Effect of Gold Nanoparticles on the TLR2-Mediated Inflammatory Responses Induced by Leptospira in TLR2-Overexpressed HEK293 Cells


 and
and
Abstract
:1. Introduction
2. Materials and Methods
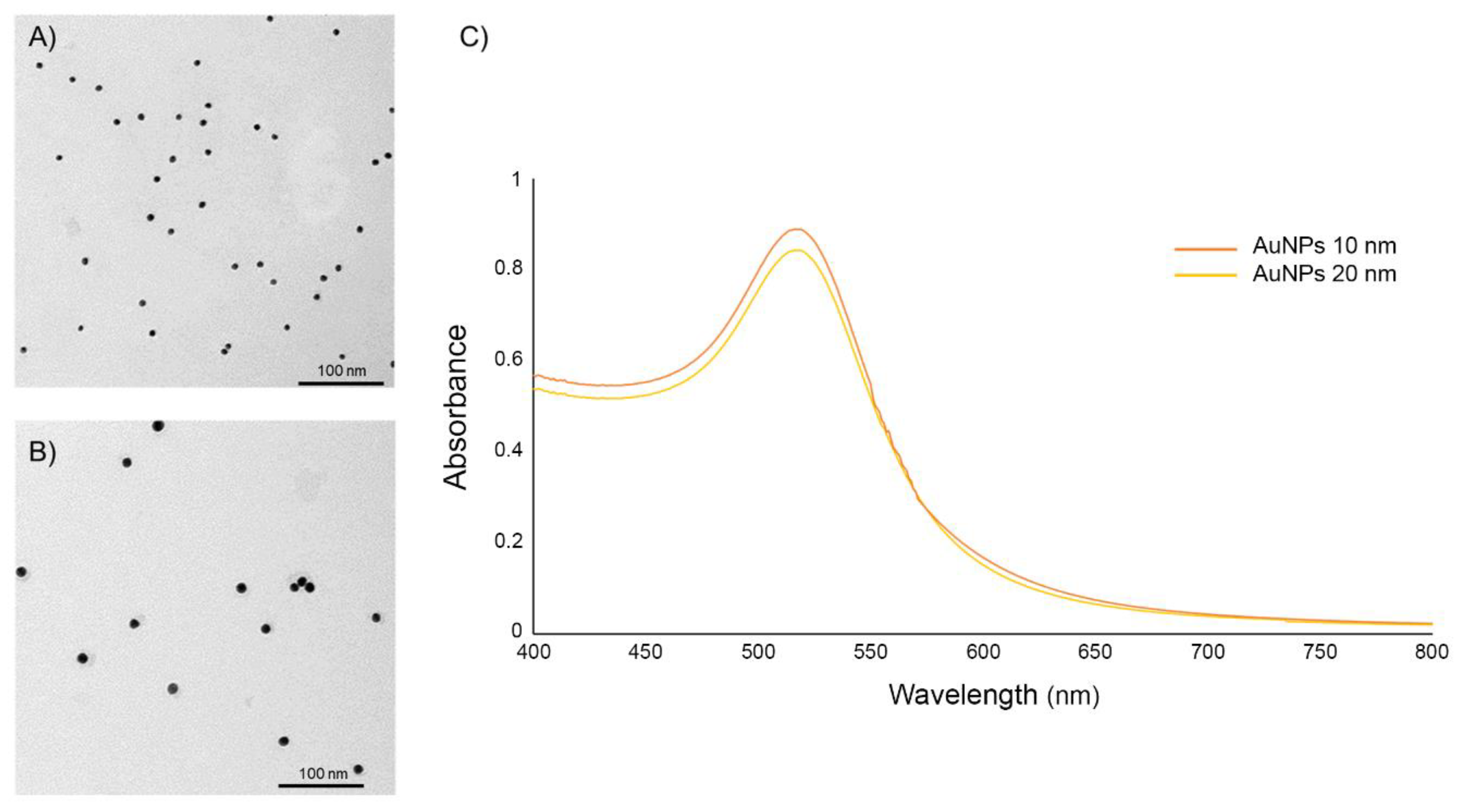
2.1. Characterization of AuNPs
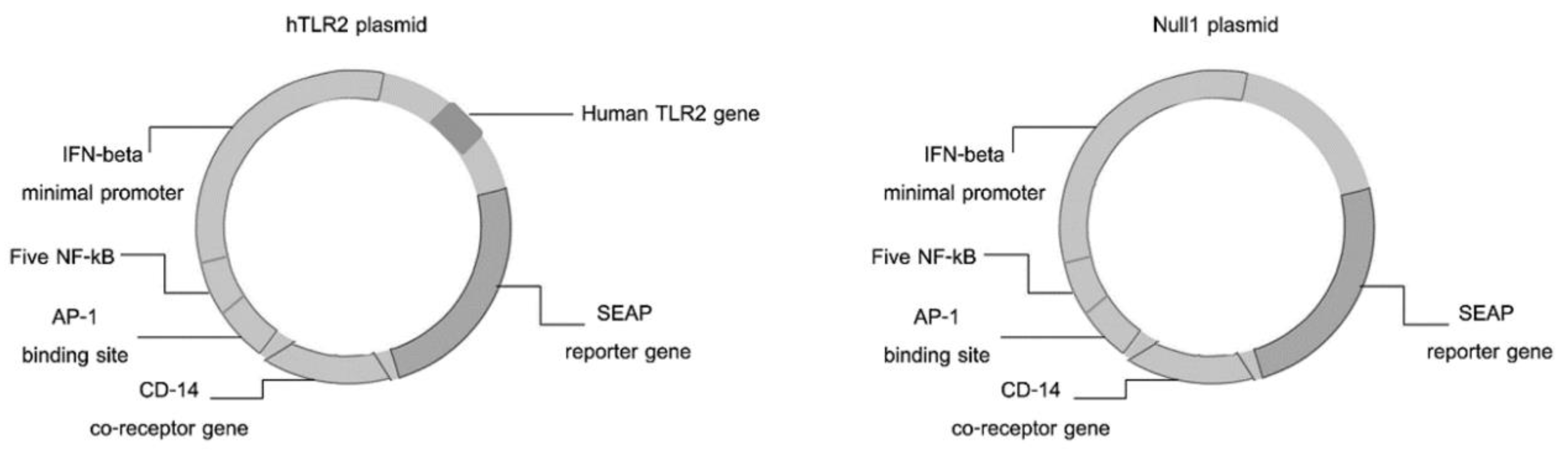
2.2. Cell Culture
2.3. Leptospira Culture Condition
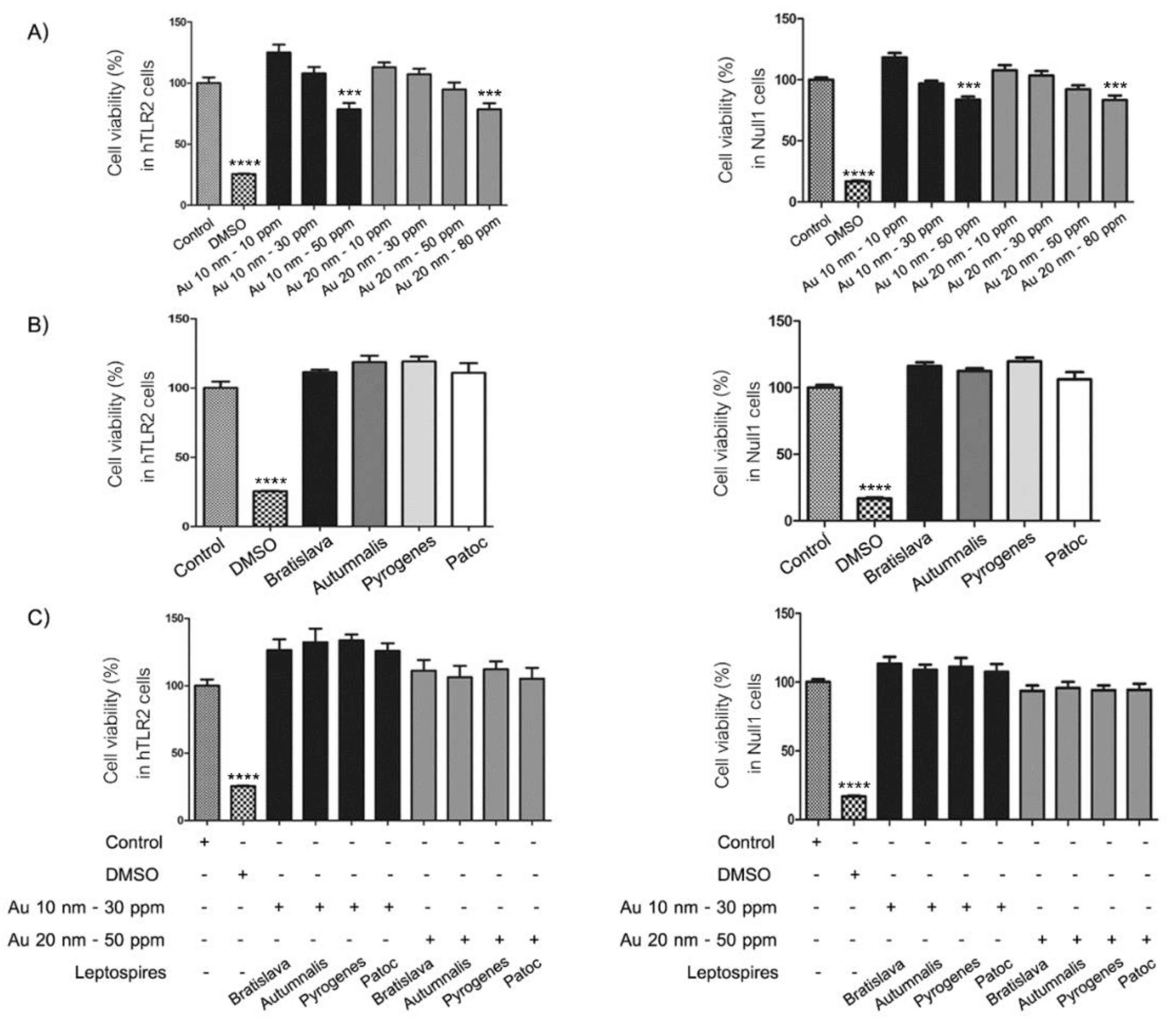
2.4. Cell Viability Assay
2.5. Cell Morphology
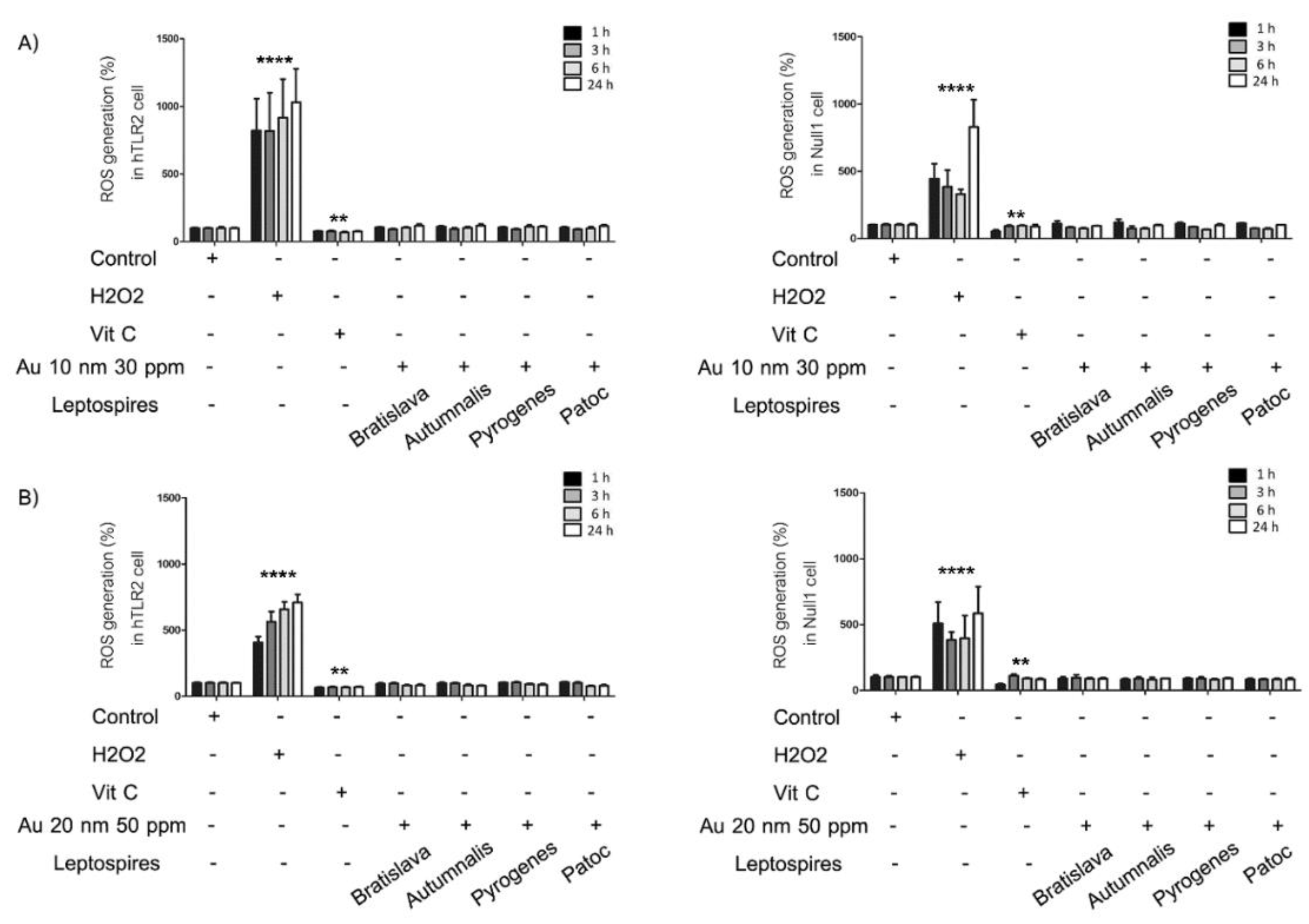
2.6. Generation of Reactive Oxygen Species (ROS)
2.7. Activation of TLR: Detection and Quantification of Alkaline Phosphatase Levels
2.8. RNA Extraction and Transcript Expression Level by Two-Stage Quantitative Real-Time Reverse Transcriptase (qrt-RT)-PCR
2.9. Statistical Analysis
3. Results
3.1. Characterization of the AuNPs
3.2. Cell Morphology and Viability Assay
3.3. Generation of ROS
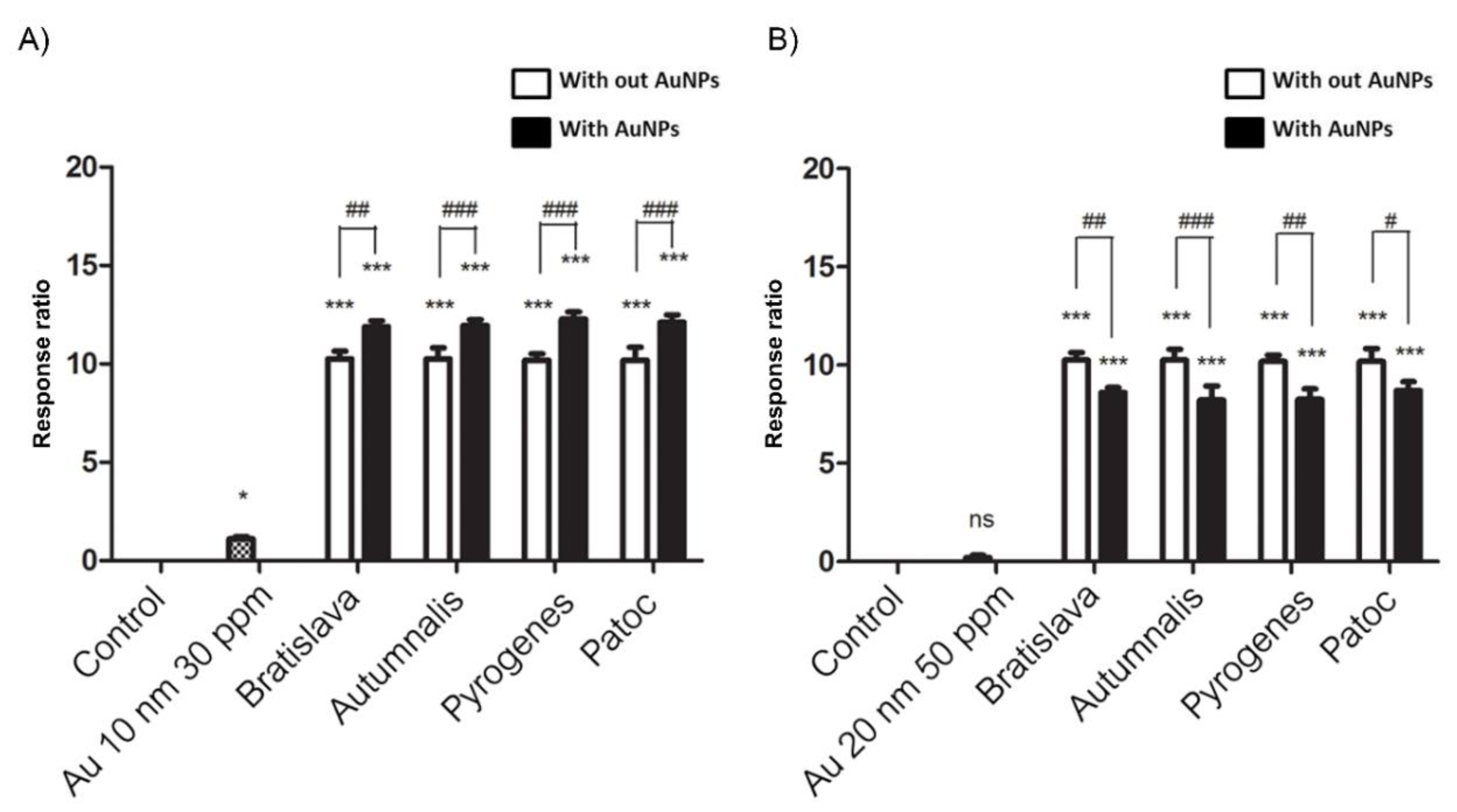
3.4. Stimulation of hTLR2, Evaluated by Monitoring NF-kB Activation
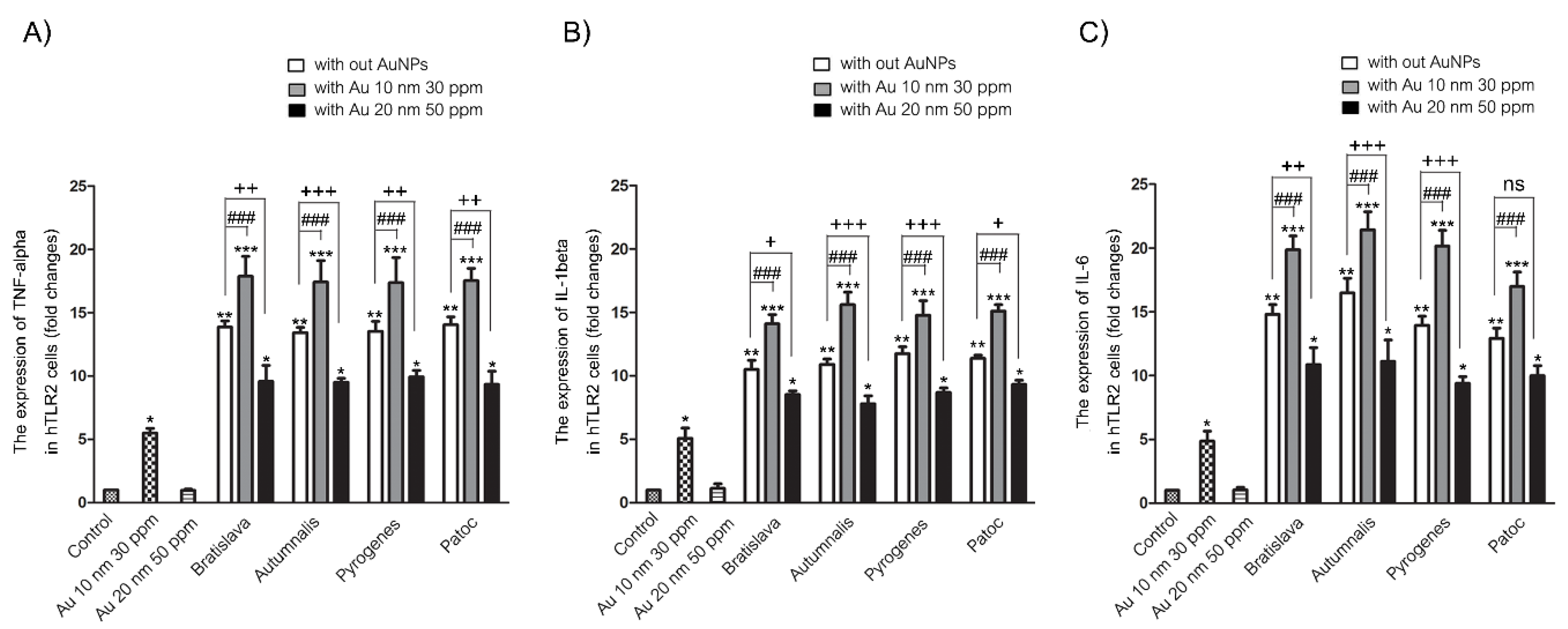
3.5. Proinflammatory mRNA Expression Levels
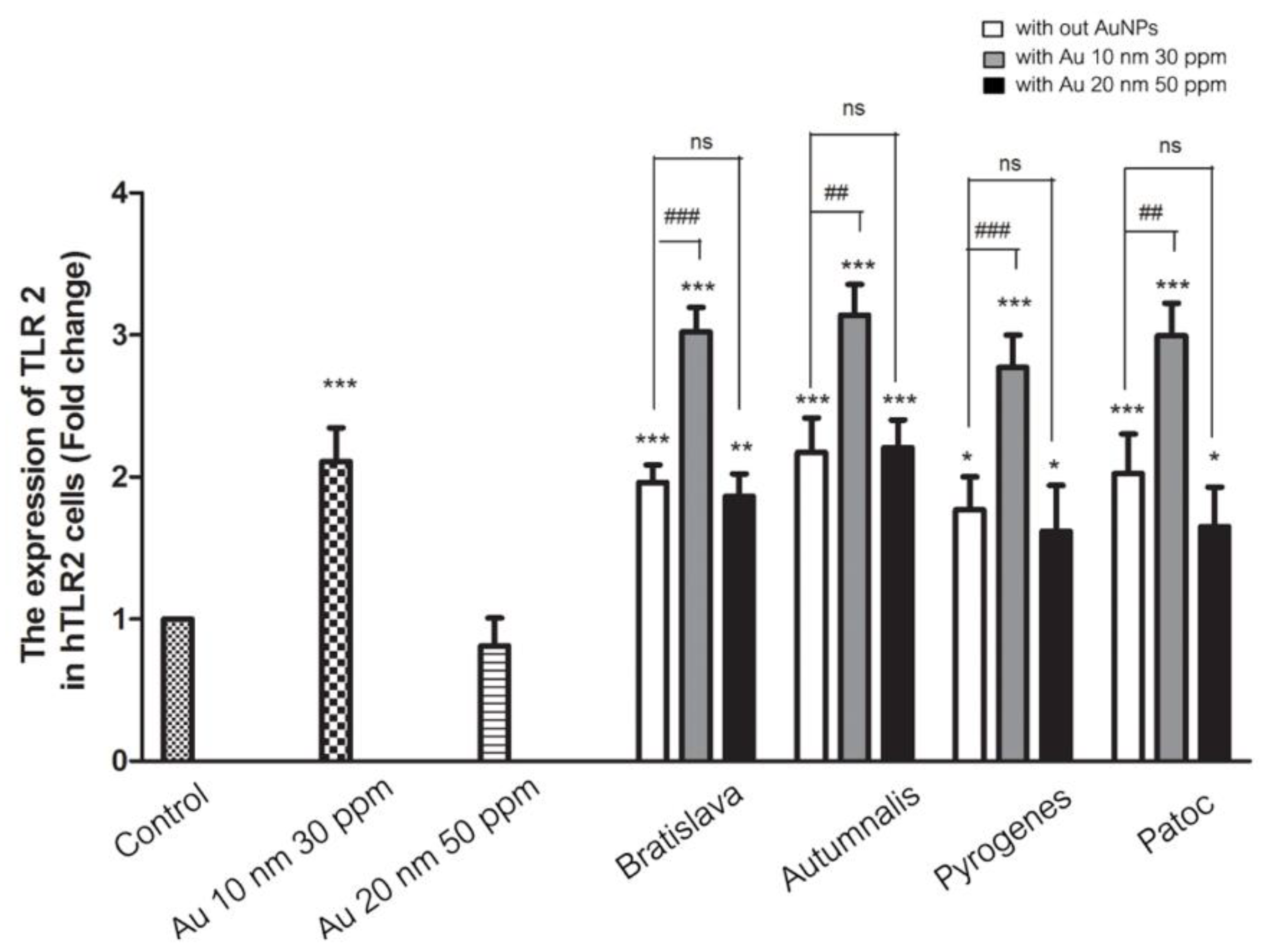
3.6. TLR2 mRNA Expression Level in Response to AuNPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meny, P.; Menendez, C.; Ashfield, N.; Quintero, J.; Rios, C.; Iglesias, T.; Schelotto, F.; Varela, G. Seroprevalence of leptospirosis in human groups at risk due to environmental, labor or social conditions. Rev. Argent. Microbiol. 2019, 51. [Google Scholar] [CrossRef] [PubMed]
- Cagliero, J.; Villanueva, S.; Matsui, M. Leptospirosis Pathophysiology: Into the Storm of Cytokines. Front. Cell. Infect. Microbiol. 2018, 8, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.R.; Peng, J.C.; Qiao, Y.Q.; Zhu, M.M.; Zhao, D.; Shen, J.; Ran, Z.H. Helicobacter pylori regulates TLR4 and TLR9 during gastric carcinogenesis. Int. J. Clin. Exp. Pathol. 2014, 7, 6950–6955. [Google Scholar] [PubMed]
- Wang, W.; Deng, Z.; Wu, H.; Zhao, Q.; Li, T.; Zhu, W.; Wang, X.; Tang, L.; Wang, C.; Cui, S.Z.; et al. A small secreted protein triggers a TLR2/4-dependent inflammatory response during invasive Candida albicans infection. Nat. Commun. 2019, 10, 1015. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, L.; Bekaert, T.; Chau, T.L.; Tavernier, J.; Chariot, A.; Beyaert, R. TLR-4, IL-1R and TNF-R signaling to NF-kappaB: Variations on a common theme. Cell. Mol. Life Sci. CMLS 2008, 65, 2964–2978. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Raffray, L.; Giry, C.; Vandroux, D.; Fayeulle, S.; Moiton, M.P.; Gerber, A.; Jaffar-Bandjee, M.C.; Gasque, P. The monocytosis during human leptospirosis is associated with modest immune cell activation states. Med. Microbiol. Immunol. 2018, 208. [Google Scholar] [CrossRef]
- Yang, H.; Fung, S.Y.; Bao, A.; Li, Q.; Turvey, S.E. Screening Bioactive Nanoparticles in Phagocytic Immune Cells for Inhibitors of Toll-like Receptor Signaling. J. Vis. Exp. JoVE 2017, 56075. [Google Scholar] [CrossRef]
- Fuller, M.A.; Koper, I. Biomedical applications of polyelectrolyte coated spherical gold nanoparticles. Nano Converg. 2019, 6, 11. [Google Scholar] [CrossRef]
- Carnovale, C.; Bryant, G.; Shukla, R.; Bansal, V. Size, shape and surface chemistry of nano-gold dictate its cellular interactions, uptake and toxicity. Prog. Mater. Sci. 2016, 83, 152–190. [Google Scholar] [CrossRef]
- Raja, V.; Prasad, M.; Bothammal, P.; Saranya, P.; Sumaiya, K.; Akino Mercy, C.S.; Natarajaseenivasan, K. Silver enhanced nano-gold dot blot immunoassay for leptospirosis. J. Microbiol. Methods 2019, 156, 20–22. [Google Scholar] [CrossRef]
- Hornos Carneiro, M.F.; Barbosa, F., Jr. Gold nanoparticles: A critical review of therapeutic applications and toxicological aspects. J. Toxicol. Environ. Health Part B Crit. Rev. 2016, 19, 129–148. [Google Scholar] [CrossRef]
- Chirathaworn, C.; Janwitthayanan, W.; Sereemaspun, A.; Lertpocasombat, K.; Rungpanich, U.; Ekpo, P.; Suwancharoen, D. Development of an immunochromatographic test with anti-LipL32-coupled gold nanoparticles for Leptospira detection. New Microbiol. 2014, 37, 201–207. [Google Scholar]
- Lopez-Campos, F.; Candini, D.; Carrasco, E.; Berenguer Frances, M.A. Nanoparticles applied to cancer immunoregulation. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Poznan Pol. Soc. Radiat. Oncol. 2019, 24, 47–55. [Google Scholar] [CrossRef]
- Hutter, E.; Boridy, S.; Labrecque, S.; Lalancette-Hebert, M.; Kriz, J.; Winnik, F.M.; Maysinger, D. Microglial response to gold nanoparticles. ACS Nano 2010, 4, 2595–2606. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.C.; Hung, C.C.; Li, Y.J.; Chen, Y.C.; Chang, M.Y.; Yen, T.H.; Hsu, H.H.; Wu, M.S.; Phillips, A.; Yang, C.W. Leptospira santorosai Serovar Shermani detergent extract induces an increase in fibronectin production through a Toll-like receptor 2-mediated pathway. Infect. Immun. 2011, 79, 1134–1142. [Google Scholar] [CrossRef] [Green Version]
- Inthasin, N.; Wongprompitak, P.; Boonwong, C.; Ekpo, P. Role of Toll-like receptor 2 in mediating the production of cytokines and human beta-defensins in oral mucosal epithelial cell response to Leptospiral infection. Asian Pac. J. Allergy Immunol. 2018, 37. [Google Scholar] [CrossRef]
- Lo, Y.Y.; Hsu, S.H.; Ko, Y.C.; Hung, C.C.; Chang, M.Y.; Hsu, H.H.; Pan, M.J.; Chen, Y.W.; Lee, C.H.; Tseng, F.G.; et al. Essential calcium-binding cluster of Leptospira LipL32 protein for inflammatory responses through the Toll-like receptor 2 pathway. J. Biol. Chem. 2013, 288, 12335–12344. [Google Scholar] [CrossRef] [Green Version]
- Werts, C.; Tapping, R.I.; Mathison, J.C.; Chuang, T.H.; Kravchenko, V.; Saint Girons, I.; Haake, D.A.; Godowski, P.J.; Hayashi, F.; Ozinsky, A.; et al. Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nat. Immunol. 2001, 2, 346–352. [Google Scholar] [CrossRef]
- Voicu, S.N.; Balas, M.; Stan, M.S.; Trica, B.; Serban, A.I.; Stanca, L.; Hermenean, A.; Dinischiotu, A. Amorphous Silica Nanoparticles Obtained by Laser Ablation Induce Inflammatory Response in Human Lung Fibroblasts. Materials 2019, 12, 1026. [Google Scholar] [CrossRef] [Green Version]
- Pedata, P.; Ricci, G.; Malorni, L.; Venezia, A.; Cammarota, M.; Volpe, M.G.; Iannaccone, N.; Guida, V.; Schiraldi, C.; Romano, M.; et al. In vitro intestinal epithelium responses to titanium dioxide nanoparticles. Food Res. Int. (Ottawa Ont.) 2019, 119, 634–642. [Google Scholar] [CrossRef]
- Jin, R.; Liu, L.; Zhu, W.; Li, D.; Yang, L.; Duan, J.; Cai, Z.; Nie, Y.; Zhang, Y.; Gong, Q.; et al. Iron oxide nanoparticles promote macrophage autophagy and inflammatory response through activation of toll-like Receptor-4 signaling. Biomaterials 2019, 203, 23–30. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, T.G.; Garcia, V.B.; de Araujo, A.A.; da Silva Gasparotto, L.H.; Silva, H.; Guerra, G.C.B.; de Castro Miguel, E.; de Carvalho Leitao, R.F.; da Silva Costa, D.V.; Cruz, L.J.; et al. Spherical neutral gold nanoparticles improve anti-inflammatory response, oxidative stress and fibrosis in alcohol-methamphetamine-induced liver injury in rats. Int. J. Pharm. 2018, 548, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kharazian, B.; Lohse, S.E.; Ghasemi, F.; Raoufi, M.; Saei, A.A.; Hashemi, F.; Farvadi, F.; Alimohamadi, R.; Jalali, S.A.; Shokrgozar, M.A.; et al. Bare surface of gold nanoparticle induces inflammation through unfolding of plasma fibrinogen. Sci. Rep. 2018, 8, 12557. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.P.; Hunter, Z.N.; Chackerian, B.; Shea, L.D.; Miller, S.D. Targeted immunomodulation using antigen-conjugated nanoparticles. Wiley interdisciplinary reviews. Nanomed. Nanobiotechnol. 2014, 6, 298–315. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.L.; Gustafsson, J.; Cronholm, P.; Moller, L. Size-dependent toxicity of metal oxide particles—A comparison between nano- and micrometer size. Toxicol. Lett. 2009, 188, 112–118. [Google Scholar] [CrossRef]
- Yang, E.J.; Kim, S.; Kim, J.S.; Choi, I.H. Inflammasome formation and IL-1beta release by human blood monocytes in response to silver nanoparticles. Biomaterials 2012, 33, 6858–6867. [Google Scholar] [CrossRef]
- Chen, H.; Dorrigan, A.; Saad, S.; Hare, D.J.; Cortie, M.B.; Valenzuela, S.M. In vivo study of spherical gold nanoparticles: Inflammatory effects and distribution in mice. PLoS ONE 2013, 8, e58208. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.A.; Alamery, S.; Ibrahim, K.E.; El-Nagar, D.M.; Al-Harbi, N.; Rusop, M.; Alrokayan, S.H. Size and time-dependent induction of proinflammatory cytokines expression in brains of mice treated with gold nanoparticles. Saudi J. Biol. Sci. 2019, 26, 625–631. [Google Scholar] [CrossRef]
- Gao, W.; Wang, Y.; Xiong, Y.; Sun, L.; Wang, L.; Wang, K.; Lu, H.Y.; Bao, A.; Turvey, S.E.; Li, Q.; et al. Size-dependent anti-inflammatory activity of a peptide-gold nanoparticle hybrid in vitro and in a mouse model of acute lung injury. Acta Biomater. 2019, 85, 203–217. [Google Scholar] [CrossRef]
- Chen, X.; Gao, C. Influences of size and surface coating of gold nanoparticles on inflammatory activation of macrophages. Colloids Surf. B Biointerfaces 2017, 160, 372–380. [Google Scholar] [CrossRef]
- Injumpa, W.; Ritprajak, P.; Insin, N. Size-dependent cytotoxicity and inflammatory responses of PEGylated silica-iron oxide nanocomposite size series. J. Magn. Magn. Mater. 2017, 427, 60–66. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Lu, S.L.; Hu, C.W.; Yeh, C.S.; Lee, G.B.; Lei, H.Y. Size-dependent attenuation of TLR9 signaling by gold nanoparticles in macrophages. J. Immunol. 2012, 188, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.V.; Petronilho, F.; Pereira, H.R.; Vuolo, F.; Mina, F.; Possato, J.C.; Vitto, M.F.; de Souza, D.R.; da Silva, L.; da Silva Paula, M.M.; et al. Effects of gold nanoparticles on endotoxin-induced uveitis in rats. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8036–8041. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | Annealing Temperature (°C) | Expected Amplicon Size (bp) |
|---|---|---|---|
| TNF-α | F: CCAGGCAGTCAGATCA | 64 | 189 |
| R: TGGGAGTAGATGAGGTACAG | |||
| IL-1β | F: TGGAGCAACAAGTGGTGT | 57 | 157 |
| R: TTGGGATCTACACTCTCCAGC | |||
| IL-6 | F: TGCAGAAAAAGGCAAA | 63 | 203 |
| R: CAACAACAATCTGAGGTG | |||
| TLR-2 | F: GGCCAGCAAATTACCTGTGTG | 60 | 67 |
| R: AGGCGGACATCCTGAACCT | |||
| GADPH | F: CAGGGGCCATCCACAGTCTTC | 59 | 357 |
| R: CATCACCATCTTCCAGGAGCG |
| Nanoparticles (Dispersed in Culture Media Supplemented with 10% FBS) | pH | Zeta Potential (mV) | Hydrodynamic Diameter (nm) | PdI |
|---|---|---|---|---|
| 10 nm AuNPs | 7.28 | −37.3 mV | 15.80 | 0.493 |
| 20 nm AuNPs | 7.31 | −31.3 mV | 27.38 | 0.501 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sooklert, K.; Boonwong, C.; Ekpo, P.; Rojanathanes, R.; Patarakul, K.; Chirathaworn, C.; Thamakaison, S.; Sereemaspun, A. Effect of Gold Nanoparticles on the TLR2-Mediated Inflammatory Responses Induced by Leptospira in TLR2-Overexpressed HEK293 Cells. Nanomaterials 2020, 10, 2522. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10122522
Sooklert K, Boonwong C, Ekpo P, Rojanathanes R, Patarakul K, Chirathaworn C, Thamakaison S, Sereemaspun A. Effect of Gold Nanoparticles on the TLR2-Mediated Inflammatory Responses Induced by Leptospira in TLR2-Overexpressed HEK293 Cells. Nanomaterials. 2020; 10(12):2522. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10122522
Chicago/Turabian StyleSooklert, Kanidta, Chawikan Boonwong, Pattama Ekpo, Rojrit Rojanathanes, Kanitha Patarakul, Chintana Chirathaworn, Sasin Thamakaison, and Amornpun Sereemaspun. 2020. "Effect of Gold Nanoparticles on the TLR2-Mediated Inflammatory Responses Induced by Leptospira in TLR2-Overexpressed HEK293 Cells" Nanomaterials 10, no. 12: 2522. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10122522