
Phytogenic Selenium Nanoparticles Elicited the Physiological, Biochemical, and Antioxidant Defense System Amelioration of Huanglongbing-Infected ‘Kinnow’ Mandarin Plants

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area, Conditions and Selection of Specimens
2.2. Sample Collection, DNA Extraction, and Polymerase Chain Reaction (PCR) Protocol
2.3. Sequencing of PCR Products for Confirmation of Candidatus Liberibacter Asiaticus
2.4. SeNPs Preparation and Characterization
2.4.1. Preparation of Garlic Cloves Extract
2.4.2. Bio-Fabrication of SeNPs
2.4.3. Spectrophotometric Analysis of SeNPs
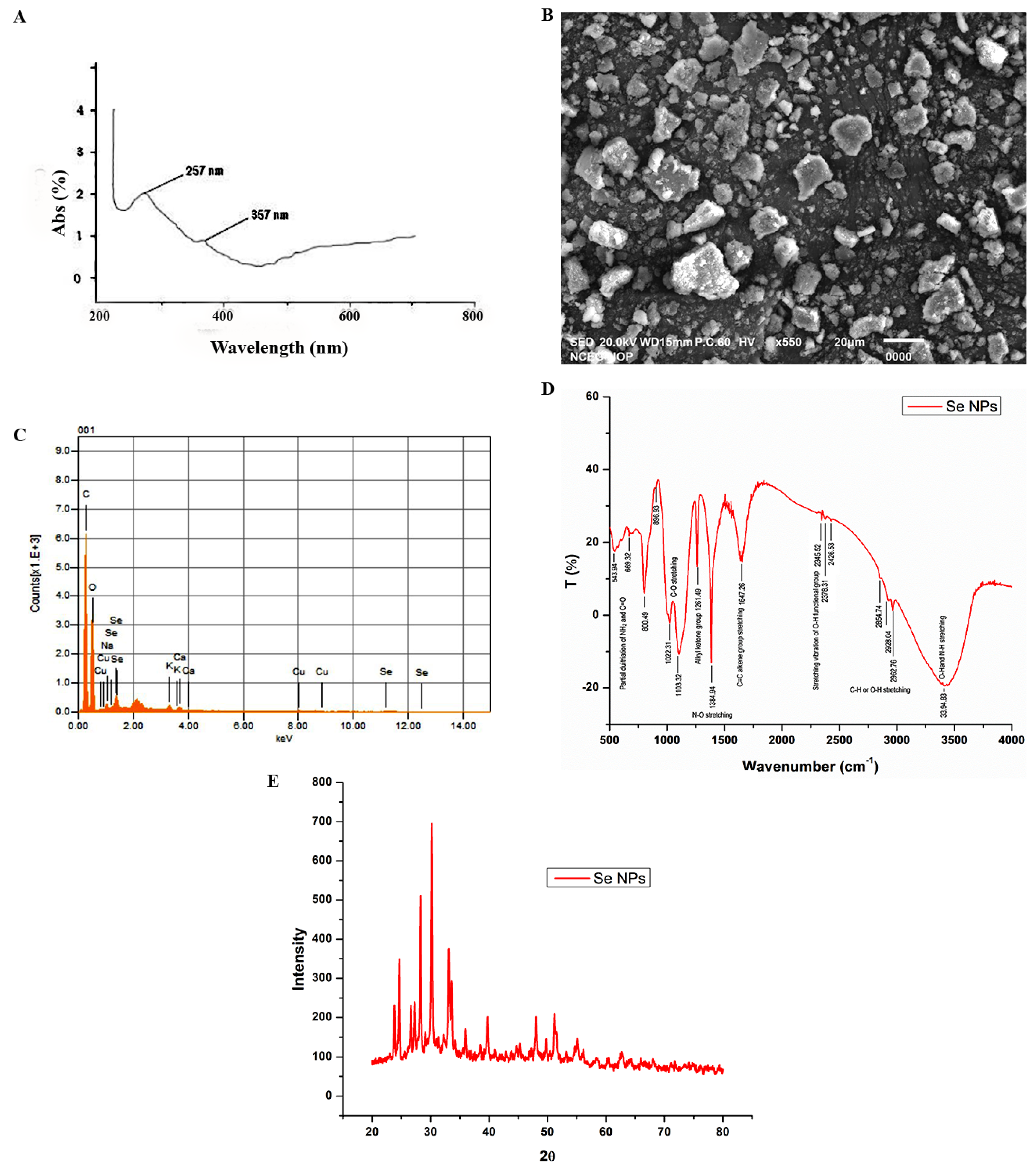
2.4.4. Scanning Electron Microscopic (SEM) Analysis of SeNPs
2.4.5. X-ray Diffraction Analysis (XRD)
2.4.6. Energy-Dispersive X-ray Spectroscopic Analysis (EDX) of SeNPs
2.4.7. Fourier Transform Infrared Spectroscopy (FTIR)
2.5. Selection and Application of Biogenic SeNPs and Experimental Plan
2.6. Measurement of Physiological Parameters
2.6.1. Chlorophyll a, Chlorophyll b, Total Chlorophyll, and Carotenoids Content
- A = Absorbance obtained at a specific wavelength;
- V = Final volume of Chl extract in 80% acetone;
- W = Weight of fresh tissue extracted.
2.6.2. Estimation of the Relative Water Content (RWC)
2.6.3. Estimation of Membrane Stability Index (MSI)
2.6.4. Estimation of Peroxidase (POD), Superoxide Dismutase (SOD), and Catalase (CAT)
2.6.5. Extraction of Flavonoid and Phenolic Contents from ‘Kinnow’ Mandarin Plants Tissues
Total Phenolic Content (TPC)
Total Flavonoids Content (TFC)
2.6.6. Estimation of Proline and Sugars Contents
2.6.7. Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2) Content
2.7. Statistical Analysis
3. Results
3.1. PCR Based Detection of Candidatus Liberibacter spp. in HLB-Infected ‘Kinnow’ Mandarin Plants
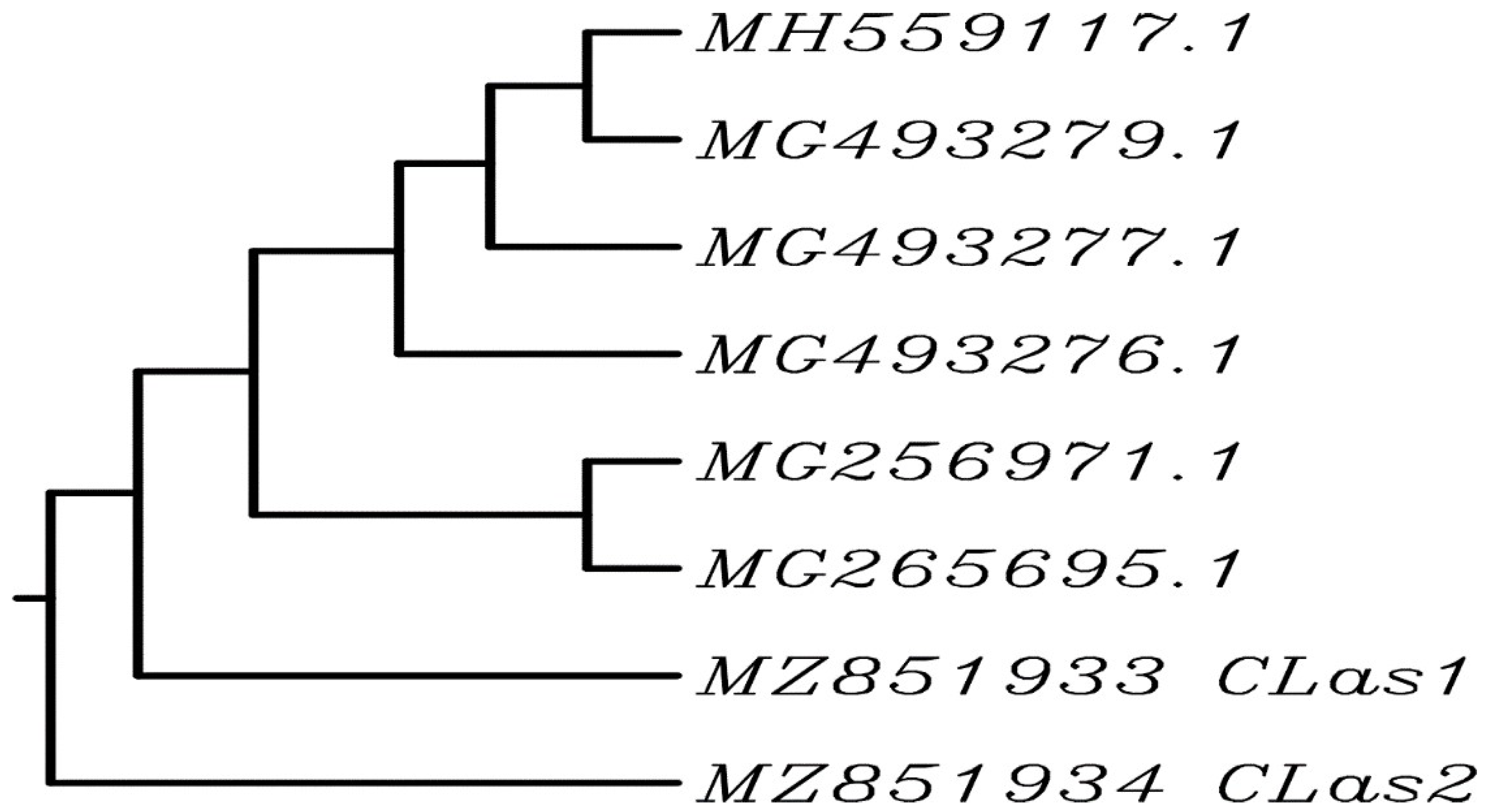
3.2. Sequencing Analysis
3.3. Morphological and Optical Characterization of Plant Mediated Selenium Nanoparticles (SeNPs)
3.4. Differential Impacts of Plant Mediated SeNPs on Physiological Profiling of HLB-Infected ‘Kinnow’ Mandarin Plants
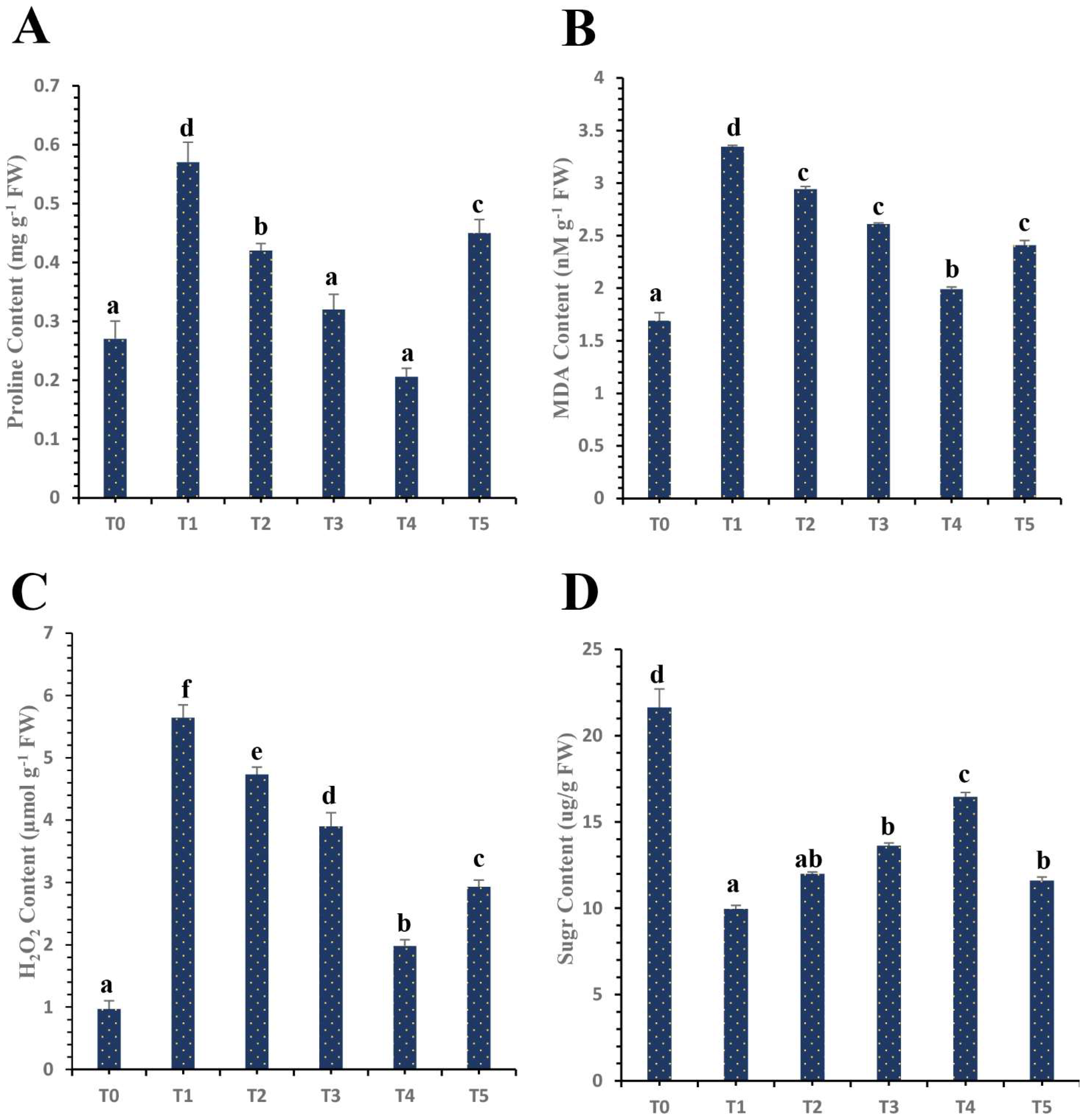
3.5. Differential Effects of Plant Mediated SeNPs on H2O2 Content and MDA Content of HLB-Diseased ‘Kinnow’ Mandarin Plants
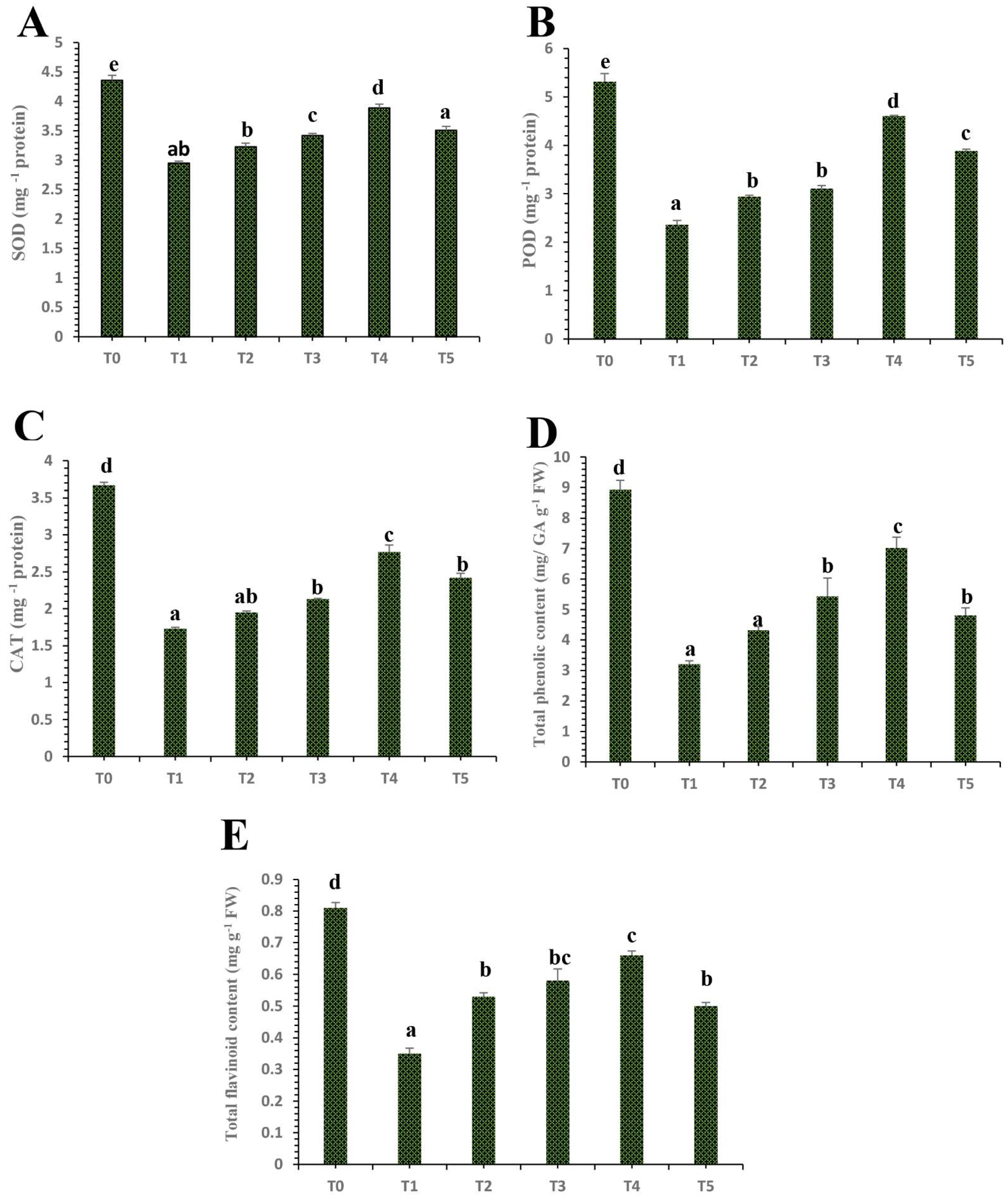
3.6. Differential Impacts of Plant Mediated Selenium Nanoparticles on Antioxidant Enzymatic and Non-Enzymatic Activity
4. Discussion
4.1. Identification of HLB Disease in ‘Kinnow’ Mandarin
4.2. Differential Effects of Plant Mediated SeNPs on Physiological Profiling of HLB-Infected ‘Kinnow’ Mandarin Plants
4.3. Differential Effects of Plant Mediated SeNPs on Antioxidant Defensive Enzymes Activities of HLB-Diseased ‘Kinnow’ Mandarin Plants
4.4. Differential Effects of Plant Mediated SeNPs on H2O2 Content and MDA Content of HLB-Diseased ‘Kinnow’ Mandarin Plants
4.5. Differential Effects of Plant-Mediated SeNPs on Proline and Total Sugars Content of HLB-Infected ‘Kinnow’ Mandarin Plants
4.6. Differential Effects of Plant Mediated SeNPs on Non-Enzymatic Antioxidant Activities of HLB-Infected ‘Kinnow’ Mandarin Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ramana, K.; Govindarajan, V.; Ranganna, S.; Kefford, J. Citrus fruits—Varieties, chemistry, technology, and quality evaluation. Part I: Varieties, production, handling, and storage. Crit. Rev. Food Sci. Nutr. 1981, 15, 353–431. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhao, C.; Shi, H.; Liao, Y.; Xu, F.; Du, H.; Xiao, H.; Zheng, J. Nutrients and bioactives in citrus fruits: Different citrus varieties, fruit parts, and growth stages. Crit. Rev. Food Sci. Nutr. 2021, 61, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.-J.; Chen, J.-B.; Cao, J.-P.; Li, X.; Sun, C.-D. Citrus flavonoids and their antioxidant evaluation. Crit. Rev. Food Sci. Nutr. 2020, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Heying, E.; Tanumihardjo, S.A. History, global distribution, and nutritional importance of citrus fruits. Compr. Rev. Food Sci. Food Saf. 2012, 11, 530–545. [Google Scholar] [CrossRef]
- Nawaz, R.; Abbasi, N.; Hafiz, I.; Khalid, A.; Ahmad, T.; Aftab, M. Impact of climate change on kinnow fruit industry of Pakistan. Agrotechnology 2019, 8, n186. [Google Scholar] [CrossRef] [Green Version]
- Jaouad, M.; Moinina, A.; Ezrzri, S.; Lahlali, R. Key pests and diseases of citrus trees with emphasis on root rot diseases: An overview. Mor. J. Agric. Sci. 2020, 1, 149–160. Available online: https://www.techagro.org/index.php/MJAS/article/view/851 (accessed on 14 December 2021).
- Tipu, M.M.H.; Rahman, M.M.; Islam, M.M.; Elahi, F.-E.; Jahan, R.; Islam, M.R. Citrus greening disease (HLB) on Citrus reticulata (Mandarin) caused by Candidatus Liberibacter asiaticus in Bangladesh. Physiol. Mol. Plant. Pathol. 2020, 112, 101558. [Google Scholar] [CrossRef]
- Pitino, M.; Sturgeon, K.; Dorado, C.; Cano, L.M.; Manthey, J.A.; Shatters Jr, R.G.; Rossi, L. Quercus leaf extracts display curative effects against Candidatus Liberibacter asiaticus that restore leaf physiological parameters in HLB-affected citrus trees. Plant. Physiol. Biochem. 2020, 148, 70–79. [Google Scholar] [CrossRef]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Ali, M.A.; Khalid, M.F.; Sohail, M.; Ercisli, S.; Zia-Ul-Haq, M.; Ahmad, S. Effect of different combinations of antibiotics on fruit quality and antioxidant defense system in Huanglongbing infected Kinnow orchards. AMB Express 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Etxeberria, E.; Narciso, C. Phloem anatomy of citrus trees: Healthy vs. greening-affected. In Proceedings of the Florida State Horticultural Society; Florida State Horticultural Society: Lake Alfred, FL, USA, 2012; pp. 67–70. [Google Scholar]
- Aritua, V.; Achor, D.; Gmitter, F.G.; Albrigo, G.; Wang, N. Transcriptional and microscopic analyses of citrus stem and root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2013, 8, e73742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razi, M.F.; Keremane, M.L.; Ramadugu, C.; Roose, M.; Khan, I.A.; Lee, R.F. Detection of citrus huanglongbing-associated ‘Candidatus Liberibacter asiaticus’ in citrus and Diaphorina citri in Pakistan, seasonal variability, and implications for disease management. Phytopathology 2014, 104, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajid, A.; Ghazanfar, M.U.; Rauf, S.; Hussain, Z.; Ahmad, S.; Iftikhar, Y. Incidence and molecular detection of greening disease in two citrus cultivars in Sargodha, Pakistan. Sarhad J. Agric. 2021, 37, 296–301. [Google Scholar] [CrossRef]
- Chen, X.D.; Gill, T.A.; Pelz-Stelinski, K.S.; Stelinski, L.L. Risk assessment of various insecticides used for management of Asian citrus psyllid, Diaphorina citri in Florida citrus, against honey bee, Apis Mellifera. Ecotoxicol. 2017, 26, 351–359. [Google Scholar] [CrossRef]
- Yang, C.; Zhong, Y.; Powell, C.A.; Doud, M.S.; Duan, Y.; Huang, Y.; Zhang, M. Antimicrobial compounds effective against Candidatus Liberibacter asiaticus discovered via graft-based assay in citrus. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Blaustein, R.A.; Lorca, G.L.; Teplitski, M. Challenges for managing Candidatus Liberibacter spp.(Huanglongbing disease pathogen): Current control measures and future directions. Phytopathology 2018, 108, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Boina, D.R.; Bloomquist, J.R. Chemical control of the Asian citrus psyllid and of huanglongbing disease in citrus. Pest. Manage. Sci. 2015, 71, 808–823. [Google Scholar] [CrossRef]
- Hussain, M.; Raja, N.I.; Naz, F.; Iqbal, M.; Aslam, S. Green synthesis and characterisation of silver nanoparticles and their effects on antimicrobial efficacy and biochemical profiling in Citrus reticulata. IET Nanobiotechnol. 2018, 12, 514–519. [Google Scholar] [CrossRef]
- Ragavan, P.; Ananth, A.; Rajan, M. Impact of selenium nanoparticles on growth, biochemical characteristics and yield of cluster bean Cyamopsis tetragonoloba. Int. J. Environ. Agric. Biotech. 2017, 2, 238983. [Google Scholar] [CrossRef]
- Zohra, E.; Ikram, M.; Omar, A.A.; Hussain, M.; Satti, S.H.; Raja, N.I.; Ehsan, M. Potential applications of biogenic selenium nanoparticles in alleviating biotic and abiotic stresses in plants: A comprehensive insight on the mechanistic approach and future perspectives. Green Process. Synth. 2021, 10, 456–475. [Google Scholar] [CrossRef]
- Ikram, M.; Javed, B.; Raja, N.I.; Mashwani, Z.-u.-R. Biomedical potential of plant-based selenium nanoparticles: A comprehensive review on therapeutic and mechanistic aspects. Int. J. Nanomed. 2021, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Alagesan, V.; Venugopal, S. Green synthesis of selenium nanoparticle using leaves extract of withania somnifera and its biological applications and photocatalytic activities. BioNano Sci. 2019, 9, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Alam, H.; Khatoon, N.; Raza, M.; Ghosh, P.C.; Sardar, M. Synthesis and characterization of nano selenium using plant biomolecules and their potential applications. BioNanoScience 2019, 9, 96–104. [Google Scholar] [CrossRef]
- Agarwal, R.; Behari, J. Effect of selenium pretreatment in chronic mercury intoxication in rats. Bull. Environ. Contam. Toxicol. 2007, 79, 306–310. [Google Scholar] [CrossRef]
- Zonaro, E.; Lampis, S.; Turner, R.J.; Qazi, S.J.S.; Vallini, G. Biogenic selenium and tellurium nanoparticles synthesized by environmental microbial isolates efficaciously inhibit bacterial planktonic cultures and biofilms. Front. Microbiol. 2015, 6, 584. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Huo, Y.; Yin, S.; Hu, H. Mechanisms of acetaminophen-induced liver injury and its implications for therapeutic interventions. Redox Biol. 2018, 17, 274–283. [Google Scholar] [CrossRef]
- Ismail, A.-W.A.; Sidkey, N.M.; Arafa, R.A.; Fathy, R.M.; El-Batal, A.I. Evaluation of in vitro antifungal activity of silver and selenium nanoparticles against Alternaria solani caused early blight disease on potato. Biotechnol. J. Int. 2016, 1–11. [Google Scholar] [CrossRef]
- Nandini, B.; Hariprasad, P.; Prakash, H.S.; Shetty, H.S.; Geetha, N. Trichogenic-selenium nanoparticles enhance disease suppressive ability of Trichoderma against downy mildew disease caused by Sclerospora graminicola in pearl millet. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Shafi, M.E.; Selem, E.; Desoky, E.-S.M.; Fouda, S.E.; El-Tahan, A.M. The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; El-Hack, A.; Mohamed, E.; Taha, A.E.; Fouda, M.M.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; Elshaer, N. Ecofriendly synthesis and insecticidal application of copper nanoparticles against the storage pest Tribolium castaneum. Nanomaterials 2020, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, M.M.; Dodamani, A.S.; Karibasappa, G.N.; Vishwakarma, P.K.; Vathar, J.B.; Sonawane, K.R.; Jadhav, H.C.; Khobragade, V.R. Antibacterial activity of garlic extract on cariogenic bacteria: An in vitro study. Ayu 2018, 39, 165. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant. Mol. Biol. Rep. 1983, 1, 19–21. Available online: https://0-link-springer-com.brum.beds.ac.uk/content/pdf/10.1007/BF02712670.pdf (accessed on 14 December 2021). [CrossRef]
- Hung, T.; Wu, n.M.; Su, H. Development of a rapid method for the diagnosis of citrus greening disease using the polymerase chain reaction. J. Phytopathol. 1999, 147, 599–604. [Google Scholar] [CrossRef]
- Hong, Y.; Luo, Y.; Yi, J.; He, L.; Dai, L.; Yi, T. Screening nested-PCR primer for ‘Candidatus Liberibacter asiaticus’ associated with citrus Huanglongbing and application in Hunan, China. PLoS ONE 2019, 14, e0212020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, K.S.; Selvaraj, K. Biogenic synthesis of selenium nanoparticles and their effect on As (III)-induced toxicity on human lymphocytes. Biol. Trace Elem. Res. 2014, 157, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Anu, K.; Singaravelu, G.; Murugan, K.; Benelli, G. Green-Synthesis of Selenium Nanoparticles Using Garlic Cloves (Allium sativum): Biophysical Characterization and Cytotoxicity on Vero Cells. J. Clust. Sci. 2017, 28, 551–563. [Google Scholar] [CrossRef]
- Ikram, M.; Raja, N.I.; Javed, B.; Hussain, M.; Hussain, M.; Ehsan, M.; Rafique, N.; Malik, K.; Sultana, T.; Akram, A. Foliar applications of bio-fabricated selenium nanoparticles to improve the growth of wheat plants under drought stress. Green Process. Synth. 2020, 9, 706–714. [Google Scholar] [CrossRef]
- Javed, B. Synergistic effects of physicochemical parameters on bio-fabrication of mint silver nanoparticles: Structural evaluation and action against HCT116 colon cancer cells. Int. J. Nanomed. 2020, 15, 3621. [Google Scholar] [CrossRef]
- Javed, B.; Nadhman, A.; Mashwani, Z.-u.-R. Phytosynthesis of Ag nanoparticles from Mentha longifolia: Their structural evaluation and therapeutic potential against HCT116 colon cancer, Leishmanial and bacterial cells. Appl. Nanosci. 2020. [Google Scholar] [CrossRef]
- Kamble, P.N.; Giri, S.P.; Mane, R.S.; Tiwana, A. Estimation of chlorophyll content in young and adult leaves of some selected plants. Univers. J. Environ. Res. Tech. 2015, 5, 306–310. Available online: http://www.environmentaljournal.org/5-6/ujert-5-6-5.pdf (accessed on 14 December 2021).
- Kumari, R.; Ashraf, S.; Bagri, G.; Khatik, S.; Bagri, D.; Bagdi, D. Extraction and estimation of chlorophyll content of seed treated lentil crop using DMSO and acetone. J. Pharm. Phytochem. 2018, 7, 249–250. Available online: https://www.phytojournal.com/archives/2018/vol7issue3/PartD/7-2-171-916.pdf (accessed on 14 December 2021).
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant. Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oktay, M.; Küfreviolu, I.; Kocaçalişkan, I.; Şaklrolu, H. Polyphenoloxidase from Amasya apple. J. Food Sci. 1995, 60, 494–496. [Google Scholar] [CrossRef]
- Behboudi, F.; Tahmasebi-Sarvestani, Z.; Kassaee, M.Z.; Modarres-Sanavy, S.A.M.; Sorooshzadeh, A.; Mokhtassi-Bidgoli, A. Evaluation of chitosan nanoparticles effects with two application methods on wheat under drought stress. J. Plant. Nutr. 2019, 42, 1439–1451. [Google Scholar] [CrossRef]
- Sairam, R. Effect of moisture-stress on physiological activities of two contrasting wheat genotypes. Indian J. Exp. Biol. 1994, 32, 594. [Google Scholar]
- Denaxa, N.-K.; Damvakaris, T.; Roussos, P.A. Antioxidant defense system in young olive plants against drought stress and mitigation of adverse effects through external application of alleviating products. Sci. Hortic. 2020, 259, 108812. [Google Scholar] [CrossRef]
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.N.; Akhtar, S.S.; Großkinsky, D.K.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant. Methods 2020, 16, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Hijaz, F.; Al-Rimawi, F.; Manthey, J.A.; Killiny, N. Phenolics, flavonoids and antioxidant capacities in Citrus species with different degree of tolerance to Huanglongbing. Plant. Signal. Behav. 2020, 15, 1752447. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. Available online: https://www.ajevonline.org/content/16/3/144 (accessed on 14 December 2021).
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Antioxidant potentials and anticholinesterase activities of methanolic and aqueous extracts of three endemic Centaurea L. species. Food Chem. Toxicol. 2013, 55, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Bhagyawant, S.S.; Narvekar, D.T.; Gupta, N.; Bhadkaria, A.; Koul, K.K.; Srivastava, N. Variations in the antioxidant and free radical scavenging under induced heavy metal stress expressed as proline content in chickpea. Physiol. Mol. Biol. Plants 2019, 25, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Gurrieri, L.; Merico, M.; Trost, P.; Forlani, G.; Sparla, F. Impact of drought on soluble sugars and free proline content in selected Arabidopsis mutants. Biology 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Ghosh, I.; Ghosh, M.; Mukherjee, A. Methods of in vitro and in vivo nanotoxicity evaluation in plants. In Environmental Toxicity of Nanomaterials; Kumar, V., Dasgupta, N., Ranjan, S., Eds.; CRC Press: New York, NY, USA, 2018; pp. 281–304. [Google Scholar]
- Menon, S.; Agarwal, H.; Shanmugam, V.K. Catalytical degradation of industrial dyes using biosynthesized selenium nanoparticles and evaluating its antimicrobial activities. Sustain. Environ. Res. 2021, 31. [Google Scholar] [CrossRef]
- Ruangwong, O.; Akarapisan, A. Detection of Candidatus Liberibacter asiaticus causing citrus huanglongbing disease. J. Agric. Sci. Technol. 2006, 2, 111–120. [Google Scholar]
- Khan, M.; Khan, A.U.; Hasan, M.A.; Yadav, K.K.; Pinto, M.; Malik, N.; Yadav, V.K.; Khan, A.H.; Islam, S.; Sharma, G.K. Agro-nanotechnology as an emerging field: A novel sustainable approach for improving plant growth by reducing biotic stress. Appl. Sci. 2021, 11, 2282. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant. Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Quiterio-Gutiérrez, T.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Hernández-Fuentes, A.D.; Sandoval-Rangel, A.; Benavides-Mendoza, A.; Cabrera-de la Fuente, M.; Juárez-Maldonado, A. The application of selenium and copper nanoparticles modifies the biochemical responses of tomato plants under stress by Alternaria solani. Int. J. Mol. Sci. 2019, 20, 1950. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.-S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Z.; Wang, Y.; Wang, S.H.; Yin, L.P.; Xu, G.J.; Zheng, C.; Lei, C.; Zhang, M.Z. Selenium increases chlorogenic acid, chlorophyll and carotenoids of Lycium chinense leaves. J. Sci. Food Agric. 2013, 93, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant. 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Kreft, I.; Osvald, J. Influence of UV-B exclusion and selenium treatment on photochemical efficiency of photosystem II, yield and respiratory potential in pumpkins (Cucurbita pepo L.). Plant. Physiol. Biochem. 2005, 43, 445–448. [Google Scholar] [CrossRef]
- Lapaz, A.d.M.; Santos, L.F.d.M.; Yoshida, C.H.P.; Heinrichs, R.; Campos, M.; Reis, A.R.d. Physiological and toxic effects of selenium on seed germination of cowpea seedlings. Bragantia 2019, 78, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Satti, S.H.; Raja, N.I.; Javed, B.; Akram, A.; Mashwani, Z.-u.-R.; Ahmad, M.S.; Ikram, M. Titanium dioxide nanoparticles elicited agro-morphological and physicochemical modifications in wheat plants to control Bipolaris sorokiniana. PLoS ONE 2021, 16, e0246880. [Google Scholar] [CrossRef]
- Tang, H.; Liu, Y.; Gong, X.; Zeng, G.; Zheng, B.; Wang, D.; Sun, Z.; Zhou, L.; Zeng, X. Effects of selenium and silicon on enhancing antioxidative capacity in ramie (Boehmeria nivea (L.) Gaud.) under cadmium stress. Environ. Sci. Pollut. Res. 2015, 22, 9999–10008. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Ma, W.; Pang, Z.; Huang, X.; Pandey, S.S.; Li, J.; Xu, J.; Achor, D.S.; Vasconcelos, F.N.; Hendrich, C.; Huang, Y. Citrus Huanglongbing is an immune-mediated disease that can be treated by mitigating reactive oxygen species triggered cell death of the phloem tissues caused by Candidatus Liberibacter asiaticus. BioRxiv 2021. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, G.; Abdelrassoul, M.; Hegazi, A.; Elsherif, M. Mitigation of saline stress adverse effects in lettuce plant using selenium and silicon. Middle East J. Agric. Res. 2016, 5, 347–361. [Google Scholar]
- Tarrahi, R.; Khataee, A.; Movafeghi, A.; Rezanejad, F.; Gohari, G. Toxicological implications of selenium nanoparticles with different coatings along with Se4+ on Lemna minor. Chemosphere 2017, 181, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, M.; Jogaiah, S.; Burritt, D.J.; Tran, L.S.P. Legume genetic resources and transcriptome dynamics under abiotic stress conditions. Plant Cell Environ. 2018, 41, 1972–1983. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, M.; Gurunathan, S.; Chung, I.-M. Physiological, metabolic, and transcriptional effects of biologically-synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma 2015, 252, 1031–1046. [Google Scholar] [CrossRef]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.-S.P. Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: Oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef]
- Mangiapane, E.; Pessione, A.; Pessione, E. Selenium and selenoproteins: An overview on differrent biological systems. Curr. Protein Pept. Sci. 2018, 19, 725. [Google Scholar] [CrossRef]
- Monreal, J.; Jimenez, E.; Remesal, E.; Morillo-Velarde, R.; García-Mauriño, S.; Echevarría, C. Proline content of sugar beet storage roots: Response to water deficit and nitrogen fertilization at field conditions. Environ. Exp. Bot. 2007, 60, 257–267. [Google Scholar] [CrossRef]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant. 2014, 36, 1607–1619. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Raja, N.I.; Wattoo, F.H.; Hussain, M.; Ejaz, M.; Saira, H. Assessment of AgNPs exposure on physiological and biochemical changes and antioxidative defence system in wheat (Triticum aestivum L) under heat stress. IET Nanobiotechnol. 2019, 13, 230–236. [Google Scholar] [CrossRef]
- Sultana, T.; Javed, B.; Raja, N.I. Silver nanoparticles elicited physiological, biochemical, and antioxidant modifications in rice plants to control Aspergillus flavus. Green Process. Synth. 2021, 10, 314–324. [Google Scholar] [CrossRef]
- Chandra, S.; Chakraborty, N.; Dasgupta, A.; Sarkar, J.; Panda, K.; Acharya, K. Chitosan nanoparticles: A positive modulator of innate immune responses in plants. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles alter secondary metabolism in plants via ROS burst. Front. Plant. Sci. 2017, 8, 832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982. [Google Scholar] [CrossRef] [Green Version]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of zinc nanoparticles on antioxidative system of potato plants. J. Environ. Biol. 2017, 38, 435. [Google Scholar] [CrossRef]
- López-Vargas, E.R.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; de Alba Romenus, K.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef] [Green Version]
- Vennila, K.; Chitra, L.; Balagurunathan, R.; Palvannan, T. Comparison of biological activities of selenium and silver nanoparticles attached with bioactive phytoconstituents: Green synthesized using Spermacoce hispida extract. Adv. Nat. Sci. Nanosci. 2018, 9, 015005. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
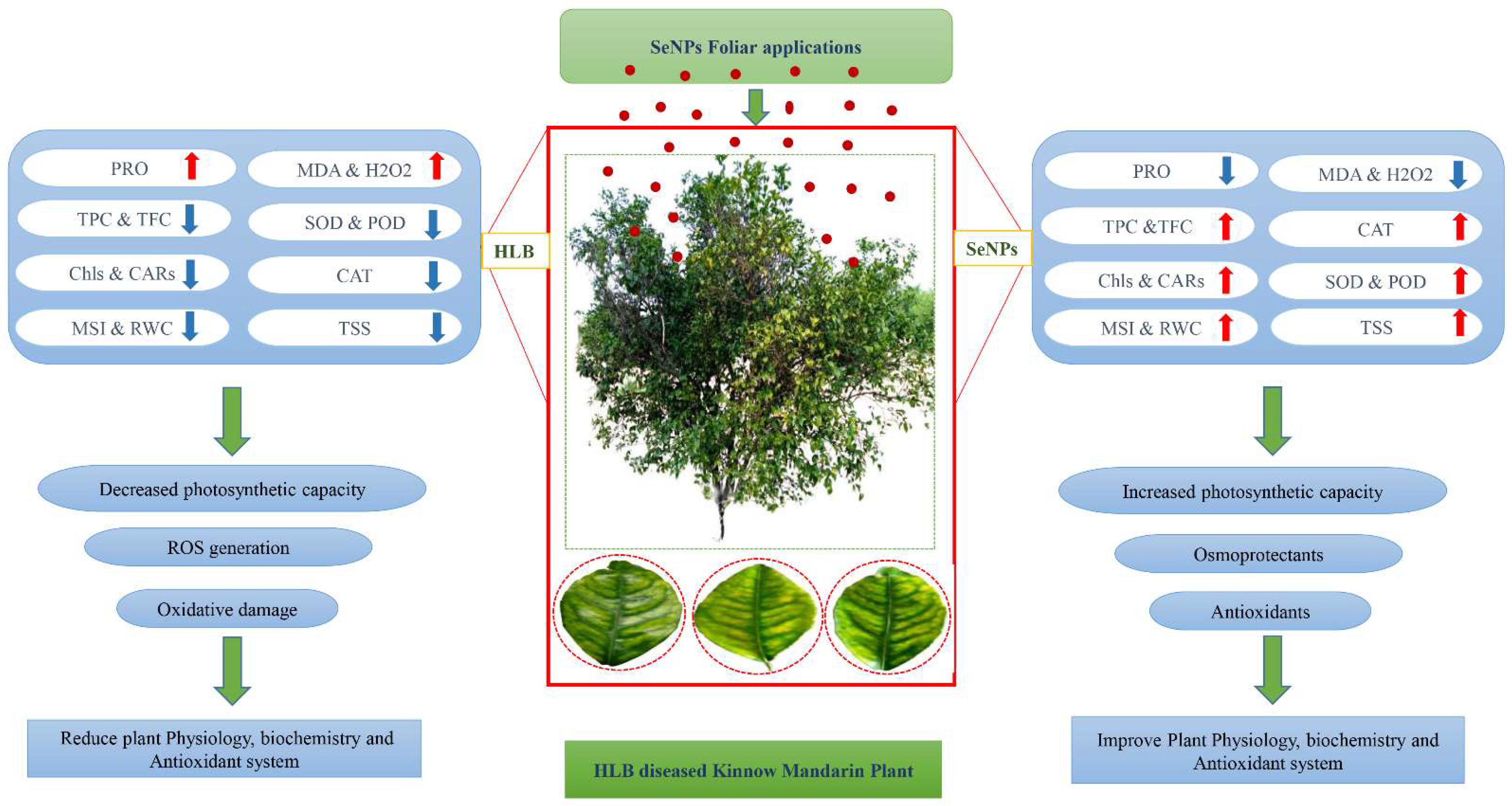{kind=link}
| Treatment | Conditions |
|---|---|
| T0 | Control (healthy ‘Kinnow’ mandarin plants) |
| T1 | HLB-infected ‘Kinnow’ mandarin plants (distillted water) |
| T2 | HLB-infected ‘Kinnow’ mandarin plants + 25 mg L−1 SeNPs |
| T3 | HLB-infected ‘Kinnow’ mandarin plants + 50 mg L−1 SeNPs |
| T4 | HLB-infected ‘Kinnow’ mandarin plants + 75 mg L−1 SeNPs |
| T5 | HLB-infected ‘Kinnow’ mandarin plants + 100 mg L−1 SeNPs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikram, M.; Raja, N.I.; Mashwani, Z.-U.-R.; Omar, A.A.; Mohamed, A.H.; Satti, S.H.; Zohra, E. Phytogenic Selenium Nanoparticles Elicited the Physiological, Biochemical, and Antioxidant Defense System Amelioration of Huanglongbing-Infected ‘Kinnow’ Mandarin Plants. Nanomaterials 2022, 12, 356. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12030356
Ikram M, Raja NI, Mashwani Z-U-R, Omar AA, Mohamed AH, Satti SH, Zohra E. Phytogenic Selenium Nanoparticles Elicited the Physiological, Biochemical, and Antioxidant Defense System Amelioration of Huanglongbing-Infected ‘Kinnow’ Mandarin Plants. Nanomaterials. 2022; 12(3):356. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12030356
Chicago/Turabian StyleIkram, Muhammad, Naveed Iqbal Raja, Zia-Ur-Rehman Mashwani, Ahmad Alsayed Omar, Azza H. Mohamed, Seema Hassan Satti, and Efat Zohra. 2022. "Phytogenic Selenium Nanoparticles Elicited the Physiological, Biochemical, and Antioxidant Defense System Amelioration of Huanglongbing-Infected ‘Kinnow’ Mandarin Plants" Nanomaterials 12, no. 3: 356. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12030356