
The Effects of the Food Additive Titanium Dioxide (E171) on Tumor Formation and Gene Expression in the Colon of a Transgenic Mouse Model for Colorectal Cancer
 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. E171 Characterization
2.2. Animals
2.3. Pilot Study: Histopathology
2.4. Phase I: Tumor Formation Study
2.5. Phase II: Gene Expression Study
2.6. mRNA Extraction from Colonic Tissues
2.7. cRNA Synthesis, Labeling, and Hybridization
2.8. Preprocessing and Data Analysis of Microarrays
2.9. Pathway Analysis
2.10. STEM Analysis
2.11. Network Analysis
3. Results
3.1. Particle Characterization
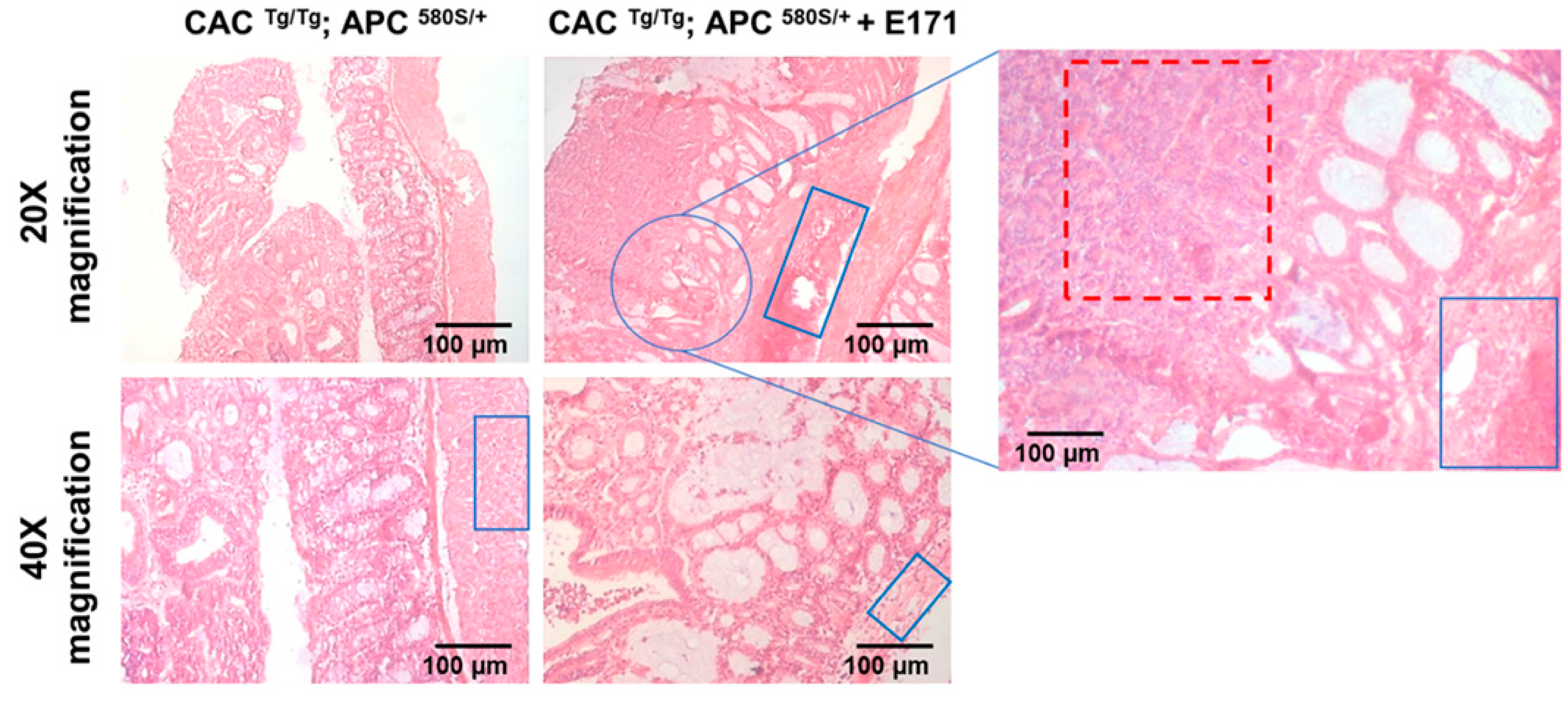
3.2. Pilot Study: Histopathology
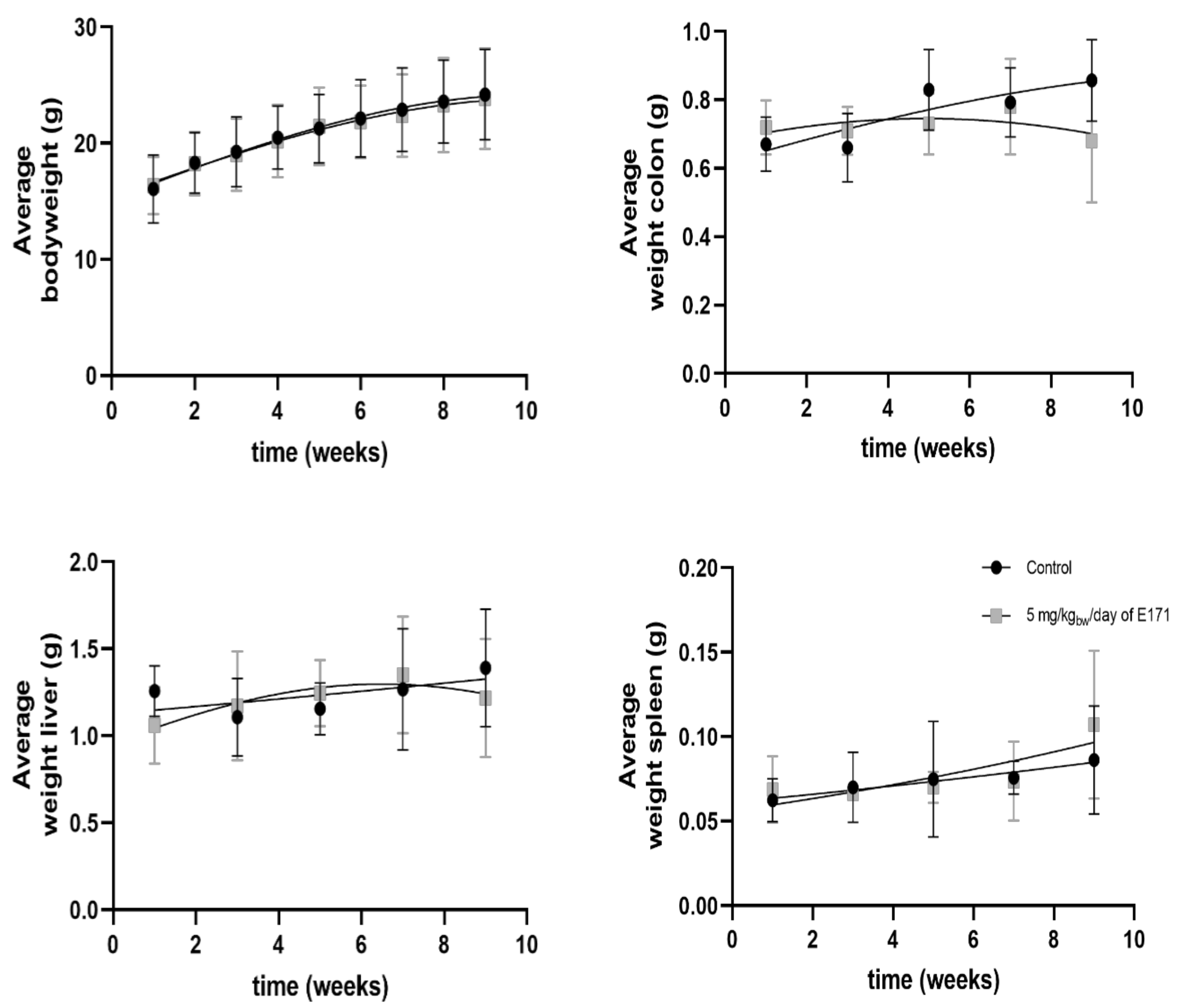
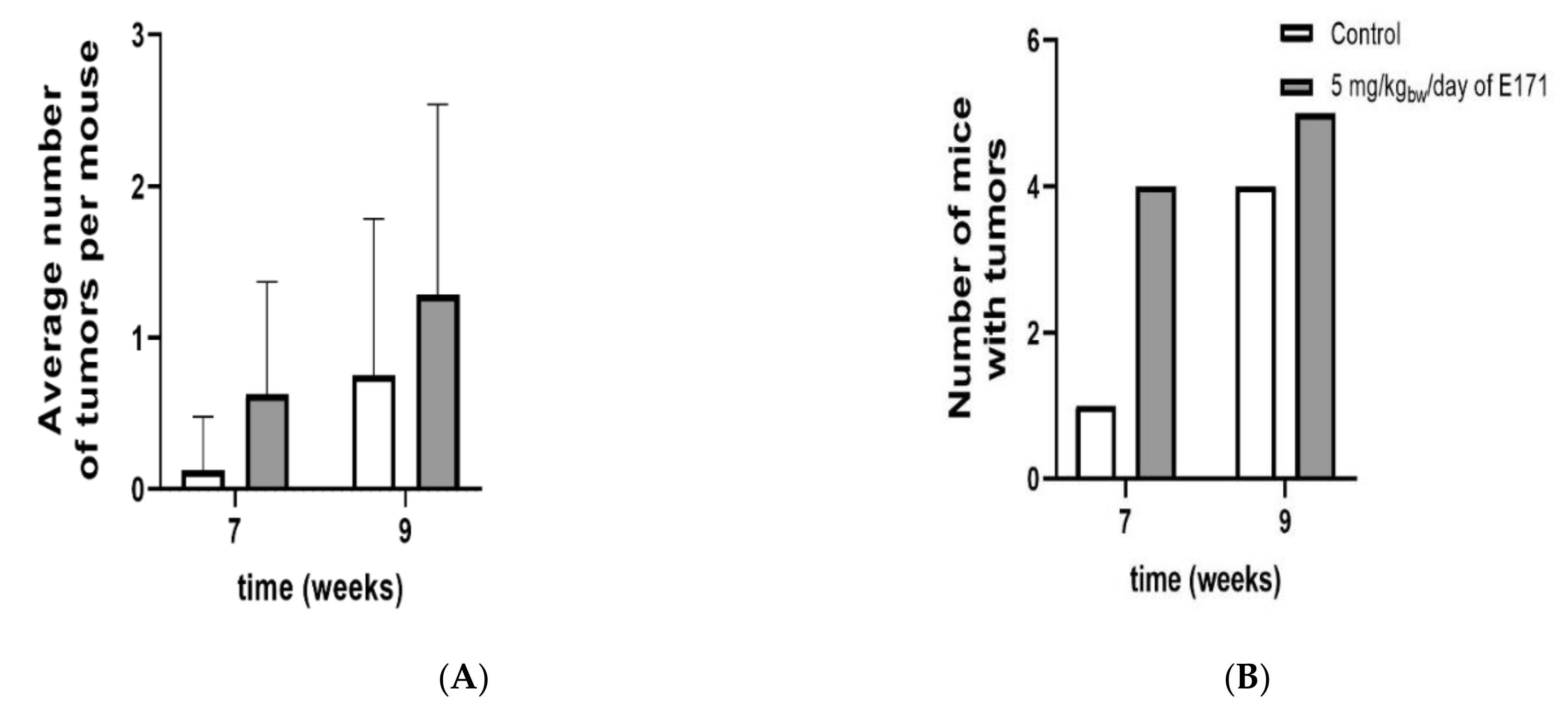
3.3. Phase I: Tumor Formation Study
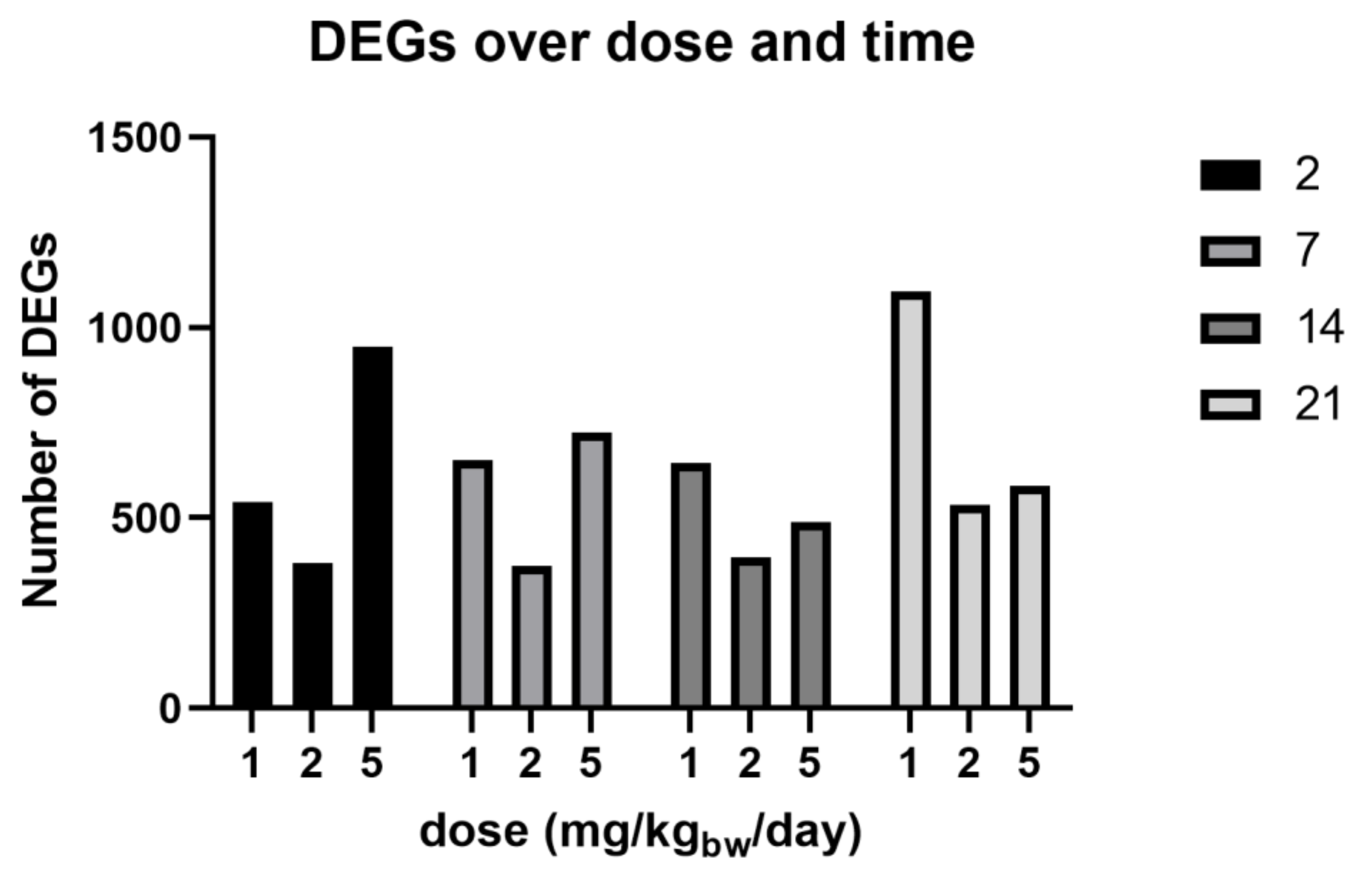
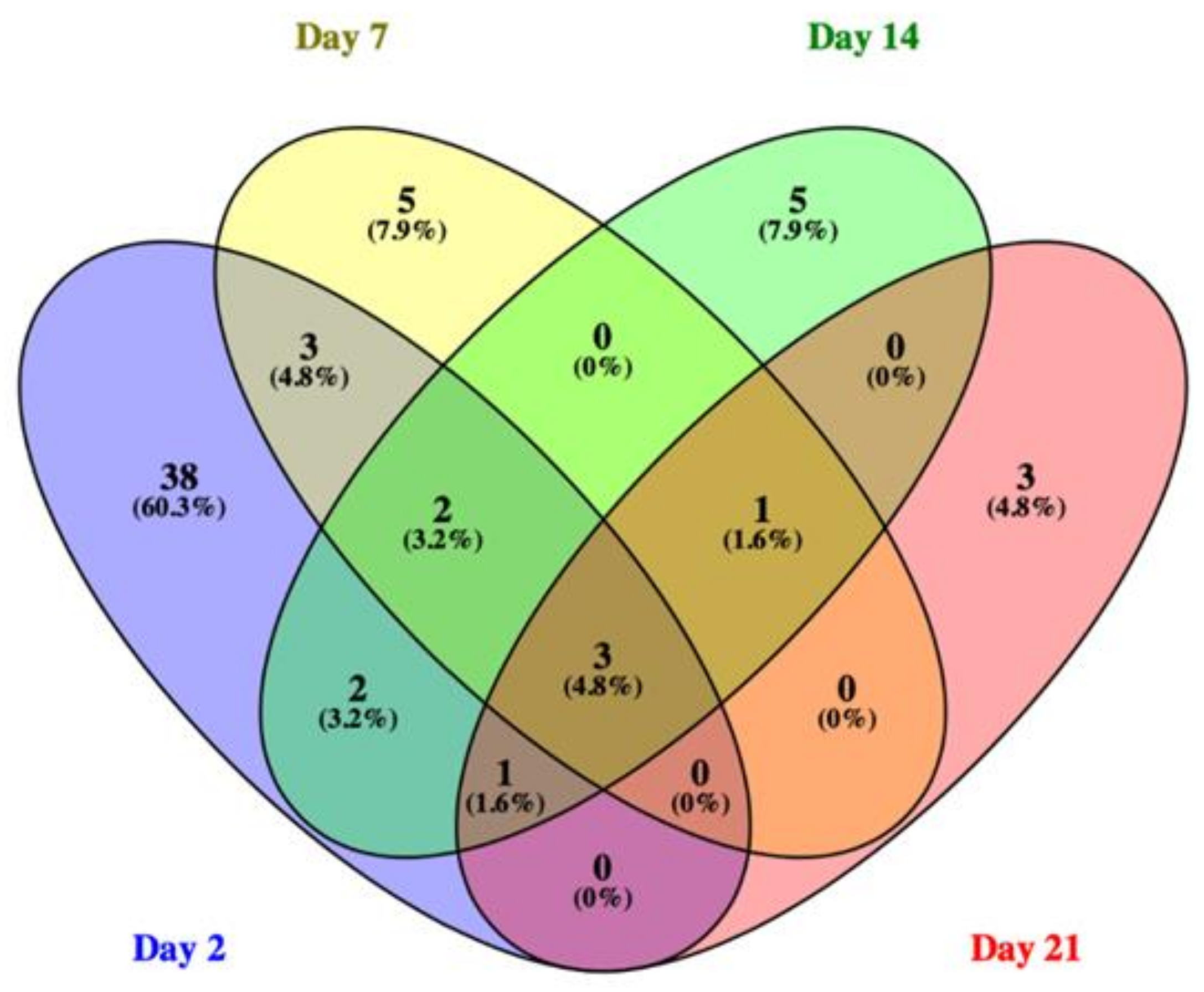
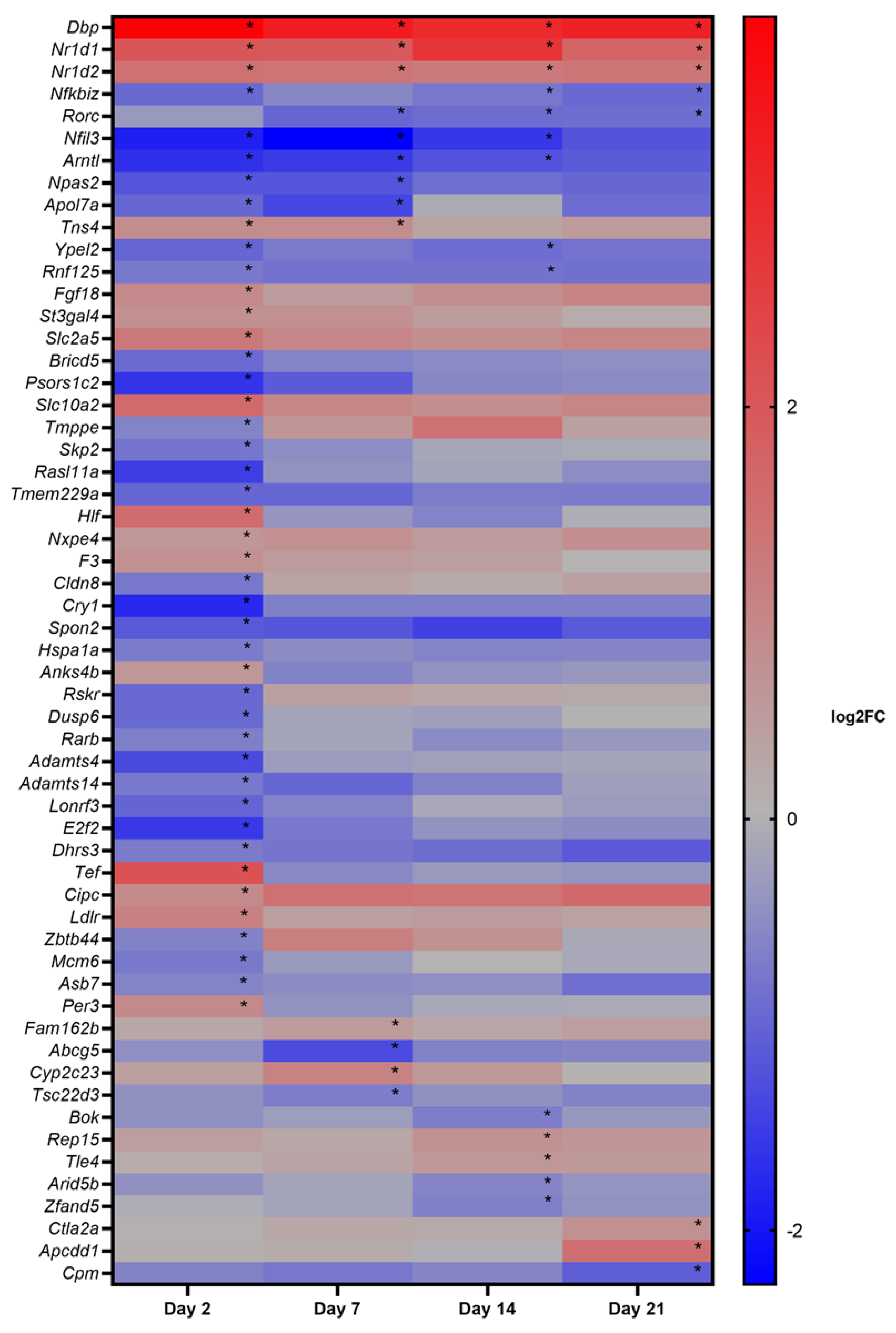
3.4. Phase II: Gene Expression Study
3.5. Pathway Analysis
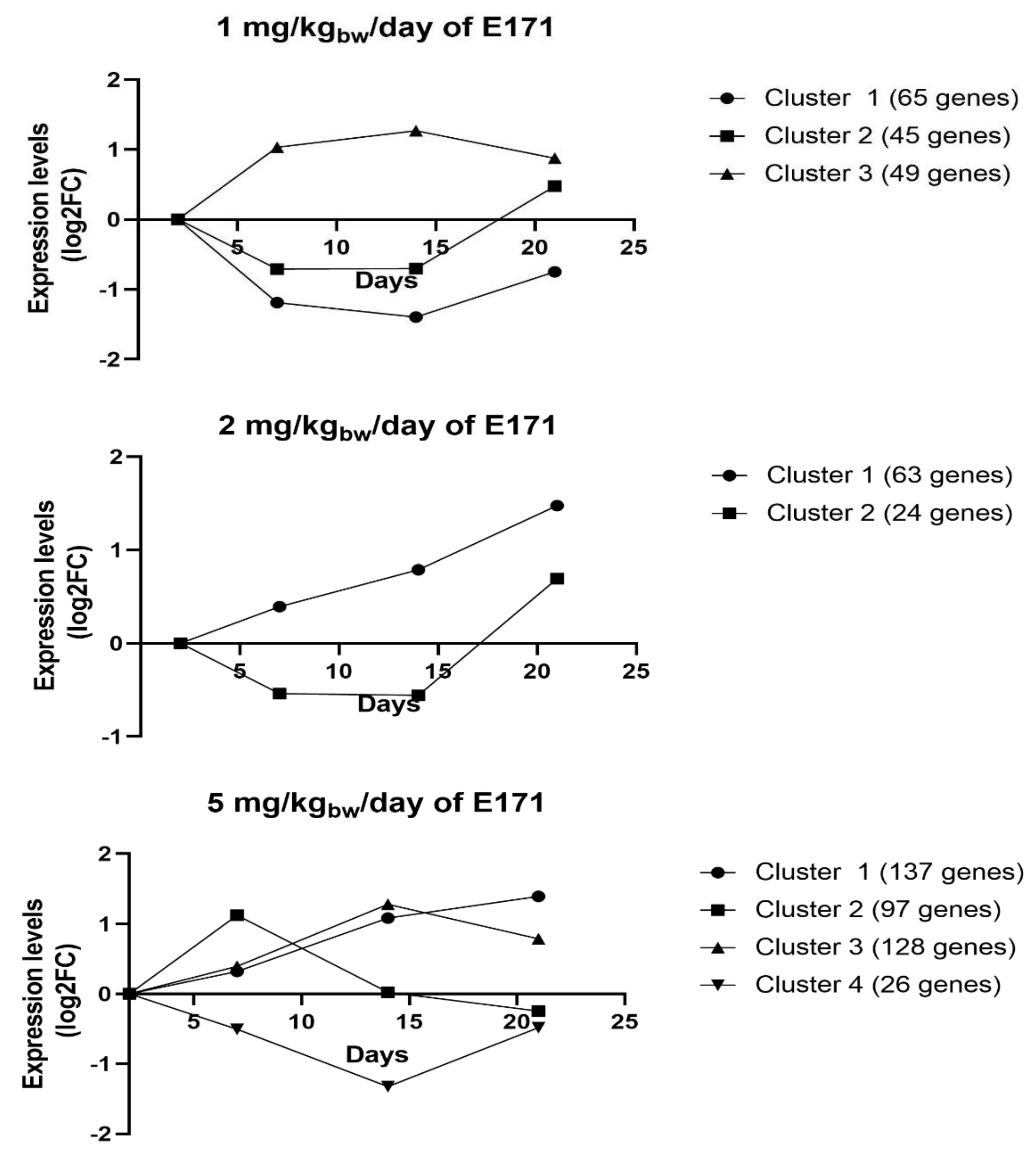
3.6. STEM Analysis
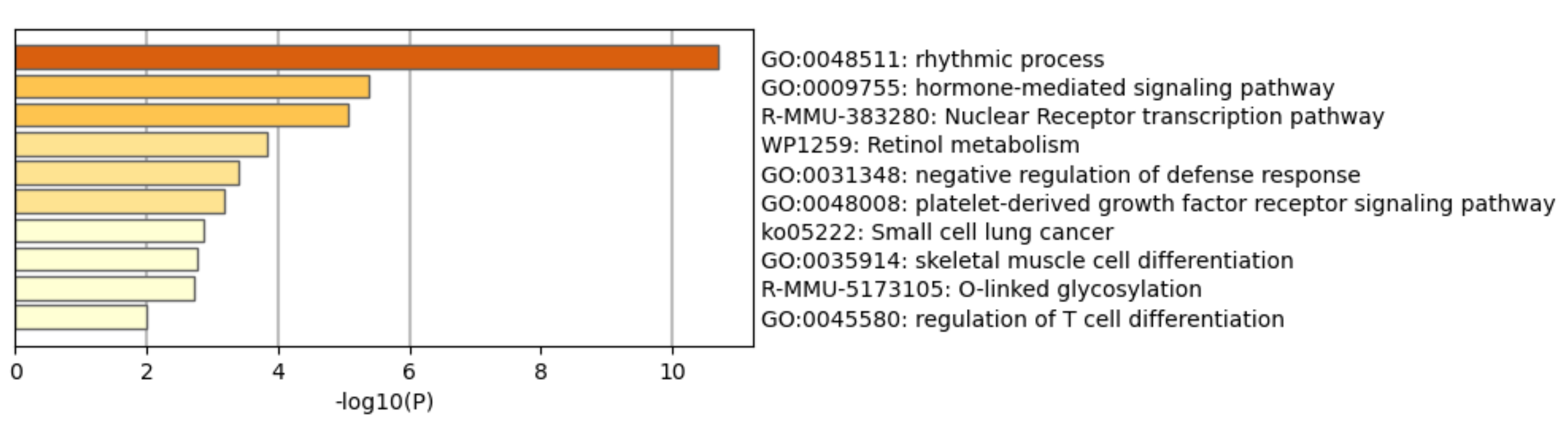
3.7. Functional Enrichment Analysis and Network Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AOM | azoxymethane |
| Apc | adenomatous polyposis coli |
| Apol7a | apolipoprotein L7a |
| Arntl | aryl hydrocarbon receptor nuclear translocator-like |
| Bmal1 | brain and muscle ARNT-like 1 |
| CAC | colitis-associated colorectal cancer |
| CI | confidence interval |
| CLP | classification, labelling, and packaging |
| COX2 | prostaglandin-endoperoxide synthase 2 |
| CPDB | ConsesusPathway database |
| CRC | colorectal cancer |
| cRNA | complementary RNA |
| Dbp | D site albumin promoter binding protein |
| DEG | differentially expressed gene |
| DNA | deoxyribonucleic acid |
| DSS | dextran sodium sulfate |
| EC | European Commission |
| EFSA | European Food Safety Authority |
| ERBB2 | Erb-B2 receptor tyrosine kinase 2 |
| ERBB4 | Erb-B2 receptor tyrosine kinase 4 |
| EU | European Union |
| FC | fold change |
| FDR | false discovery rate |
| FES | feature extraction software |
| FGFR1 | fibroblast growth factor receptor 1 |
| FIB | focused ion beam |
| GPCR | G-protein-coupled receptor |
| HE | hematoxylin and eosin |
| IARC | International Agency for Research and Cancer |
| IL | interleukin |
| MAPK | mitogen-activated protein kinase |
| MMP | matrixmetalloproteinase |
| mRNA | messenger ribonucleic acid |
| Nfil3 | nuclear factor interleukin 3, regulated |
| Nfkbiz | nuclear factor of kappa light polypeptide gene enhancer in B-cell inhibitor zeta |
| NLRP3 | NOD-, LRR- and pyrin-domain-containing protein 3 |
| Npas2 | neuronal PAS domain 2 |
| Nr1d1 | nuclear receptor subfamily 1, group D, member 1 |
| Nr1d2 | nuclear receptor subfamily 1, group D, member 2 |
| ORA | over-representation analysis |
| PCR | polymerase chain reaction |
| PI3K | phosphoinositide 3-kinase |
| PLK1 | polo-like kinase 1 |
| PMT | photomultiplier tubes |
| RAF | rapidly accelerated fibrosarcoma |
| RARE | retinoic acid response element |
| RIN | RNA integrity number |
| Rnf125 | ring finger protein 125 |
| Rorc | RAR-related orphan receptor gamma |
| SD | standard deviation |
| SEM | scanning electron microscopy |
| STEM | short time-series expression miner |
| TEM | transmission electron microscopy |
| Tg | transgenic |
| TiO2 | titanium dioxide |
| Tns4 | tensin 4 |
| TP53 | tumor protein P53 |
| TRP | transient receptor potential |
| UK | United Kingdom |
| UV | ultraviolet |
| WNT | wingless-related integration site |
| Ypel2 | protein yippee-like 2 |
References
- Siegel, R.L.; Jakubowski, C.D.; Fedewa, S.A.; Davis, A.; Azad, N.S. Colorectal Cancer in the Young: Epidemiology, Prevention, Management. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, e75–e88. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Sauer, A.G.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Veettil, S.K.; Wong, T.Y.; Loo, Y.S.; Playdon, M.C.; Lai, N.M.; Giovannucci, E.L.; Chaiyakunapruk, N. Role of Diet in Colorectal Cancer Incidence: Umbrella Review of Meta-analyses of Prospective Observational Studies. JAMA Netw. Open. 2021, 4, e2037341. [Google Scholar] [CrossRef]
- McClements, D.J.; Xiao, H. Is nano safe in foods? Establishing the factors impacting the gastrointestinal fate and toxicity of organic and inorganic food-grade nanoparticles. NPJ Sci. Food 2017, 1, 6. [Google Scholar] [CrossRef]
- EFSA. Safety assessment of titanium dioxide (E171) as a food additive. EFSA J. 2021, 19, 6585. [Google Scholar]
- Verleysen, E.; Waegeneers, N.; Brassinne, F.; De Vos, S.; Jimenez, I.O.; Mathioudaki, S.; Mast, J. Physicochemical Characterization of the Pristine E171 Food Additive by Standardized and Validated Methods. Nanomaterials 2020, 10, 592. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, N.S.; De Kok, T.M.; Sijm, D.T.; Van Breda, S.G.; Briedé, J.J.; Castenmiller, J.J.; Opperhuizen, A.; Chirino, Y.I.; Dirven, H.; Gott, D.; et al. Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System. Int. J. Mol. Sci. 2020, 22, 207. [Google Scholar] [CrossRef]
- Peters, R.J.B.; van Bemmel, G.; Herrera-Rivera, Z.; Helsper, H.P.F.G.; Marvin, H.J.P.; Weigel, S.; Tromp, P.C.; Oomen, A.G.; Rietveld, A.G.; Bouwmeester, H. Characterization of Titanium Dioxide Nanoparticles in Food Products: Analytical Methods To Define Nanoparticles. J. Agric. Food Chem. 2014, 62, 6285–6293. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15–33. [Google Scholar] [CrossRef]
- IARC. IARC Moographs on the Evaluation of Carcinogenic Risks to Humans: Carbon Black, Titanium Dioxide and Talc. 2010, Volume 93. Available online: https://publications.iarc.fr/Book-And-Report-Series/Iarc-Monographs-On-The-Identification-Of-Carcinogenic-Hazards-To-Humans/Carbon-Black-Titanium-Dioxide-And-Talc-2010 (accessed on 17 December 2021).
- ECHA. Opinion Proposing Harmonised Classification and Labelling a EU Level of Titanium Dioxide. 2017. Available online: https://echa.europa.eu/documents/10162/682fac9f-5b01-86d3-2f70-3d40277a53c2 (accessed on 17 December 2021).
- Urrutia-Ortega, I.M.; Garduño-Balderas, L.G.; Delgado-Buenrostro, N.L.; Freyre-Fonseca, V.; Flores-Flores, J.O.; González-Robles, A.; Pedraza-Chaverri, J.; Hernández-Pando, R.; Rodríguez-Sosa, M.; León-Cabrera, S.; et al. Food-grade titanium dioxide exposure exacerbates tumor formation in colitis associated cancer model. Food Chem. Toxicol. 2016, 93, 20–31. [Google Scholar] [CrossRef]
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Coméra, C.; Gaultier, E.; Dupuy, J.; Naud, N.; Taché, S.; Grysan, P.; Reguer, S.; et al. Food-grade TiO2 impairs intestinal and systemic immune homeostasis, initiates preneoplastic lesions and promotes aberrant crypt development in the rat colon. Sci. Rep. 2017, 7, srep40373. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Jetten, M.J.; Jonkhout, M.; Garduño-Balderas, L.G.; Briedé, J.; de Kok, T.M.; Chirino, Y.I.; van Loveren, H. Gene expression profiling in colon of mice exposed to food additive titanium dioxide (E171). Food Chem. Toxicol. 2018, 111, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Jetten, M.J.; Jonkhout, M.C.M.; Garduño-Balderas, L.G.; Briedé, J.J.; De Kok, T.M.; Van Loveren, H.; Chirino, Y.I. Transcriptomics analysis reveals new insights in E171-induced molecular alterations in a mouse model of colon cancer. Sci. Rep. 2018, 8, 9738. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Johnson, R.; Desmet, M.; Snyder, P.W.; Fleet, J.C. Generation of a Transgenic Mouse for Colorectal Cancer Research with Intestinal Cre Expression Limited to the Large Intestine. Mol. Cancer Res. 2010, 8, 1095–1104. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.; Im, S.K.; Fang, S. Mouse Cre-LoxP system: General principles to determine tissue-specific roles of target genes. Lab. Anim. Res. 2018, 34, 147–159. [Google Scholar] [CrossRef]
- Terzic, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- NanoDefine. Development of an Integrated Approach Based on Validated and Standardized Methods to Support the Implementation of the EC Recommendations for a Definition of Nanomaterials. FP7-NMP-2013-LARGE (NMP.2013.1.4–3 2013–2016). 2016. Available online: www.nanodefine.eu (accessed on 17 December 2021).
- Verleysen, E.; Brassinne, F.; Van Steen, F.; Waegeneers, N.; Cheyns, K.; Machiels, R.; Mathioudaki, S.; Jimenez, I.O.; Ledecq, M.; Mast, J. Towards a generic protocol for measuring the constituent particle size distribution of E171 in food by electron microscopy. Food Control 2021, 132, 108492. [Google Scholar] [CrossRef]
- MacNicoll, A.K.M.; Aksoy, H.; Kramer, E.; Bouwmeester, H.; Chaudhry, Q. A study of the uptake and biodistribution of nano-titanium dioxide using in vitro and in vivo models of oral intake. J. Nanopart. Res. Interdiscip. Forum Nanoscale Sci. Technol. 2015, 17, 66. [Google Scholar] [CrossRef]
- Qiagen. RNeasy Mini Handbook. 2012. Available online: www.bea.ki.se/documents/EN-RNeasy%20handbook.pdf (accessed on 17 December 2021).
- Agilent. One-Color Microarray Based Gene Expression Analysis. 2015. Available online: G4140-90040_GeneExpression_OneColor_6.9.pdf (accessed on 17 December 2021).
- Reich, M.; Liefeld, T.; Gould, J.; Lerner, J.; Tamayo, P.; Mesirov, J.P. GenePattern 2.0. Nat. Genet. 2006, 38, 500–501. [Google Scholar] [CrossRef]
- Alonso, R.; Salavert, F.; Garcia-Garcia, F.; Carbonell-Caballero, J.; Bleda, M.; Garcia-Alonso, L.; Sanchis-Juan, A.; Perez-Gil, D.; Marin-Garcia, P.; Sanchez, R.; et al. Babelomics 5.0: Functional interpretation for new generations of genomic data. Nucleic Acids Res. 2015, 43, W117–W121. [Google Scholar] [CrossRef]
- Smyth, G.K. limma: Linear Models for Microarray Data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2005; pp. 397–420. ISBN 1431-8776. [Google Scholar]
- Herwig, R.; Hardt, C.; Lienhard, M.; Kamburov, A. Analyzing and interpreting genome data at the network level with ConsensusPathDB. Nat. Protoc. 2016, 11, 1889–1907. [Google Scholar] [CrossRef] [PubMed]
- Kamburov, A.; Stelzl, U.; Lehrach, H.; Herwig, R. The ConsensusPathDB interaction database: 2013 update. Nucleic Acids Res. 2012, 41, D793–D800. [Google Scholar] [CrossRef] [PubMed]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Kim, E.S.; Kim, J.Y.; Hong, S.T.; Chun, H.J.; Kang, D.E.; Cho, B.R. Measurement of the Nucleus Area and Nucleus/Cytoplasm and Mitochondria/Nucleus Ratios in Human Colon Tissues by Dual-Colour Two-Photon Microscopy Imaging. Sci. Rep. 2015, 5, 18521. [Google Scholar] [CrossRef]
- Proquin, H.; Rodríguez-Ibarra, C.; Moonen, C.; Ortega, I.M.U.; Briedé, J.J.; de Kok, T.M.; van Loveren, H.; Chirino, Y.I. Titanium dioxide food additive (E171) induces ROS formation and genotoxicity: Contribution of micro and nano-sized fractions. Mutagenesis 2018, 33, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Momma, T.; Okayama, H.; Saitou, M.; Sugeno, H.; Yoshimoto, N.; Takebayashi, Y.; Ohki, S.; Takenoshita, S. Expression of circadian clock genes in human colorectal adenoma and carcinoma. Oncol. Lett. 2017, 14, 5319–5325. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, H.; Ji, M.; Wang, Z.; Wu, W. Low circadian clock genes expression in cancers: A meta-analysis of its association with clinicopathological features and prognosis. PLoS ONE 2020, 15, e0233508. [Google Scholar] [CrossRef]
- Yang, S.-F.; Xu, M.; Yang, H.-Y.; Li, P.-Q.; Chi, X.-F. Expression of circadian gene NPAS2 in colorectal cancer and its prognostic significance. Nan Fang Yi Ke Da Xue Xue Bao 2016, 36, 714–718. [Google Scholar]
- Mazzoccoli, G.; Vinciguerra, M.; Papa, G.; Piepoli, A. Circadian clock circuitry in colorectal cancer. World J. Gastroenterol. 2014, 20, 4197–4207. [Google Scholar] [CrossRef]
- Haus, E. Circadian disruption in shiftwork is probably carcinogenic to humans. Chronobiol. Int. 2007, 24, 1255–1256. [Google Scholar]
- Yu, H.; Meng, X.; Wu, J.; Pan, C.; Ying, X.; Zhou, Y.; Liu, R.; Huang, W. Cryptochrome 1 Overexpression Correlates with Tumor Progression and Poor Prognosis in Patients with Colorectal Cancer. PLoS ONE 2013, 8, e61679. [Google Scholar] [CrossRef]
- Mazzoccoli, G.; Colangelo, T.; Panza, A.; Rubino, R.; De Cata, A.; Tiberio, C.; Valvano, M.R.; Pazienza, V.; Merla, G.; Augello, B.; et al. Deregulated expression of cryptochrome genes in human colorectal cancer. Mol. Cancer 2016, 15, 6. [Google Scholar] [CrossRef]
- Ray, S.; Valekunja, U.K.; Stangherlin, A.; Howell, S.A.; Snijders, A.P.; Damodaran, G.; Reddy, A.B. Circadian rhythms in the absence of the clock gene Bmal1. Science 2020, 367, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.-L.; Luo, H.-Y.; Yang, J.; Wu, W.-J.; Chen, D.-L.; Huang, P.; Xu, R.-H. Overexpression of the Circadian Clock Gene Bmal1 Increases Sensitivity to Oxaliplatin in Colorectal Cancer. Clin. Cancer Res. 2014, 20, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Devocelle, A.; Souza, L.; Foudi, A.; Tenreira Bento, S.; Desterke, C.; Sherrard, R.; Ballesta, A.; Adam, R.; Giron-Michel, J.; et al. BMAL1 knockdown triggers different colon carcinoma cell fates by altering the delicate equilibrium between AKT/mTOR and P53/P21 pathways. Aging 2020, 12, 8067–8083. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Li, X.; Li, B.; Li, Y.; Xia, K.; Yang, Y.; Aman, S.; Wang, M.; Wu, H. Circadian protein BMAL1 promotes breast cancer cell invasion and metastasis by up-regulating matrix metalloproteinase9 expression. Cancer Cell Int. 2019, 19, 182. [Google Scholar] [CrossRef]
- Said, A.H.; Raufman, J.-P.; Xie, G. The Role of Matrix Metalloproteinases in Colorectal Cancer. Cancers 2014, 6, 366–375. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Zhou, Y.; Qin, S.; Qiu, Y.; Cui, R.; Yu, M.; Qin, J.; Zhong, M. Promotion of Tumor Growth by ADAMTS4 in Colorectal Cancer: Focused on Macrophages. Cell. Physiol. Biochem. 2018, 46, 1693–1703. [Google Scholar] [CrossRef]
- Filou, S.; Korpetinou, A.; Kyriakopoulou, D.; Bounias, D.; Stavropoulos, M.; Ravazoula, P.; Papachristou, D.J.; Theocharis, A.D.; Vynios, D.H. ADAMTS Expression in Colorectal Cancer. PLoS ONE 2015, 10, e0121209. [Google Scholar] [CrossRef]
- Schmid, F.; Wang, Q.; Huska, M.; A Andrade-Navarro, M.; Lemm, M.; Fichtner, I.; Dahlmann, M.; Kobelt, D.; Walther, W.; Smith, J.; et al. SPON2, a newly identified target gene of MACC1, drives colorectal cancer metastasis in mice and is prognostic for colorectal cancer patient survival. Oncogene 2016, 35, 5942–5952. [Google Scholar] [CrossRef]
- D’Errico, I.; Moschetta, A. Nuclear receptors, intestinal architecture and colon cancer: An intriguing link. Cell. Mol. Life Sci. 2008, 65, 1523–1543. [Google Scholar] [CrossRef]
- Applegate, C.C.; A Lane, M. Role of retinoids in the prevention and treatment of colorectal cancer. World J. Gastrointest. Oncol. 2015, 7, 184–203. [Google Scholar] [CrossRef]
- Wang, W.; Liu, S.; Jiang, C.; Wang, Y.; Zhu, H.; Wang, X. High Expression of RARbeta Is a Favorable Factor in Colorectal Cancer. Dis. Markers 2019, 2019, 7138754. [Google Scholar] [CrossRef] [PubMed]
- Koni, M.; Pinnarò, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Park, J.I. Wnt signaling in cancer: Therapeutic targeting of Wnt signaling beyond beta-catenin and the destruction complex. Exp. Mol. Med. 2020, 52, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Schatoff, E.M.; Leach, B.I.; Dow, L.E. WNT Signaling and Colorectal Cancer. Curr. Color. Cancer Rep. 2017, 13, 101–110. [Google Scholar] [CrossRef]
- Fan, J.; Lv, Z.; Yang, G.; Liao, T.T.; Xu, J.; Wu, F.; Huang, Q.; Guo, M.; Hu, G.; Zhou, M.; et al. Retinoic Acid Receptor-Related Orphan Receptors: Critical Roles in Tumorigenesis. Front. Immunol. 2018, 9, 1187. [Google Scholar] [CrossRef]
- Kathania, M.; Khare, P.; Zeng, M.; Cantarel, B.; Zhang, H.; Ueno, H.; Venuprasad, K. Itch inhibits IL-17-mediated colon inflammation and tumorigenesis by ROR-gammat ubiquitination. Nat. Immunol. 2016, 17, 997–1004. [Google Scholar] [CrossRef]
- Capone, A.; Volpe, E. Transcriptional Regulators of T Helper 17 Cell Differentiation in Health and Autoimmune Diseases. Front. Immunol. 2020, 11, 348. [Google Scholar] [CrossRef]
- Basti, A.; Fior, R.; Yalçin, M.; Póvoa, V.; Astaburuaga, R.; Li, Y.; Naderi, J.; Ferreira, M.G.; Relógio, A. The Core-Clock Gene NR1D1 Impacts Cell Motility In Vitro and Invasiveness in a Zebrafish Xenograft Colon Cancer Model. Cancers 2020, 12, 853. [Google Scholar] [CrossRef]
- Sumei, S.; Xiangyun, K.; Fenrong, C.; Xueguang, S.; Sijun, H.; Bin, B.; Xiaolei, S.; Yongjiu, T.; Kaichun, W.; Qingchuan, Z.; et al. Hypermethylation of DHRS3 as a Novel Tumor Suppressor Involved in Tumor Growth and Prognosis in Gastric Cancer. Front. Cell Dev. Biol. 2021, 9, 624871. [Google Scholar] [CrossRef]
- Cook, D.; Kang, H.S.; Jetten, A.A.M. Retinoic Acid-Related Orphan Receptors (RORs): Regulatory Functions in Immunity, Development, Circadian Rhythm, and Metabolism. Nucl. Recept. Res. 2015, 2, 101185. [Google Scholar] [CrossRef]
- Jetten, A.M. Retinoid-Related Orphan Receptors (RORs): Critical Roles in Development, Immunity, Circadian Rhythm, and Cellular Metabolism. Nucl. Recept. Signal. 2009, 7, e003. [Google Scholar] [CrossRef] [PubMed]
- Bochis, O.V.; Irimie, A.; Pichler, M.; Neagoe, I.B. The Role of Skp2 and its Substrate CDKN1B (p27) in Colorectal Cancer. J. Gastrointest. Liver Dis. 2015, 24, 225–234. [Google Scholar] [CrossRef]
- Hendricks, A.; Gieseler, F.; Nazzal, S.; Bräsen, J.H.; Lucius, R.; Sipos, B.; Claasen, J.H.; Becker, T.; Hinz, S.; Burmeister, G.; et al. Prognostic relevance of topoisomerase II α and minichromosome maintenance protein 6 expression in colorectal cancer. BMC Cancer 2019, 19, 429. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Lu, F.; Shao, Y. The E2F family as potential biomarkers and therapeutic targets in colon cancer. PeerJ 2020, 8, e8562. [Google Scholar] [CrossRef] [PubMed]
- Greally, M.; Kelly, C.M.; Cercek, A. HER2: An emerging target in colorectal cancer. Curr. Probl. Cancer 2018, 42, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Bitar, L.; Zouein, J.; Haddad, F.G.; Eid, R.; Kourie, H.R. HER2 in metastatic colorectal cancer: A new to target to remember. Biomark. Med. 2021, 15, 135–138. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
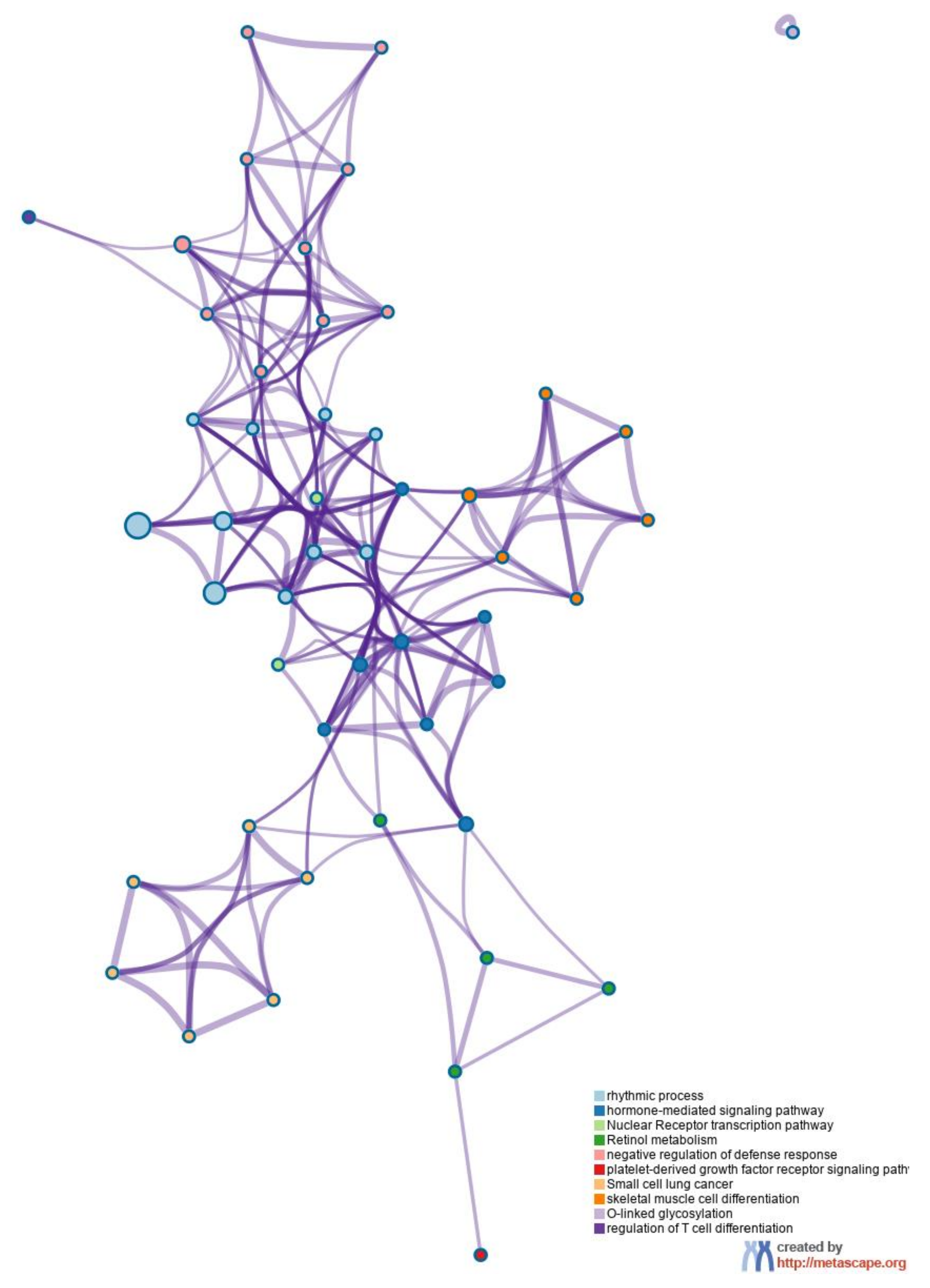{kind=link}
| TEM | DLS | Zeta-Potential | ||||
|---|---|---|---|---|---|---|
| Fmax (nm) Median ± 95% CI | Fmin (nm) Median ± 95% CI | Particles < 100 nm (%) | Z-Average (nm) Mean ± SD | PDI Mean ± SD | (mV) Mean ± SD | |
| 1 mg/mL | 107 (±8.3) | 79 (±5.4) | ~64 | 315.3 (±92.4) | 0.246 (±0.02) | −29.9 (±6.0) |
| 2 mg/mL | 106 (±11.4) | 75 (±7.2) | ~64 | 318.1 (±96.6) | 0.218 (±0.02) | −28.7 (±7.8) |
| 5 mg/mL | 110 (±7.7) | 79 (±4.8) | ~63 | 348.7 (±140.5) | 0.192 (±0.01) | −27.1 (±9.4) |
| Day 2 | Day 7 | Day 14 | Day 21 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (mg/kgbw/day) | 1 | 2 | 5 | 1 | 2 | 5 | 1 | 2 | 5 | 1 | 2 | 5 |
| absolute FC ≥ 1.5 | 542 | 382 | 950 | 650 | 373 | 724 | 643 | 396 | 489 | 1095 | 534 | 584 |
| Upregulated | 231 | 124 | 262 | 360 | 202 | 341 | 202 | 188 | 168 | 499 | 352 | 386 |
| Downregulated | 311 | 258 | 688 | 290 | 171 | 383 | 441 | 208 | 321 | 596 | 182 | 198 |
| p-value < 0.05 | 1657 | 752 | 1534 | 1403 | 988 | 1025 | 1421 | 501 | 1180 | 1517 | 562 | 1172 |
| q-value < 0.05 | 61 | 0 | 1 | 15 | 1 | 0 | 16 | 2 | 7 | 8 | 0 | 0 |
| absolute FC ≥ 1.5 and p-value < 0.05 | 311 | 129 | 337 | 328 | 156 | 357 | 282 | 135 | 238 | 489 | 129 | 246 |
| absolute FC ≥ 1.5 and q-value < 0.05 | 49 | 0 | 1 | 14 | 1 | 0 | 14 | 1 | 6 | 8 | 0 | 0 |
| Biological Function | Day | Pathway | Source | p-Value | q-Value |
|---|---|---|---|---|---|
| Cancer | 2 | Small cell lung cancer–Mus musculus (mouse) | KEGG | 2.16 × 10−3 | 1.73 × 10−2 |
| 2 | Gastric cancer–Mus musculus (mouse) | KEGG | 6.63 × 10−3 | 2.84 × 10−2 | |
| Cell cycle | 2 | Cell cycle | Wikipathways | 1.83 × 10−3 | 1.73 × 10−2 |
| 2 | Mitotic G1-G1/S phases | Reactome | 2.16 × 10−3 | 1.73 × 10−2 | |
| 2 | Orc1 removal from chromatin | Reactome | 2.43 × 10−3 | 1.73 × 10−2 | |
| 2 | Cell cycle–Mus musculus (mouse) | KEGG | 4.96 × 10−3 | 2.64 × 10−2 | |
| 2 | Cyclin D associated events in G1 | Reactome | 6.77 × 10−3 | 2.84 × 10−2 | |
| 2 | G1 Phase | Reactome | 6.77 × 10−3 | 2.84 × 10−2 | |
| 2 | Switching of origins to a post-replicative state | Reactome | 7.09 × 10−3 | 2.84 × 10−2 | |
| Circadian rhythm | 2 | Circadian rhythm–Mus musculus (mouse) | KEGG | 1.48 × 10−8 | 9.49 × 10−7 |
| 2 | Exercise-induced circadian regulation | Wikipathways | 1.10 × 10−5 | 3.53 × 10−4 | |
| Disease | 2 | Hepatitis C–Mus musculus (mouse) | KEGG | 8.93 × 10−3 | 3.17 × 10−2 |
| Gene expression (transcription) | 2 | Nuclear receptor transcription pathway | Reactome | 3.25 × 10−4 | 4.16 × 10−3 |
| 2 | Nuclear receptors | Wikipathways | 4.74 × 10−3 | 2.64 × 10−2 | |
| Metabolism | 2 | Retinol metabolism | Wikipathways | 3.73 × 10−3 | 2.39 × 10−2 |
| 2 | Signaling by retinoic acid | Reactome | 7.74 × 10−3 | 2.92 × 10−2 | |
| Post-translational modification/ metabolism | 2 | O-glycosylation of TSR-domain-containing proteins | Reactome | 1.20 × 10−4 | 2.12 × 10−3 |
| 2 | O-linked glycosylation | Reactome | 1.33 × 10−4 | 2.12 × 10−3 | |
| Circadian rhythm | 7 | Circadian rhythm–Mus musculus (mouse) | KEGG | 1.11 × 10−8 | 8.87 × 10−8 |
| 7 | Exercise-induced circadian regulation | Wikipathways | 1.29 × 10−5 | 3.45 × 10−5 | |
| Gene expression (transcription) | 7 | Nuclear receptor transcription pathway | Reactome | 1.14 × 10−5 | 3.45 × 10−5 |
| 7 | Nuclear receptors | Wikipathways | 5.48 × 10−4 | 1.10 × 10−3 | |
| Circadian rhythm | 14 | Circadian rhythm–Mus musculus (mouse) | KEGG | 2.48 × 10−6 | 1.98 × 10−5 |
| 14 | Exercise-induced circadian regulation | Wikipathways | 1.29 × 10−5 | 3.45 × 10−5 | |
| Gene expression (transcription) | 14 | Nuclear receptor transcription pathway | Reactome | 1.14 × 10−5 | 3.45 × 10−5 |
| 14 | Nuclear receptors | Wikipathways | 5.48 × 10−4 | 1.10 × 10−3 | |
| Circadian rhythm | 21 | Circadian rhythm–Mus musculus (mouse) | KEGG | 7.84 × 10−5 | 2.74 × 10−4 |
| 21 | Exercise-induced circadian regulation | Wikipathways | 2.33 × 10−4 | 4.08 × 10−4 | |
| Gene expression (transcription) | 21 | Nuclear receptor transcription pathway | Reactome | 9.63 × 10−7 | 6.74 × 10−6 |
| 21 | Nuclear receptors | Wikipathways | 1.23 × 10−4 | 2.87 × 10−4 | |
| 21 | Generic transcription pathway | Reactome | 3.31 × 10−3 | 4.64 × 10−3 | |
| 21 | RNA polymerase II transcription | Reactome | 5.19 × 10−3 | 6.06 × 10−3 | |
| 21 | Gene expression (transcription) | Reactome | 7.44 × 10−3 | 7.44 × 10−3 |
| Biological Function | Dose (mg/kgbw/Day) | Cluster | Pathway | Source | p-Value | q-Value |
|---|---|---|---|---|---|---|
| Signaling | 1 | 1 | G alpha (s) signaling events | Reactome | 1.77 × 10−3 | 1.40 × 10−2 |
| 1 | 1 | Olfactory transduction–Mus musculus (mouse) | KEGG | 2.33 × 10−3 | 1.40 × 10−2 | |
| 1 | 1 | Olfactory signaling pathway | Reactome | 4.45 × 10−3 | 1.78 × 10−2 | |
| Signaling | 1 | 2 | Olfactory transduction–Mus musculus (mouse) | KEGG | 3.59 × 10−7 | 4.67 × 10−6 |
| 1 | 2 | Neuroactive ligand–receptor interaction–Mus musculus (mouse) | KEGG | 3.02 × 10−3 | 1.96 × 10−2 | |
| 1 | 2 | Olfactory signaling pathway | Reactome | 5.72 × 10−3 | 2.48 × 10−2 | |
| Signaling | 1 | 3 | RAF/MAP kinase cascade | Reactome | 5.33 × 10−3 | 2.89 × 10−2 |
| 1 | 3 | MAPK1/MAPK3 signaling | Reactome | 5.79 × 10−3 | 2.89 × 10−2 | |
| 1 | 3 | MAPK family signaling cascades | Reactome | 8.20 × 10−3 | 3.59 × 10−2 | |
| Circadian rhythm | 1 | 3 | Circadian rhythm–Mus musculus (mouse) | KEGG | 2.53 × 10−5 | 8.86 × 10−4 |
| 1 | 3 | Exercise-induced circadian regulation | Wikipathways | 4.62 × 10−3 | 2.89 × 10−2 | |
| Disease | 1 | 3 | Lung fibrosis | Wikipathways | 4.81 × 10−3 | 2.89 × 10−2 |
| Immune response | 1 | 3 | Cytokine–cytokine receptor interaction–Mus musculus (mouse) | KEGG | 9.41 × 10−3 | 3.66 × 10−2 |
| Post-translational modification/Metabolism | 1 | 3 | O-glycosylation of TSR domain-containing proteins | Reactome | 2.30 × 10−3 | 2.89 × 10−2 |
| Signaling | 1 | 3 | Hedgehog signaling pathway–Mus musculus (mouse) | KEGG | 3.88 × 10−3 | 2.89 × 10−2 |
| Disease | 2 | 1 | Epstein–Barr virus infection–Mus musculus (mouse) | KEGG | 4.85 × 10−3 | 2.18 × 10−2 |
| Immune response | 2 | 1 | Cell adhesion molecules (CAMs)–Mus musculus (mouse) | KEGG | 1.46 × 10−3 | 1.39 × 10−2 |
| 2 | 1 | Regulation of complement cascade | Reactome | 1.59 × 10−3 | 1.39 × 10−2 | |
| 2 | 1 | Complement cascade | Reactome | 2.31 × 10−3 | 1.39 × 10−2 | |
| 2 | 1 | B cell receptor signaling pathway–Mus musculus (mouse) | KEGG | 6.42 × 10−3 | 2.21 × 10−2 | |
| 2 | 1 | Hematopoietic cell lineage–Mus musculus (mouse) | KEGG | 7.36 × 10−3 | 2.21 × 10−2 | |
| 2 | 1 | Immunoregulatory interactions between a lymphoid and a non-lymphoid cell | Reactome | 8.76 × 10−3 | 2.25 × 10−2 | |
| Disease | 5 | 1 | Type I diabetes mellitus–Mus musculus (mouse) | KEGG | 2.20 × 10−3 | 9.83 × 10−3 |
| Immune response | 5 | 1 | Immunoregulatory interactions between a lymphoid and a non-lymphoid cell | Reactome | 4.33 × 10−7 | 2.03 × 10−5 |
| 5 | 1 | Hematopoietic cell lineage–Mus musculus (mouse) | KEGG | 4.47 × 10−6 | 1.40 × 10−4 | |
| 5 | 1 | Immune system | Reactome | 1.22 × 10−5 | 2.86 × 10−4 | |
| 5 | 1 | Adaptive immune system | Reactome | 4.45 × 10−5 | 5.23 × 10−4 | |
| 5 | 1 | Chemokine receptors bind chemokines | Reactome | 5.05 × 10−5 | 5.27 × 10−4 | |
| 5 | 1 | Cell adhesion molecules (CAMs)–Mus musculus (mouse) | KEGG | 1.16 × 10−4 | 9.65 × 10−4 | |
| 5 | 1 | Cytokine–cytokine receptor interaction—Mus musculus (mouse) | KEGG | 1.87 × 10−4 | 1.33 × 10−3 | |
| 5 | 1 | B cell receptor signaling pathway | Wikipathways | 2.82 × 10−4 | 1.76 × 10−3 | |
| 5 | 1 | B cell receptor signaling pathway–Mus musculus (mouse) | KEGG | 7.17 × 10−4 | 3.97 × 10−3 | |
| 5 | 1 | Regulation of complement cascade | Reactome | 1.03 × 10−3 | 5.40 × 10−3 | |
| 5 | 1 | Complement cascade | Reactome | 1.79 × 10−3 | 8.85 × 10−3 | |
| 5 | 1 | Complement and coagulation cascades | Wikipathways | 2.50 × 10−3 | 1.07 × 10−2 | |
| 5 | 1 | Signaling by the B cell receptor (BCR) | Reactome | 2.99 × 10−3 | 1.22 × 10−2 | |
| 5 | 1 | TNF receptor superfamily (TNFSF) members mediating non-canonical NF-kB pathway | Reactome | 3.44 × 10−3 | 1.35 × 10−2 | |
| 5 | 1 | Costimulation by the CD28 family | Reactome | 5.31 × 10−3 | 1.85 × 10−2 | |
| 5 | 1 | Complement and coagulation cascades–Mus musculus (mouse) | KEGG | 6.08 × 10−3 | 1.98 × 10−2 | |
| 5 | 1 | Innate immune system | Reactome | 6.10 × 10−3 | 1.98 × 10−2 | |
| Signaling | 5 | 1 | Olfactory transduction–Mus musculus (mouse) | KEGG | 6.64 × 10−8 | 6.24 × 10−6 |
| 5 | 1 | GPCR downstream signaling | Reactome | 2.39 × 10−5 | 4.50 × 10−4 | |
| 5 | 1 | Signaling by GPCR | Reactome | 3.42 × 10−5 | 5.15 × 10−4 | |
| 5 | 1 | Olfactory signaling pathway | Reactome | 3.84 × 10−5 | 5.15 × 10−4 | |
| 5 | 1 | Peptide-ligand-binding receptors | Reactome | 9.38 × 10−5 | 8.82 × 10−4 | |
| 5 | 1 | GPCR ligand binding | Reactome | 1.23 × 10−4 | 9.65 × 10−4 | |
| 5 | 1 | Class A/1 (rhodopsin-like receptors) | Reactome | 1.98 × 10−4 | 1.33 × 10−3 | |
| 5 | 1 | G alpha (s) signaling events | Reactome | 3.46 × 10−4 | 2.03 × 10−3 | |
| 5 | 1 | Peptide GPCRs | Wikipathways | 2.20 × 10−3 | 9.83 × 10−3 | |
| 5 | 1 | Signal transduction | Reactome | 3.64 × 10−3 | 1.37 × 10−2 | |
| 5 | 1 | GPCRs, other | Wikipathways | 5.06 × 10−3 | 1.83 × 10−2 | |
| 5 | 1 | Generation of second messenger molecules | Reactome | 8.05 × 10−3 | 2.52 × 10−2 | |
| Gene expression | 5 | 2 | Regulation of TP53 activity through phosphorylation | Reactome | 1.07 × 10−4 | 3.70 × 10−4 |
| Cell cycle | 5 | 2 | Cell cycle | Reactome | 1.25 × 10−24 | 1.69 × 10−22 |
| 5 | 2 | Cell cycle, mitotic | Reactome | 1.70 × 10−23 | 1.14 × 10−21 | |
| 5 | 2 | Resolution of sister chromatid cohesion | Reactome | 3.13 × 10−23 | 1.41 × 10−21 | |
| 5 | 2 | Mitotic prometaphase | Reactome | 3.49 × 10−22 | 1.18 × 10−20 | |
| 5 | 2 | Cell cycle checkpoints | Reactome | 6.95 × 10−21 | 1.88 × 10−19 | |
| 5 | 2 | M phase | Reactome | 6.48 × 10−19 | 1.46 × 10−17 | |
| 5 | 2 | Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal | Reactome | 2.53 × 10−17 | 4.27 × 10−16 | |
| 5 | 2 | Amplification of signal from the kinetochores | Reactome | 2.53 × 10−17 | 4.27 × 10−16 | |
| 5 | 2 | Separation of sister chromatids | Reactome | 4.59 × 10−17 | 6.88 × 10−16 | |
| 5 | 2 | Mitotic anaphase | Reactome | 6.62 × 10−17 | 8.94 × 10−16 | |
| 5 | 2 | Mitotic metaphase and anaphase | Reactome | 7.46 × 10−17 | 9.16 × 10−16 | |
| 5 | 2 | Mitotic spindle checkpoint | Reactome | 2.52 × 10−16 | 2.83 × 10−15 | |
| 5 | 2 | G2/M transition | Reactome | 5.16 × 10−10 | 4.35 × 10−9 | |
| 5 | 2 | Mitotic G2-G2/M phases | Reactome | 6.08 × 10−10 | 4.82 × 10−9 | |
| 5 | 2 | APC/C-mediated degradation of cell cycle proteins | Reactome | 1.25 × 10−9 | 8.86 × 10−9 | |
| 5 | 2 | Regulation of mitotic cell cycle | Reactome | 1.25 × 10−9 | 8.86 × 10−9 | |
| 5 | 2 | The role of GTSE1 in G2/M progression after G2 checkpoint | Reactome | 1.54 × 10−9 | 1.04 × 10−8 | |
| 5 | 2 | Cyclin A/B1/B2-associated events during G2/M transition | Reactome | 2.04 × 10−9 | 1.31 × 10−8 | |
| 5 | 2 | Activation of NIMA kinases NEK9, NEK6, and NEK7 | Reactome | 2.42 × 10−8 | 1.48 × 10−7 | |
| 5 | 2 | Activation of APC/C- and APC/C:Cdc20-mediated degradation of mitotic proteins | Reactome | 4.40 × 10−7 | 2.48 × 10−6 | |
| 5 | 2 | G2/M DNA replication checkpoint | Reactome | 1.39 × 10−6 | 7.22 × 10−6 | |
| 5 | 2 | Cell cycle–Mus musculus (mouse) | KEGG | 2.43 × 10−6 | 1.22 × 10−5 | |
| 5 | 2 | Phosphorylation of Emi1 | Reactome | 2.77 × 10−6 | 1.34 × 10−5 | |
| 5 | 2 | Regulation of PLK1 activity at G2/M transition | Reactome | 4.21 × 10−6 | 1.96 × 10−5 | |
| 5 | 2 | Cell cycle | Wikipathways | 4.52 × 10−6 | 2.03 × 10−5 | |
| 5 | 2 | TP53-regulated transcription of genes Involved in G2 cell cycle arrest | Reactome | 4.83 × 10−6 | 2.10 × 10−5 | |
| 5 | 2 | APC/C:Cdc20-mediated degradation of mitotic proteins | Reactome | 1.51 × 10−5 | 6.35 × 10−5 | |
| 5 | 2 | TP53-regulates transcription of cell cycle genes | Reactome | 2.25 × 10−5 | 9.22 × 10−5 | |
| 5 | 2 | AURKA activation by TPX2 | Reactome | 2.97 × 10−5 | 1.18 × 10−4 | |
| 5 | 2 | Regulation of APC/C activators between G1/S and early anaphase | Reactome | 3.24 × 10−5 | 1.25 × 10−4 | |
| 5 | 2 | Condensation of prophase chromosomes | Reactome | 3.86 × 10−5 | 1.45 × 10−4 | |
| 5 | 2 | G2/M checkpoints | Reactome | 7.26 × 10−5 | 2.65 × 10−4 | |
| 5 | 2 | Nuclear envelope breakdown | Reactome | 8.12 × 10−5 | 2.88 × 10−4 | |
| 5 | 2 | Phosphorylation of the APC/C | Reactome | 1.28 × 10−4 | 4.32 × 10−4 | |
| 5 | 2 | Regulation of TP53 activity | Reactome | 1.46 × 10−4 | 4.80 × 10−4 | |
| 5 | 2 | Polo-like-kinase-mediated events | Reactome | 2.72 × 10−4 | 8.76 × 10−4 | |
| 5 | 2 | APC/C:Cdh1-mediated degradation of Cdc20 and other APC/C:Cdh1-targeted proteins in late mitosis/early G1 | Reactome | 2.97 × 10−4 | 8.91 × 10−4 | |
| 5 | 2 | Deposition of new CENPA-containing nucleosomes at the centromere | Reactome | 2.97 × 10−4 | 8.91 × 10−4 | |
| 5 | 2 | Nucleosome assembly | Reactome | 2.97 × 10−4 | 8.91 × 10−4 | |
| 5 | 2 | p53 signaling | Wikipathways | 3.73 × 10−4 | 1.08 × 10−3 | |
| 5 | 2 | APC:Cdc20-mediated degradation of cell cycle proteins prior to satisfation of the cell cycle checkpoint | Reactome | 3.75 × 10−4 | 1.08 × 10−3 | |
| 5 | 2 | Mitotic prophase | Reactome | 4.19 × 10−4 | 1.18 × 10−3 | |
| 5 | 2 | p53 signaling pathway–Mus musculus (mouse) | KEGG | 4.69 × 10−4 | 1.28 × 10−3 | |
| 5 | 2 | Transcriptional regulation by TP53 | Reactome | 4.76 × 10−4 | 1.28 × 10−3 | |
| 5 | 2 | Nuclear pore complex (NPC) disassembly | Reactome | 7.47 × 10−4 | 1.96 × 10−3 | |
| 5 | 2 | Establishment of sister chromatid cohesion | Reactome | 1.47 × 10−3 | 3.48 × 10−3 | |
| 5 | 2 | Depolymerisation of the nuclear lamina | Reactome | 1.47 × 10−3 | 3.48 × 10−3 | |
| 5 | 2 | Chk1/Chk2(Cds1)-mediated inactivation of the cyclin B:Cdk1 complex | Reactome | 1.76 × 10−3 | 4.09 × 10−3 | |
| 5 | 2 | Chromosome maintenance | Reactome | 2.44 × 10−3 | 5.58 × 10−3 | |
| 5 | 2 | G2/M DNA damage checkpoint | Reactome | 3.70 × 10−3 | 8.32 × 10−3 | |
| 5 | 2 | Transcriptional regulation by RUNX2 | Reactome | 3.99 × 10−3 | 8.83 × 10−3 | |
| 5 | 2 | Loss of Nlp from mitotic centrosomes | Reactome | 5.07 × 10−3 | 1.07 × 10−2 | |
| 5 | 2 | Loss of proteins required for interphase microtubule organization from the centrosome | Reactome | 5.07 × 10−3 | 1.07 × 10−2 | |
| 5 | 2 | Hedgehog signaling pathway | Wikipathways | 5.42 × 10−3 | 1.13 × 10−2 | |
| 5 | 2 | APC/C:Cdc20-mediated degradation of cyclin B | Reactome | 5.94 × 10−3 | 1.22 × 10−2 | |
| 5 | 2 | HDR through homologous recombination (HR) or single-strand annealing (SSA) | Reactome | 6.97 × 10−3 | 1.36 × 10−2 | |
| 5 | 2 | Recruitment of mitotic centrosome proteins and complexes | Reactome | 6.97 × 10−3 | 1.36 × 10−2 | |
| 5 | 2 | Centrosome maturation | Reactome | 6.97 × 10−3 | 1.36 × 10−2 | |
| 5 | 2 | Recruitment of NuMA to mitotic centrosomes | Reactome | 7.24 × 10−3 | 1.40 × 10−2 | |
| 5 | 2 | COPI-dependent Golgi-to-ER retrograde traffic | Reactome | 8.06 × 10−3 | 1.53 × 10−2 | |
| 5 | 2 | Cdc20:Phospho-APC/C-mediated degradation of Cyclin A | Reactome | 8.25 × 10−3 | 1.55 × 10−2 | |
| Development | 5 | 2 | Progesterone-mediated oocyte maturation–Mus musculus (mouse) | KEGG | 2.15 × 10−7 | 1.26 × 10−6 |
| 5 | 2 | Oocyte meiosis–Mus musculus (mouse) | KEGG | 7.86 × 10−7 | 4.24 × 10−6 | |
| DNA damage | 5 | 2 | miRNA regulation of DNA damage response | Wikipathways | 4.06 × 10−3 | 8.85 × 10−3 |
| 5 | 2 | Homology-directed repair | Reactome | 8.64 × 10−3 | 1.60 × 10−2 | |
| Haemostasis | 5 | 2 | Kinesins | Reactome | 1.12 × 10−3 | 2.80 × 10−3 |
| Senescence | 5 | 2 | Cellular senescence–Mus musculus (mouse) | KEGG | 1.31 × 10−3 | 3.22 × 10−3 |
| Signaling | 5 | 2 | RHO GTPase effectors | Reactome | 5.91 × 10−16 | 5.93 × 10−15 |
| 5 | 2 | RHO GTPases-activated formins | Reactome | 6.15 × 10−16 | 5.93 × 10−15 | |
| 5 | 2 | Signaling by RHO GTPases | Reactome | 7.96 × 10−13 | 7.17 × 10−12 | |
| 5 | 2 | RHO-GTPases-activated CIT | Reactome | 7.55 × 10−4 | 1.96 × 10−3 | |
| 5 | 2 | Signal transduction | Reactome | 8.16 × 10−4 | 2.08 × 10−3 | |
| Extracellular matrix organization | 5 | 3 | Activation of matrix metalloproteinases | Reactome | 9.91 × 10−3 | 5.25 × 10−2 |
| Haemostasis | 5 | 3 | Blood clotting cascade | Wikipathways | 1.35 × 10−3 | 1.02 × 10−2 |
| 5 | 3 | Dissolution of fibrin clot | Reactome | 3.52 × 10−3 | 2.34 × 10−2 | |
| 5 | 3 | Response to elevated platelet cytosolic Ca2+ | Reactome | 5.05 × 10−3 | 2.97 × 10−2 | |
| Signaling | 5 | 3 | Olfactory transduction–Mus musculus (mouse) | KEGG | 1.64 × 10−26 | 8.69 × 10−25 |
| 5 | 3 | Olfactory signaling pathway | Reactome | 2.26 × 10−9 | 5.99 × 10−8 | |
| 5 | 3 | G alpha (s) signaling events | Reactome | 2.66 × 10−6 | 4.70 × 10−5 | |
| 5 | 3 | Signaling by GPCR | Reactome | 8.96 × 10−5 | 1.19 × 10−3 | |
| 5 | 3 | GPCR downstream signaling | Reactome | 2.94 × 10−4 | 2.70 × 10−3 | |
| Transport | 5 | 3 | Transport of fatty acids | Reactome | 3.06 × 10−4 | 2.70 × 10−3 |
| Immune response | 5 | 4 | Inflammatory mediator regulation of TRP channels–Mus musculus (mouse) | KEGG | 6.85 × 10−3 | 2.74 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bischoff, N.S.; Proquin, H.; Jetten, M.J.; Schrooders, Y.; Jonkhout, M.C.M.; Briedé, J.J.; van Breda, S.G.; Jennen, D.G.J.; Medina-Reyes, E.I.; Delgado-Buenrostro, N.L.; et al. The Effects of the Food Additive Titanium Dioxide (E171) on Tumor Formation and Gene Expression in the Colon of a Transgenic Mouse Model for Colorectal Cancer. Nanomaterials 2022, 12, 1256. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12081256
Bischoff NS, Proquin H, Jetten MJ, Schrooders Y, Jonkhout MCM, Briedé JJ, van Breda SG, Jennen DGJ, Medina-Reyes EI, Delgado-Buenrostro NL, et al. The Effects of the Food Additive Titanium Dioxide (E171) on Tumor Formation and Gene Expression in the Colon of a Transgenic Mouse Model for Colorectal Cancer. Nanomaterials. 2022; 12(8):1256. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12081256
Chicago/Turabian StyleBischoff, Nicolaj S., Héloïse Proquin, Marlon J. Jetten, Yannick Schrooders, Marloes C. M. Jonkhout, Jacco J. Briedé, Simone G. van Breda, Danyel G. J. Jennen, Estefany I. Medina-Reyes, Norma L. Delgado-Buenrostro, and et al. 2022. "The Effects of the Food Additive Titanium Dioxide (E171) on Tumor Formation and Gene Expression in the Colon of a Transgenic Mouse Model for Colorectal Cancer" Nanomaterials 12, no. 8: 1256. https://0-doi-org.brum.beds.ac.uk/10.3390/nano12081256