A Rapid and Easy-to-Perform Method of Nucleic-Acid Based Dengue Virus Diagnosis Using Fluorescence-Based Molecular Beacons



Abstract
:
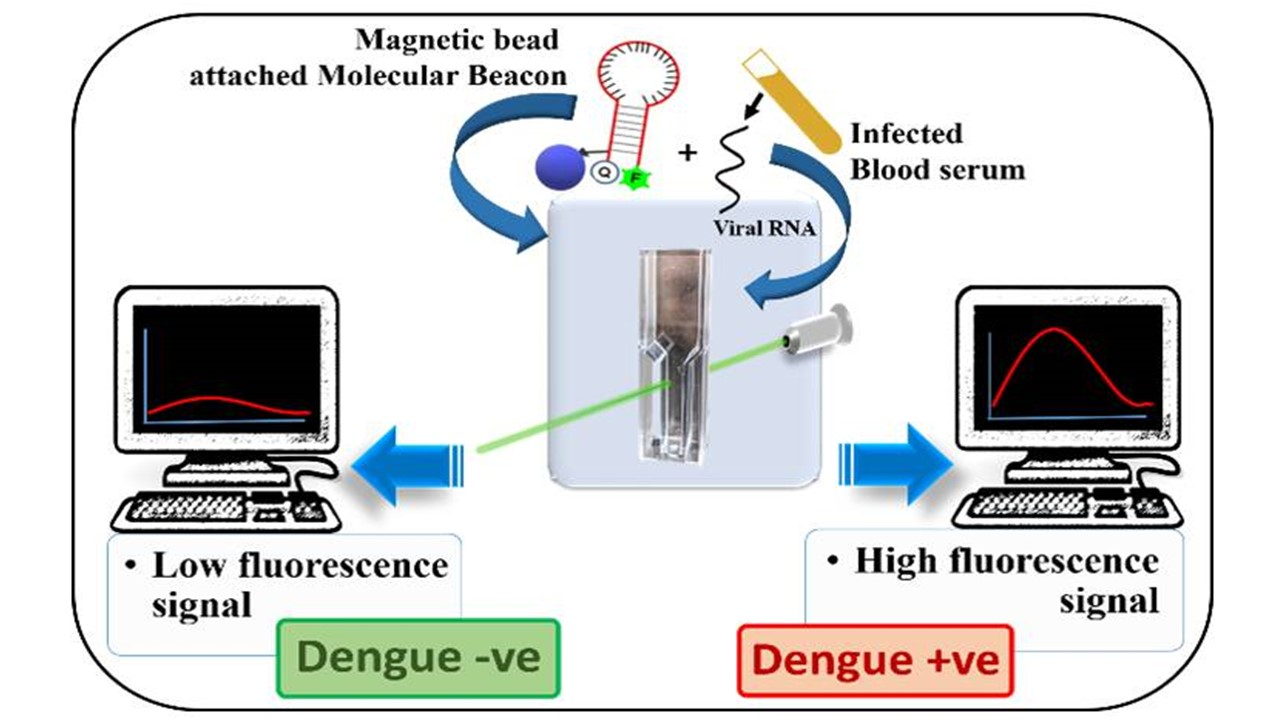{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Dengue NS1 ELISA-Positive Human Serum Samples
2.3. Virus Culture
2.4. Sequence Selection for Designing Molecular Beacon
2.5. Fluorophore-Quencher Selection
2.6. Molecular Beacon Designing
2.7. Standardization of Different Parameters in Fluorometer
- Excitation slit width: 1.25 mm
- Emission slit Width: 1.25 mm
- Voltage: 121 V
- Integration time: 0.1 s
- Excitation wavelength: 495 nm
- Emission range: 505–650 nm
- Maximum emission wavelength: 519 nm
- Volume of the mixture: 20 µL
- Incubation temperature: 50 °C
- Incubation time: 15 min
2.8. Molecular Beacon Experiments
2.8.1. Checking for the Sensitivity
2.8.2. Checking for the Specificity
2.8.3. Testing of Viral RNA as a Target
2.9. Experiments with Biotinylated Molecular Beacons
2.10. Immobilization of the Molecular Beacons with the Magnetic Beads
2.11. Testing Viral RNA with the Conjugated Molecular Beacons and Magnetic Beads
2.12. Detection of Viral RNA by Processing the Virus In Situ
2.13. Detailed Protocol for the Detection of Dengue Virus in Serum Samples Directly
3. Results
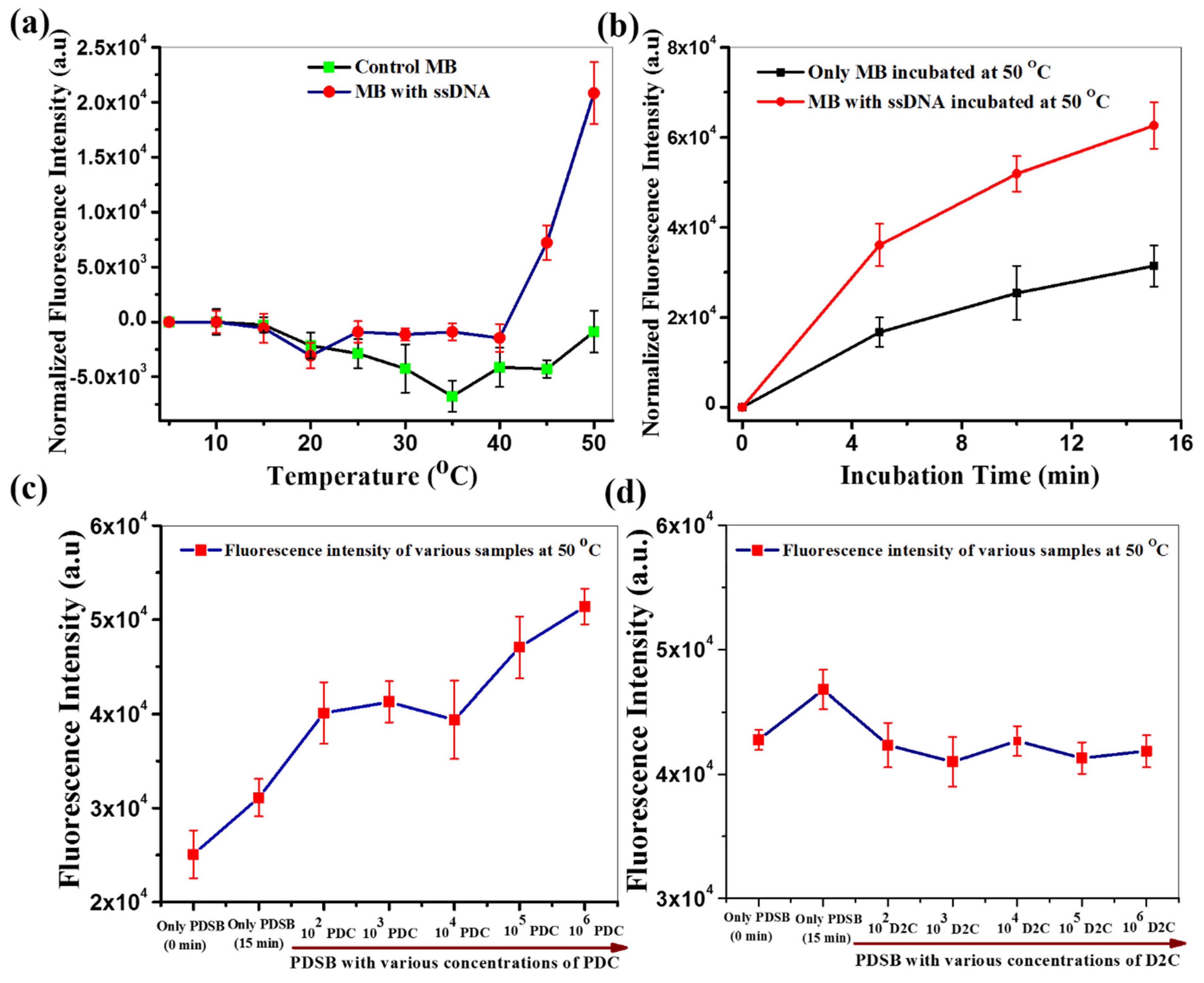
3.1. Standardization of Molecular Beacon
3.1.1. Checking for the Sensitivity
3.1.2. Checking for the Specificity
3.1.3. Testing of Viral RNA as a Target Using Molecular Beacon Only
3.2. Experiments with Biotinylated Molecular Beacons
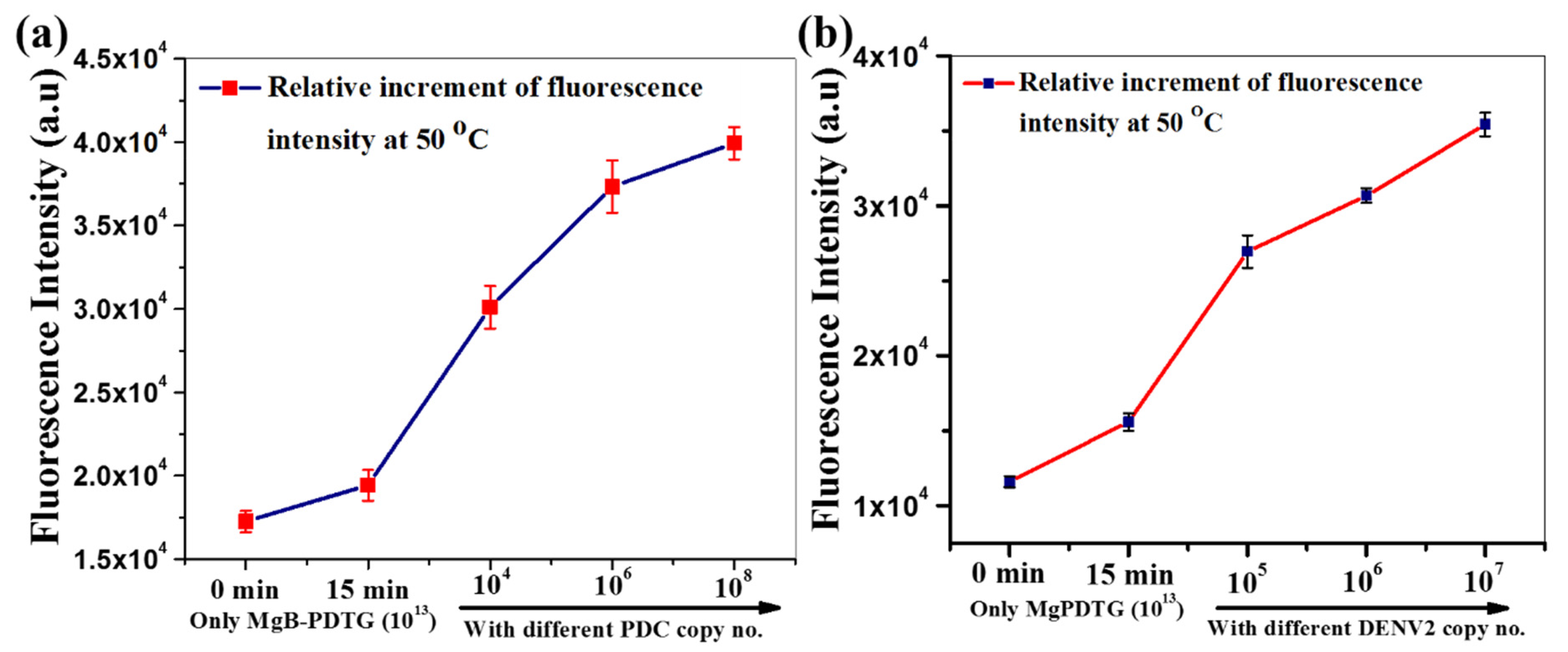
3.2.1. Testing Viral RNA with the Conjugated Molecular Beacons and Magnetic Beads
3.2.2. Detection of Viral RNA by Processing the Virus In Situ
3.3. Clinical Samples Testing
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Improving Data for Dengue. Available online: https://www.who.int/activities/improving-data-for-dengue (accessed on 10 September 2021).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Nath, H.; Mallick, A.; Roy, S.; Sukla, S.; Biswas, S. Computational modelling supports that dengue virus envelope antibodies can bind to SARS-CoV-2 receptor binding sites: Is pre-exposure to dengue virus protective against COVID-19 severity? Comput. Struct. Biotechnol. J. 2021, 19, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Keler, S.; Kolodny, R.; Ben-Tal, N.; Atias-Varon, D.; Shlush, E.; Gerlic, M.; Munitz, A.; Doolman, R.; Asraf, K.; et al. Potential Antigenic cross-reactivity between severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and dengue viruses. Clin. Infect. Dis. 2021, 73, e2444–e2449. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Lee, C.K.; Lam, L.T.M.; Yan, B.; Chua, Y.X.; Lim, A.Y.N.; Phang, K.F.; Kew, G.S.; Teng, H.; Ngai, C.H.; et al. Covert COVID-19 and false-positive dengue serology in Singapore. Lancet Infect. Dis. 2020, 20, 536. [Google Scholar] [CrossRef] [Green Version]
- Nath, H.; Mallick, A.; Roy, S.; Sukla, S.; Basu, K. Dengue antibodies can cross-react with SARS-CoV-2 and vice versa-Antibody detection kits can give false-positive results for both viruses in regions where both COVID-19 and Dengue co-exist. MedRxiv 2020. [Google Scholar] [CrossRef]
- Kembuan, G.J. Dengue serology in Indonesian COVID-19 patients: Coinfection or serological overlap? IDCases 2020, 22, e00927. [Google Scholar] [CrossRef]
- Bicudo, N.; Bicudo, E.; Costa, J.D.; Castro, J.A.L.P.; Barra, G.B. Co-infection of SARS-CoV-2 and dengue virus: A clinical challenge. Braz. J. Infect. Dis. 2020, 24, 452–454. [Google Scholar] [CrossRef]
- Butt, M.H.; Ahmad, A.; Misbah, S.; Mallhi, T.H.; Khan, Y.H. Dengue fever and COVID-19 coinfection; a threat to public health for coepidemic in Pakistan. J. Med. Virol. 2020, 93, 671–672. [Google Scholar] [CrossRef] [PubMed]
- Parkash, O.; Shueb, R.H. Diagnosis of dengue infection using conventional and biosensor based techniques. Viruses 2015, 7, 5410–5427. [Google Scholar] [CrossRef]
- Muller, D.A.; Depelsenaire, A.C.; Young, P.R. Clinical and laboratory diagnosis of dengue virus infection. J. Infect. Dis. 2017, 215, S89–S95. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Waggoner, J.J.; Abeynayake, J.; Sahoo, M.K.; Gresh, L.; Tellez, Y.; Gonzalez, K.; Ballesteros, G.; Pierro, A.M.; Gaibani, P.; Guo, F.P.; et al. Single-reaction, multiplex, real-time RT-PCR for the detection, quantitation, and serotyping of dengue viruses. PLoS Negl. Trop. Dis. 2013, 7, e2116. [Google Scholar] [CrossRef] [PubMed]
- Sukla, S.; Ghosh, A.; Saha, R.; De, A.; Adhya, S.; Biswas, S. In-depth molecular analysis of a small cohort of human and Aedes mosquito (adults and larvae) samples from Kolkata revealed absence of Zika but high prevalence of dengue virus. J. Med. Microbiol. 2018, 67, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.S.; Mathews, D.H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Marras, S.A.E. Interactive fluorophore and quencher pairs for labeling fluorescent nucleic acid hybridization probes. Mol. Biotechnol. 2008, 38, 247–255. [Google Scholar] [CrossRef]
- Marras, S.A.E. Selection of fluorophore and quencher pairs for fluorescent nucleic acid hybridization probes. Methods Mol. Biol. 2006, 335, 3–16. [Google Scholar]
- Mukherjee, S.; Dutta, S.K.; Sengupta, S.; Tripathi, A. Evidence of dengue and chikungunya virus co-infection and circulation of multiple dengue serotypes in a recent Indian outbreak. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2273–2279. [Google Scholar] [CrossRef]
- Kulkarni, R.; Sapkal, G.N.; Kaushal, H.; Mourya, D.T. Japanese Encephalitis: A brief review on Indian perspectives. Open Virol. J. 2018, 31, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, P.D.; Malhotra, B.; Sapkal, G.; Nyayanit, D.A.; Deshpande, G.; Gupta, N.; Padinjaremattathil, U.T.; Sharma, H.; Sahay, R.R.; Sharma, P.; et al. Zika virus outbreak in Rajasthan, India in 2018 was caused by a virus endemic to Asia. Infect. Genet. Evol. 2019, 69, 199–202. [Google Scholar] [CrossRef]
- Solanke, V.N.; Karmarkar, M.G.; Mehta, P.R. Early dengue diagnosis: Role of rapid NS1 antigen, NS1 early ELISA, and PCR assay. Trop. J. Med. Res. 2015, 18, 95. [Google Scholar] [CrossRef]
- Ambrose, J.H.; Sekaran, S.D.; Azizan, A. Dengue Virus NS1 Protein as a Diagnostic Marker: Commercially Available ELISA and Comparison to qRT-PCR and Serological Diagnostic Assays Currently Used by the State of Florida. J. Trop. Med. 2017, 2017, 1–6. [Google Scholar] [CrossRef]
- Hermann, L.L.; Thaisomboonsuk, B.; Poolpanichupatam, Y.; Jarman, R.G.; Kalayanarooj, S.; Nisalak, A.; Yoon, I.-K.; Fernandez, S. Evaluation of a dengue NS1 antigen detection assay sensitivity and specificity for the diagnosis of acute dengue virus infection. PLoS Negl. Trop. Dis. 2014, 8, e3193. [Google Scholar] [CrossRef] [PubMed]
- Hunsperger, E.A.; Yoksan, S.; Buchy, P.; Nguyen, V.C.; Sekaran, S.D.; Enria, D.A.; Vazquez, S.; Cartozian, E.; Pelegrino, J.L.; Artsob, H.; et al. Evaluation of commercially available diagnostic tests for the detection of dengue virus NS1 antigen and anti-dengue virus IgM antibody. PLoS Negl. Trop. Dis. 2014, 8, 3171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryati, A.; Trimarsanto, H.; Yohan, B.; Wardhani, P.; Fahri, S.; Sasmono, R.T. Performance of commercial dengue NS1 ELISA and molecular analysis of NS1 gene of dengue viruses obtained during surveillance in Indonesia. BMC Infect. Dis. 2013, 13, 611. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.D.R.Q.; Nogueira, R.M.R.; Schatzmayr, H.G.; dos Santos, F.B. Comparison of three commercially available dengue NS1 antigen capture assays for acute diagnosis of dengue in Brazil. PLoS Negl. Trop. Dis. 2010, 4, e738. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Jaenisch, T.; Gaczkowski, R.; Ty Hang, V.T.; Sekaran, S.D.; Kroeger, A.; Vazquez, S.; Ruiz, D.; Martinez, E.; Mercado, J.C.; et al. Multi-country evaluation of the sensitivity and specificity of two commercially-available NS1 ELISA assays for dengue diagnosis. PLoS Negl. Trop. Dis. 2010, 4, e811. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.; Wang, X.; Yuan, Q.; Zheng, Y.; Zhang, H.; Wu, Y.; Yang, J. Evaluation of the diagnostic accuracy of nonstructural pro-tein 1 Ag-based tests for dengue virus in Asian population: A meta-analysis. BMC Infect. Dis. 2015, 15, 360. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, V.G.; Marques-Silva, A.C.; Moreli, M.L. A meta-analysis of the diagnostic accuracy of two commercial NS1 antigen ELISA tests for early dengue virus detection. PLoS ONE 2014, 9, e94655. [Google Scholar] [CrossRef] [Green Version]
- Sea, V.R.; Cruz, A.C.; Gurgel, R.Q.; Nunes, B.T.; Silva, E.V.; Dolabella, S.S.; dos Santos, R.L. Underreporting of dengue-4 in Brazil due to low sensitivity of the NS1 Ag test in routine control programs. PLoS ONE 2013, 8, e64056. [Google Scholar] [CrossRef]
- Wang, W.K.; Sung, T.L.; Tsai, Y.C.; Kao, C.L.; Chang, S.M.; King, C.C. Detection of dengue virus replication in peripheral blood mononuclear cells from dengue virus type 2-infected patients by a reverse transcription-real-time PCR assay. J. Clin. Microbiol. 2002, 40, 4472–4478. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukla, S.; Mondal, P.; Biswas, S.; Ghosh, S. A Rapid and Easy-to-Perform Method of Nucleic-Acid Based Dengue Virus Diagnosis Using Fluorescence-Based Molecular Beacons. Biosensors 2021, 11, 479. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120479
Sukla S, Mondal P, Biswas S, Ghosh S. A Rapid and Easy-to-Perform Method of Nucleic-Acid Based Dengue Virus Diagnosis Using Fluorescence-Based Molecular Beacons. Biosensors. 2021; 11(12):479. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120479
Chicago/Turabian StyleSukla, Soumi, Prasenjit Mondal, Subhajit Biswas, and Surajit Ghosh. 2021. "A Rapid and Easy-to-Perform Method of Nucleic-Acid Based Dengue Virus Diagnosis Using Fluorescence-Based Molecular Beacons" Biosensors 11, no. 12: 479. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120479