

Liposomal Formulations Loaded with a Eugenol Derivative for Application as Insecticides: Encapsulation Studies and In Silico Identification of Protein Targets

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Viability Assessment
2.3. Inverted Virtual Screening (IVS) Studies
2.4. Molecular Dynamics Simulations and Free Energy Calculations
2.5. Nanoencapsulation Studies and Release Assays
2.6. Statistical Analysis
3. Results and Discussion
3.1. Toxicity of Eugenol Derivatives towards Insect Cells Have a Time-Dependent Effect
3.2. Inverted Virtual Screening Results
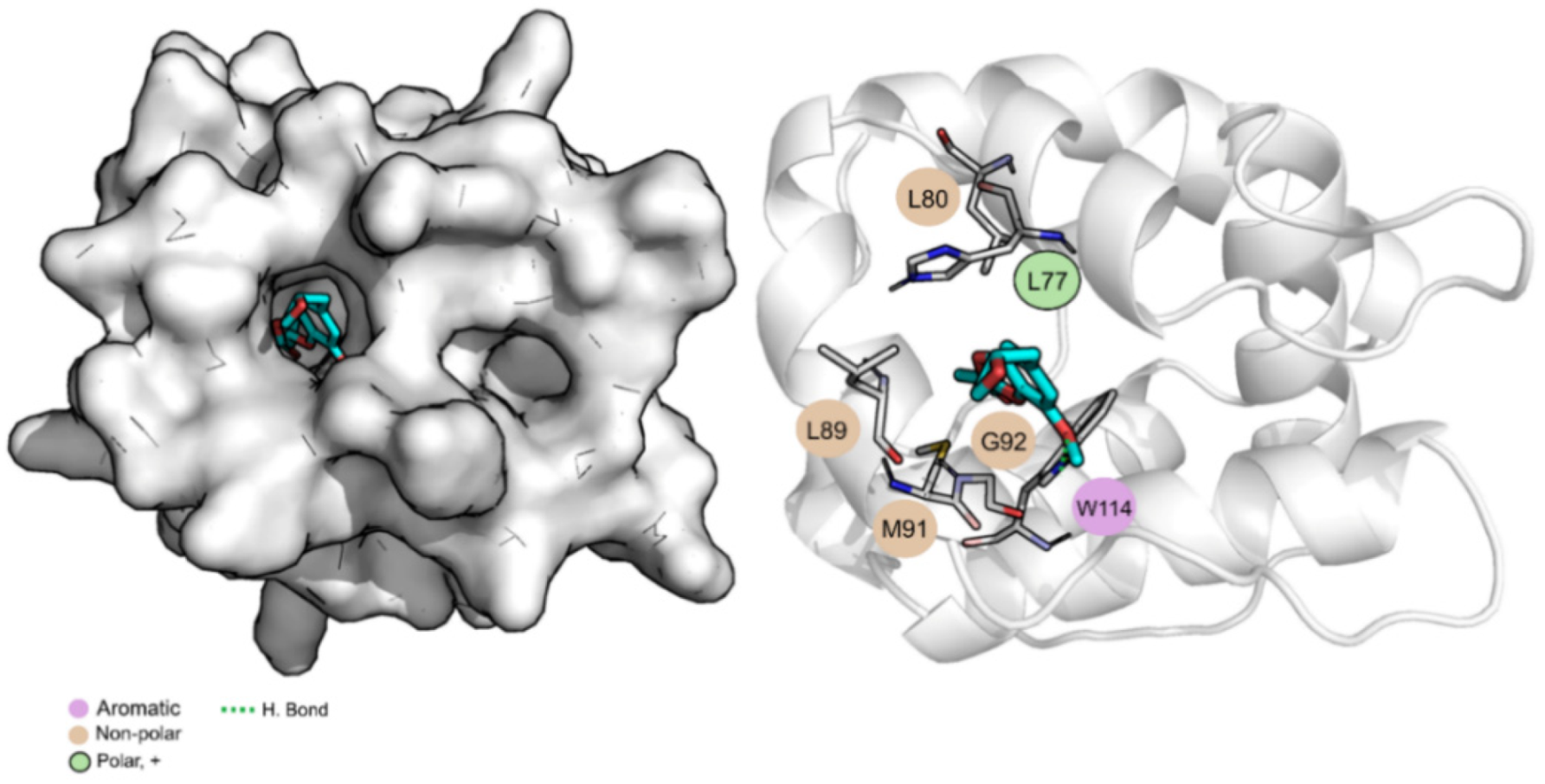
3.3. Molecular Dynamics Simulations and Free Energy Calculations Results
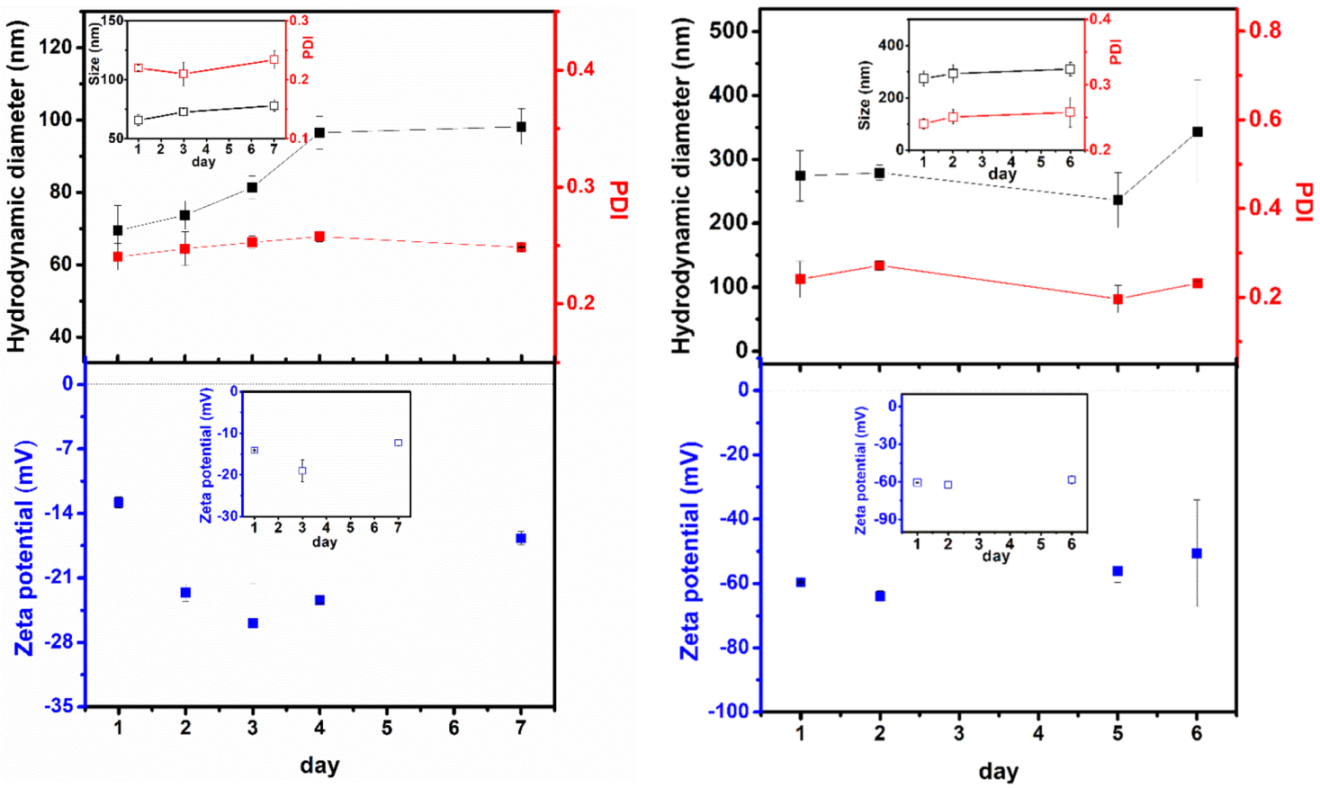
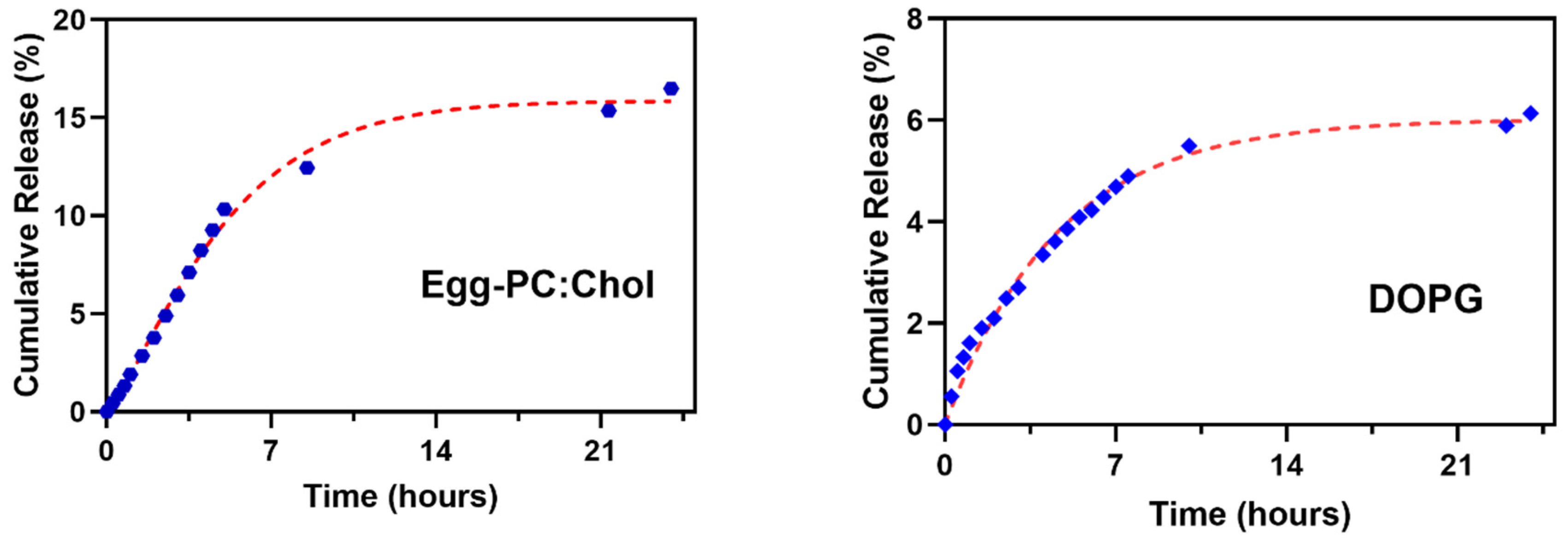
3.4. Nanoencapsulation Assays in Liposomal Formulations
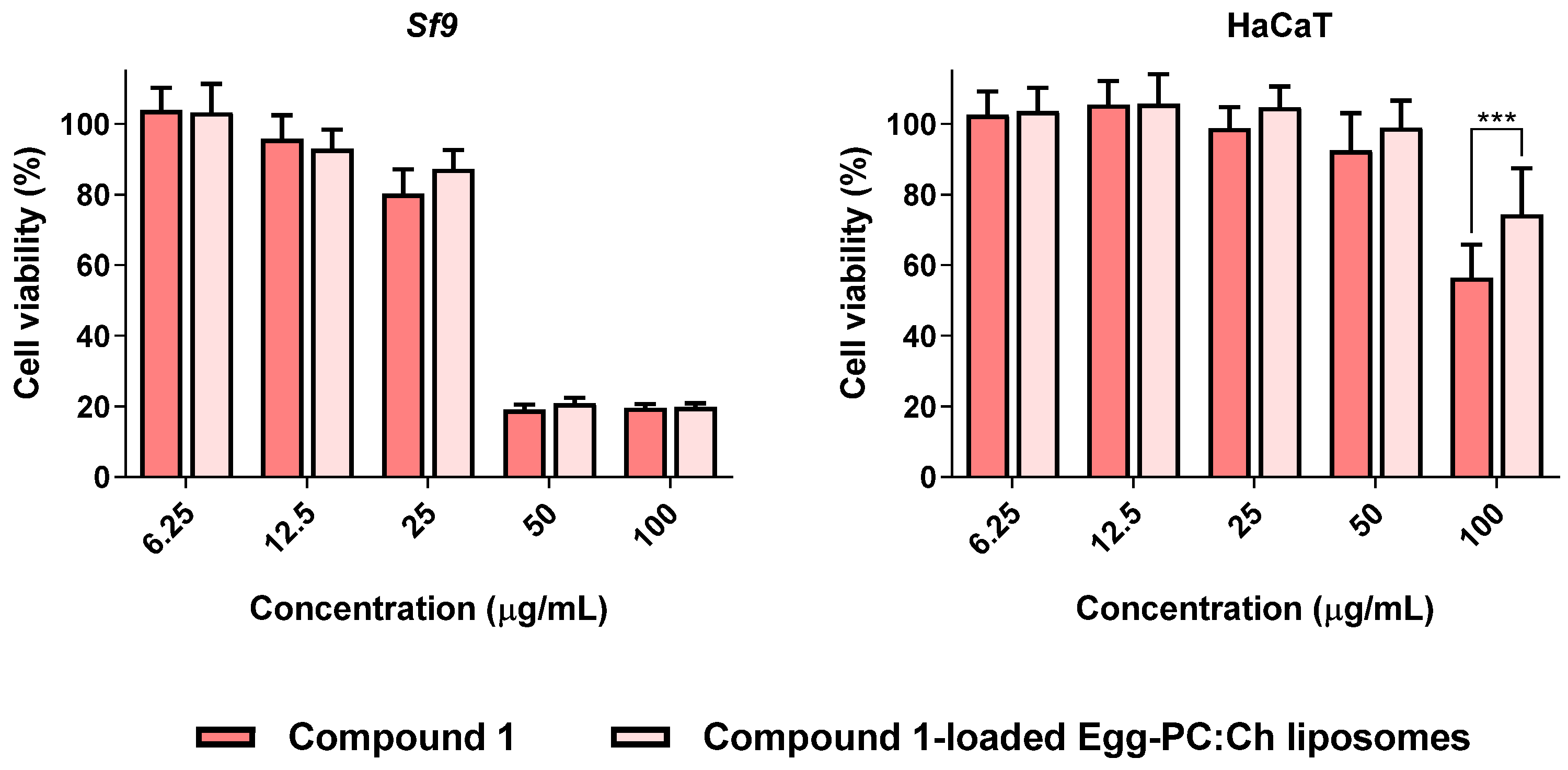
3.5. Compound 1-Loaded Egg-PC:Ch Liposomes Maintained Insecticidal Activity with Decreased Toxicity towards Human Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khallaf, A.; Wang, P.; Zhuo, S.; Zhu, H.; Liu, H. Synthesis, insecticidal activities, and structure–activity relationships of 1,3,4-oxadiazole-ring-containing pyridylpyrazole-4-carboxamides as novel insecticides of the anthranilic diamide family. J. Heterocycl. Chem. 2021, 58, 2189–2202. [Google Scholar] [CrossRef]
- Abdou, A.; Elmakssoudi, A.; El Amrani, A.; Jamal Eddine, J.; Dakir, M. Recent advances in chemical reactivity and biological activities of eugenol derivatives. Med. Chem. Res. 2021, 30, 1011–1030. [Google Scholar] [CrossRef]
- Guenette, S.A.; Beaudry, F.; Marier, J.F.; Vachon, P. Pharmacokinetics and anesthetic activity of eugenol in male Sprague-Dawley rats. J. Vet. Pharm. Ther. 2006, 29, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Danie, A.N.; Sartoretto, S.M.; Schmidt, G.; Caparroz-Assef, S.M.; Bersani-Amado, C.A.; Cuman, R.K.N. Anti-inflammatory and antinociceptive activities of eugenol essential oil in experimental animal models. Braz. J. Pharmacogn. 2009, 19, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Lahlou, S.; Figueiredo, A.F.; Magalhães, P.J.C.; Leal-Cardoso, J.H.; Gloria, P.D. Cardiovascular effects of methyleugenol, a natural constituent of many plant essential oils, in normotensive rats. Life Sci. 2004, 74, 2401–2412. [Google Scholar] [CrossRef]
- Zheng, G.Q.; Kenney, P.M.; Lam, L.K.T. Sesquiterpenes from clove (Eugenia caryophyllata) as potential anticarcinogenic agents. J. Nat. Prod. 1992, 55, 999–1003. [Google Scholar] [CrossRef]
- Hidalgo, M.E.; De la Rosa, C.; Carrasco, H.; Cardona, W.; Gallardo, C.; Espinoza, L. Antioxidant capacity of eugenol derivatives. Quim. Nova. 2009, 32, 1467–1470. [Google Scholar] [CrossRef]
- Barceloux, D.G. Medical toxicology of natural substances. In Foods, Fungi, Medicinal Herbs, Plants and Venomous Animals; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008. [Google Scholar] [CrossRef]
- Machado, M.; Dinis, A.M.; Salgueiro, L.; Custódio, J.B.A.; Cavaleiro, C.; Sousa, M.C. Anti-Giardia activity of Syzygium aromaticum essential oil and eugenol: Effects on growth, viability, adherence and ultrastructure. Exp. Parasitol. 2011, 127, 732–739. [Google Scholar] [CrossRef]
- Wang, C.; Fan, Y. Eugenol enhances the resistance of tomato against tomato yellow leaf curl virus. J. Sci. Food Agric. 2014, 94, 677–682. [Google Scholar] [CrossRef]
- Santos, T.D.; Coelho, C.M.; Elias, T.C.; Siqueira, F.S.; Nora, E.S.; Campos, M.; de Souza, G.A.; Coelho, L.F.; Carvalho, D.T. Synthesis and biological evaluation of new eugenol-derived 1,2,3-triazoles as antimycobacterial agents. J. Braz. Chem. Soc. 2019, 30, 1425–1436. [Google Scholar] [CrossRef]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and Dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef]
- Yadav, M.K.; Chae, S.W.; Im, G.J.; Chung, J.W.; Song, J.J. Eugenol: A phyto-compound effective against methicillin-resistant and methicillin-sensitive Staphylococcus aureus clinical strain biofilms. PLoS ONE 2015, 10, e0119564. [Google Scholar] [CrossRef] [Green Version]
- da Camara, C.A.G.; Doboszewski, B.; Melo, J.B.R.; Nazarenko, A.Y.; dos Santos, R.B.; Moraes, M.M. Novel Insecticides from Alkylated and Acylated Derivatives of Thymol and Eugenol for the Control of Plutella xylostella (Lepidoptera: Plutellidae). J. Braz. Chem. Soc. 2022, 33, 196–204. [Google Scholar] [CrossRef]
- Fernandes, M.J.G.; Pereira, R.B.; Pereira, D.M.; Fortes, A.G.; Castanheira, E.M.S.; Gonçalves, M.S.T. New Eugenol Derivatives with Enhanced Insecticidal Activity. Int. J. Mol. Sci. 2020, 21, 9257. [Google Scholar] [CrossRef]
- El Asbahani, A.; Miladi, K.; Badri, W.; Sala, M.; Aït Addi, E.H.; Casabianca, H.; El Mousadik, A.; Hartmann, D.; Jilale, A.; Renaud, F.N.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef]
- Pinho, E.; Grootveld, M.; Soares, G.; Henriques, M. Cyclodextrins as encapsulation agents for plant bioactive compounds. Carbohydr. Polym. 2014, 101, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal formulations in clinical use: An updated review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Shi, F.; Zhao, J.-H.; Liu, Y.; Wang, Z.; Zhang, Y.-T.; Feng, N.-P. Preparation and characterization of solid lipid nanoparticles loaded with frankincense and myrrh oil. Int. J. Nanomed. 2012, 7, 2033–2043. [Google Scholar] [CrossRef] [Green Version]
- Liolios, C.C.; Gortzi, O.; Lalas, S.; Tsaknis, J.; Chinou, I. Liposomal incorporation of carvacrol and thymol isolated from the essential oil of Origanum dictamnus L. and in vitro antimicrobial activity. Food Chem. 2009, 112, 77–83. [Google Scholar] [CrossRef]
- Varona, S.; Martín, A.; Cocero, M.J. Liposomal Incorporation of Lavandin Essential Oil by a Thin-Film Hydration Method and by Particles from Gas-Saturated Solutions. Ind. Eng. Chem. Res. 2011, 50, 2088–2097. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Rahman, M.M.; Liu, Y.; Naidu, R. Nanoencapsulation, Nano-guard for Pesticides: A New Window for Safe Application. J. Agric. Food Chem. 2016, 64, 1447–1483. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.B.; Pinto, N.F.S.; Fernandes, M.J.G.; Vieira, T.F.; Rodrigues, A.R.O.; Pereira, D.M.; Sousa, S.F.; Castanheira, E.M.S.; Fortes, A.G.; Gonçalves, M.S.T. Amino Alcohols from Eugenol as Potential Semisynthetic Insecticides: Chemical, Biological, and Computational Insights. Molecules 2021, 26, 6616. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yin, B.; Cappelle, K.; Swevers, L.; Smagghe, G.; Yang, X.; Zhang, L. Identification of novel agonists and antagonists of the ecdysone receptor by virtual screening. J. Mol. Graph. Model. 2018, 81, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Nakagawa, Y.; Ogura, T.; Yamada, Y.; Ohe, T.; Miyagawa, H. Virtual screening for ligands of the insect molting hormone receptor. J. Chem. Inf. Model. 2011, 51, 296–305. [Google Scholar] [CrossRef]
- Dong, Y.; Jiang, X.; Liu, T.; Ling, Y.; Yang, Q.; Zhang, L.; He, X. Structure-Based Virtual Screening, Compound Synthesis, and Bioassay for the Design of Chitinase Inhibitors. J. Agric. Food Chem. 2018, 66, 3351–3357. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Yao, Y.; Wang, J.; Li, Y.; Li, G.; Wang, Y. Identification of novel potential β-N-acetyl-D-hexosaminidase inhibitors by virtual screening, molecular dynamics simulation and MM-PBSA calculations. Int. J. Mol. Sci. 2012, 13, 4545–4563. [Google Scholar] [CrossRef]
- Dong, L.; Shen, S.; Xu, Y.; Wang, L.; Yang, Q.; Zhang, J.; Lu, H. Identification of novel insect β-N-acetylhexosaminidase OfHex1 inhibitors based on virtual screening, biological evaluation, and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2021, 39, 1735–1743. [Google Scholar] [CrossRef]
- Min, J.; Lin, D.; Zhang, Q.; Zhang, J.; Yu, Z. Structure-based virtual screening of novel inhibitors of the uridyltransferase activity of Xanthomonas oryzae pv. oryzae GlmU. Eur. J. Med. Chem. 2012, 53, 150–158. [Google Scholar] [CrossRef]
- Ramos, R.D.S.; Costa, J.D.S.; Silva, R.C.; Costa, G.V.; Rodrigues, A.B.L.; Rabelo, E.M.; Souto, R.N.P.; Taft, C.A.; Silva, C.H.T.P.; Rosa, J.M.C.; et al. Identification of Potential Inhibitors from Pyriproxyfen with Insecticidal Activity by Virtual Screening. Pharmaceuticals 2019, 12, 20. [Google Scholar] [CrossRef]
- Riva, C.; Suzanne, P.; Charpentier, G.; Dulin, F.; Halm-Lemeille, M.-P.; Santos, J.S.-O. In silico chemical library screening and experimental validation of novel compounds with potential varroacide activities. Pestic. Biochem. Physiol. 2019, 160, 11–19. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Jiang, H.; Deng, J. Crystal structure of Manduca sexta prophenoloxidase provides insights into the mechanism of type 3 copper enzymes. Proc. Natl. Acad. Sci. USA 2009, 106, 17002–17006. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Liu, Y.X.; Kang, T.; Sun, Y.N.; Li, J.Z.; Ye, F. Identification of novel inhibitors of p-hydroxyphenylpyruvate dioxygenase using receptor-based virtual screening. J. Taiwan Inst. Chem. Eng. 2019, 103, 33–34. [Google Scholar] [CrossRef]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Cristofori-Armstrong, B.; Smith, J.J.; Chin, Y.K.Y.; Lei, J.; Zhou, Q.; et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 2018, 362, eaau2596. [Google Scholar] [CrossRef] [Green Version]
- Offermann, L.R.; Chan, S.L.; Osinski, T.; Tan, Y.W.; Chew, F.T.; Sivaraman, J.; Mok, Y.-K.; Minor, W.; Chruszcz, M. The major cockroach allergen Bla g 4 binds tyramine and octopamine. Mol. Immunol. 2014, 60, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Du, X.; Wang, C.; Lin, J.; Du, X. Identification of Potential Helicoverpa armigera (Lepidoptera: Noctuidae) Sterol Carrier Protein-2 Inhibitors Through High-Throughput Virtual Screening. J. Econ. Entomol. 2017, 110, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Joshi, T.; Sharma, P.; Chandra, S.; Pande, V. Molecular docking and molecular dynamics simulation approach to screen natural compounds for inhibition of Xanthomonas oryzae pv. Oryzae by targeting peptide deformylase. J. Biomol. Struct. Dyn. 2021, 39, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Correy, G.J.; Zaidman, D.; Harmelin, A.; Carvalho, S.; Mabbitta, P.D.; Calaora, V.; James, P.J.; Kotz, A.C.; Jackson, C.J.; London, N. Overcoming insecticide resistance through computational inhibitor design. Proc. Natl. Acad. Sci. USA 2019, 116, 21012–21021. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliferenko, P.V.; Oliferenko, A.A.; Poda, G.I.; Osolodkin, D.I.; Pillai, G.G.; Bernier, U.R.; Tsikolia, M.; Agramonte, M.N.; Clark, G.G.; Linthicum, K.J.; et al. Promising Aedes aegypti repellent chemotypes identified through integrated QSAR, virtual screening, synthesis, and bioassay. PLoS ONE 2013, 8, e64547. [Google Scholar] [CrossRef] [PubMed]
- Lill, M.A.; Danielson, M.L. Computer-aided drug design platform using PyMOL. J. Comput. Aided Mol. Des. 2011, 25, 13–19. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading, J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sander, T.; Freyss, J.; von Korff, M.; Rufener, C. DataWarrior: An open-source program for chemistry aware data visualization and analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Natal, C.M.; Fernandes, M.J.G.; Pinto, N.F.S.; Pereira, R.B.; Vieira, T.F.; Rodrigues, A.R.O.; Pereira, D.M.; Sousa, S.F.; Fortes, A.G.; Castanheira, E.M.S.; et al. New carvacrol and thymol derivatives as potential insecticides: Synthesis, biological activity, computational studies and nanoencapsulation. RSC Adv. 2021, 11, 34024–34035. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, D.A.; Cheatham III, T.E.; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comp. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Frisch, M.; Trucks, G.; Schlegel, H.E.A.; Scuseria, G.W.; Robb, M.A.; Cheeseman, J.R.; Montgomery, J.A., Jr.; Vreven, T.K.K.N.; Kudin, K.N.; Burant, J.C.; et al. Gaussian 09, Revision A.02; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Magalhães, R.P.; Vieira, T.F.; Melo, A.; Sousa, S.F. Identification of novel candidates for inhibition of LasR, a quorum-sensing receptor of multidrug resistant Pseudomonas aeruginosa, through a specialized multi-level in silico approach. Mol. Syst. Des. Eng. 2022, 7, 434–446. [Google Scholar] [CrossRef]
- Vieira, T.F.; Magalhães, R.P.; Simões, M.; Sousa, S.F. Drug Repurposing Targeting Pseudomonas aeruginosa MvfR Using Docking, Virtual Screening, Molecular Dynamics, and Free-Energy Calculations. Antibiotics 2022, 11, 185. [Google Scholar] [CrossRef]
- Vieira, T.F.; Martins, F.G.; Moreira, J.P.; Barbosa, T.; Sousa, S.F. In Silico Identification of Possible Inhibitors for Protein Kinase B (PknB) of Mycobacterium tuberculosis. Molecules 2021, 26, 6162. [Google Scholar] [CrossRef]
- Martins, F.G.; Melo, A.; Sousa, S.F. Identification of new potential inhibitors of quorum sensing through a specialized multi-level computational approach. Molecules 2021, 26, 2600. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Jaafar-Maalej, C.; Diab, R.; Andrieu, V.; Elaissari, A.; Fessi, H. Ethanol injection method for hydrophilic and lipophilic drug-loaded liposome preparation. J. Liposome Res. 2010, 20, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Noyes, A.A.; Whitney, W.R. The rate of solution of solid substances in their own solutions. J. Am. Chem. Soc. 1987, 19, 930–934. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, V.; Kosmidis, K.; Vlachou, M.; Macheras, P. On the use of the Weibull function for the discernment of drug release mechanisms. Int. J. Pharm. 2006, 309, 44–50. [Google Scholar] [CrossRef]
- Simão, A.M.S.; Bolean, M.; Cury, T.A.C.; Stabeli, R.G.; Itri, R.; Ciancaglini, P. Liposomal systems as carriers for bioactive compounds. Biophys Rev. 2015, 7, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Kondratowicz, A.; Weiss, M.; Juzwa, W.; Majchrzycki, Ł.; Lewandowicz, G. Characteristics of liposomes derived from egg yolk. Open Chem. 2019, 17, 763–778. [Google Scholar] [CrossRef]
- Crisma, M.; Barazza, A.; Formaggio, F.; Kaptein, B.; Broxterman, Q.B.; Kamphuis, J.; Toniolo, C. Peptaibolin: Synthesis, 3D-structure, and membrane modifying properties of the natural antibiotic and selected analogues. Tetrahedron 2001, 57, 2813–2825. [Google Scholar] [CrossRef]
- Castro, V.I.B.; Carvalho, C.M.; Fernandes, R.V.D.; Pereira-Lima, S.M.M.A.; Castanheira, E.M.S.; Costa, S.P.G. Peptaibolin analogues by incorporation of α,α-dialkylglycines: Synthesis and study of their membrane permeating ability. Tetrahedron 2016, 72, 1024–1030. [Google Scholar] [CrossRef]
- Vincent, J.S.; Revak, S.D.; Cochrane, C.D.; Levin, I.W. Interactions of model human pulmonary surfactants with a mixed phospholipid bilayer assembly: Raman spectroscopic studies. Biochemistry 1993, 32, 8228–8238. [Google Scholar] [CrossRef]
- Dargel, C.; Gräbitz-Bräuer, F.; Geisler, R.; Fandrich, P.; Hannappel, Y.; Porcar, L.; Hellweg, T. Stable DOPG/Glycyrrhizin Vesicles with a Wide Range of Mixing Ratios: Structure and Stability as Seen by Scattering Experiments and Cryo-TEM. Molecules 2021, 26, 4959. [Google Scholar] [CrossRef]
- Abreu, A.S.; Hermenegildo, B.F.C.; Ferreira, P.M.T.; Queiroz, M.J.R.P.; Castanheira, E.M.S. Interaction of fluorescent quinolin-2-one and coumarin derivatives including dipeptides with lipid bilayers. RSC Adv. 2016, 6, 72141–72148. [Google Scholar] [CrossRef]









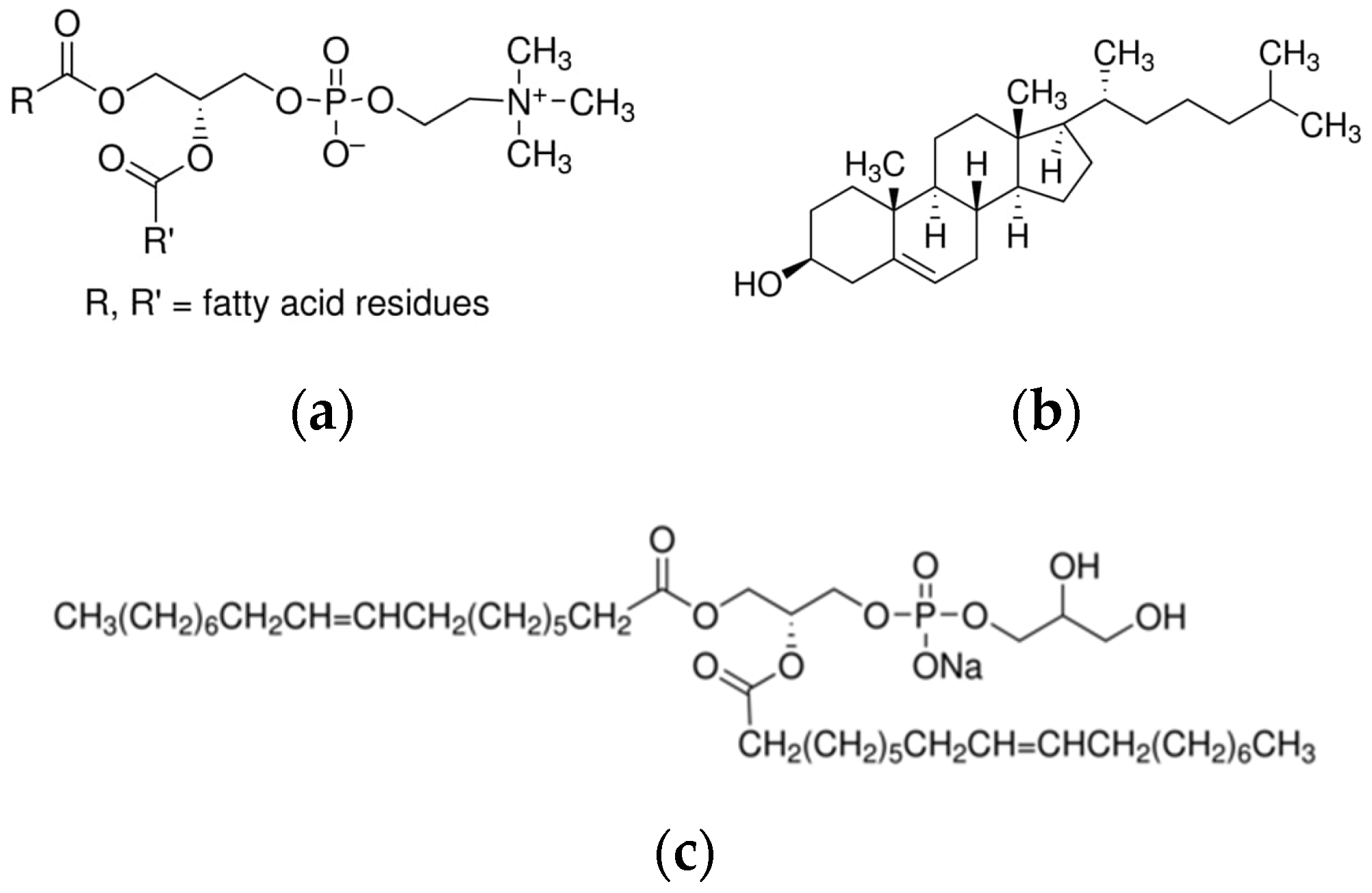{kind=link}
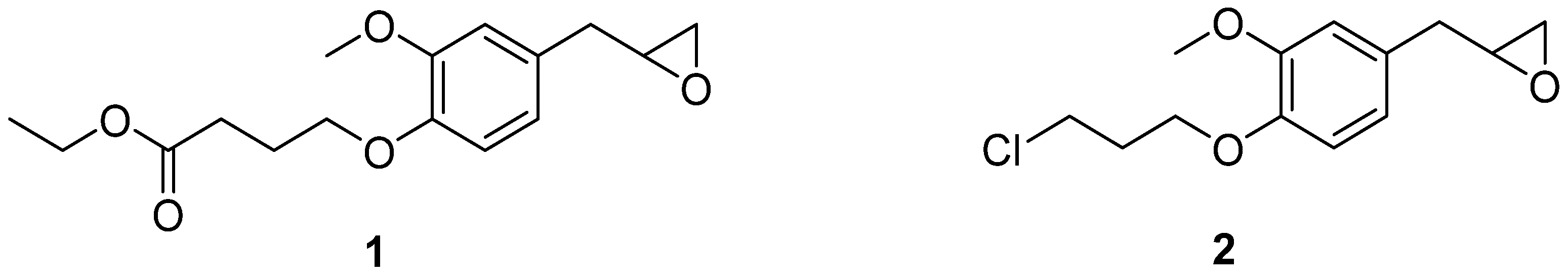{kind=link}
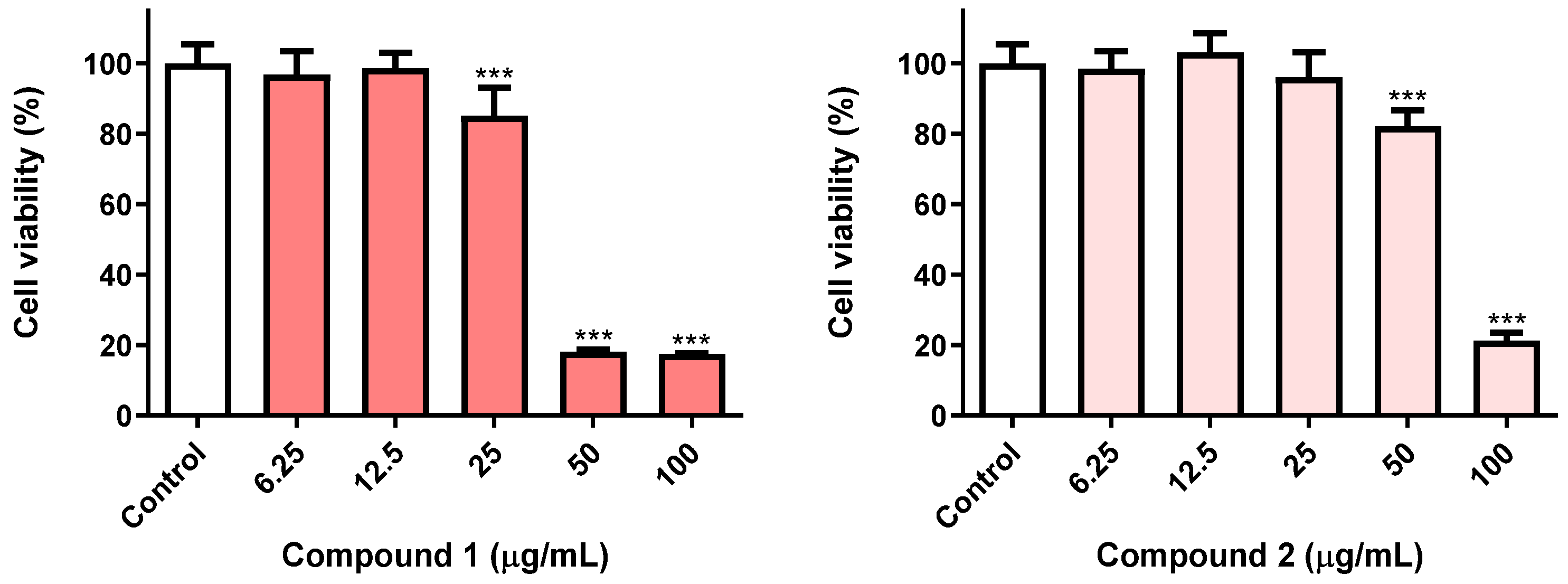{kind=link}
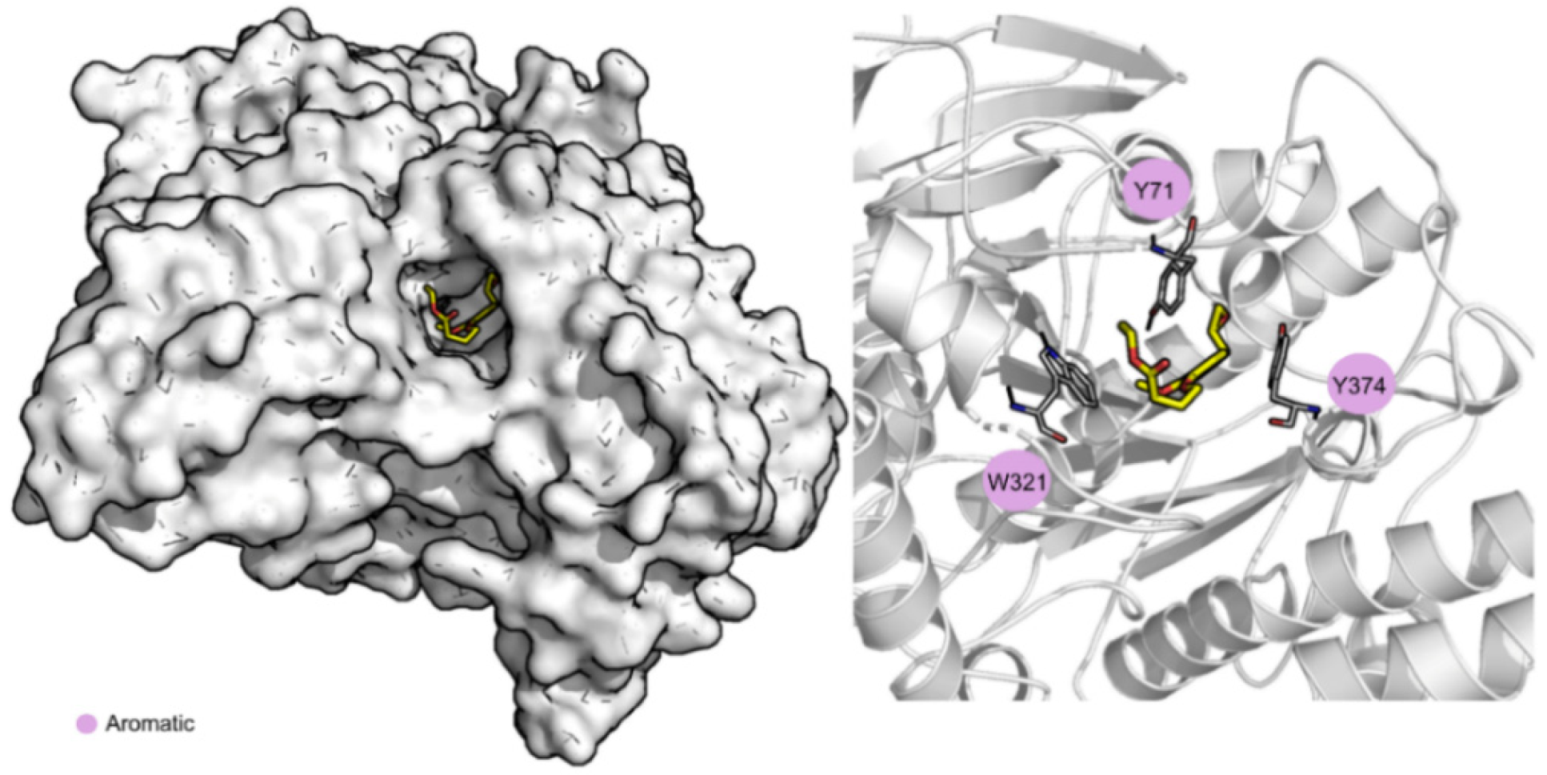{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Organism | PDB Target | Resolution (Å) | Description | Ref. |
|---|---|---|---|---|---|
| Ecdysone receptor | Heliothis virescens | 1R20 | 3.00 | VS based on 1R20 bound to an agonist as a model for the development of a receptor-based pharmacophore model. | [24] |
| 1R1K | 2.90 | VS of 2 million compounds against 1R1K, an ecdysone receptor structure bound to its known ligand ponasterone A. | [25] | ||
| Chitinase | Ostrinia furnacalis | 3WL1 | 1.77 | Pharmacophore-based screening using two crystal structures of chitinases: 3WL1 bound to its reaction product and 3WQV bound to an inhibitor. | [26] |
| 3WQV | 2.04 | ||||
| β-N-Acetyl-D-hexosaminidase OfHex1 | 3NSN | 2.10 | VS of the ZINC database to identify OfHex1 inhibitors using 3NSN crystal structure bound to a known inhibitor. | [27] | |
| 3OZP | 2.00 | VS of the ZINC database targeting 3OZP, a crystal structure of OfHex1 bound to an inhibitor. | [28] | ||
| N-Acetylgluco-samine-1-phosphate uridyltransferase (GlmU) | Xanthomonas oryzae | 2V0K | 2.30 | Homology model built for docking using 2V0K and 2VD4 as templates. The 2V0K crystal structure is bound to its known ligand and 2VD4 is bound to a possible inhibitor. | [29] |
| 2VD4 | 1.90 | ||||
| Acetyl- cholinesterase | Aedes aegypti | 1QON | 2.72 | Search for new molecules with insecticidal activity against Ae. Aegypti using acetylcholinesterase crystal structures 1QON and 4EY6 as targets, both bound to possible inhibitors. | [30] |
| 4EY6 | 2.40 | ||||
| Drosophila melanogaster | 1DX4 | 2.70 | Homology 3D model built for VS using 1DX4 as a template. 1DX4 crystal structure is bound to a potent inhibitor. | [31] | |
| Polyphenol- oxidase | Manduca sexta | 3HSS | 2.70 | Crystal structure of a prophenoloxidase from Manduca sexta. | [32] |
| p-Hydroxyphenyl-pyruvate dioxygenase | Arabidopsis thaliana | 6ISD | 2.40 | Development of a receptor-ligand pharmacophore model based on the crystal structure 6ISD bound to a commonly used pesticide. The best model created was then used for VS studies. | [33] |
| Voltage-gated sodium channel | Periplaneta americana | 6A95 | 2.60 | Crystallographic structure of a voltage-gated sodium channel, NavPaS, bound to a pore blocker, tetrodotoxin (TTX) | [34] |
| Octopamine receptor | Blattella germanica | 4N7C | 1.75 | Crystal structure of Bla g 4, an octopamine receptor, bound to tyramine. | [35] |
| Sterol carrier protein-2 (HaSCP-2) | Helicoverpa armigera | 4UEI | Solution NMR | Structure-based VS of a database of commercially available compounds to find potential inhibitors of HaSCP-2. The residues Phe53, Thr128, and Gln131 were selected for thebinding cavity. | [36] |
| Peptide deformylase | Xanthomonas oryzae | 5CY8 | 2.38 | Docking and VS of a library of 318 phytochemicals. The 5CY8 crystal structure is bound to a possible inhibitor. | [37] |
| Alpha-esterase-7 (αE7) | Lucilia cuprina | 5TYJ | 1.75 | Computational design of potent and selective covalent inhibitors of αE7. The 5TYJ and 5TYP crystal structures are bound to inhibitors: (3-bromo-5-phenoxylphenyl)boronic acid and (3-bromo-4-methylphenyl)boronic acid, respectively. | [38] |
| 5TYP | 1.88 | ||||
| Odorant Binding Protein | Aedes aegypti | 5V13 | 1.84 | Search for new molecules with insecticidal activity against Ae. Aegypti using a crystal structure of a mosquito juvenile hormone-binding protein, 5V13, bound to its natural hormone. | [30] |
| Drosophila melanogaster | 2GTE | 1.40 | The 2GTE crystal structure is bound to its natural ligand | [39] | |
| Anopheles gambiae | 3N7H | 1.60 | QSAR and docking studies for the rational design of mosquito repellents using the crystal structure 3K1E bound to a polyethylene glycol molecule. The 3N7H crystal structure is bound to a commonly used repellent. | [40] | |
| Aedes aegypti | 3K1E | 1.85 | [40] |
| Target | PDB | PLP | ASP | ChemScore | GoldScore | Vina | Overall Ranking |
|---|---|---|---|---|---|---|---|
| Acetylcholinesterase | 1QON | 81.19 | 59.26 | 37.75 | 69.05 | −8.30 | 1 |
| 1DX4 | 77.08 | 51.68 | 36.22 | 62.54 | −7.70 | ||
| 4EY6 | 76.18 | 48.57 | 32.96 | 59.66 | −7.60 | ||
| Alpha-esterase-7 | 5TYJ | 59.68 | 33.92 | 32.74 | 54.09 | −6.20 | 8 |
| 5TYP | 63.72 | 39.54 | 31.84 | 56.73 | −6.20 | ||
| Beta-N-acetyl-D-hexosaminidase OfHex1 | 3OZP | 71.22 | 49.42 | 29.92 | 63.78 | −7.30 | 4 |
| 3NSN | 76.67 | 54.51 | 32.45 | 67.93 | −6.50 | ||
| Chitinases | 3WQV | 68.53 | 49.59 | 31.24 | 65.25 | −6.60 | 3 |
| 3WL1 | 70.09 | 48.98 | 31.76 | 60.37 | −7.00 | ||
| Ecdysone receptor (EcR) | 1R1K | 66.83 | 31.02 | 32.21 | 61.61 | −7.70 | 5 |
| 1R20 | 63.97 | 29.6 | 29.98 | 58.93 | −6.80 | ||
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | 2V0K | 58.12 | 28.56 | 23.72 | 59.67 | −5.90 | 13 |
| 2VD4 | 52.43 | 25.36 | 20.76 | 48.33 | −5.20 | ||
| Octopamine receptor | 4N7C | 60.27 | 39.29 | 34.34 | 69.10 | −5.90 | 7 |
| Odorant Binding Protein | 2GTE | 70.46 | 39.20 | 30.71 | 66.39 | −6.30 | 2 |
| 3K1E | 83.01 | 44.98 | 37.85 | 66.89 | −5.90 | ||
| 5V13 | 80.20 | 49.37 | 40.18 | 63.84 | −7.70 | ||
| 3N7H | 74.95 | 39.20 | 30.71 | 66.39 | −6.30 | ||
| Peptide deformylase | 5CY8 | 72.08 | 30.11 | 25.48 | 71.01 | −7.00 | 6 |
| p-hydroxyphenylpyruvate dioxygenase | 6ISD | 68.60 | 33.78 | 26.22 | 51.19 | −7.00 | 9 |
| Polyphenol oxidase (PPO) | 3HHS | 62.37 | 34.09 | 29.03 | 66.32 | −5.70 | 11 |
| Sterol carrier protein-2 (HaSCP-2) | 4UEI | 64.27 | 32.22 | 31.10 | 50.51 | −6.20 | 10 |
| Voltage-gated sodium channel | 6A95 | 63.20 | 28.28 | 21.92 | 61.46 | −6.10 | 12 |
| Average RMSD of the Complex (Å) | Average RMSD of the Ligand (Å) | Ligand SASA (Å2) | Percentage of Potential Ligand SASA Buried (%) | Average Number H-Bonds | ΔGbind (kcal/mol) | Main Contributors (kcal/mol) | |
|---|---|---|---|---|---|---|---|
| AChE | 3.4 ± 0.3 | 1.7 ± 0.4 | 111.2 ± 53.2 | 79 | 0.1 ± 0.2 | −22.6 ± 0.2 | Tyr71 (−2.3 ± 0.9) |
| Trp321 (−1.6 ± 1.0) | |||||||
| Tyr374 (−1.5 ± 0.8) | |||||||
| OBP1 | 2.1 ± 0.3 | 1.2 ± 0.2 | 63.7 ± 16.3 | 88 | 0.01 ± 0.1 | −32.4 ± 0.2 | Met91 (−1.3 ± 0.5) |
| Gly92 (−1.8 ± 0.6) | |||||||
| Trp114 (−2.5 ± 0.6) |
| Liposomes | EE(%) ± SD(%) |
|---|---|
| Egg-PC:Ch (70:30) | 88.8 ± 2.7 |
| DOPG (100%) | 79.8 ± 2.6 |
| Liposomes | First-Order Kinetics | Weibull | |||
|---|---|---|---|---|---|
| k | R2 | b | a | R2 | |
| Egg-PC:Ch (70:30) | 0.16 | 0.988 | 0.1258 | 1.244 | 0.995 |
| DOPG (100%) | 0.21 | 0.985 | 0.2469 | 0.8457 | 0.9904 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, M.J.G.; Pereira, R.B.; Rodrigues, A.R.O.; Vieira, T.F.; Fortes, A.G.; Pereira, D.M.; Sousa, S.F.; Gonçalves, M.S.T.; Castanheira, E.M.S. Liposomal Formulations Loaded with a Eugenol Derivative for Application as Insecticides: Encapsulation Studies and In Silico Identification of Protein Targets. Nanomaterials 2022, 12, 3583. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12203583
Fernandes MJG, Pereira RB, Rodrigues ARO, Vieira TF, Fortes AG, Pereira DM, Sousa SF, Gonçalves MST, Castanheira EMS. Liposomal Formulations Loaded with a Eugenol Derivative for Application as Insecticides: Encapsulation Studies and In Silico Identification of Protein Targets. Nanomaterials. 2022; 12(20):3583. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12203583
Chicago/Turabian StyleFernandes, Maria José G., Renato B. Pereira, Ana Rita O. Rodrigues, Tatiana F. Vieira, A. Gil Fortes, David M. Pereira, Sérgio F. Sousa, M. Sameiro T. Gonçalves, and Elisabete M. S. Castanheira. 2022. "Liposomal Formulations Loaded with a Eugenol Derivative for Application as Insecticides: Encapsulation Studies and In Silico Identification of Protein Targets" Nanomaterials 12, no. 20: 3583. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12203583