Development of Multiplex PCR Coupled DNA Chip Technology for Assessment of Endogenous and Exogenous Allergens in GM Soybean


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction
2.3. Bioinformatic Analysis, Design of Oligonucleotide Primers and Probes
2.4. PCR Analysis
2.5. Asymmetric Multiplex PCR and Labelling
2.6. Biochip Preparation and Analysis
3. Results
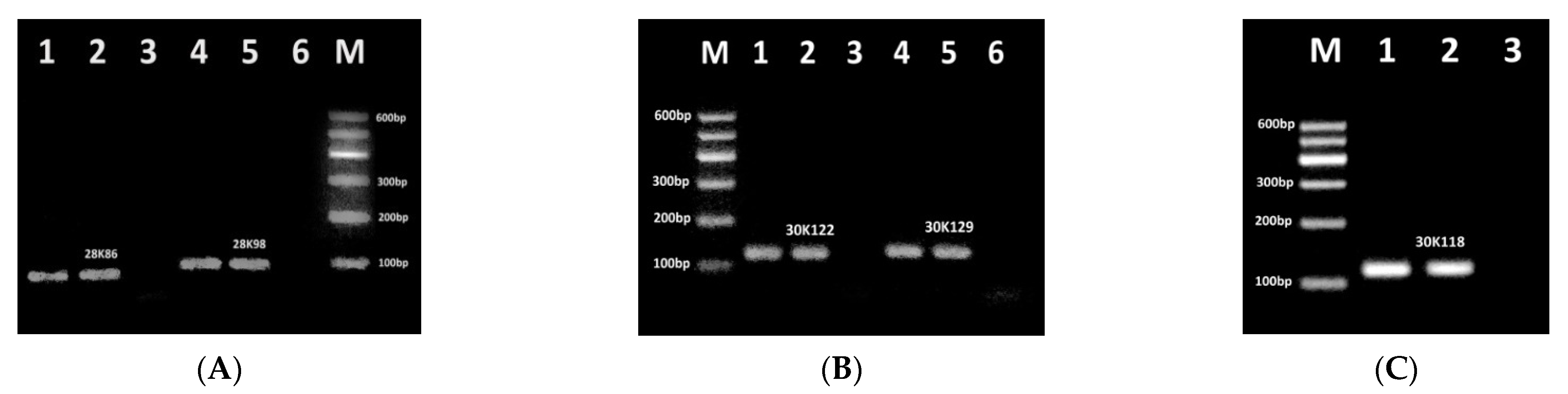
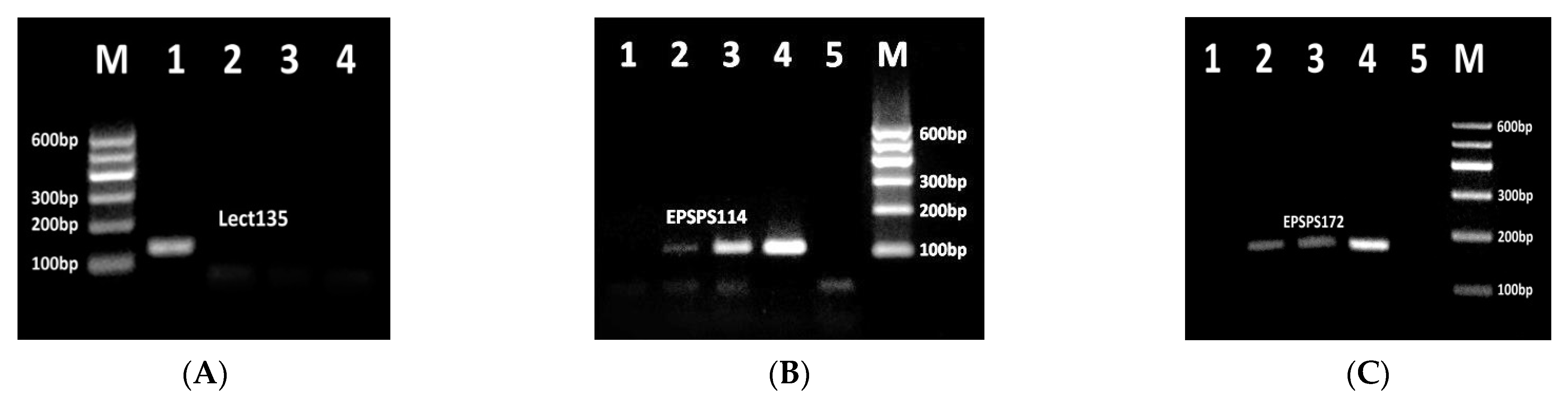
3.1. Identification of Effective PCR Primers for Detection of RRS Allergens
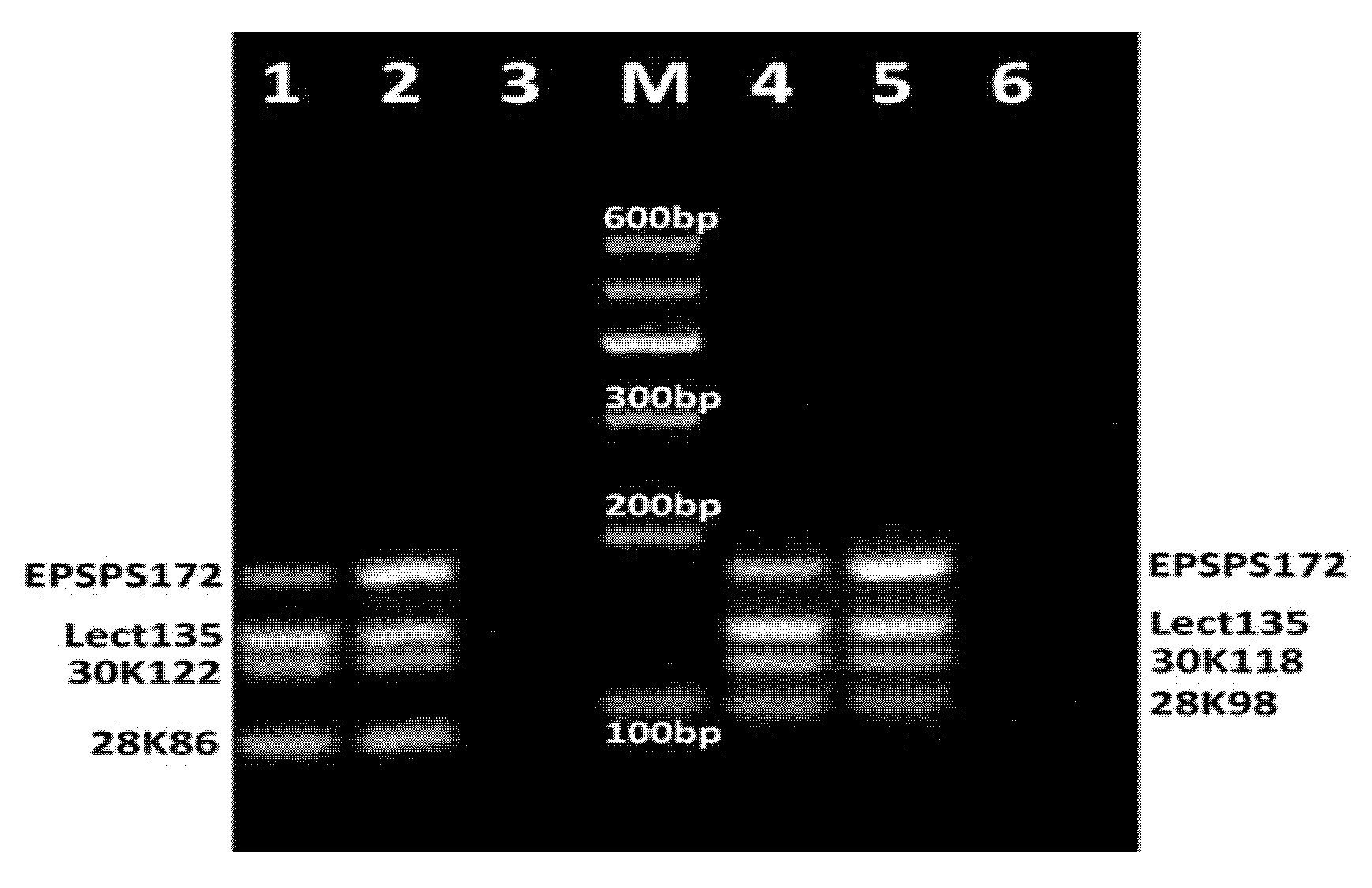
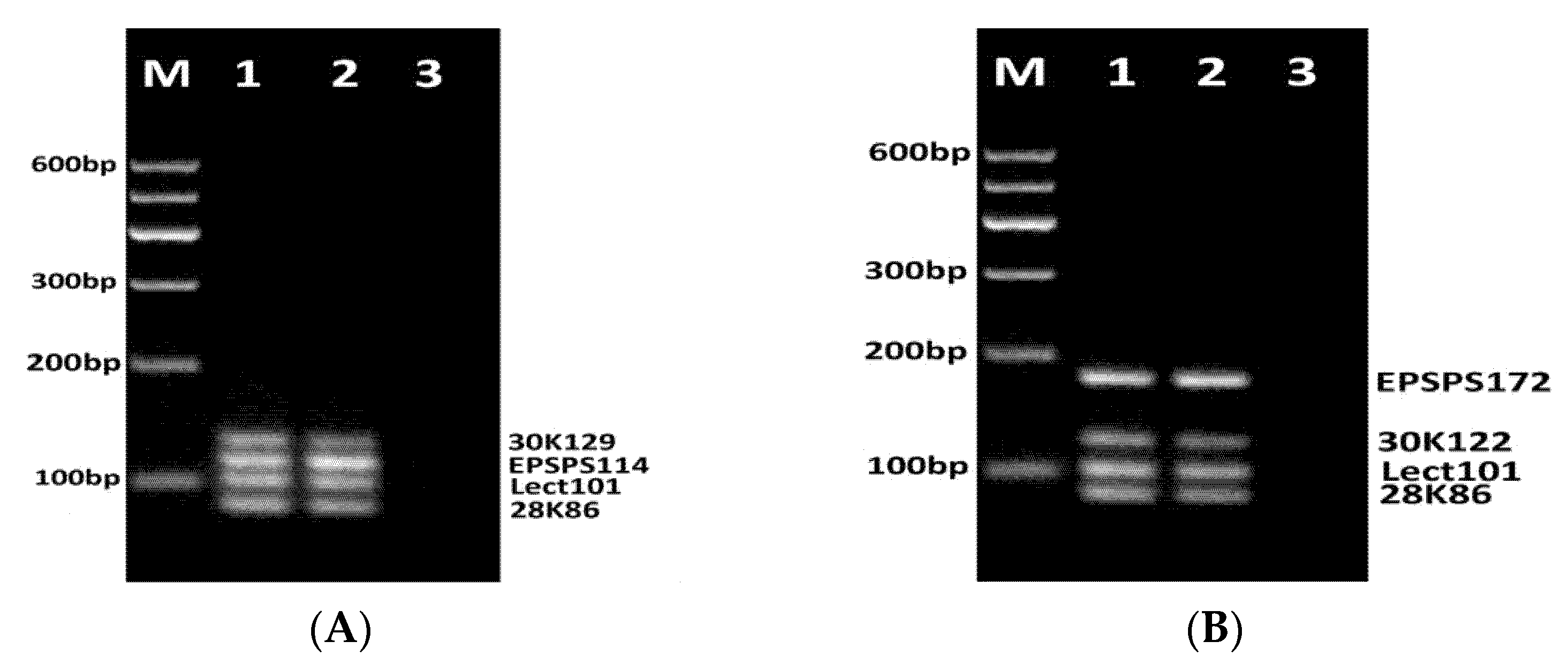
3.2. Development of Fourplex PCR
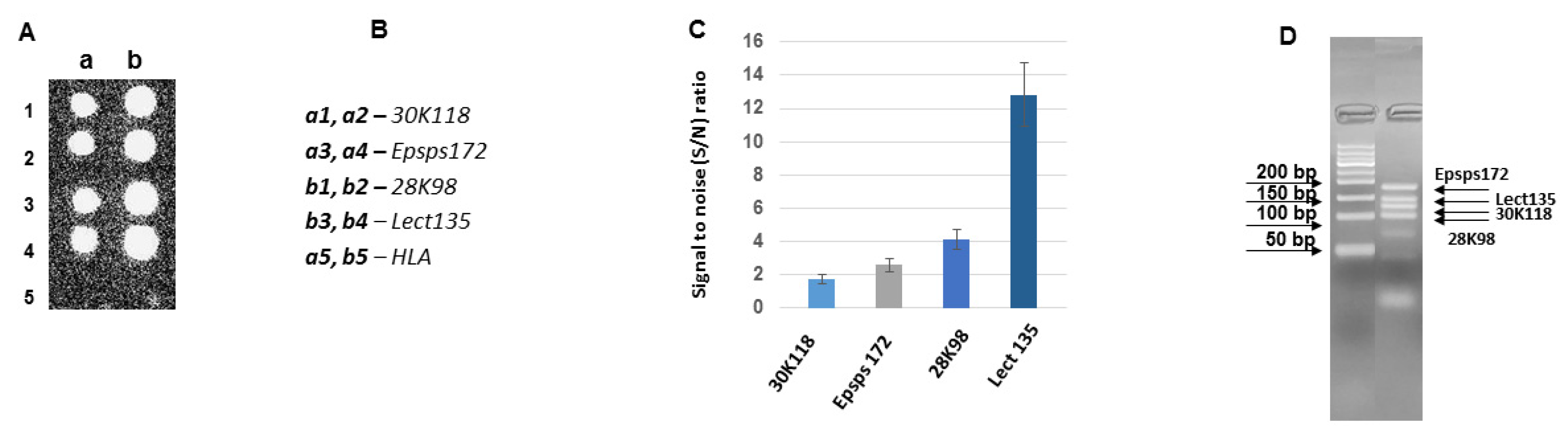
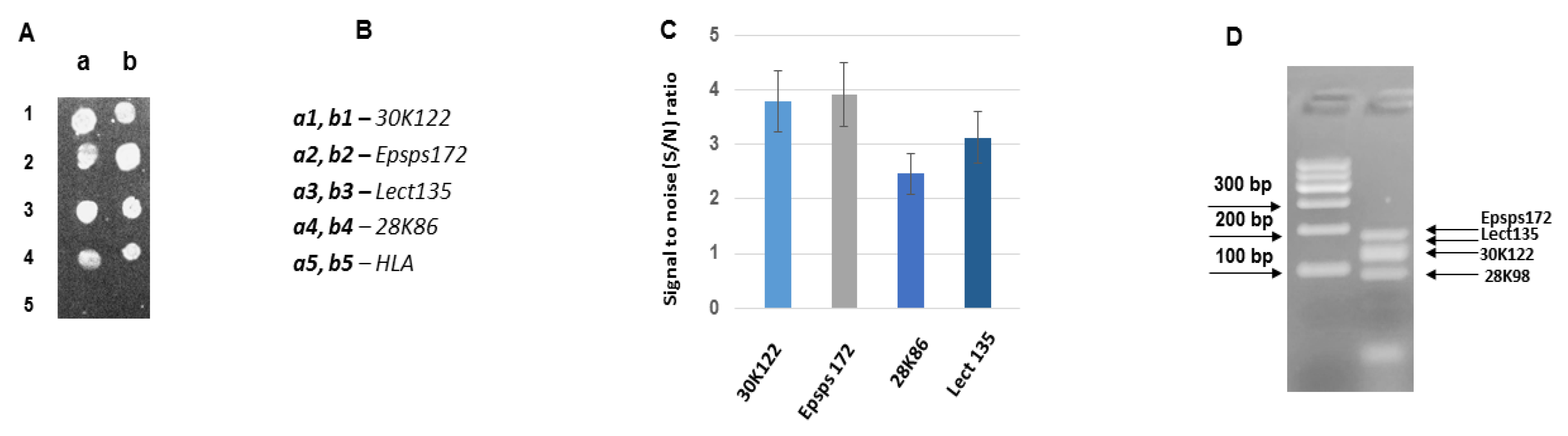
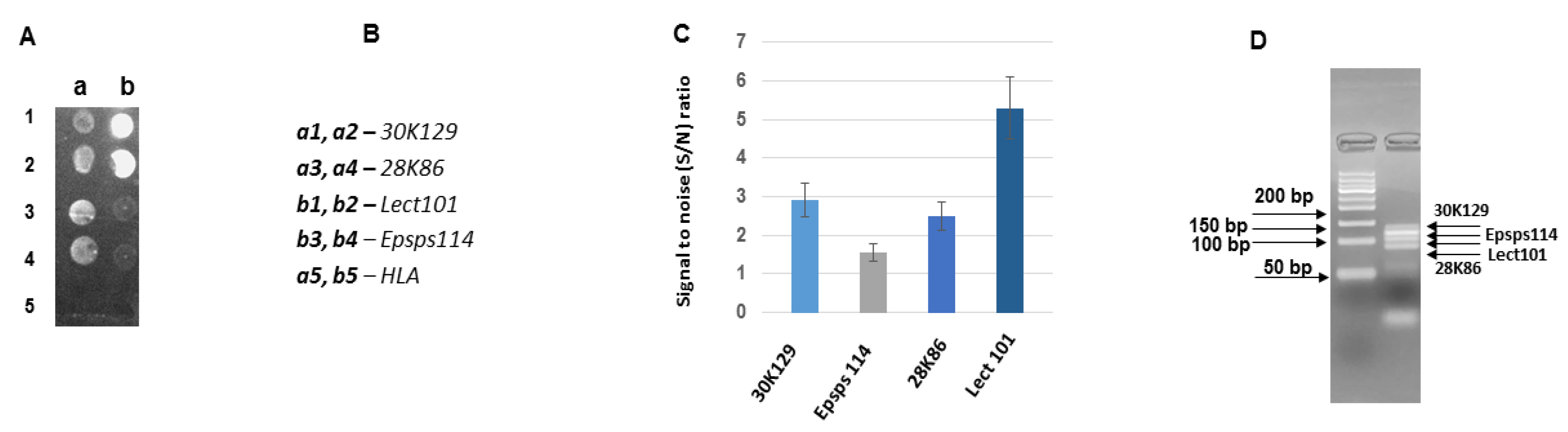
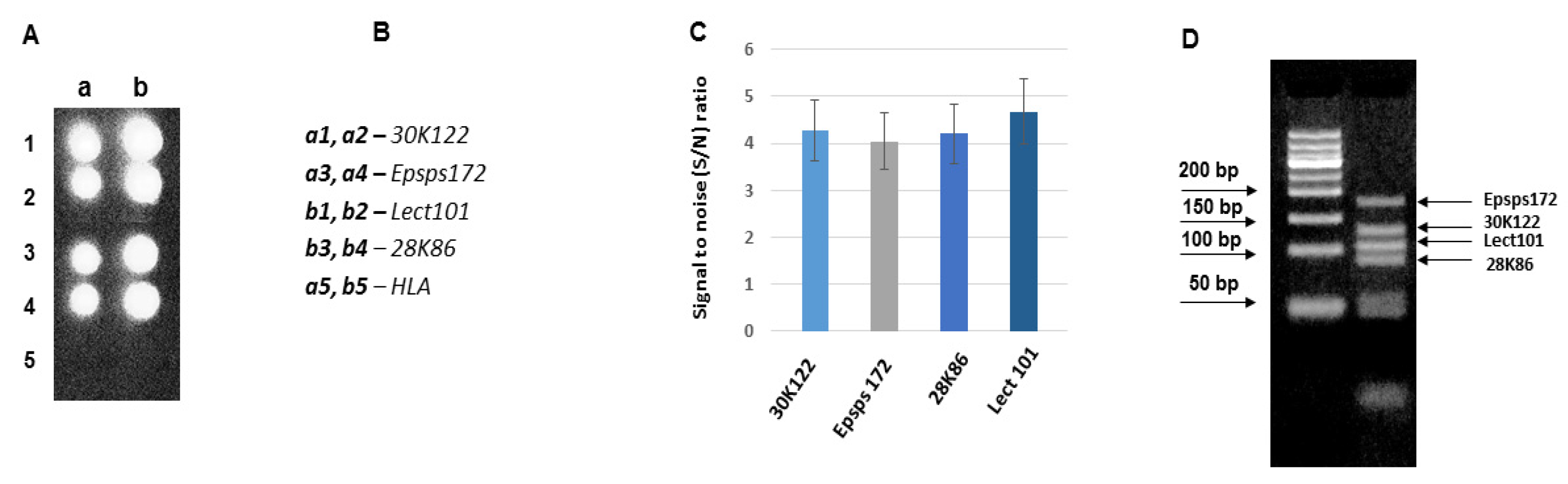
3.3. Development of DNA Chips for GM Soya Allergens
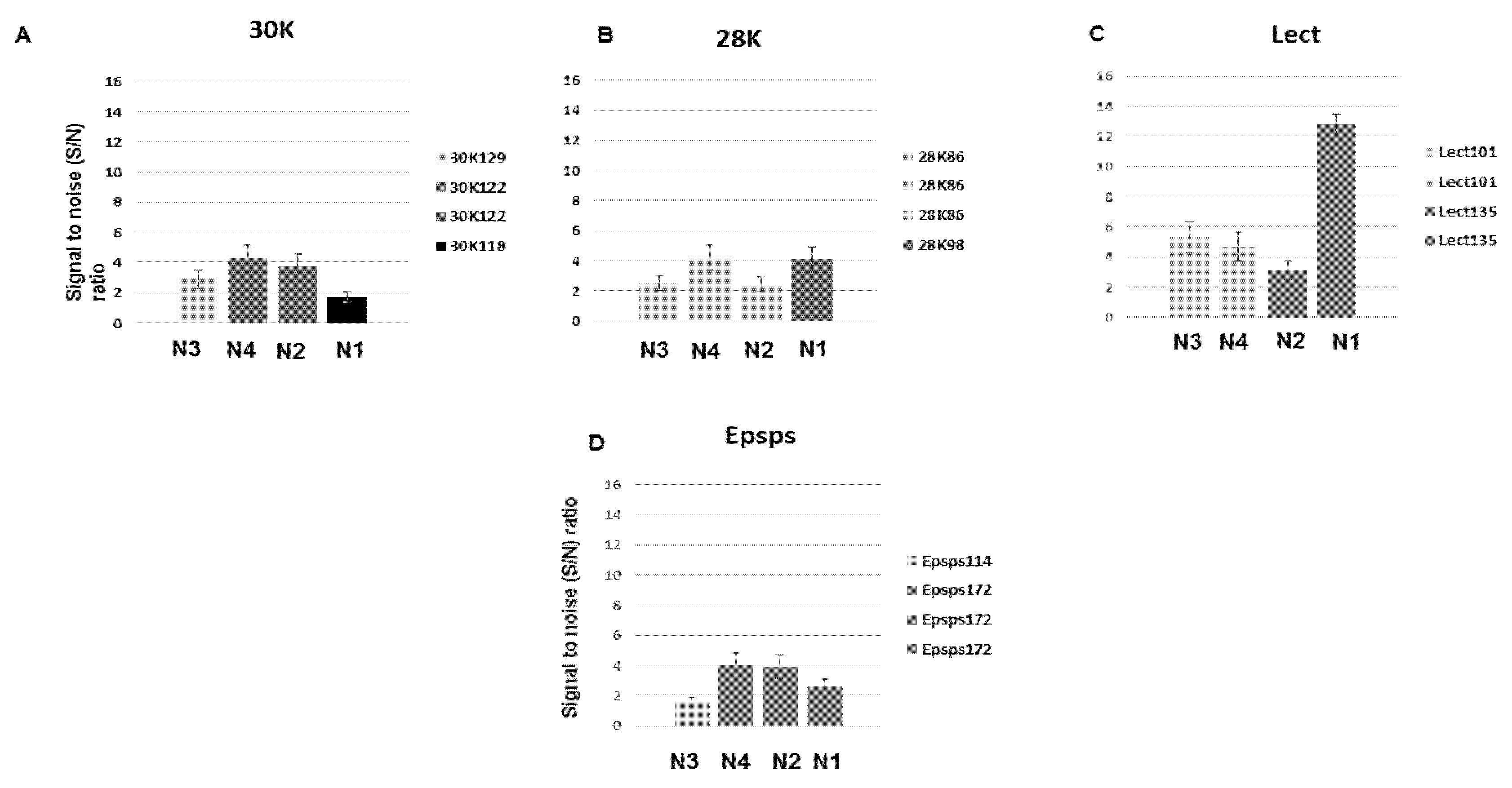
3.4. Analysis of the Developed DNA Chips
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzo, G.; Baroni, L. Soy, Soy Foods and Their Role in Vegetarian Diets. Nutrients 2018, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Seth, D.; Poowutikul, P.; Pansare, M.; Kamat, D. Food Allergy: A Review. Pediatr. Ann. 2020, 49, e50–e58. [Google Scholar] [CrossRef]
- FAO/WHO and A. C. Codex, General Standard for the Labelling of Prepackaged Foods. Codex Standard 1-1985. 1985. Available online: http://www.fao.org/3/y2770e/y2770e02.htm#fn1 (accessed on 7 September 2021).
- Gendel, S.M. Comparison of international food allergen labeling regulations. Regul. Toxicol. Pharmacol. 2012, 63, 279–285. [Google Scholar] [CrossRef] [PubMed]
- ISAAA. Global Status of Commercalized Biotech/GM Crops: 2019. ISAAA Brief No. 55; ISAAA: Ithaca, NY, USA, 2019; Available online: https://www.isaaa.org/resources/publications/briefs/55/ (accessed on 7 September 2021).
- Zhang, C.; Wohlhueter, R.; Zhang, H. Genetically modified foods: A critical review of their promise and problems. Food Sci. Hum. Wellness 2016, 5, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Codex Alimentarius Commission (Codex). Alinorm 03/34: Joint FAO/WHO Food Standard Program, Codex Alimentarius Commission, 25th Session, Rome, Italy, 30 June–5 July 2003. Appendix III, Guideline for the Conduct of Food Safety Assessment of Foods Derived from Recombinant- DNA Plants; Appendix IV, Annex on Assessment of Possible Allergenicity. 2003, pp. 47–60. Available online: http://www.fao.org/fileadmin/user_upload/gmfp/resources/al03_34e.pdf (accessed on 7 September 2021).
- EFSA Panel on Genetically Modified Organisms (GMO). Guidance on allergenicity assessment of genetically modified plants. EFSA J. 2017, 15, e04862. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.M.; Khuda, S.E.; Parker, C.H.; Eischeid, A.C.; Pereira, M. Detection of Allergen Markers in Food: Analytical Methods. Food Drug Adm. Pap. 2017, 6. Available online: http://digitalcommons.unl.edu/usfda/6 (accessed on 9 September 2021).
- Tuzimski, T.; Petruczynik, A. Review of New Trends in the Analysis of Allergenic Residues in Foods and Cosmetic Products. J. AOAC Int. 2020, 103, 997–1028. [Google Scholar] [CrossRef]
- Benedé, S.; Lozano-Ojalvo, D.; Cristobal, S.; Costa, J.; D’Auria, E.; Velickovic, T.C.; Garrido-Arandia, M.; Karakaya, S.; Mafra, I.; Mazzucchelli, G.; et al. New applications of advanced instrumental techniques for the characterization of food allergenic proteins. Crit. Rev. Food Sci. Nutr. 2021, 1–17. [Google Scholar] [CrossRef]
- Allgöwer, S.M.; Hartmann, C.A.; Holzhauser, T. The Development of Highly Specific and Sensitive Primers for the Detection of Potentially Allergenic Soybean (Glycine max) Using Loop-Mediated Isothermal Amplification Combined with Lateral Flow Dipstick (LAMP-LFD). Foods 2020, 9, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Samoto, M.; Takahashi, K. Soybean allergens and hypoallergenic soybean products. J. Nutr. Sci. Vitaminol. 2000, 46, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Holzhauser, T.; Wackermann, O.; Ballmer-Weber, B.K.; Bindslev-Jensen, C.; Scibilia, J.; Perono-Garoffo, L.; Utsumi, S.; Poulsen, L.K.; Vieths, S. Soybean (Glycine max) allergy in Europe: Gly m 5 (b-conglycinin) and Gly m 6 (glycinin) are potential diagnostic markers for severe allergic reactions to soy. J. Allergy Clin. Immunol. 2009, 123, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hei, W.; Li, Z.; Ma, X.; He, P. Determination of beta-conglycinin in soybean and soybean products using a sandwich enzyme-linked immunosorbent assay. Anal. Chim. Acta 2012, 734, 62–68. [Google Scholar] [CrossRef]
- Chen, J.; Wang, J.; Song, P.; Ma, X. Determination of glycinin in soybean and soybean products using a sandwich enzyme-linked immunosorbent assay. Food Chem. 2014, 162, 27–33. [Google Scholar] [CrossRef]
- Liu, B.; Teng, D.; Wang, X.M.; Wang, J.H. Detection of the soybean allergenic protein Gly m Bd 28K by an indirect enzyme-linked immunosorbent assay. J. Agric. Food Chem. 2013, 61, 822–828. [Google Scholar] [CrossRef]
- Morishita, N.; Kamjya, K.; Matsumoto, T.; Sakai, S.; Teshima, R.; Urisu, A.; Moriyama, T.; Ogawa, T.; Akiyama, H.; Morimatsu, F. Reliable enzymelinked immunosorbent assay for the determination of soybean proteins in processed foods. J. Agric. Food Chem. 2008, 56, 6818–6824. [Google Scholar] [CrossRef]
- Wang, B.P.; Li, Z.X.; Pawar, R.; Wang, X.F.; Lin, H. PCR-Based Detection of Traces of Potentially Allergenic Soybean (Glycine max) in Food Matrices. Adv. Mater. Res. 2011, 343–344, 618–624. [Google Scholar] [CrossRef]
- Torp, A.M.; Olesen, A.; Sten, E.; Stahl Skov, P.; Bindslev-Jensen, U.; Poulsen, L.K.; Bindslev-Jensen, C.; Andersen, S.B. Specific, semi-quantitative detection of the soybean allergen Gly m Bd 30K DNA by PCR. Food Control 2006, 17, 30–36. [Google Scholar] [CrossRef]
- Espiñeira, M.; Herrero, B.; Vieites, J.M.; Santaclara, F.J. Validation of end-point and real-time PCR methods for the rapid detection of soy allergen in processed products. Food Addit. Contam. 2010, 27, 426–432. [Google Scholar] [CrossRef]
- Pedersen, M.H.; Holzhauser, T.; Bisson, C.; Conti, A.; Jensen, L.B.; Skov, P.S.; Bindslev-Jensen, C.; Brinch, D.T.; Poulsen, L.K. Soybean allergen detection methods—A comparison study. Mol. Nutr. Food Res. 2008, 52, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.E.; Vicini, J.L.; Gleen, K.C.; Fleischer, D.M.; Greenhawt, M.J. The allergenicity of genetically modified foods from genetically engineered crops A narrative and systematic review. Ann. Allergy Asthma Immunol. 2017, 119, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, R.E.; Panda, R.; Ariyarathna, H. Evaluation of endogenous allergens for the safety evaluation of genetically engineered food crops: Review of potential risks, test methods, examples and relevance. J. Agric. Food Chem. 2013, 61, 8317–8332. [Google Scholar] [CrossRef]
- Ladics, G.S. Assessment of the potential allergenicity of genetically-engineered food crops. J. Immunotoxicol. 2019, 16, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, A.; Matsushita, K.; Fukuzumi, A.; Tokumasu, N.; Yano, E.; Zaima, N.; Moriyama, T. Comparison of Various Soybean Allergen Levels in Genetically and Non-Genetically Modified Soybeans. Foods 2020, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Tortajada-Genaro, L.A.; Santiago-Felipe, S.; Morais, S.; Gabaldón, J.A.; Puchades, R.; Maquieira, A. Multiplex DNA Detection of Food Allergens on a Digital Versatile Disk. J. Agric. Food Chem. 2012, 60, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Weng, X.; Gaur, G.; Neethirajan, S. Rapid Detection of Food Allergens by Microfluidics ELISA-Based Optical Sensor. Biosensors 2016, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashley, J.; D’Aurelio, R.; Piekarska, M.; Temblay, J.; Pleasants, M.; Trinh, L.; Rodgers, T.L.; Tothill, I.E. Development of a β-Lactoglobulin Sensor Based on SPR for Milk Allergens Detection. Biosensors 2018, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Aquino, A.; Conte-Junior, C.A. A Systematic Review of Food Allergy: Nanobiosensor and Food Allergen Detection. Biosensors 2020, 10, 194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhao, Y.; Xu, Q.; Chen, Q. Development of a triplex real-time PCR for simultaneous detection of allergenic ingredients in processed food. Czech J. Food Sci. 2018, 36, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.C.; Oman, T.J.; Wang, X.; Shan, G.; Schafer, B.; Herman, R.A.; Tobias, R.; Shippar, J.; Malayappan, B.; Sheng, L.; et al. Development, Validation, and Inter-laboratory Evaluation of a Quantitative Multiplexing Method to assess levels of Ten Endogenous Allergens in Soybean Seed and its Application to Field Trials Spanning Three Growing Seasons. J. Agric. Food Chem. 2017, 65, 5531–5554. [Google Scholar] [CrossRef]
- Datukishvili, N.; Kutateladze, T.; Gabriadze, I.; Bitskinashvili, K.; Vishnepolsky, B. New multiplex PCR methods for rapid screening of genetically modified organisms in foods. Front. Microbiol. 2015, 6, 757. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishnepolsky, B.; Pirtskhalava, M. Align_MTX—An optimal pairwise textual sequence alignment program, adapted for using in sequence-structure alignment. Comput. Biol. Chem. 2009, 33, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Lee, D.; Schulman, A.H. Java web tools for PCR, in silico PCR, and oligonucleotide assembly and analysis. Genomics 2011, 98, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Kaplinski, L.; Andreson, R.; Puurand, T.; Remm, M. MultiPLX: Automatic grouping and evaluation of PCR primers. Bioinformatics 2005, 21, 1701–1702. [Google Scholar] [CrossRef] [Green Version]
- Kutateladze, T.; Gabriadze, I.; Vishnepolsky, B.; Karseladze, M.; Datukishvili, N. Development of triplex PCR for simultaneous detection of maize, wheat and soybean. Food Control 2013, 34, 698–702. [Google Scholar] [CrossRef]
- Asatiani, N.; Abuladze, M.; Kartvelishvili, T.; Osepashvili, M.; Shengelaya, A.; Daraselia, D.; Japaridze, D.; Khatisashvili, G.; Varazi, T.; Holman, H.-Y.; et al. Copper (II) ion action on soil bacteria. Water Air Soil Pollut. 2021, 232, 355. [Google Scholar] [CrossRef]
- Barre, A.; Van Damme, E.J.M.; Simplicien, M.; Benoist, H.; Rougé, P. Are Dietary Lectins Relevant Allergens in Plant Food Allergy? Foods 2020, 9, 1724. [Google Scholar] [CrossRef]
- Beier, M.; Hoheisel, J.D. Versatile derivatization of solid support media for covalent bonding on DNA-microchips. Nucleic Acids Res. 1999, 27, 1970–1977. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Jiang, S.-M.; Zhang, M.; Wang, J.; Guo, S.-J.; Li, Y.; Jiang, H.-W.; Liu, C.-X.; Zhang, D.-B.; Yang, L.-T.; et al. MACRO: A combined microchip-PCR and microarray system for high-throughput monitoring of genetically modified organisms. Anal. Chem. 2014, 86, 1269–1276. [Google Scholar] [CrossRef]
- Nimse, S.B.; Song, K.-S.; Warkad, S.D.; Kim, T. A novel method that allows SNP discrimination with 160:1 ratio for biosensors based on DNA-DNA hybridization. Biosensors 2021, 11, 265. [Google Scholar] [CrossRef]
- Balzani, V. Dendrimers: Order, complexity, functions. Aust. J. Chem. 2011, 64, 129–130. [Google Scholar] [CrossRef]
- Lee, C.; MacKay, J.; Fréchet, J.; Szoka, F.C. Designing dendrimers for biological applications. Nat. Biotechnol. 2005, 23, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Peytavi, R.; Tang, L.-Y.; Raymond, F.R.; Boissinot, K.; Bissonnette, L.; Boissinot, M.; Picard, F.J.; Huletsky, A.; Ouellette, M.; Bergeron, M.G. Correlation between microarray DNA hybridization efficiency and the position of short capture probe on the target nucleic acid. BioTechniques 2005, 39, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Qamhieh, K.; Pettitt, B.M. Controlling microarray DNA hybridization efficiency by probe-surface distance and external surface electrostatics. In AIP Conference Proceedings, Proceedings of the 4th International Congress in Advances in Applied Physics and Materials Science (APMAS), Fethiye, Turkey, April 2014; AIP Publishing LLC: Melville, NY, USA, 2015; Volume 1653, p. 020090. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Probe | Sequence 5′→3 | 5′- Label | Target Gene | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| 28K86f 28K86r 28K86 probe | ACTGCATGGAGGCGAGTATC GGACATGTTTGCATACCGCT CCACTCAGCGAACCGGATATTGGA | CY3 AC6 | Gly m Bd 28K | 86 | This study |
| 28K98f 28K98r 28K98 probe | GGAGGAGAAGCAAACAAGTAGG GGGAACCAGCAGTGTCTTT ATG GAG GAA GCT CTT GGA AAC CGT | CY3 AC6 | Gly m Bd 28K | 98 | This study |
| 30K129f 30K129r 30K129 probe | GCTACGAGGGAACTTCTTCAGT AGGAAACCCATAACTTGGTGGA ACGACATGCTACAAGTGAAGTGACCA | CY3 AC6 | Gly m Bd 30K | 129 | This study |
| 30K118f 30K118r 30K118 probe | GGACCTTGACCTAACCAAGTT CTCAAGTCTCTTTGCCTCTTCT ATGGAAGAGTGAGCATGGACGTGT | CY3 AC6 | Gly m Bd 30K | 118 | This study |
| 30K122f 30K122r 30K122 probe | CAGGAGACCTTGTTAGCCTTT GGCAATCCCACCATGTTCTA AGACTGTGTGGAAGAAAGCGAAGGT | CY3 AC6 | Gly m Bd 30K | 122 | This study |
| Lect101f Lect101r Lect101 probe | ACGGCACCCCAAAACCCTCG GGAAGCGGCGAAGCTGGCAA CCGGTAGCGTTGCCAGCTTC | CY3 AC6 | lectin | 101 | [38] |
| lect135f lect135r lect135 probe | TCACAGAGAACCAGCAATATCC AGACCAAGAAAGCACGTCAT AGACCAAGAAAGCACGTCAT | CY3 AC6 | lectin | 135 | This study |
| epsps114f epsps114r epsps114 probe | TACGATTTCGACAGCACCTTC GTCACCGTCTTCCGATTTCA TTGAACCCGCTGCGCGAAATG | CY3 AC6 | epsps | 114 | This study |
| epsps172f epsps172r epsps172 probe | CGCTCGATTTCGGCAATG TCACCGTCTTCCGATTTCAC AAGGTGCTGTCGAAATCGTAGACCC | CY3 AC6 | epsps | 172 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutateladze, T.; Bitskinashvili, K.; Sapojnikova, N.; Kartvelishvili, T.; Asatiani, N.; Vishnepolsky, B.; Datukishvili, N. Development of Multiplex PCR Coupled DNA Chip Technology for Assessment of Endogenous and Exogenous Allergens in GM Soybean. Biosensors 2021, 11, 481. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120481
Kutateladze T, Bitskinashvili K, Sapojnikova N, Kartvelishvili T, Asatiani N, Vishnepolsky B, Datukishvili N. Development of Multiplex PCR Coupled DNA Chip Technology for Assessment of Endogenous and Exogenous Allergens in GM Soybean. Biosensors. 2021; 11(12):481. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120481
Chicago/Turabian StyleKutateladze, Tamara, Kakha Bitskinashvili, Nelly Sapojnikova, Tamar Kartvelishvili, Nino Asatiani, Boris Vishnepolsky, and Nelly Datukishvili. 2021. "Development of Multiplex PCR Coupled DNA Chip Technology for Assessment of Endogenous and Exogenous Allergens in GM Soybean" Biosensors 11, no. 12: 481. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11120481