
Low Concentrations of Silver Nanoparticles Inhibit Spore Germination and Disturb Gender Differentiation of Ceratopteris thalictroides (L.) Brongn


Abstract
:1. Introduction
2. Materials and Methods
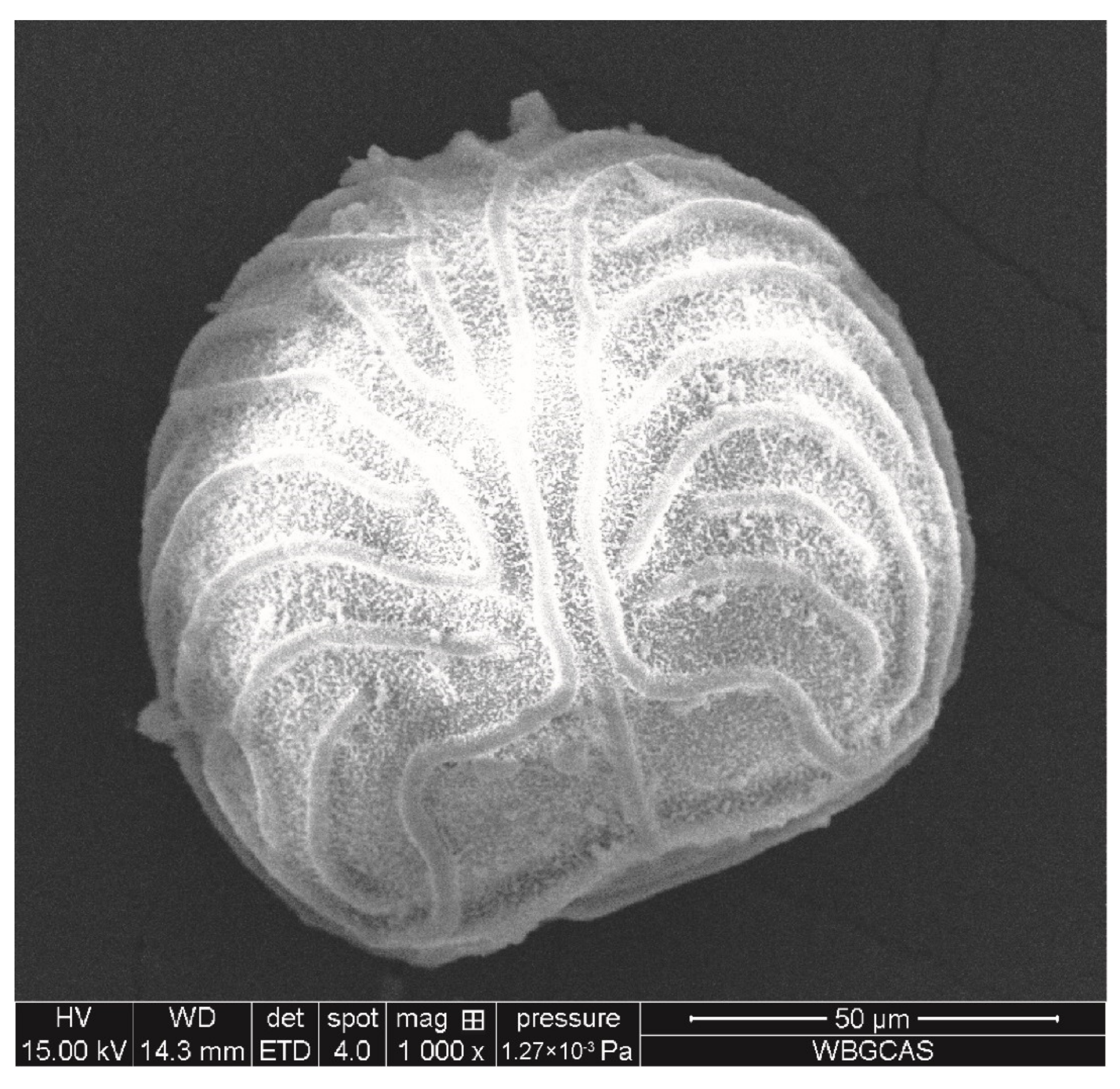
2.1. Plant Material
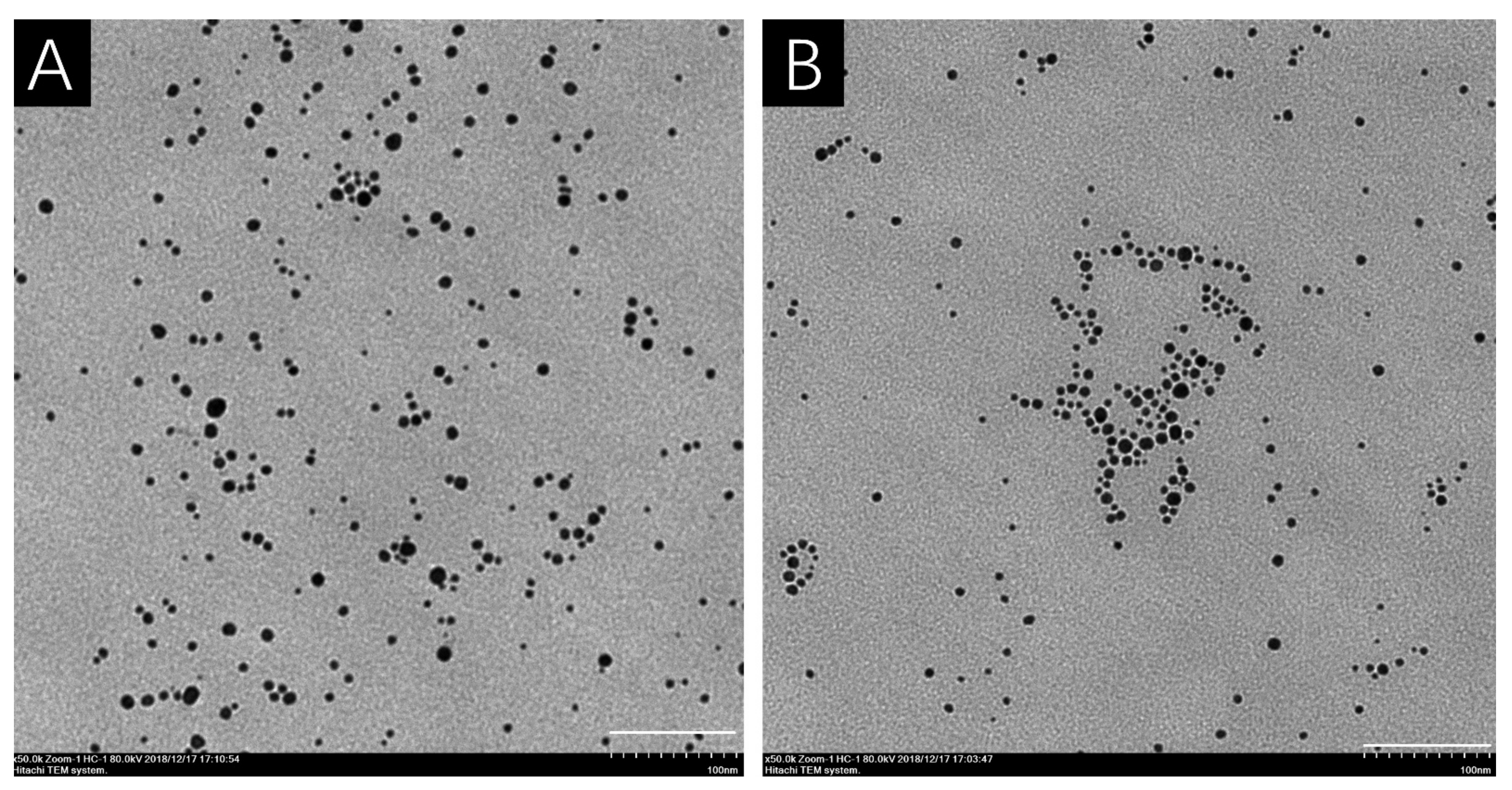
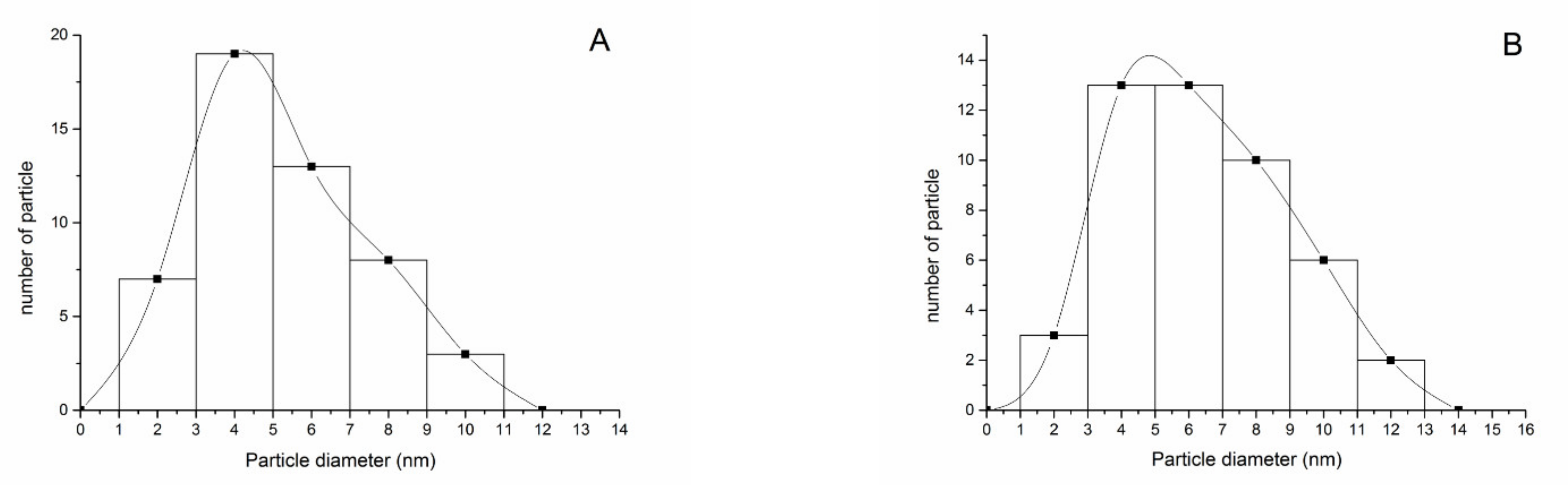
2.2. Characterization of AgNPs
2.3. Effects of AgNPs on Spore Germination and Gametophyte Differentiation in C. thalictroides
2.4. Toxicity Sources of AgNPs on Spore Germination and Gametophyte Differentiation in C. thalictroides
2.5. Statistics Analysis
3. Results
3.1. Characterization of AgNPs
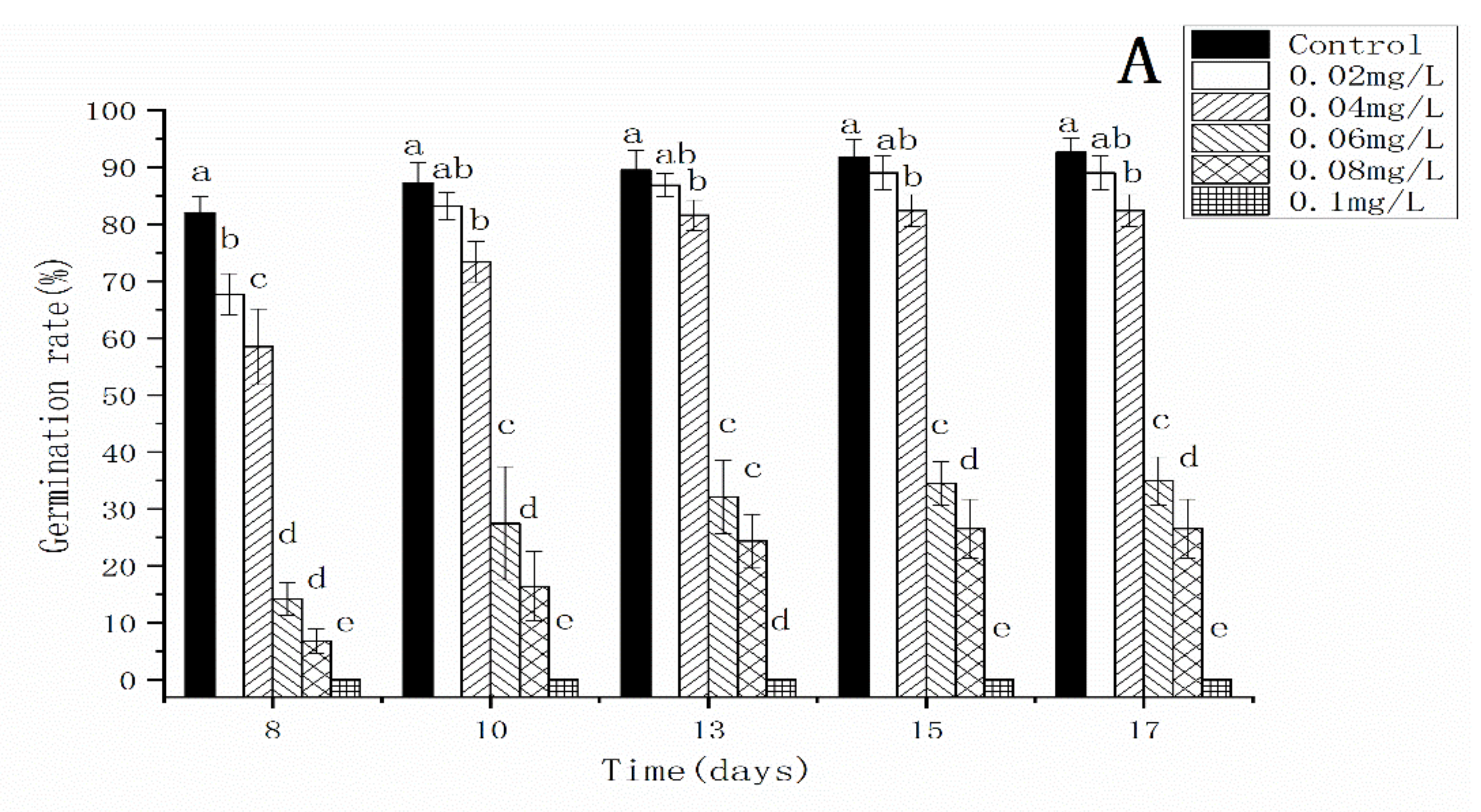
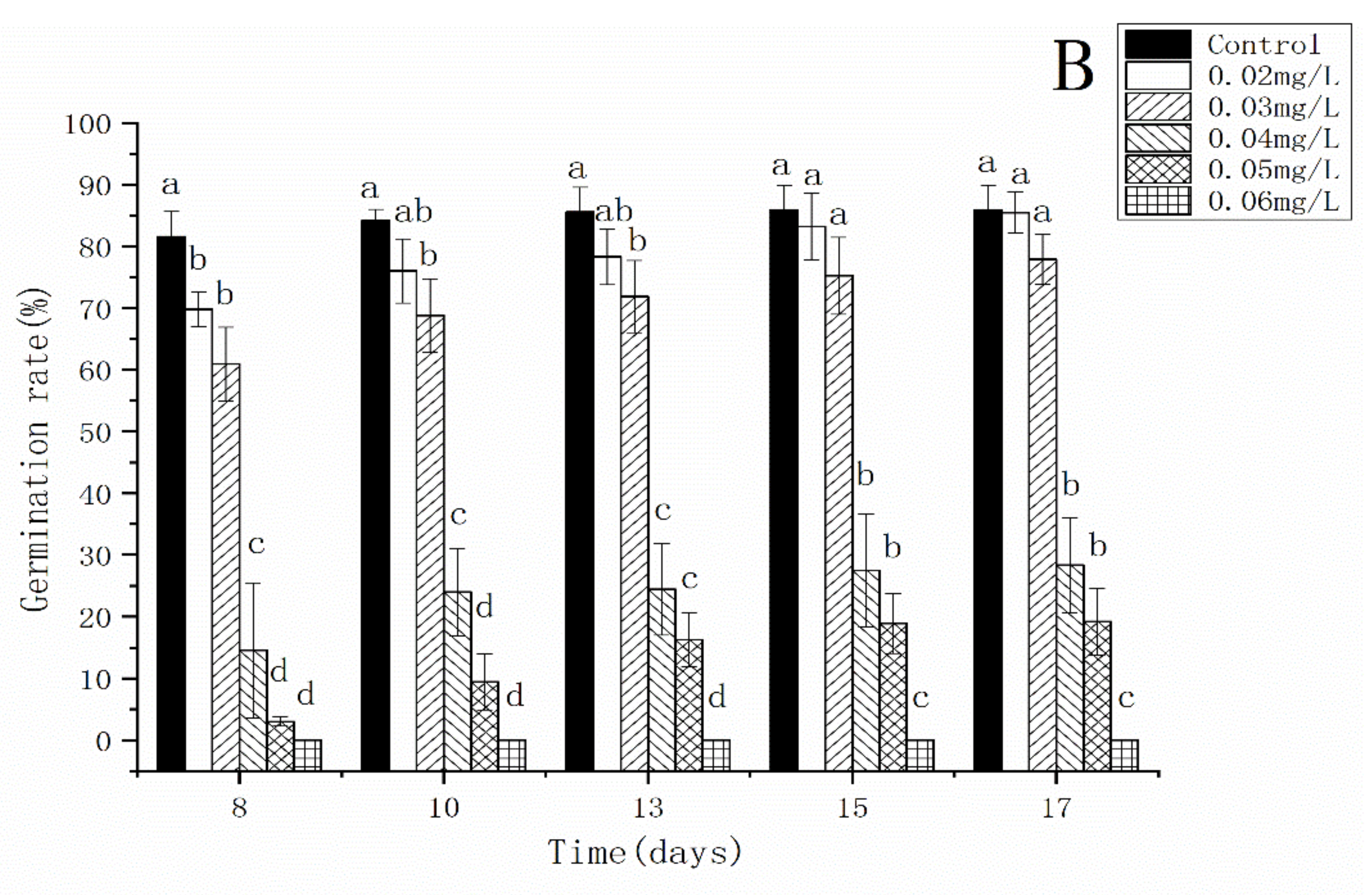
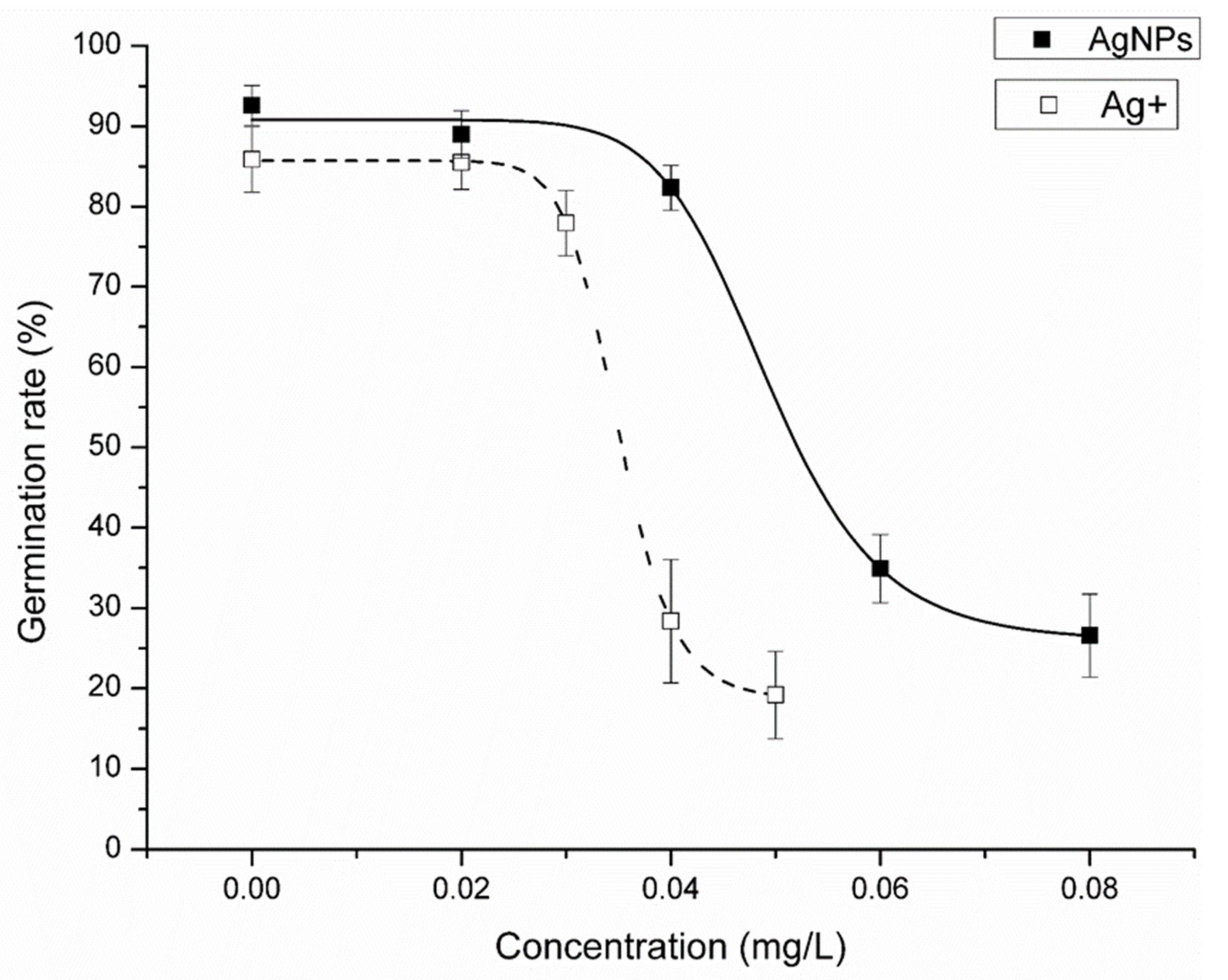
3.2. Effects of AgNPs and Ag+ on the Spore Germination Rate of C. thalictroides
3.3. Effects of AgNPs and Ag+ on Gametophytes of C. thalictroides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capek, I. Preparation of metal nanoparticles in water-in-oil (w/o) microemulsions. Adv. Colloid Interface Sci. 2004, 110, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Frattini, A.; Pellegri, N.; Nicastro, D.; Sanctis, O.D. Effect of amine groups in the synthesis of Ag nanoparticles using amino silanes. Mater. Chem. Phys. 2005, 94, 148–152. [Google Scholar] [CrossRef]
- Mueller, N.C.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanopart. Res. 2012, 14, 1109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu-Rui, L.; Robert, A.; Michael, R. Inventory of engineered nanoparticle-containing consumer products available in the Singapore retail market and likelihood of release into the aquatic environment. Int. J. Environ. Res. Public Health 2015, 12, 8717. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Liu, S.M.; Ko, C.L.; Chen, W.C. Advances of Hydroxyapatite Hybrid Organic Composite Used as Drug or Protein Carriers for Biomedical Applications: A Review. Polymers 2022, 14, 976. [Google Scholar] [CrossRef]
- Baldino, L.; Aragón, J.; Mendoza, G.; Irusta, S.; Cardea, S.; Reverchon, E. Production, characterization and testing of antibacterial PVA membranes loaded with HA-Ag3PO4 nanoparticles, produced by SC-CO2 phase inversion. J. Chem. Technol. Biotechnol. 2019, 94, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Prihandana, G.S.; Sriani, T.; Muthi’ah, A.D.; Machmudah, A.; Mahardika, M.; Miki, N. Study Effect of nAg Particle Size on the Properties and Antibacterial Characteristics of Polysulfone Membranes. Nanomaterials 2022, 12, 388. [Google Scholar] [CrossRef]
- Handy, R.D.; Cornelis, G.; Fernandes, T.; Tsyusko, O.; Decho, A.; Sabo-Attwood, T.; Metcalfe, C.; Steevens, J.A.; Klaine, S.J.; Koelmans, A.A. Ecotoxicity test methods for engineered nanomaterials: Practical experiences and recommendations from the bench. Environ. Toxicol. Chem. 2012, 31, 15–31. [Google Scholar] [CrossRef]
- Geranio, L.; Heuberger, M.; Nowack, B. The behavior of silver nanoparticles during washing. Environ. Sci. Technol. 2009, 43, 8113–8118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, F.; Sonderer, T.; Scholz, R.W.; Nowack, B. Modeled environmental concentrations of engineered nanomaterials (TiO2, ZnO, Ag, CNT, fullerenes) for different regions. Environ. Sci. Technol. 2009, 43, 9216–9222. [Google Scholar] [CrossRef] [PubMed]
- Edition, F. Guidelines for drinking-water quality. WHO Chron. 2011, 38, 104–108. [Google Scholar]
- Hund-Rinke, K.; Simon, M. Ecotoxic effect of photocatalytic active nanoparticles (TiO2) on algae and daphnids (8 pp). Environ. Sci. Pollut. Res. 2006, 13, 225–232. [Google Scholar] [CrossRef]
- Lee, W.F.; Huang, Y.C. Swelling and antibacterial properties for the superabsorbent hydrogels containing silver nanoparticles. J. Appl. Polym. Sci. 2007, 106, 1992–1999. [Google Scholar] [CrossRef]
- Shahverdi, A.R.; Fakhimi, A.; Shahverdi, H.R.; Minaian, S. Synthesis and effect of silver nanoparticles on the antibacterial activity of different antibiotics against Staphylococcus aureus and Escherichia coli. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 168–171. [Google Scholar] [CrossRef]
- Shoults-Wilson, W.A.; Reinsch, B.C.; Tsyusko, O.V.; Bertsch, P.M.; Lowry, G.V.; Unrine, J.M. Effect of silver nanoparticle surface coating on bioaccumulation and reproductive toxicity in earthworms (Eisenia fetida). Nanotoxicology 2011, 5, 432–444. [Google Scholar] [CrossRef]
- Xiu, Z.; Zhang, Q.; Puppala, H.L.; Colvin, V.L.; Alvarez, P. Negligible particle-specific antibacterial activity of silver nanoparticles. Nano Lett. 2012, 12, 4271–4275. [Google Scholar] [CrossRef]
- Zhao, C.M.; Wang, W.X. Importance of surface coatings and soluble silver in silver nanoparticles toxicity to Daphnia magna. Nanotoxicology 2012, 6, 361–370. [Google Scholar] [CrossRef]
- Yin, L.Y.; Cheng, Y.; Colman, B.P.; Auffan, M.; Wiesner, M.; Bernhardt, E.S. More than the ions: The effects of silver nanoparticles on Lolium multiflorum. Environ. Sci. Technol. 2011, 45, 2360–2367. [Google Scholar] [CrossRef]
- Jiang, H.S.; Ming, L.; Chang, F.Y.; Wei, L.; Yin, L.Y. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 2012, 31, 1880–1886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Qiu, X.N.; Li, G.B.; Yin, L.Y. Silver nanoparticles induced the accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 2014, 33, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Yin, L.Y.; Ren, N.N.; Zhao, S.T.; Li, Z.; Zhi, Y.; Shao, H.; Li, W.; Gontero, B. Silver nanoparticles induced reactive oxygen species via photosynthetic energy transport imbalance in an aquatic plant. Nanotoxicology 2017, 11, 157–167. [Google Scholar] [CrossRef]
- Jiang, X.; Miclăuş, T.; Wang, L.; Foldbjerg, R.; Sutherland, D.S.; Autrup, H.; Chen, C.; Beer, C. Fast intracellular dissolution and persistent cellular uptake of silver nanoparticles in CHO-K1 cells: Implication for cytotoxicity. Nanotoxicology 2015, 9, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Wang, Q.; Cai, W.; Zhou, D.M.; Xing, B. Uptake kinetics of silver nanoparticles by plant: Relative importance of particles and dissolved ions. Nanotoxicology 2020, 14, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Zhang, Y.Z.; Lu, Z.W.; Lebrun, R.; Li, W. Interaction between Silver Nanoparticles and Two Dehydrogenases: Role of Thiol Groups. Small 2019, 15, 1900860. [Google Scholar] [CrossRef]
- Van Aerle, R.; Lange, A.; Moorhouse, A.; Paszkiewicz, K.; Ball, K.; Johnston, B.D.; De-Bastos, E.; Booth, T.; Tyler, C.R.; Santons, E.M. Molecular mechanisms of toxicity of silver nanoparticles in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 8005–8014. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, H.; Poortman, J.; Peters, R.J.; Wijma, E.; Kramer, E.; Makama, S.; Puspitaninganindita, K.; Marvin, H.J.P.; Peijnenburg, A.A.C.M.; Hendriksen, P.J.M. Characterization of translocation of silver nanoparticles and effects on whole-genome gene expression using an in vitro intestinal epithelium coculture model. ACS Nano 2011, 5, 4091–4103. [Google Scholar] [CrossRef]
- Sakamoto, M.; Ha, J.Y.; Yoneshima, S.; Kataoka, C.; Tatsuta, H.; Kashiwada, S. Free silver ion as the main cause of acute and chronic toxicity of silver nanoparticles to cladocerans. Arch. Environ. Contam. Toxicol. 2015, 68, 500–509. [Google Scholar] [CrossRef]
- Shen, M.H.; Zhou, X.X.; Yang, X.Y.; Chao, J.B.; Liu, R.; Liu, J.F. Exposure medium: Key in identifying free Ag+ as the exclusive species of silver nanoparticles with acute toxicity to Daphnia magna. Sci. Rep. 2015, 5, 9674. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.D. Phytotoxicity of Silver Nanoparticles to Arabidopsis thaliana in Hydroponic and Soil Systems; Southern Illinois University at Carbondale: Carbondale, IL, USA, 2011. [Google Scholar]
- Peng, X.F. The Effects of Silver Nanoparticles on the Vegetative Growth and Flowing of Arabidopsis thaliana. Master’s Thesis, Zhejiang University of Technology, Hangzhou, China, 2013. [Google Scholar]
- Wu, D.; Fan, W.; Kishen, A.; Gutmann, J.L.; Bing, F. Evaluation of the antibacterial efficacy of silver nanoparticles against Enterococcus faecalis, biofilm. J. Endod. 2014, 40, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Akaighe, N.; Depner, S.W.; Banerjee, S.; Gutmann, J.L.; Bing, F. The effects of monovalent and divalent cations on the stability of silver nanoparticles formed from direct reduction of silver ions by Suwannee River humic acid/natural organic matter. Sci. Total Environ. 2012, 441, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Herth, S. Plant nanotoxicology. Trends Plant Sci. 2011, 16, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Xiao, W.B. On the development of gametophyte and sporophyte of Ceratopteris. J. Hunan Agric. Univ. 2008, 34, 306–310. [Google Scholar]
- Dai, X.L.; Jin, Q.; Wang, Q.X. Studies on the development of gametophyte of Ceratopteris thalictroides. Bull. Bot. Res. 2005, 25, 274–276. [Google Scholar]
- Zhan, Z.G.; Li, Y.J. Research progress on spore germination and gametophyte development of Water Fern. North Hortic. 2011, 9, 181–185. [Google Scholar]
- Chen, X. Study on the Interaction between Nano-Silver Particles and Chlamydomonas reinhardtii in Water Environment. Master’s Thesis, Shandong Agricultural University, Taian, China, 2019. [Google Scholar]
- Becaro, A.A.; Jonsson, C.M.; Puti, F.C.; Siqueira, M.C. Toxicity of PVA-stabilized silver nanoparticles to algae and microcrustaceans. Environ. Nanotechnol. Monit. Manag. 2015, 3, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.H.; Xu, W.Y.; Jiang, M.S.; Chen, C. The toxicity of nano-silver to 15 varieties plant pathogenic fungi. Chin. Agric. Sci. Bull. 2007, 23, 220–224. [Google Scholar]
- Yu, P.; Liu, H.; Mou, Z.; Lu, Z.; Li, H.; Zhuang, D.; He, H. Acute Toxic Effects of Silver Nanoparticles and Silver Ion on Two Microalgae. Asian J. Ecotoxicol. 2017, 12, 188–198. [Google Scholar]
- Pereira, S.; Jesus, F.; Aguiar, S.; Oliveira, R.D.; Nogueira, A. Phytotoxicity of silver nanoparticles to Lemna minor: Surface coating and exposure period-related effects. Sci. Total Environ. 2018, 618, 1389–1399. [Google Scholar] [CrossRef]
- Pham, T.L. Toxicity of Silver Nanoparticles to Tropical Microalgae Scenedesmus acuminatus, Chaetoceros gracilis and Crustacean Daphnia lumholtzi. Turk. J. Fish. Aquat. Sci. 2019, 19, 1009–1016. [Google Scholar] [CrossRef]
- Wang, Y. Neurotoxic Effects of Silver Nanoparticles on Early Life-Stage Development of Zebrafish (Danio rerio). Master’s Thesis, Shanxi University, Taiyuan, China, 2019. [Google Scholar]
- Lalau, C.M.; Simioni, C.; Vicentini, D.S.; Ouriques, L.C.; Matias, W. Toxicological effects of AgNPs on duckweed (Landoltia punctata). Sci. Total Environ. 2020, 710, 136318. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.T. Toxic Effects of Liposome-Encapsulated Silver (AgNPs, Ag+) on Daphnia magna. Master’s Thesis, China Mining University, Xuzhou, China, 2019. [Google Scholar]
- Lv, Y. A Study on the Cytotoxicity of Two Types of Nanoparticles on Ciliated Protozoa Pseudourostyla cristata. Master’s Thesis, East China Normal University, Shanghai, China, 2019. [Google Scholar]
- Tan, C.L. The Biological Effect of Nanomaterials on Bacteria. Master’s Thesis, Tianjin University of Technology, Tianjin, China, 2012. [Google Scholar]
- Chasan, R. Ceratopteris: A model plant for the 90s. Plant Cell 1992, 4, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Tao, L. Effects of Herbicides on Sexual Reproduction of Ceratopteris pteridoides and Vallisneria natans. Master’s Thesis, Graduate University of Chinese Academy of Sciences (Wuhan Botanical Garden), Wuhan, China, 2007. [Google Scholar]
- Yuan, W.; Zhou, Y.; Liu, X.; Wang, J. New Perspective on the Nanoplastics Disrupting the Reproduction of an Endangered Fern in Artificial Freshwater. Environ. Sci. Technol. 2019, 53, 12715–12724. [Google Scholar] [CrossRef]
- Austin, C.A.; Umbreit, T.H.; Brown, K.M.; Barber, D.S.; Dair, B.J.; Francke-Carroll, S.; Feswick, A.; Saint-Louis, M.A.; Hikawa, H.; Siebein, K.N. Distribution of silver nanoparticles in pregnant mice and developing embryos. Nanotoxicology 2012, 6, 912–922. [Google Scholar] [CrossRef]
- Chatterjee, N.; Bhattacharjee, B. Revelation of ZnS Nanoparticles Induces Follicular Atresia and Apoptosis in the Ovarian Preovulatory Follicles in the Catfish Mystus tengara (Hamilton, 1822). Scientifica 2016, 2016, 3927340. [Google Scholar] [CrossRef] [Green Version]
- Yeon-Jin, L.; Jong-Hye, C.; Pil-Je, K.; Kyung-Hee, C.; Su-Hyon, K.; Woo-Chan, S.; Kwang-Sik, P. A transfer of silver nanoparticles from pregnant rat to offspring. Toxicol. Res. 2012, 28, 139–141. [Google Scholar]
- Liu, X.Q.; Zhang, H.F.; Zhang, W.D.; Zhang, P.F.; Hao, Y.N.; Song, R.; Li, L.; Feng, Y.N.; Hao, Z.H.; Shen, W. Regulation of neuroendocrine cells and neuron factors in the ovary by zinc oxide nanoparticles. Toxicol. Lett. 2016, 256, 19–32. [Google Scholar] [CrossRef]
- Morishita, Y.; Yoshioka, Y.; Takimura, Y.; Shimizu, Y.; Namba, Y.; Nojiri, N.; Ishizaka, T.; Takao, K.; Yamashita, F.; Takuma, K. Distribution of silver nanoparticles to breast milk and their biological effects on breast-fed offspring mice. ACS Nano 2016, 10, 8180–8191. [Google Scholar] [CrossRef]
- Tabatabaei, S.R.F.; Moshrefi, M.; Askaripour, M. Prenatal exposure to silver nanoparticles causes depression-like responses in mice. Indian J. Pharm. Sci. 2015, 77, 681. [Google Scholar]
- Zhang, W.D.; Zhao, Y.; Zhang, H.F.; Wang, S.K.; Hao, Z.H.; Liu, J.; Yuan, Y.Q.; Zhang, P.F.; Yang, H.D.; Shen, W. Alteration of gene expression by zinc oxide nanoparticles or zinc sulfate in vivo and comparison with in vitro data: A harmonious case. Theriogenology 2016, 86, 850–861.e1. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.J.; Lin, L.; Wei, A.; Sepulveda, M.S. Protein Corona Analysis of Silver Nanoparticles Exposed to Fish Plasma. Environ. Sci. Technol. Lett. 2017, 4, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.M.; Yau, S.K.W.; Lok, C.N.; So, M.H.; Che, C.M. Oxidative dissolution of silver nanoparticles by biologically relevant oxidants: A kinetic and mechanistic study. Chem. Asian J. 2010, 5, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lenhart, J.J.; Walker, H.W. Dissolution-accompanied aggregation kinetics of silver nanoparticles. Langmuir 2010, 26, 16690–16698. [Google Scholar] [CrossRef]
- Ma, R.; Levard, C.; Marinakos, S.M.; Cheng, Y.; Lowry, G.V. Size-controlled dissolution of organic-coated silver nanoparticles. Environ. Sci. Technol. 2012, 46, 752–759. [Google Scholar] [CrossRef]
- Ferreira, L.A.; Dos Reis, S.B.; da Silva, E.D.N.; Cadore, S.; da Silva Bernardes, J.; Duran, N.; de Jesus, M.B. Thiol-antioxidants interfere with assessing silver nanoparticle cytotoxicity. Nanomedicine 2020, 24, 102130. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter (μm) | Density (g/cm3) | Weight (mg) | Number of Spores per Milligram |
|---|---|---|---|
| 105.68 ± 3.83 | 1.05 ± 0.04 | 0.41 ± 0.03 × 10−6 | 2.45 ± 0.15 × 106 |
| Solution | Core Diameter * (nm) | Hydrodynamic Diameter * (nm) | PDI | Zeta Potential (mV) |
|---|---|---|---|---|
| H2O | 6.2 ± 2.0 | 20.3 ± 3.1 | 0.637 | −10.7 ± 0.4 a |
| 10% Hoagland’s solution | 7.8 ± 2.7 | 27.1 ± 4.1 | 0.791 | −2.1 ± 0.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Yin, L.; Li, W.; Jiang, H.-S. Low Concentrations of Silver Nanoparticles Inhibit Spore Germination and Disturb Gender Differentiation of Ceratopteris thalictroides (L.) Brongn. Nanomaterials 2022, 12, 1730. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12101730
Lu Z, Yin L, Li W, Jiang H-S. Low Concentrations of Silver Nanoparticles Inhibit Spore Germination and Disturb Gender Differentiation of Ceratopteris thalictroides (L.) Brongn. Nanomaterials. 2022; 12(10):1730. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12101730
Chicago/Turabian StyleLu, Zhenwei, Liyan Yin, Wei Li, and Hong-Sheng Jiang. 2022. "Low Concentrations of Silver Nanoparticles Inhibit Spore Germination and Disturb Gender Differentiation of Ceratopteris thalictroides (L.) Brongn" Nanomaterials 12, no. 10: 1730. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12101730