
| Dohle et al., 2018 [29] | 4 | 3 | 6 | 3 | 2 | 18 |
| Douglas et al., 2012 [30] | 4 | 1 | 5 | 3 | 2 | 15 |
| Duan et al., 2018 [31] | 4 | 1 | 5 | 3 | 2 | 15 |
| Ehrenfest et al., 2009 [32] | 4 | 1 | 5 | 3 | 2 | 15 |
| Esmaeilnejad et al., 2022 [33] | 4 | 3 | 6 | 3 | 2 | 18 |
| Fernandez-Medina et al., 2019 [34] | 4 | 3 | 6 | 3 | 2 | 18 |
| Gassling et al., 2009 [35] | 4 | 1 | 5 | 3 | 2 | 15 |
| Gassling et al., 2010 [36] | 4 | 3 | 5 | 3 | 2 | 17 |
| Gassling et al., 2013 [37] | 4 | 3 | 6 | 3 | 2 | 18 |
| Gassling et al., 2013 [38] | 4 | 3 | 5 | 3 | 2 | 17 |
| Girija and Kavitha, 2020 [39] | 2 | 3 | 5 | 2 | 1 | 13 |
| He et al., 2009 [40] | 4 | 3 | 5 | 3 | 2 | 17 |
| Hong, Chen, and Jiang, 2018 [41] | 4 | 3 | 6 | 3 | 2 | 18 |
| Huang et al., 2010 [42] | 4 | 3 | 6 | 3 | 2 | 18 |
| Irastorza et al., 2019 [43] | 4 | 3 | 6 | 3 | 2 | 18 |
| Isobe et al., 2017 [44] | 4 | 1 | 6 | 3 | 2 | 16 |
| Ji et al., 2015 [45] | 4 | 3 | 6 | 3 | 2 | 18 |
| Kang et al., 2011 [46] | 4 | 3 | 6 | 3 | 2 | 18 |
| Kardos et al., 2018 [47] | 4 | 2 | 5 | 3 | 2 | 16 |
| Kim et al., 2017 [48] | 4 | 2 | 5 | 3 | 2 | 16 |
| Kim et al., 2017 [49] | 4 | 3 | 5 | 3 | 2 | 17 |
| Kosmidis et al., 2023 [50] | 4 | 3 | 6 | 3 | 2 | 18 |
| Koyanagi et al., 2022 [51] | 4 | 3 | 5 | 3 | 2 | 17 |
| Kyyak et al., 2020 [52] | 4 | 3 | 6 | 3 | 2 | 18 |
| Kyyak et al., 2021 [53] | 4 | 3 | 6 | 3 | 2 | 18 |
| Li et al., 2013 [54] | 4 | 2 | 6 | 3 | 2 | 17 |
| Li et al., 2014 [55] | 4 | 3 | 6 | 3 | 2 | 18 |
| Li et al., 2018 [56] | 4 | 3 | 6 | 3 | 2 | 18 |
| Li et al., 2018 [57] | 4 | 3 | 6 | 3 | 2 | 18 |
| Liang et al., 2021 [58] | 3 | 3 | 6 | 3 | 2 | 17 |
| Liu et al., 2019 [59] | 4 | 3 | 6 | 3 | 2 | 18 |
| Liu et al., 2022 [60] | 4 | 3 | 6 | 3 | 2 | 18 |
| Lo Monaco et al. 2020 [61] | 4 | 3 | 6 | 3 | 2 | 18 |
| Marchetti, 2020 [62] | 4 | 3 | 6 | 3 | 2 | 18 |
| Moradian et al., 2017 [63] | 4 | 2 | 5 | 3 | 2 | 16 |
| Nguyen et al., 2022 [64] | 4 | 3 | 6 | 3 | 2 | 18 |
| Nie et al., 2020 [65] | 4 | 3 | 6 | 3 | 2 | 18 |
| Nugraha et al., 2018 [66] | 4 | 2 | 5 | 3 | 2 | 16 |
| Nugraha et al., 2018 [67] | 4 | 2 | 5 | 3 | 2 | 16 |
| Nugraha et al., 2018 [68] | 4 | 2 | 5 | 3 | 2 | 16 |
| Nugraha et al., 2019 [69] | 4 | 2 | 5 | 3 | 2 | 16 |
| Rastegar et al., 2021 [70] | 4 | 3 | 6 | 3 | 2 | 18 |
| Shah et al., 2021 [71] | 4 | 2 | 4 | 3 | 2 | 15 |
| Song et al., 2018 [72] | 4 | 3 | 6 | 3 | 2 | 18 |
| Steller et al., 2019 [73] | 4 | 3 | 6 | 3 | 2 | 18 |
| Steller et al., 2019 [74] | 4 | 3 | 6 | 3 | 2 | 18 |
| Sui et al., 2023 [75] | 4 | 3 | 6 | 3 | 2 | 18 |
| Thanasrisuebwong et al., 2020 [76] | 4 | 2 | 6 | 3 | 2 | 16 |
| Verboket et al., 2019 [77] | 4 | 2 | 6 | 3 | 2 | 17 |
| Wang et al., 2015 [78] | 4 | 3 | 6 | 3 | 2 | 18 |
| Wang et al., 2018 [79] | 4 | 3 | 6 | 3 | 2 | 18 |
| Wang et al., 2022 [80] | 4 | 3 | 5 | 3 | 2 | 17 |
| Wang et al., 2023 [81] | 4 | 3 | 5 | 3 | 2 | 17 |
| Wong et al., 2021 [7] | 4 | 3 | 6 | 3 | 2 | 18 |
| Wong et al., 2021 [82] | 4 | 3 | 6 | 3 | 2 | 18 |
| Woo et al., 2016 [83] | 4 | 1 | 5 | 3 | 2 | 15 |
| Wu et al., 2012 [84] | 4 | 3 | 5 | 3 | 2 | 16 |
| Yu et al., 2016 [85] Yu et al., 2023 [86] | 4 | 3 | 6 | 3 | 2 | 18 |
| Zhang et al., 2019 [87] | 4 | 3 | 6 | 3 | 2 | 18 |
| Zhang et al., 2023 [88] | 4 | 3 | 5 | 3 | 2 | 17 |
| Zhao et al., 2013 [89] | 4 | 2 | 5 | 3 | 2 | 16 |
| Zheng et al., 2015 [90] | 4 | 1 | 5 | 3 | 2 | 15 |
| Zheng et al., 2020 [91] | 4 | 3 | 6 | 3 | 2 | 18 |
| Publication | PRF Protocol | Mineralizing Cell Type | Exposure Time | Exposure Method | Biological Parameters | Results |
|---|---|---|---|---|---|---|
| Al-Maawi et al., 2021 [17] | A-PRF | Primary human osteoblasts | After 3 and 7 days | Eluate | VEGF; TGF-β1; PDGF, OPG, IL-8; OPN; ALP activity. | PRF produced according to the low-speed centrifugation concept, associated with a polymeric scaffold, had a significant effect on osteogenic markers of osteoblasts. |
| Al-Maawi et al., 2022 [18] | L-PRF and H-PRF | Primary human osteoblasts (pOBs) | 24 h | Eluate | Cell adhesion. | Osteoblasts exposed to PRF produced with fixed-angle rotors presented higher adhesion than those exposed to PRF produced with variable angles. |
| Bagio et al., 2021 [19] | A-PRF | Human dental pulp stem cells | 5, 12 and 24 h | Eluate | VEGF-A. | 5% A-PRF extracts increased VEGF-A expression by hDPSCs. |
| Banyatworakul et al., 2021 [20] | L-PRF | Canine periodontal ligament cells | 1, 3 and 7 days | Eluate | Proliferation; migration; in vitro mineralization. | PRF derived from Thai buffalo blood promoted the proliferation, migration, and increased mineral deposition in vitro of canine periodontal ligament cells. |
| Bi et al., 2020 [21] | L-PRF | Stem cells from the apical papilla (SCAP) | 1, 3 and 5 days | Eluate | Proliferation; migration; in vitro mineralization; ERK; pERK; ALP; DMP-1. | PRF improved the proliferation, migration, and the osteo-/odontogenic differentiation of SCAPs by activating the ERK pathway. |
| Blatt et al., 2021 [22] | A-PRF and iPRF | Human osteoblasts (HOBs) | 24 h | Co-culture | Cell viability; proliferation; migration; ALP; Col-I; BMP2; Runx2. | The combination of PRF with bone substitute materials increased the viability, early proliferation, and migration potential of human osteoblasts via Runx2 alkaline phosphatase, collagen, and BMP2. |
| Chang, Tsai, and Chang, 2010 [23] | L-PRF | Human osteosarcoma osteoblast-like cells (U2OS) | 1, 3 and 5 days | Co-culture | Proliferation; p-ERK, RANKL; OPG. | PRF stimulated osteoblast proliferation with positive regulation of the expression of p-ERK and increased the secretion of OPG. |
| Chen et al., 2015 [24] | L-PRF | Canine dental pulp stem cells (DPSCs) | 7, 14 and 21 days | Co-culture | Proliferation; ALP activity; ALP; DSPP; DMP 1; BSP. | PRF not only provides a well-organized scaffold for cell adhesion and migration but also induces DPSC proliferation and differentiation markers. |
| Cheng et al., 2022 [25] | L-PRF | Rabbit bone marrow mesenchymal stem cells (BMSCs) | Up to 28 days | Co-culture | Mineralization; adipogenic differentiation; aggrecan, Col-II; Sox9; b-catenin; P-GSK3b; CaMKII; PKC. | Co-culture with PRF reversed the activation of Wnt/Ca2+ signaling in BMSCs under hydrostatic pressure, with increased expression of chondrogenic differentiation markers. |
| Chi et al., 2019 [26] | Decellularized PRF | Bone marrow stem cells | every 24 h for 9 days | Cultured on PRF | Adhesion; proliferation; Col I; ALP; OPN; OCN; Runx-2. | Decellularized PRF combined with chitosan/gelatin scaffolds accelerate the attachment, proliferation, and osteogenesis-related marker expression of bone marrow stem cells. |
| Clipet et al., 2012 [27] | L-PRF | Human osteosarcoma osteoblast-like cells (Saos-2) | 1 and 2 days | Eluate | Cytotoxicity; proliferation; Cell Cycle; cbfa1, Col1, OCN; OPN | Exposure to a PRF-conditioned medium increased the cell viability, proliferation, and expression of the late and early markers of osteogenesis. |
| Dohan Ehrenfest et al., 2010 [28] | L-PRF | Human bone mesenchymal stem cells (BMSCs) | 7, 14, 21 and 28 days | Co-culture | Cytotoxicity; proliferation; ALP activity; mineralization; morphology (SEM). | Increased proliferation and differentiation of BMSC when exposed to Choukroun’s PRF. |
| Dohle et al., 2018 [29] | iPRF | Primary osteoblasts (pOBs) | 1 and 7 days | Cultured on PRF | VEGF; ICAM-1; ALP. | The expression of E-selectin, ICAM-1, VEGF, and ALP was significantly higher in the co-culture of primary osteoblasts and outgrowth endothelial cells cultured in PRF in vitro, in addition to improving the angiogenesis process. |
| Douglas et al., 2012 [30] | L-PRF | Human osteosarcoma osteoblast-like cells (Saos-2) | 3, 5 and 7 days | Co-culture | Cytocompatibility; migration. | PRF functionalized with ALP and induced to mineralization was not cytotoxic and promoted colonization by human osteoblasts. |
| Duan et al., 2018 [31] | L-PRF | Rat periodontal ligament stem cells (PDLSCs) | 1, 2, 3, 4, 7 and 14 days | Cultured on PRF | Proliferation; BSP; OCN; Runx2; ALP activity. | PRF enhanced cell proliferation and the expression of osteogenic markers in rat PDLSCs. |
| Ehrenfest et al., 2009 [32] | L-PRF | Human maxillofacial osteoblasts | 7, 14, 21 and 28 days | Co-culture | Proliferation; mineralization; ALP activity. | PRF stimulates the proliferation of several very different cell types, and the effects on osteoblastic differentiation are highly significant. |
| Esmaeilnejad et al., 2023 [33] | A-PRF and L-PRF | Human osteosarcoma osteoblast-like cells (MG-63) | 24 and 72 h | Eluate | Proliferation; mineralization. | L-PRF increased proliferation, while A-PRF increased the in vitro mineralization of MG-63 cells. |
| Fernandez-Medina et al., 2019 [34] | A-PRF and I-PRF | Primary human osteoblasts | 24 and 72 h | Eluate | Proliferation; migration; mineralization; cytokine release. | Cell viability and migration assays have demonstrated a detrimental effect when the concentration was ≥60. i-PRF demonstrated superior induction of mineralization. A negative impact of A-PRF was demonstrated at high concentrations. |
| Gassling et al., 2009 [35] | L-PRF | Human osteoblasts (HOBs) and osteosarcoma (Saos-2) | 10 days | Co-culture | PDGF; IGF; TGF-Beta. | PRF exposure led to an increased secretion of growth factors by osteoblasts. |
| Gassling et al., 2010 [36] | L-PRF | Human periosteal cells | 10 min, 1 h and 1 day. | Eluate | Cell viability; proliferation. | PRF appears to be superior to collagen (Bio-Gide) as a scaffold for human periosteal cell proliferation. |
| Gassling et al., 2013 [37] | Mg-enhanced and enzymatically mineralized PRF | Human osteosarcoma osteoblast-like cells (Saos-2) | 1, 3 and 7 days | Co-culture | Cell viability; proliferation; morphology. | The enzymatic mineralization of PRF did not affect osteoblast viability and the proliferation on the membrane. |
| Gassling et al., 2013 [38] | L-PRF | Human osteoblasts | 1, 5, 7 and 36 days | Eluate | Cell viability; proliferation, ALP activity. | The PRF membrane supports the proliferation of human osteoblast cells, in addition to being an adequate support for the cultivation of human osteoblasts in vitro. |
| Girija and Kavitha, 2020 [39] | PRF (undefined protocol) | Dental pulp cells | 2 h, overnight | Eluate | IL-6; IL-8; DMP-1, DSPP, STRO-1; mineralization. | The addition of bioactive radiopacifiers into PRF has a synergistic effect on the stimulation of odontoblastic differentiation of HDPCs, inducing mineralization. |
| He et al., 2009 [40] | L-PRF | Rat calvaria osteoblasts | 1, 7, 14, 21 and 28 days | Exudates | Proliferation; mineralization; ALP activity; cytokine release. | PRF released autologous growth factors gradually and expressed stronger and more durable effects on the proliferation and differentiation of rat osteoblasts than PRP in vitro. |
| Hong, Chen, and Jiang, 2018 [41] | Freeze-dried L-PRF | Apical papilla (SCAPs) | 7 and 14 days | Membrane dissolved in 10 mL of DMEM | Proliferation; migration; morphology; differentiation (CD45, CD90, and CD146), mineralization; ALP; BSP; DMP-1; DSPP. | Freeze-dried PRF promotes the proliferation, migration, and differentiation of SCAPs |
| Huang et al., 2010 [42] | L-PRF | Dental pulp cells (DPCs) | 0, 1, 3 and 5 days | Co-culture | Viability; proliferation; OPG; ALP activity. | PRF stimulates cell proliferation and the differentiation of DPCs by upregulating OPG and ALP expression. |
| Irastorza et al., 2019 [43] | L-PRF | Pulp stem cells | 4 days | Co-culture | Mineralization; ALP activity; ALP; Col-I; Osteonectin; Runx2, OSX. | Osteoblastic differentiation from human pulp stem cells was achieved with a combination of biomimetic rough titanium surfaces (BASTMs) with autologous plasma-derived fibrin-clot membranes. |
| Isobe et al., 2017 [44] | L-PRF from stored (frozen) blood | Human periosteal cells | 3 days | Eluate | Proliferation. | The quality of PRF clots prepared from stored whole blood samples is not significantly reduced and induced similar proliferation of periosteal cells as fresh PRF. |
| Ji et al., 2015 [45] | L-PRF | Periodontal ligament stem cells and bone marrow mesenchymal stem cells | 1, 2, 3, 4, 5, 6 and 7 days | Transwell inserts | Migration; proliferation; BSP; OCN; OPN; Col-III. | The association of PRF and TDM (treated dentin matrix) induced cell differentiation according to different markers. |
| Kang et al., 2011 [46] | L-PRF | Human alveolar bone marrow stem cells (hABMSCs) | 0, 0.5, 3, 6, and 12 h. 1, 7, 14, 21, 28 and 35 days | Eluate | Proliferation; mineralization; migration; MMP9 activity. | PRF increased the proliferation, aggregation, activation of MMP9, and mineralization by decreasing the migration of the hABMSCs. |
| Kardos et al., 2018 [47] | Fresh, frozen, and freeze-dried L-PRF | Mesenchymal stem cells | 1, 7 and 14 days | Co-culture | Viability; proliferation; adhesion. | Preserved PRF membranes presented the same biological properties as fresh samples. |
| Kim et al., 2017 [48] | L-PRF | Human primary osteoblasts | 1, 2, 3 and 7 days | Co-culture | Proliferation; ALP activity. | PRF presented significantly higher data on DNA quantification, synthesis and proliferation, differentiation, and bone generation of osteoblasts, PDGFs, and TGF-b. |
| Kim et al., 2017 [49] | L-PRF | Human dental pulp cells (HDPCs) | 1, 2 and 3 days | Eluate | Viability; IL-1b; IL-6, and IL-8; VCAM-1, DSP; DMP-1; ALP activity; mineralization. | PRF presents odontogenic capacity in inflamed HDPCs. |
| Koyanagi et al., 2022 [50] | Arterial blood-derived PRF (Ar-PRF) and venous blood-derived PRF (Ve-PRF) | Primary rabbit osteoblasts | 1, 3 and 5 days | Eluate | Viability; Col-1; OCN; mineralization. | Exposed osteoblasts presented greater differentiation potential, including higher osteocalcin expression and mineralization with no difference between Ar and Ve-PRF. |
| Kosmidis et al., 2023 [51] | A-PRF, i-PRF, and L-PRF | Human osteosarcoma osteoblast-like cells (U2OS) | Up to 28 days | Eluate | Mineralization; ALP activity; ALP; OCN; ON; ICAM-1; Runx2; Col 1a. | The three PRF preparations increased the osteogenic potential of U2OS cells. A-PRF presented the highest effect on mineralization, and i-PRF had the highest potential for early cell differentiation. |
| Kyyak et al., 2020 [52] | i-PRF | Human osteoblasts (HOBs) | 3, 7 and 10 days | Eluate | Viability; migration; proliferation; ALP; BMP-2; OCN. | i-PRF in combination with allogenic biomaterials enhances human osteoblast activity compared to xenogenic bone substitute material + i-PRF. |
| Kyyak et al., 2021 [53] | i-PRF | Human osteoblasts (HOBs) | 3, 7 and 10 days | Eluate | Viability; migration; proliferation; ALP; BMP-2; OCN. | The combination of four bovine bone substitute materials with i-PRF improved all cellular parameters, ALP and BMP-2 expression at earlier stages, and osteonectin expression at later stages. |
| Li et al., 2013 [54] | L-PRF | Dental follicle (DF), alveolar bone (AB), and periodontal ligament (PDL) | 7, 14 and 21 days | Co-culture | Proliferation; migration; mineralization; ALP; MGP; Runx2. | PRF induced an increase in the early osteoblast transcription factor Runx2 and a reduction in the mineralization inhibitor MGP. |
| Li et al., 2014 [55] | Lyophilized PRF | Rat alveolar bone cells | 7, 14 and 21 days | Co-culture | Mineralization; proliferation; ALP; Runx2. | Lyophilized PRF caused greater proliferation and elevation in the Runx2 expression in alveolar bone cells compared to fresh PRF and a more than 10-fold rise of ALP and in vitro mineralization. |
| Li et al., 2018 [56] | L-PRF | Human periodontal ligament stem cells (hPDLCs) | 21 days | Exudates | Adhesion; proliferation; mineralization; ALP activity; ALP; OCN; OSX; Runx2. | PRF exudate enhances hPDLC adhesion and proliferation and induces the differentiation of hPDLCs into mineralized tissue formation cells. |
| Li et al., 2018 [57] | L-PRF | Human periodontal ligament cells (PDLSCs) | 1, 2, 3, 7 and 14 days | Co-culture | Proliferation; Runx2; MAPK; ERK1/2; pERK1/2; JNK1/2/3; pJNK1/2/3; P38; OSX; OCN; ALP activity. | PRF and IGF-1 can promote the osteogenic differentiation of PDLSCs and enhance their osteogenic mineralization through the regulation of the MAPK pathway. |
| Liu et al., 2022 [59] | Lyophilized L-PRF, crosslinked with genipin | Pulp stem cells from human exfoliated deciduous teeth (SHEDs) | Up to 14 days | Eluate | Proliferation; mineralization; Runx2; Col 1; OCN. | Genipin crosslinked L-PRF induced cell proliferation and enhanced the expression of key genes in osteogenesis. |
| Liang et al., 2021 [58] | A-PRFe | Adipose-derived stem cells (ASCs) | 7 days | Eluate | Roliferation; mineralization; adipogenesis; ALP, OPN; OCN; Runx2. | A-PRF stimulated ASC proliferation and adipogenic and osteogenic differentiation in a dose-dependent manner. |
| Liu et al., 2019 [60] | Fresh/lyophilized PRF | Bone mesenchymal stem cells (BMSCs) | 1–7 days | Eluate | Proliferation; mineralization. | Fresh/lyophilized PRF (1:1) increased BMSC proliferation and in vitro mineralization. |
| Lo Monaco et al., 2020 [61] | L-PRF | Dental pulp stem cells (DPSCs) | 24, 48 and 72 h | Eluate | Chondrogenic differentiation; TIMP-1; proliferation. | L-PRF induced differential chondrogenesis on DPSCs. |
| Marchetti et al., 2020 [62] | L-PRF | Periodontal ligament fibroblasts | 24 h, 72 h and 7 days | Eluate | Proliferation; viability; morphology. | L-PRF stimulated the onset of the growth of the periodontal ligament fibroblasts. |
| Moradian et al., 2017 [63] | L-PRF | Bone marrow mesenchymal stem cells (BMMSCs) | 1, 5, 7, 9 and 12 days | Cultured on PRF | Proliferation; adhesion. | PRF significantly induced BMMSC proliferation. Scanning electron microscopy showed that BMMSCs tightly adhered to the fibrin scaffold after seeding. |
| Nguyen et al., 2022 [64] | A-PRF | Human periodontal ligament stem cells (hPDLSCs) | 1 h, 6 h, 24 h, 3 days, 5 days and 7 days | Exudates | Proliferation; migration. | A-PRF in combination with xenogenic bone induced hPDLSC migration or proliferation, depending on the exudate concentration. |
| Nie et al., 2020 [65] | Lyophilized L-PRF | MC3T3-E1 murine preosteoblasts | 1, 3 and 5 days | Eluate | Proliferation; mineralization; OCN; OPN. | Eluates from lyophilized PRF added as a component for electrospinning preparation enhanced the proliferation, mineralization, and expression of the OCN and OPN of MEC3T3-E1 cells. |
| Nugraha et al., 2018a [66] | L-PRF | Rat gingival mesenchymal stem cells (GMSCs) | 7, 14 and 21 days | Cultured on PRF | ALP; OC. | PRF induced increased ALP and OCN expression on GMSCs. |
| Nugraha et al., 2018b [67] | L-PRF | Rat gingival somatic cells (GSCs) | 7, 14 and 21 days | Cultured on PRF | BSP-1. | PRF increases and stimulates GSC BSP-1 expression. |
| Nugraha et al., 2018c [68] | L-PRF | Rat gingival stromal progenitor cells (GSPCs) | 7, 14 and 21 days | Cultured on PRF | Cbfa-1; sox9. | GSPCs cultured in PRF possessed a potential osteogenic differentiation ability, as predicted by the cbfa-1/sox9 expression ratio. |
| Nugraha et al., 2019 [69] | L-PRF | Rat gingival mesenchymal stem cells (GMSCs) | 7, 14 and 21 days | Cultured on PRF | Aggrecan. | Platelet-rich fibrin increases the aggrecan expression of GMSCs during osteogenic differentiation. |
| Rastegar et al., 2021 [70] | L-PRF | Human osteosarcoma osteoblast-like cells (MG-63) | 3 days | Co-culture | ALP activity; mineralization. | PRF loaded into PCL/chitosan core-shell fibers promoted in vitro mineralization and increased ALP activity. |
| Shah et al., 2021 [71] | i-PRF | Human osteosarcoma osteoblast-like cells (MG-63) | 1, 7, 14 and 21 days | Co-culture | Proliferation; ALP activity; mineralization. | Coating titanium discs with i-PRF causes increased proliferation, alkaline phosphatase production, and mineralization at days 1, 7, 14, and 21. |
| Song et al., 2018 [72] | L-PRF | Rabbit bone marrow-derived mesenchymal stem cells (BMSCs) | 7 days | Eluate | Adhesion; proliferation; ALP; Col-1; OPN; Runx2. | Printed scaffolds of BCP/PVA associated with PRF promoted the adhesion, proliferation, and differentiation of BMSCs. |
| Steller et al., 2019 [73] | L-PRF | Human osteoblasts (HOBs) | 72 h | Eluate | Proliferation; migration; viability. | The use of PRF improves the behavior of osteoblasts treated with zoledronic acid. |
| Steller et al., 2019 [74] | PRF | Human primary osteoblasts | 24 h | Eluate | Adhesion; viability; morphology. | Zoledronic acid decreased osteoblast adhesion on implant surfaces. PRF increased the primary adhesion of zoledronic acid-treated osteoblasts on implant surfaces in vitro. |
| Sui et al., 2023 [75] | 3D-printed L-PRF composite scaffolds | MC3T3-E1 murine preosteoblasts | 1 to 3 days | Cultured on a PRF composite scaffold | Proliferation. | The proliferation of preosteoblasts into the scaffolds increased with the release of GFs, indicating that L-PRF remains bioactive after 3D printing. |
| Thanasrisuebwong et al., 2020 [76] | Subfractioned (red and yellow) i-PRF | Periodontal ligament stem cells | 0, 3 and 5 days | Eluate | Proliferation; migration; mineralization; ALP activity. | The factors released from the red i-PRF had a greater effect on cell proliferation and cell migration, while yellow i-PRF stimulated earlier osteogenic differentiation of periodontal ligament stem cells. |
| Verboket et al., 2019 [77] | High (208 g) and low (60 g) RCF PRF | Bone marrow mononuclear cells (BMCs) | 2, 7 and 14 days | Eluate | Viability; apoptosis; VEGFA; ICAM3; MMP2; MMP7; MMP9; TGF-β1; BCL2; BAX; ALP; COL-1; FGF2; SPP1. | PRF produced with low RCF significantly increased mediator contents and stimulatory effects on BMC with regard to the gene expression of MMPs and metabolic activity/viability. |
| Wang et al., 2015 [78] | L-PRF | Rabbit mesenchymal stem cells (MSCs) | 1, 2, 3, 4, 5, 6, 7, 8 and 14 days | Eluate | Proliferation; ALP; BMP2; OCN; OPN; Col-1. | PRF significantly stimulated MSC proliferation and osteogenesis in vitro. |
| Wang et al., 2018 [79] | i-PRF | Human primary osteoblasts | 1, 3, 5, 7 and 14 days | Co-culture | Proliferation; migration; adhesion; mineralization; ALP activity; ALP; Col-1; OC. | i-PRF was able to influence osteoblast behavior, including migration, proliferation, and differentiation, at higher levels than PRP. |
| Wang et al., 2022 [80] | i-PRF | Human bone marrow stem cells (hBMSCs) | 1 to 7 days | Eluate | Proliferation; survival; migration; mineralization; Col 1; OCN; OPN; Runx2; ERK 1/2; p-ERK. | i-PRF improved the proliferation and migration of hBMSCs, with an increased expression of osteogenic markers, mineralization, and activation of the ERK pathway. |
| Wang et al. 2022 [81] | L-PRF | Rabbit mesenchymal stem cells from the Schneiderian membrane (SM-MSCs) | 1 to 14 days | Eluate | Proliferation; migration; mineralization; ALP activity; ALP; Col 1; Runx2; ERK 1/2; p-ERK. | PRF stimulated proliferation, migration, and osteogenic differentiation of SM-MSCs, with the upregulation of the ERK 1/2 signaling pathway. |
| Wong et al., 2021 [7] | Large-pore PRF (LPPRF) | MC3T3-E1 preosteoblasts | 6 days | Eluate | Proliferation; migration; mineralization. | Large-pore LPPRF combined with a Mg ring increased preoteoblast proliferation, migration, and in vitro calcium deposition. |
| Wong et al., 2021 [82] | L-PRF | Rabbit primary osteoblasts | 3 and 6 days | Eluate | Viability; ALP activity; Col-1; OPN; ALP. | L-PRF positively affected primary osteoblast behavior and induced bone formation when associated with TCP. |
| Woo et al., 2016 [83] | L-PRF | Human dental pulp cells (HDPCs) | 12 h. 1, 2 and 7 days | Eluate | Viability; ALP activity; DSP; DMP1; BMP 2/4; pSmad1/5/8. | A combination of MTA and PRF synergistically stimulated odontoblastic differentiation of HDPCs by the modulation of the BMP/Smad pathway. |
| Wu et al., 2012 [84] | L-PRF | Human osteosarcoma osteoblast-like cells (U2OS) | 2 h. 1, 3 and 5 days | Co-culture | Adhesion; proliferation p-Akt; HSP47; LOX. | PRF increased cell attachment and proliferation by the Akt pathway and matrix synthesis via HSP47 and LOX accumulation. |
| Yu et al., 2016 [85] | L-PRF | Canine deciduous and permanent dental pulp cells (DPCs) | 1, 4, 7 and 11 days | Co-culture | Cytotoxicity; proliferation; ALP activity; mineralization; Col-1; OCN; OPN; Runx2, ALP. | PRF stimulated the proliferation and differentiation of both deciduous and permanent DPCs, and deciduous pulp cells were more responsive to the effects of PRF. |
| Yu et al., 2023 [86] | H-PRF | Human osteoblasts (hFOBs) | 3 days | Transwell inserts | Migration. | The culture medium from H-PRF bone blocks markedly promoted the migration of osteoblasts. |
| Zhang et al., 2019 [87] | L-PRF | Dental pulp stem cells (DPSCs) | 1, 3, 5 and 7 days | Eluate | Migration; morphology; ALP activity; mineralization (SEM); OPN; Col-1; ALP. | Multifunctional triple-layered scaffolds combined with PRF significantly increased ALP activity and the expression of differentiation markers on DPSCs. |
| Zhang et al., 2023 [88] | i-PRF | Human dental pulp stem cells (hDPSCs) | Up to 21 days | Eluate | Proliferation; mineralization; ALP activity; Runx2; DSPP; DMP1; BSP; Notch 1; Jagged 1; Hes 1. | I-PRF induced a dose-dependent increase in the proliferation of hDPSCs and the expression of osteo-/odontoblastic differentiation markers, as well as key proteins in the Notch signaling. |
| Zhao et al., 2013 [89] | L-PRF | Periodontal ligament stem cells (PDLSCs) | 7, 14 and 21 days | Co-culture | Proliferation; mineralization (SEM); ALP activity; BSP; OCN; Col-I; CP23. | PRF induced proliferation in PDLSCs while suppressing the osteoblastic differentiation of PDLSCs by decreasing ALP activity and the gene expression of BSP and OCN while upregulating the mRNA expression levels of Col-I and CP23 during the testing period. |
| Zheng et al., 2015 [90] | Lyophilized PRF | Human osteosarcoma osteoblast-like cells (MG63) | 1, 3 and 5 days | Co-culture | Viability; adhesion; proliferation. | A combination of hydrogel and a nanostructured scaffold loaded with PRF improved the adhesion and proliferation of MG63 cells compared to the controls. |
| Zheng et al., 2020 [91] | i-PRF | Human periodontal ligament cells (hPDLCs) | 1, 3 and 5 days | Eluate | Migration; proliferation; ALP activity; mineralization; Runx2; Col-1; OCN; IL-1β; TNF-α and p65 (in the presence of LPS). | Liquid PRF promoted hPDLÇ proliferation and differentiation and attenuated the inflammatory state induced by LPS. |
| References | Associated Material | Relevance | Results |
|---|---|---|---|
| Al-Maawi, 2021 [17] | OsteoporeTM (OP), a commercially available PCL mesh | Combination of differently centrifuged PRF matrices with a polymeric resorbable scaffold to influence their biological properties on bone regeneration. | The presented results suggest that PRF produced according to the low-speed centrifugation concept exhibits autologous blood cells and growth factors and seems to have a significant effect on osteogenesis, showing promising results to support bone regeneration. |
| Chi, 2019 [26] | Chitosan/gelatin (C/G) | Test whether decellularized PRF (DPRF) maintains its bioactive effects to improve chitosan/gelatin (C/G) base scaffolds, which display appropriate biocompatibility and mechanical properties but lack biological activity to promote soft and hard tissue repair. | C/G/DPRF scaffolds accelerated attachment, proliferation, and osteogenesis-related marker expression of bone marrow stem cells. In vivo, C/G/DPRF scaffolds led to enhanced bone healing and defect closure in a rat calvarial defect model. Thus, it was concluded that DPRF remains bioactive, and the prepared C/G/DPRF scaffold is a promising material for bone regeneration. |
| Douglas, 2012 [30]; Gassling, 2013 [37] | Calcium glycerophosphate (CaGP) and ALP | Induce the mineralization of PRF membranes to achieve mechanical reinforcement of the gel and stability as a barrier membrane in guided bone regeneration. | The mineralization was confirmed, and WST test results showed that cell proliferation was inferior on PRF after the addition of ALP, confirming its properties as a barrier. |
| Girija and Kavitha, 2020 [39] | Bioactive radiopacifiers—nanohydroxyapatite (nHA) and dentin chips (DCs) | Combine bioactive radiopacifiers, nanohydroxyapatite (nHA) and dentin chips (DC), to PRF, aiming to produce a traceable material for endodontic procedures while still inducing adequate biological responses. | The results suggest that the addition of bioactive radiopacifiers into PRF has a synergistic effect on the stimulation of odontoblastic differentiation of HDPCs, inducing mineralization. |
| Ji B, 2015 [45] | Treated dentin matrix (TDM) | Associate endogenous stem cells, PRF, and TDM in the local microenvironment to contribute to the regeneration of periodontal tissues around the tooth root. | The study confirmed the role of PRF as a bioactive agent with TDM as an inductive scaffold for cells of the tooth socket microenvironment involved in endogenous tooth root regeneration. |
| Kyyak, 2020 [52] | Allogenic (ABSM) and xenogenic bone substitute material (XBSM) | The comparison of allogenic and xenogenic bone substitutes with i-PRF for the production of the more bioactive composite material for bone treatment. | i-PRF in combination with ABSM enhances HOB activity compared to XBSM-i-PRF or untreated BSM in vitro. Therefore, the addition of i-PRF to ABSM and—to a lower extent—XBSM may influence osteoblast activity in vivo in an interesting way for bone therapy. |
| Kyyak, 2021 [52] | Cerabone R (CB), Bio-Oss R (BO), Creos Xenogain R (CX), and MinerOSS R X (MO) | Four bovine bone substitute materials (XBSMs) were associated with i-PRF and aimed to increase their osteoinductive properties. | XBSM sintered under high temperatures showed increased HOB viability and metabolic activity throughout the whole period compared to XBSM manufactured at lower temperatures. Overall, the combination of XBSM with i-PRF improved all cellular parameters related to osteogenesis. |
| Nguyen, 2022 [64] | Xenogenic bone substitute material (XBSM) | Advanced platelet-rich fibrin (A-PRF) and xenogenic bone substitute material (XBSM) were associated and aimed to increase periodontal tissue regeneration. | The PRF-XBSM mixture continuously released growth factors over 7 days and enhanced human ligament stem cell proliferation and migration. |
| Nie, 2020 [65] | Polyvinyl alcohol/sodium alginate | The addition of lyophilized PRF as a component for electrospinning preparations to increase the proliferation and osteogenesis of osteogenic precursor cells for bioengineering purposes. | The resulting material presented adequate physicochemical properties and was able to increase osteogenic markers on bone cells. |
| Rastegar, 2021 [70] | PCL/chitosan | Platelet-rich fibrin (PRF)-loaded PCL/chitosan (PCL/CS-PRF) core-shell nanofibrous scaffold was made through a coaxial electrospinning method. The goal was to evaluate the effect of CS-RPF in the core layer of the nanofibrous on the osteogenic differentiation of human mesenchymal stem cells (HMSCs). | The formation of Ca-P on the surface of the scaffold immersed in a simulated body fluid solution indicated the suitable osteoconductivity of the PCL/CS-PRF core-shell nanofibrous scaffold. Due to the higher hydrophilicity and porosity of the PCL/CS-PRF core-shell nanofibrous scaffold compared to the PCL/CS scaffold, better bone cell growth on the surface of the PCL/CS-PRF scaffold was observed. |
| Song, 2018 [72] | Nano-biphasic calcium phosphate (BCP) and polyvinyl alcohol (PVA) | The low-temperature 3D printing of BCP/ PVA/PRF scaffolds would preserve the biological activity of PRF and provide an innovative biomaterial for restoring segmental bone defects. | The biological activity of PRF was retained during the 3D printing process, and the presence of PRF in the biocompatible microenvironment of the scaffold provided cell binding sites and promoted the adhesion, proliferation, and the differentiation of BMSCs. |
| Steller, 2019 [73] | Zoledronic acid | An investigation of the effects or bone cells treated with bisphosphonates as a potential mitigator of osteonecrosis associated with treatments with these drugs. | The negative effects of ZA on osteoblast survival and behavior (proliferation, morphology, adhesion to implant surface) were especially reduced using PRF, indicating that the autologous material may have positive effects in the therapy of bisphosphonate-related osteonecrosis of the kaw. |
| Sui et al., 2023 [75] | Chitosan (CS)–hydroxyapatite (HAP) scaffolds | A study aiming to identify if the 3D printing of a CS-HAP-PRF would compromise the biological properties of the platelet aggregate. | Based on the presented experimental results, it is possible to infer that the 2.5% P-C-H scaffold exhibits remarkable biological activity. And, therefore, it is not negatively affected by 3D printing. |
| Woo, 2016 [83] | Mineral trioxide aggregate (MTA) | Combined PRF as a bioactive matrix and MTA as a root-filling material beneficial for the endodontic management of an open apex. | The combination of MTA and PRF was proven as an odontogenic inducer in human dental pulp cells (HDPCs) in vitro. |
| Wong, 2021 [7] | Magnesium rings | The freeze-drying enlarges the pores of PRF to engineer large-pore PRF (LPPRF), a type of PRF that has expanded pores for cell migration. Biodegradable Mg rings were used to provide stability to these pores and release Mg ions during degradation, with the potential to enhance osteoconduction and osteoinduction. | The results revealed that cell migration was more extensive when LPPRF was used rather than PRF. Moreover, the Mg ions released from the Mg rings significantly enhanced the calcium deposition by preosteoblasts, evidencing in vitro osteoinduction. |
| Wong, 2021 [82] | Tricalcium phosphate (TCP) | The development of a composite biomaterial combining the osteoconductive TCP incorporated with bioactive PRF for bio-synergistic bone regeneration. | The in vitro results showed that PRF plus TCP had excellent biosafety and was favorable for increasing osteoblast activity related to bone repair. |
| Zhang, 2019 [87] | Polycaprolactone, chitosan, and hydroxyapatite | Polycaprolactone/gelatin (PG) nanofiber films by electrospinning chitosan/poly (γ-glutamic acid)/hydroxyapatite (CPH) hydrogels were formed by electrostatic interaction and lyophilization to exert osteoconduction, and platelet-rich fibrin (PRF) was added to promote bone induction through the release of growth factors. | This study provided evidence that the composite biomaterial positively affects dental pulp stem cells, with great potential for endodontics and wider applications, such as calvarial repair and oral alveolar bone regeneration. |
| Zheng, 2015 [90] | Copolymer poly-polyethylene glycol (PEG)-PLGA (PLGA-PEG-PLGA) | A combination of PRF with PLGA and nano-hydroxyapatite (nHA/PLGA) might produce a scaffold with high porosity, controlled pore size to better mimic natural bone, and improved osteogenic ability. | The resulting scaffold provided a good substrate for osteoblast proliferation with sustained-release growth factors, producing a promising therapeutic agent for local applications in bone tissue engineering. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lima Barbosa, R.; Stellet Lourenço, E.; de Azevedo dos Santos, J.V.; Rodrigues Santiago Rocha, N.; Mourão, C.F.; Alves, G.G. The Effects of Platelet-Rich Fibrin in the Behavior of Mineralizing Cells Related to Bone Tissue Regeneration—A Scoping Review of In Vitro Evidence. J. Funct. Biomater. 2023, 14, 503. https://0-doi-org.brum.beds.ac.uk/10.3390/jfb14100503
de Lima Barbosa R, Stellet Lourenço E, de Azevedo dos Santos JV, Rodrigues Santiago Rocha N, Mourão CF, Alves GG. The Effects of Platelet-Rich Fibrin in the Behavior of Mineralizing Cells Related to Bone Tissue Regeneration—A Scoping Review of In Vitro Evidence. Journal of Functional Biomaterials. 2023; 14(10):503. https://0-doi-org.brum.beds.ac.uk/10.3390/jfb14100503
Chicago/Turabian Stylede Lima Barbosa, Renata, Emanuelle Stellet Lourenço, Julya Vittoria de Azevedo dos Santos, Neilane Rodrigues Santiago Rocha, Carlos Fernando Mourão, and Gutemberg Gomes Alves. 2023. "The Effects of Platelet-Rich Fibrin in the Behavior of Mineralizing Cells Related to Bone Tissue Regeneration—A Scoping Review of In Vitro Evidence" Journal of Functional Biomaterials 14, no. 10: 503. https://0-doi-org.brum.beds.ac.uk/10.3390/jfb14100503