
Enhanced Bioaccumulation and Toxicity of Arsenic in Marine Mussel Perna viridis in the Presence of CuO/Fe3O4 Nanoparticles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Nanoparticles Characterization
2.2. Experimental Design
2.3. Analysis of As in Mussels
2.4. As Bioconcentration Factor (BCF)
2.5. As(V) Adsorption to MNPs in Seawater
2.6. Biomarkers Determination
2.7. Statistical Analysis
3. Results and Discussion
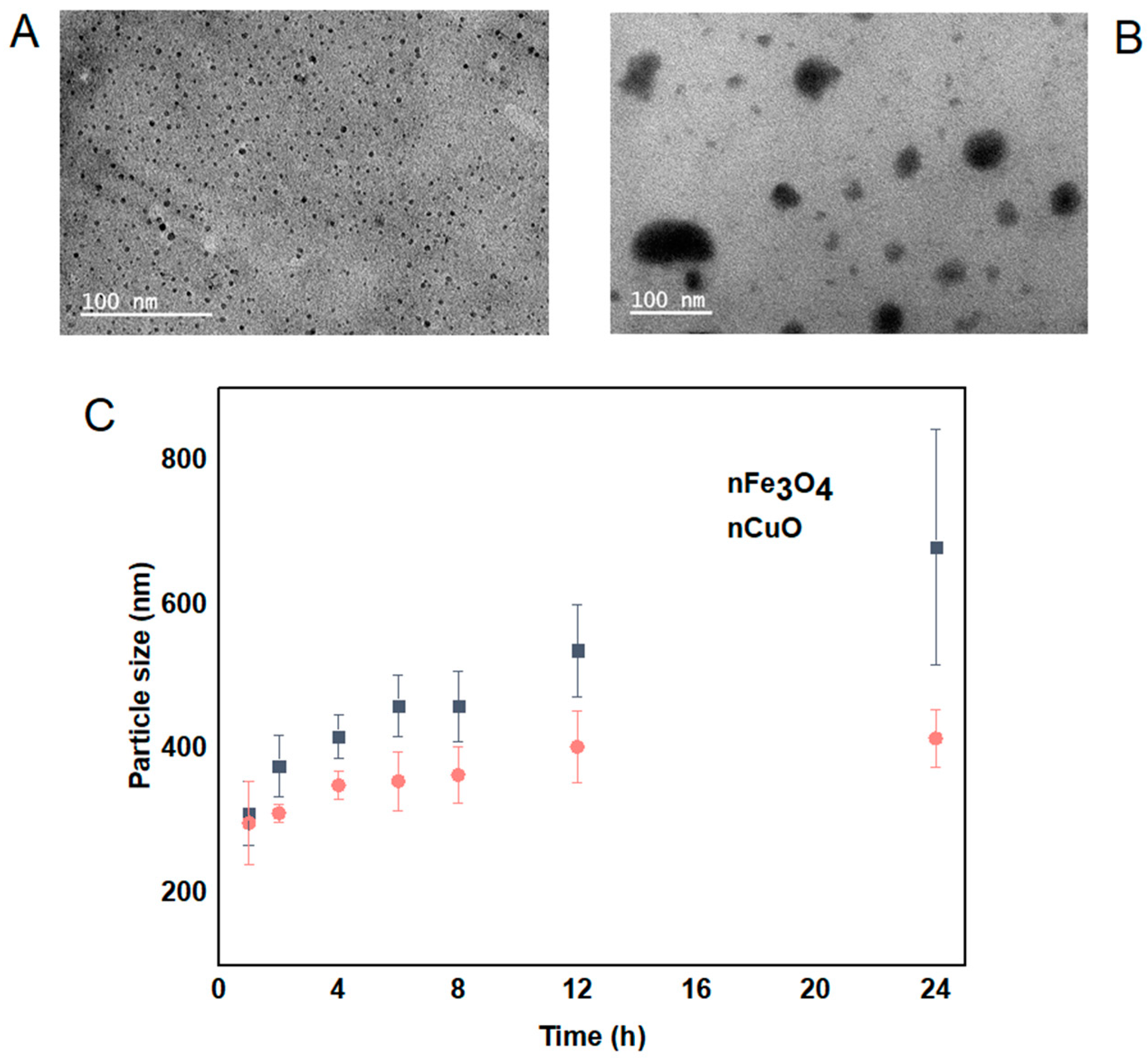
3.1. MNPs Characterization
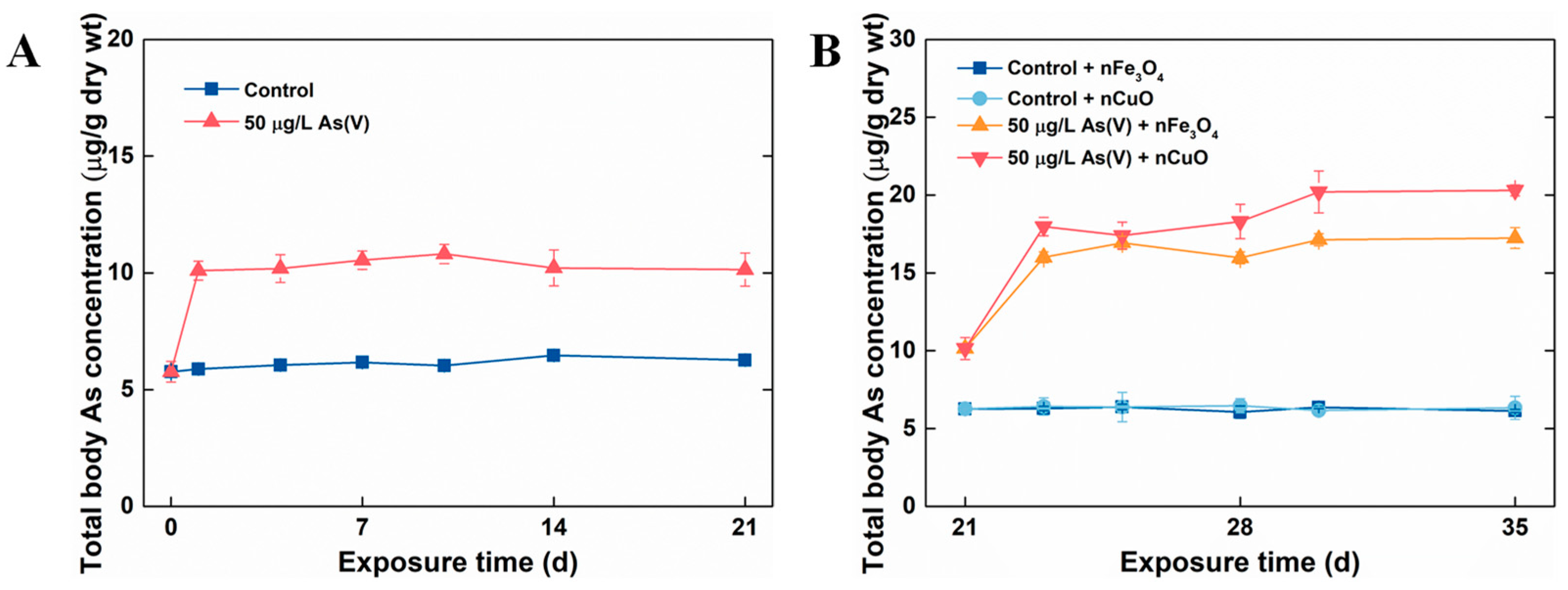
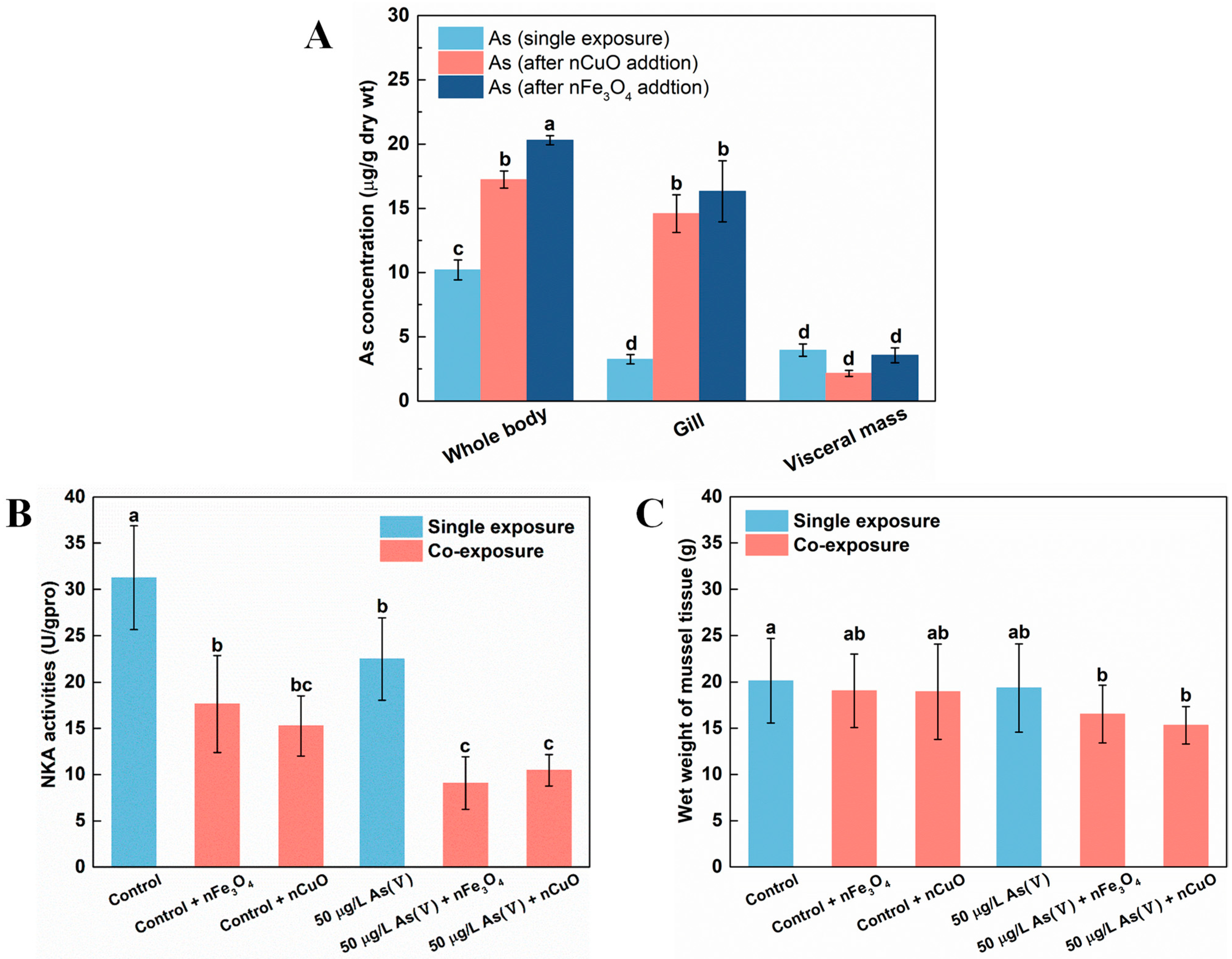
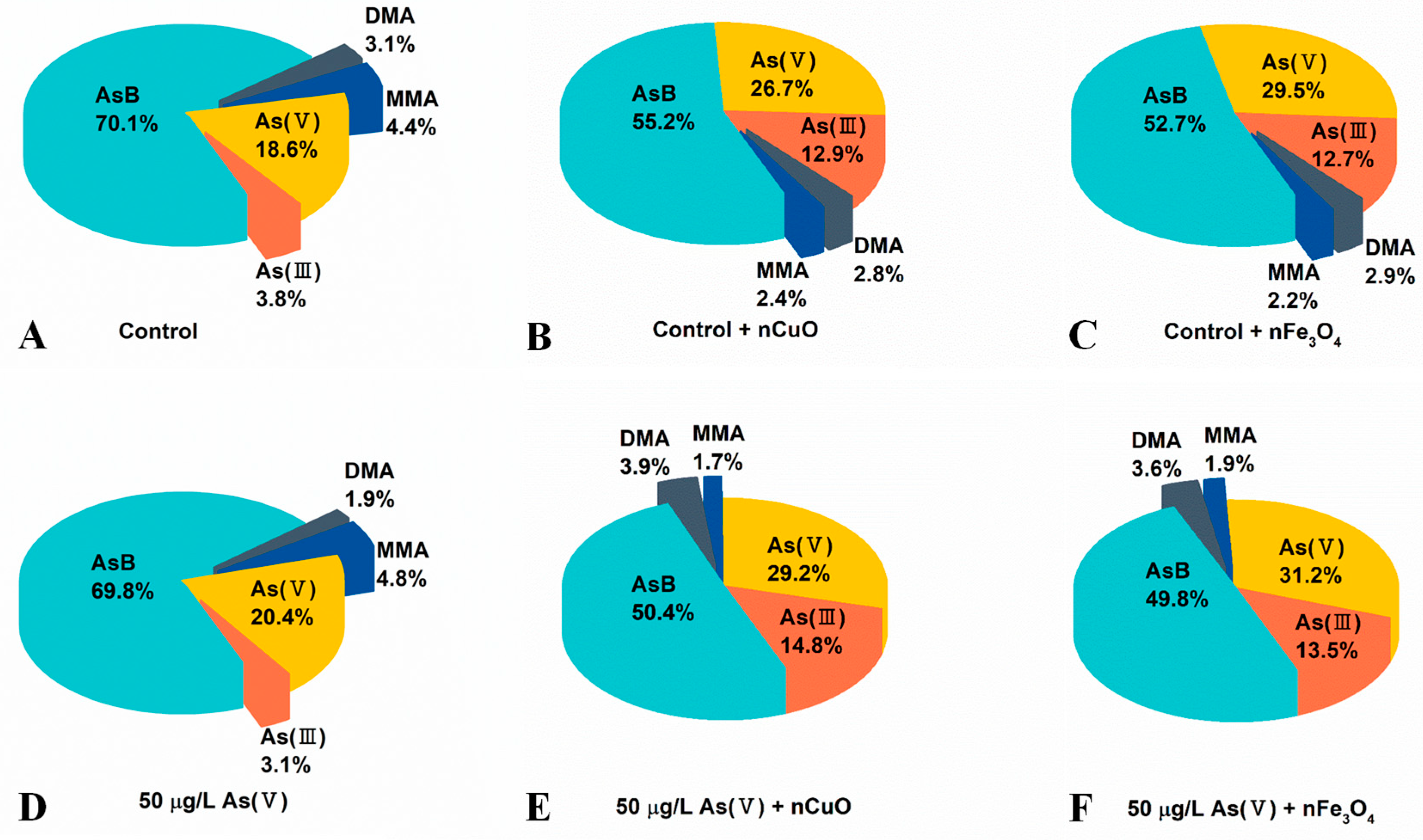
3.2. Exposure to MNPs Increased As Bioaccumulation and Altered As Distribution in Mussels
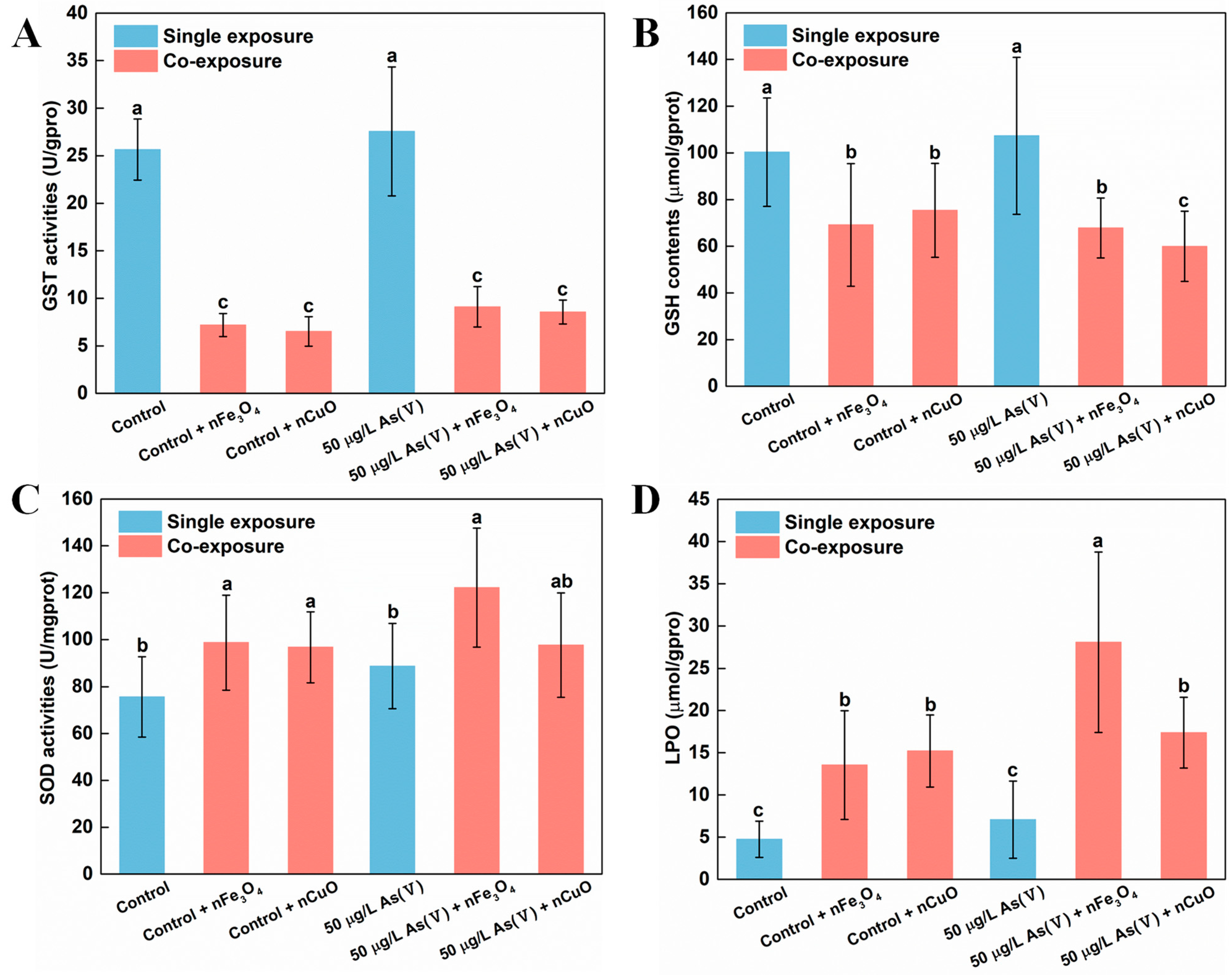
3.3. Exposure to MNPs Enhanced the Toxicity of As in Mussels
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Yoon, K.Y.; Hoon Byeon, J.; Park, J.H.; Hwang, J. Susceptibility constants of Escherichia coli and Bacillus subtilis to silver and copper nanoparticles. Sci. Total Environ. 2007, 373, 572–575. [Google Scholar] [CrossRef]
- Fahmy, B.; Cormier, S.A. Copper oxide nanoparticles induce oxidative stress and cytotoxicity in airway epithelial cells. Toxicol. Vitr. 2009, 23, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Gomes, T.; Pinheiro, J.P.; Cancio, I.; Pereira, C.G.; Cardoso, C.; Bebianno, M.J. Effects of copper nanoparticles exposure in the mussel Mytilus galloprovincialis. Environ. Sci. Technol. 2011, 45, 9356–9362. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Cui, B.; Li, G.; Yang, J.; Peng, H.; Wang, Y.; Li, N.; Gao, R.; Chang, Z.; Wang, Y. Novel Fe3O4@ZnO@mSiO2 Nanocarrier for targeted drug delivery and controllable release with microwave irradiation. J. Phys. Chem. C 2014, 118, 14929–14937. [Google Scholar] [CrossRef]
- Sadri, F.; Ramazani, A.; Massoudi, A.; Khoobi, M.; Tarasi, R.; Shafiee, A.; Azizkhani, V.; Dolatyari, L.; Joo, S.W. Green oxidation of alcohols by using hydrogen peroxide in water in the presence of magnetic Fe3O4 nanoparticles as recoverable catalyst. Green Chem. Lett. Rev. 2014, 7, 257–264. [Google Scholar] [CrossRef]
- Zhang, Y.R.; Su, P.; Huang, J.; Wang, Q.R.; Zhao, B.X. A magnetic nanomaterial modified with poly-lysine for efficient removal of anionic dyes from water. Chem. Eng. J. 2015, 262, 313–318. [Google Scholar] [CrossRef]
- Abbas, Q.; Yousaf, B.; Ullah, H.; Ali, M.U.; Ok, Y.S.; Rinklebe, J. Environmental transformation and nano-toxicity of engineered nano-particles (ENPs) in aquatic and terrestrial organisms. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2523–2581. [Google Scholar] [CrossRef]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F.; Hull, D.R. Nanotechnology in the real world: Redeveloping the nanomaterial consumer products inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Deng, R.; Lin, D.; Zhu, L.; Majumdar, S.; White, J.C.; Gardea-Torresdey, J.L.; Xing, B. Nanoparticle interactions with co-existing contaminants: Joint toxicity, bioaccumulation and risk. Nanotoxicology 2017, 11, 591–612. [Google Scholar] [CrossRef]
- Smedley, P.L.; Kinniburgh, D.G. A review of the source, behaviour and distribution of arsenic in natural waters. Appl. Geochem. 2002, 17, 517–568. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Gao, X.; Gu, Y.; Wang, R.; Xie, P.; Liang, M.; Ming, H.; Su, J. Comprehensive large-scale investigation and assessment of trace metal in the coastal sediments of Bohai Sea. Mar. Pollut. Bull. 2018, 129, 126–134. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, Z.; Song, D.; Du, S.; Zhang, L. Arsenic speciation in wild marine organisms and a health risk assessment in a subtropical bay of China. Sci. Total Environ. 2018, 626, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Byeon, E.; Kang, H.; Yoon, C.; Lee, J. Toxicity mechanisms of arsenic compounds in aquatic organisms. Aquat. Toxicol. 2021, 237, 105901. [Google Scholar] [CrossRef]
- Baumann, J.; Köser, J.; Arndt, D.; Filser, J. The coating makes the difference: Acute effects of iron oxide nanoparticles on Daphnia magna. Sci. Total Environ. 2014, 484, 176–184. [Google Scholar] [CrossRef]
- Rotini, A.; Tornambè, A.; Cossi, R.; Iamunno, F.; Benvenuto, G.; Berducci, M.T.; Maggi, C.; Thaller, M.C.; Cicero, A.M.; Manfra, L.; et al. Salinity-based toxicity of CuO nanoparticles, CuO-bulk and Cu ion to vibrio anguillarum. Front. Microbiol. 2017, 8, 2076. [Google Scholar] [CrossRef]
- Maurya, R.; Pandey, A.K. Importance of protozoa Tetrahymena in toxicological studies: A review. Sci. Total Environ. 2020, 741, 140058. [Google Scholar] [CrossRef] [PubMed]
- Braz-Mota, S.; Campos, D.F.; MacCormack, T.J.; Duarte, R.M.; Val, A.L.; Almeida-Val, V.M.F. Mechanisms of toxic action of copper and copper nanoparticles in two Amazon fish species: Dwarf cichlid (Apistogramma agassizii) and cardinal tetra (Paracheirodon axelrodi). Sci. Total Environ. 2018, 630, 1168–1180. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Chen, C.C.; Zhou, S.; Huang, Y.; Zhu, X.; Wang, Z.; Cai, Z. TiO2 Nanoparticles in the marine environment: Enhancing bioconcentration, while limiting biotransformation of arsenic in the Mussel Perna viridis. Environ. Sci. Technol. 2020, 54, 12254–12261. [Google Scholar] [CrossRef]
- Zhelyazkov, G.; Yankovska-Stefanova, T.; Mineva, E.; Stratev, D.; Vashin, I.; Dospatliev, L.; Valkova, E.; Popova, T. Risk assessment of some heavy metals in mussels (Mytilus galloprovincialis) and veined rapa whelks (Rapana venosa) for human health. Mar. Pollut. Bull. 2018, 128, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Coppola, F.; Almeida, Â.; Henriques, B.; Soares, A.M.V.M.; Figueira, E.; Pereira, E.; Freitas, R. Biochemical responses and accumulation patterns of Mytilus galloprovincialis exposed to thermal stress and Arsenic contamination. Ecotoxicol. Environ. Saf. 2018, 147, 954–962. [Google Scholar] [CrossRef]
- Hanna, S.; Miller, R.; Lenihan, H. Accumulation and toxicity of copper oxide engineered nanoparticles in a marine mussel. Nanomaterials 2014, 4, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Guo, Z.; Zhou, Y.; Liu, H.; Zhang, L. Biotransformation and detoxification of inorganic arsenic in Bombay oyster Saccostrea cucullata. Aquat. Toxicol. 2015, 158, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Whaley-Martin, K.J.; Koch, I.; Moriarty, M.; Reimer, K.J. Arsenic speciation in blue mussels (Mytilus edulis) along a highly contaminated arsenic gradient. Environ. Sci. Technol. 2012, 46, 3110–3118. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.; Araújo, O.; Pereira, R.; Almeida, A.C.; Cravo, A.; Bebianno, M.J. Genotoxicity of copper oxide and silver nanoparticles in the mussel Mytilus galloprovincialis. Mar. Environ. Res. 2013, 84, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Dringen, R.; Petters, C.; Rastedt, W.; Köser, J.; Filser, J.; Stolte, S. Toxicity of dimercaptosuccinate-coated and un-functionalized magnetic iron oxide nanoparticles towards aquatic organisms. Environ. Sci. Nano. 2016, 3, 754–767. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Huang, L.; Wang, W.X. Biotransformation and detoxification of inorganic arsenic in a marine juvenile fish Terapon jarbua after waterborne and dietborne exposure. J. Hazard. Mater. 2012, 221–222, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, D.; Zhang, W.; Zhang, C.; Zhang, L. The dynamic changes of arsenic bioaccumulation and antioxidant responses in the marine medaka Oryzias melastigma during chronic exposure. Aquat. Toxicol. 2019, 212, 110–119. [Google Scholar] [CrossRef]
- Suhendrayatna; Ohki, A.; Nakajima, T.; Maeda, S. Studies on the accumulation and transformation of arsenic in freshwater organisms I. Accumulation, transformation and toxicity of arsenic compounds on the Japanese Medaka, Oryzias latipes. Chemosphere 2002, 46, 319–324. [Google Scholar] [CrossRef]
- Hua, M.; Zhang, S.; Pan, B.; Zhang, W.; Lv, L.; Zhang, Q. Heavy metal removal from water/wastewater by nanosized metal oxides: A review. J. Hazard. Mater. 2012, 211–212, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Amde, M.; Liu, J.; Tan, Z.Q.; Bekana, D. Transformation and bioavailability of metal oxide nanoparticles in aquatic and terrestrial environments: A review. Environ. Pollut. 2017, 230, 250–267. [Google Scholar] [CrossRef]
- Fan, W.; Cui, M.; Liu, H.; Wang, C.; Shi, Z.; Tan, C.; Yang, X. Nano-TiO2 enhances the toxicity of copper in natural water to Daphnia magna. Environ. Pollut. 2011, 159, 729–734. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, J.; Cai, Z. TiO2 nanoparticles in the marine environment: Impact on the toxicity of tributyltin to abalone (Haliotis diversicolor supertexta) embryos. Environ. Sci. Technol. 2011, 45, 3753–3758. [Google Scholar] [CrossRef]
- Lu, J.; Tian, S.; Lv, X.; Chen, Z.; Chen, B.; Zhu, X.; Cai, Z. TiO2 nanoparticles in the marine environment: Impact on the toxicity of phenanthrene and Cd2+ to marine zooplankton Artemia salina. Sci. Total Environ. 2018, 615, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Chandra Sekhar, K.; Chary, N.S.; Kamala, C.T.; Suman Raj, D.S.; Sreenivasa Rao, A. Fractionation studies and bioaccumulation of sediment-bound heavy metals in Kolleru lake by edible fish. Environ. Int. 2004, 29, 1001–1008. [Google Scholar] [CrossRef]
- Zhao, S.; Meng, F.; Fu, H.; Xiao, J.; Gao, Y. Metallothionein levels in gills and visceral mass of Ruditapes philippinarum exposed to sublethal doses of cadmium and copper. In Proceedings of the International Conference on Challenges in Environmental Science and Computer Engineering, CESCE, Wuhan, China, 6–7 March 2010; Volume 2. [Google Scholar] [CrossRef]
- Hwang, P.P.; Lee, T.H. New insights into fish ion regulation and mitochondrion-rich cells. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Rosen, B.P. Families of arsenic transporters. Trends Microbiol. 1999, 7, 207–212. [Google Scholar] [CrossRef]
- Sörös, C.; Bodó, E.T.; Fodor, P.; Morabito, R. The potential of arsenic speciation in molluscs for environmental monitoring. Anal. Bioanal. Chem. 2003, 377, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Vahter, M. Mechanisms of arsenic biotransformation. Toxicology 2002, 181–182, 211–217. [Google Scholar] [CrossRef]
- Bagnyukova, T.V.; Luzhna, L.I.; Pogribny, I.P.; Lushchak, V.I. Oxidative stress and antioxidant defenses in goldfish liver in response to short-term exposure to arsenite. Environ. Mol. Mutagen. 2007, 48, 658–659. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Bhattacharya, S. Induction of oxidative stress by arsenic in Clarias batrachus: Involvement of peroxisomes. Ecotoxicol. Environ. Saf. 2007, 66, 178–187. [Google Scholar] [CrossRef]
- Sampayo-Reyes, A.; Zakharyan, R.A. Inhibition of human glutathione s-transferase omega by tocopherol succinate. Biomed. Pharmacother. 2006, 60, 238–244. [Google Scholar] [CrossRef]
- Vasken Aposhian, H.; Zakharyan, R.A.; Avram, M.D.; Sampayo-Reyes, A.; Wollenberg, M.L. A review of the enzymology of arsenic metabolism and a new potential role of hydrogen peroxide in the detoxication of the trivalent arsenic species. Toxicol. Appl. Pharmacol. 2004, 198, 327–335. [Google Scholar] [CrossRef]
- Griffitt, R.J.; Weil, R.; Hyndman, K.A.; Denslow, N.D.; Powers, K.; Taylor, D.; Barber, D.S. Exposure to copper nanoparticles causes gill injury and acute lethality in zebrafish (Danio rerio). Environ. Sci. Technol. 2007, 41, 8178–8186. [Google Scholar] [CrossRef] [PubMed]
- Heinlaan, M.; Ivask, A.; Blinova, I.; Dubourguier, H.C.; Kahru, A. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria Vibrio fischeri and crustaceans Daphnia magna and Thamnocephalus platyurus. Chemosphere 2008, 71, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Unfried, K.; Albrecht, C.; Klotz, L.O.; Von Mikecz, A.; Grether-Beck, S.; Schins, R.P.F. Cellular responses to nanoparticles: Target structures and mechanisms. Nanotoxicology 2007, 1, 52–71. [Google Scholar] [CrossRef]
- Han, J.; Won, E.J.; Hwang, D.S.; Rhee, J.S.; Kim, I.C.; Lee, J.S. Effect of copper exposure on GST activity and on the expression of four GSTs under oxidative stress condition in the monogonont rotifer, Brachionus koreanus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 158, 91–100. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Qian, W.; Ning, Z.; Zhu, X. Enhanced Bioaccumulation and Toxicity of Arsenic in Marine Mussel Perna viridis in the Presence of CuO/Fe3O4 Nanoparticles. Nanomaterials 2021, 11, 2769. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11102769
Zhou S, Qian W, Ning Z, Zhu X. Enhanced Bioaccumulation and Toxicity of Arsenic in Marine Mussel Perna viridis in the Presence of CuO/Fe3O4 Nanoparticles. Nanomaterials. 2021; 11(10):2769. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11102769
Chicago/Turabian StyleZhou, Shuang, Wei Qian, Zigong Ning, and Xiaoshan Zhu. 2021. "Enhanced Bioaccumulation and Toxicity of Arsenic in Marine Mussel Perna viridis in the Presence of CuO/Fe3O4 Nanoparticles" Nanomaterials 11, no. 10: 2769. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11102769