Bioluminescence Color-Tuning Firefly Luciferases: Engineering and Prospects for Real-Time Intracellular pH Imaging and Heavy Metal Biosensing

Abstract
:1. Introduction
2. An Overview of Current Luminescent Intracellular Sensors for pH and Heavy Metals
3. The Firefly Luciferases pH Sensitivity
4. Identification of the pH-Sensor and Metal Binding Site of Firefly Luciferases
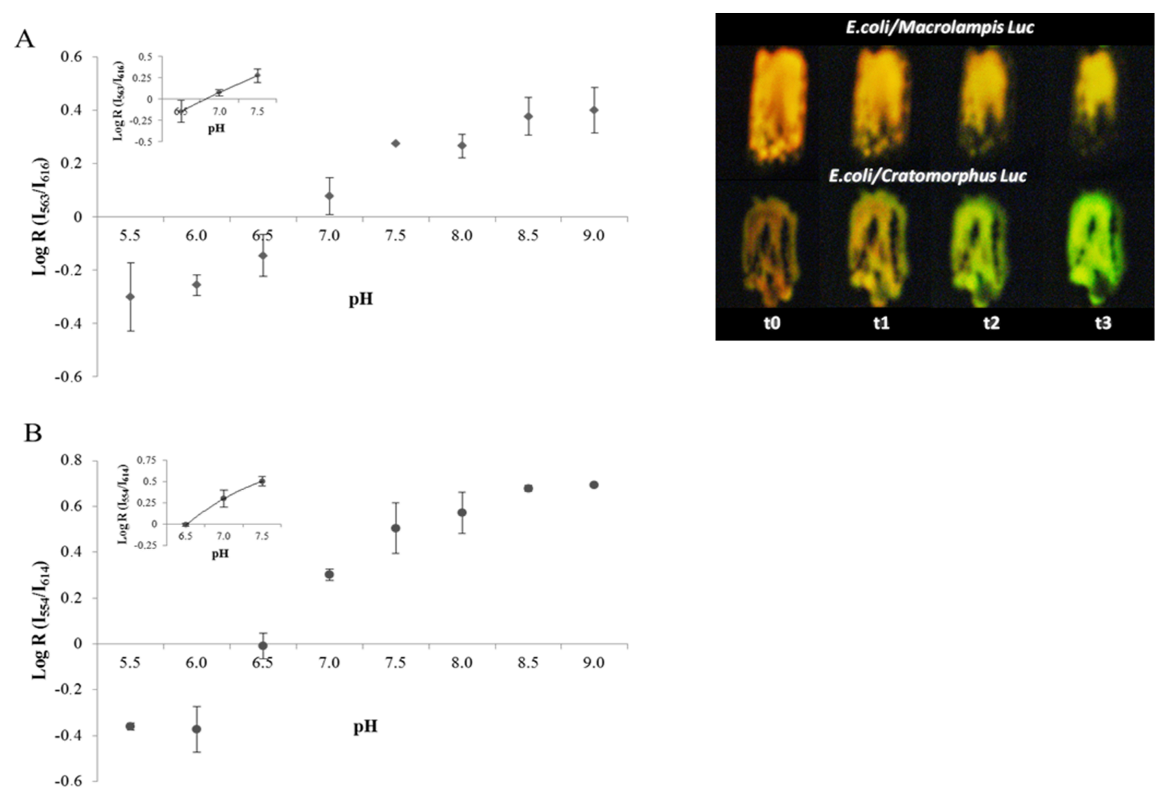
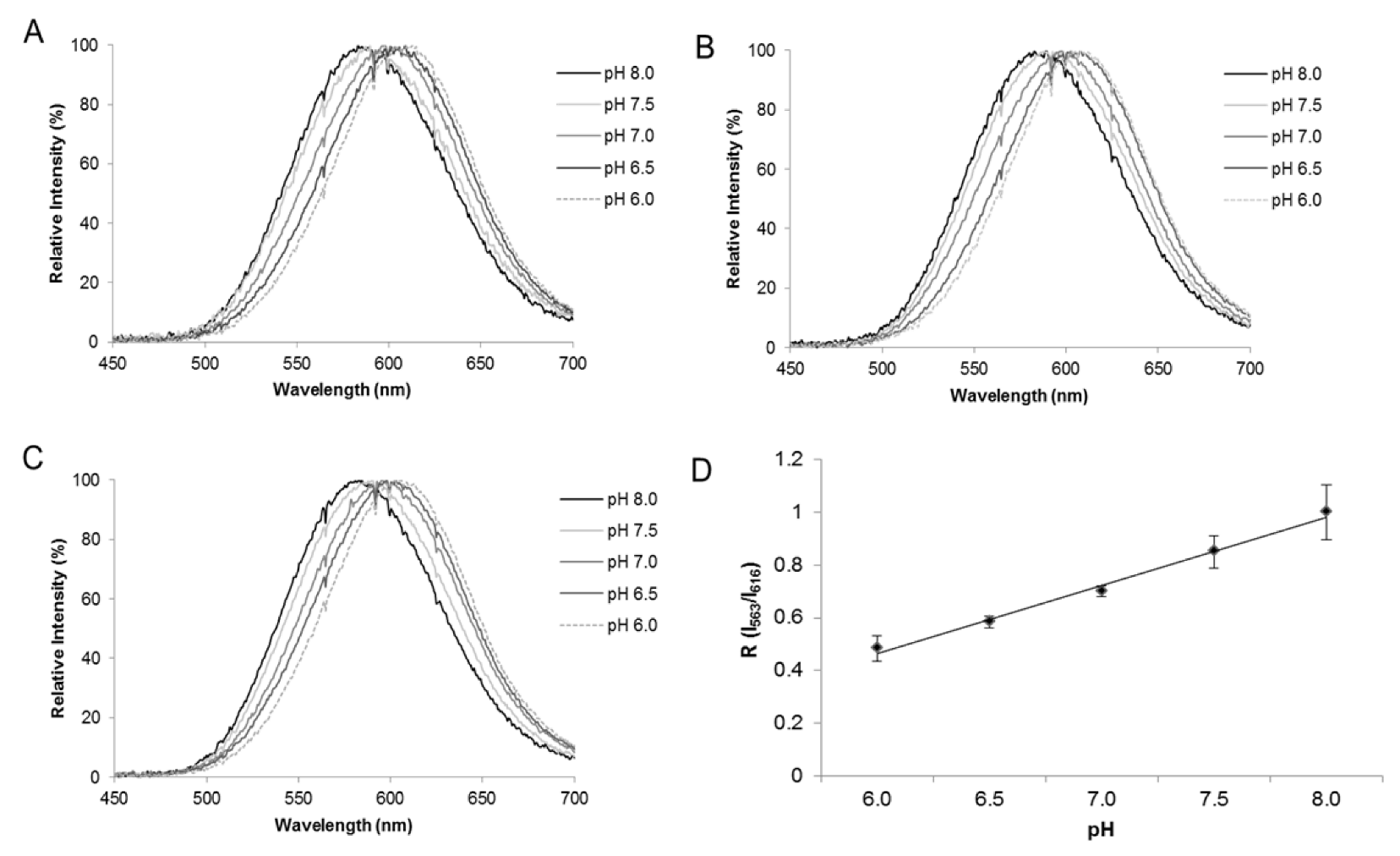
5. Use of Firefly Luciferases as Color-Tuning Indicators of Intracellular pH
6. Ratiometric Analysis of Temperature
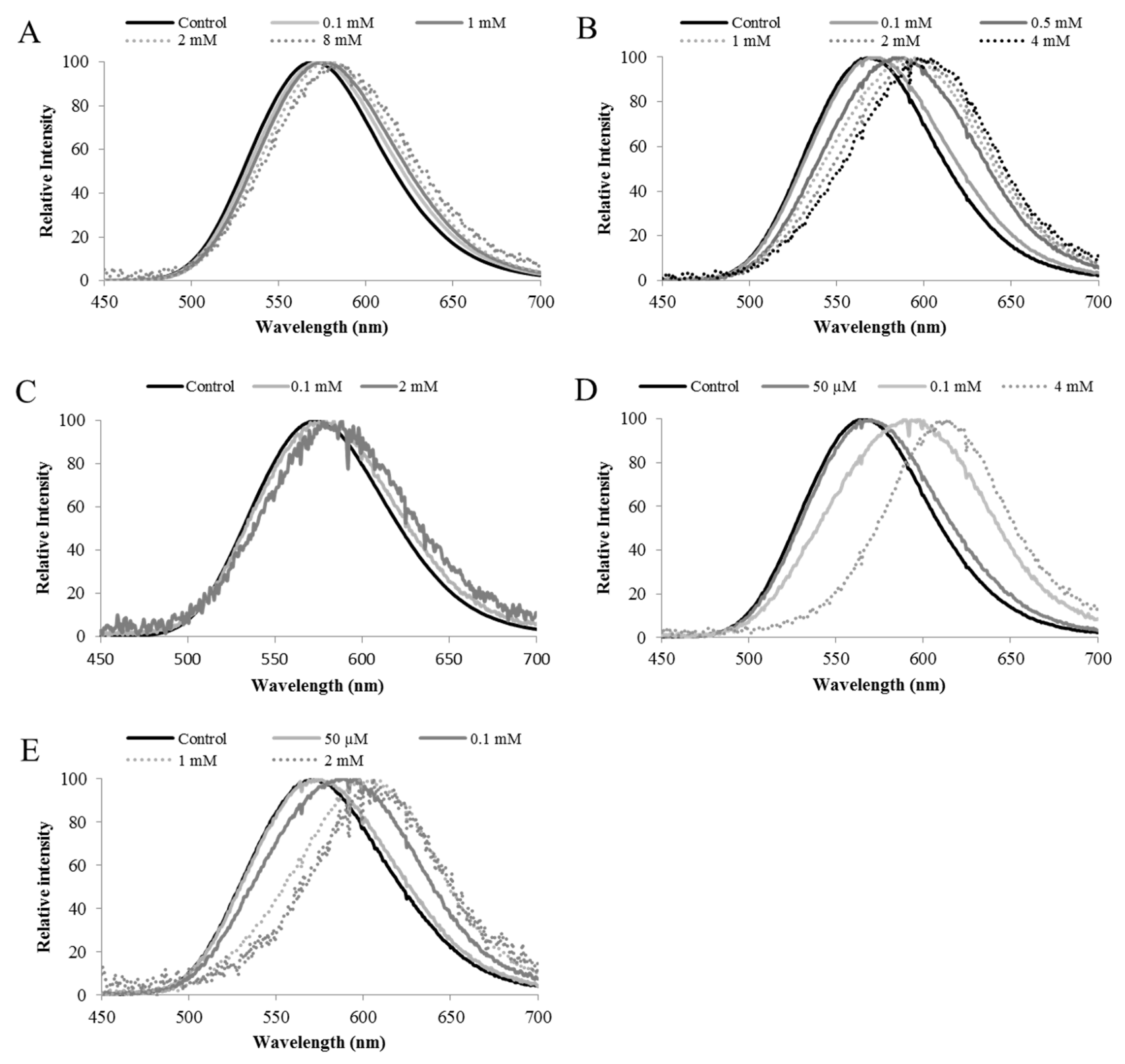
7. Use of Firefly Luciferases as Color-Tuning Sensors for Heavy Metals
8. Selection of Metal-Sensitive Luciferases and Engineering of the Metal-Binding Site
9. Smartphone Detection of Cadmium Contamination in Water
10. Is it Possible to Report in Two Dimensions?
11. Drawbacks and Perspectives: Comparison with Other Luminescent Biosensors
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Viviani, V.R.; Ohmiya, Y. Beetle luciferases: Colorful lights on biological processes and diseases. In Photoproteins in Bioanalysis; Wiley-VCH: Weinheim, Germany, 2006; pp. 49–63. [Google Scholar] [CrossRef]
- Roda, A.; Mezzanotte, L.; Aldini, R.; Michelini, E.; Cevenini, L. A new gastric-emptying mouse model based on in vivo non-invasive bioluminescence imaging. Neurogastroenterol. Motil. 2010, 22, 1117–1288. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.J.; Anderson, J.C. Applications of bioluminescence in biotechnology and beyond. Chem. Soc. Rev. 2021, 50, 5668–5705. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wei, L.; Niu, P. The novel coronavirus outbreak in Wuhan, China. Glob. Health Res. Policy 2020, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Niu, J.; Wang, C.; Huang, B.; Wang, W.; Zhu, N.; Deng, Y.; Wang, H.; Ye, F.; Cen, S.; et al. High-throughput screening and identification of potent broad-spectrum inhibitors of coronaviruses. J. Virol. 2019, 93, e00023-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirasoli, M.; Michelini, E. Analytical bioluminescence and chemiluminescence. Anal. Bioanal. Chem. 2014, 406, 5529–5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nat. Rev. Microbiol. 2011, 9, 330–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarante-Mendes, G.P. Apoptose: Programa molecular de morte celular. Einstein 2003, 1, 15–18. [Google Scholar]
- Maret, W. Analyzing free zinc (II) ion concentrations in cell biology with fluorescent chelating molecules. Metallomics 2015, 7, 202–211. [Google Scholar] [CrossRef]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef]
- Howard, W.; Leonard, B.; Moody, W.; Kochhar, T.S. Induction of chromosome changes by metal compounds in cultured CHO cells. Toxicol. Lett. 1991, 56, 179–186. [Google Scholar] [CrossRef]
- Breeuwer, P.; Drocourt, J.L.; Rombouts, F.M.; Abee, T. A novel method for continuous determination of the intracellular pH in bacteria with the internally conjugated fluorescent probe 5 (and 6)-carboxyfluoresceinsuccinimidyl ester. Appl. Environ. Microbiol. 1996, 62, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneen, M.; Farinas, J.; Li, Y.; Verkman, A.S. Green fluorescent protein as a noninvasive intracellular pH indicator. Biophys. J. 1998, 74, 1591–1599. [Google Scholar] [CrossRef] [Green Version]
- Bizzarri, R.; Arcangeli, C.; Arosio, D.; Ricci, F.; Faraci, P.; Cardarelli, F.; Beltram, F. Development of a novel GFP-based ratiometric excitation and emission pH indicator for intracellular studies. Biophys. J. 2006, 90, 3300–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Qian, X.; Liu, F.; Zhang, R. Novel fluorescent pH sensors based on intramolecular hydrogen bonding ability of naphthalimide. Organicletters 2004, 6, 2757–2760. [Google Scholar] [CrossRef]
- Mahon, M.J. pHluorin2: An enhanced, ratiometric, pH-sensitive green florescent protein. Adv. Biosci. Biotechnol. 2011, 2, 132. [Google Scholar] [CrossRef] [Green Version]
- Valeur, B. Fluorescent Molecular Sensors of Ions and Molecules. In Molecular Fluorescence: Principles and Applications; Wiley-VCH: Weinheim, Germany, 2001; pp. 273–349. [Google Scholar] [CrossRef]
- Chen, G.; Guo, Z.; Zeng, G.; Tang, L. Fluorescent and colorimetric sensors for environmental mercury detection. Analyst 2015, 140, 5400–5443. [Google Scholar] [CrossRef]
- Kim, H.N.; Ren, W.X.; Kim, J.S.; Yoon, J. Fluorescent and colorimetric sensors for detection of lead, cadmium, and mercury ions. Chem. Soc. Rev. 2012, 41, 3210–3244. [Google Scholar] [CrossRef]
- Eltzov, E.; Marks, R.S. Whole-cell aquatic biosensors. Anal. Bioanal. Chem. 2011, 400, 895–913. [Google Scholar] [CrossRef]
- Gabriel, G.V.; Lopes, P.S.; Viviani, V.R. Suitability of Macrolampis firefly and Pyrearinus click beetle luciferases for bacterial light off toxicity biosensor. Anal. Biochem. 2014, 445, 73–79. [Google Scholar] [CrossRef]
- Selifonova, O.; Burlage, R.; Barkay, T. Bioluminescent sensors for detection of bioavailable Hg (II) in the environment. Appl. Environ. Microbiol. 1993, 59, 3083–3090. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Adil, M.; Ehtisham-ul-Haque, S.; Munir, B.; Yameen, M.; Ghaffar, A.; Shar, G.A.; Tahir, M.A.; Iqbal, M. Vibrio fischeri bioluminescence inhibition assay for ecotoxicity assessment: A review. Sci. Total Environ. 2018, 626, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Tauriainen, S.M.; Virta MP, J.; Karp, M.T. Detecting bioavailable toxic metals and metalloids from natural water samples using luminescent sensor bacteria. Water Res. 2000, 34, 2661–2666. [Google Scholar] [CrossRef]
- Seliger, H.H.; McElroy, W.D. The colors of firefly bioluminescence: Enzyme configuration and species specificity. Proc. Natl. Acad. Sci. USA 1964, 52, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viviani, V.R.; Bechara, E.J. Bioluminescence of Brazilian fireflies (Coleoptera: Lampyridae): Spectral distribution and pH effect on luciferase-elicited colors. Comparison with elaterid and phengodid luciferases. Photochem. Photobiol. 1995, 62, 490–495. [Google Scholar] [CrossRef]
- Viviani, V.R.; Arnoldi, F.G.; Brochetto-Braga, M.; Ohmiya, Y. Cloning and characterization of the cDNA for the Brazilian Cratomorphus distinctus larval firefly luciferase: Similarities with European Lampyris noctiluca and Asiatic Pyrocoelia luciferases. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Viviani, V.R.; Oehlmeyer, T.L.; Arnoldi, F.G.C.; Brochetto-Braga, M.R. A New Firefly Luciferase with Bimodal Spectrum: Identification of Structural Determinants of Spectral pH-Sensitivity in Firefly Luciferases. Photochem. Photobiol. 2005, 81, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Viviani, V.R.; Amaral, D.; Prado, R.; Arnoldi, F.G. A new blue-shifted luciferase from the Brazilian Amydetes fanestratus (Coleoptera: Lampyridae) firefly: Molecular evolution and structural/functional properties. Photochem. Photobiol. Sci. 2011, 10, 1879–1886. [Google Scholar] [CrossRef]
- Carvalho, M.C.; Tomazini, A.; Amaral, D.T.; Murakami, M.T.; Viviani, V.R. Luciferase isozymes from the Brazilian Aspisoma lineatum (Lampyridae) firefly: Origin of efficient pH-sensitive lantern luciferases from fat body pH-insensitive ancestors. Photochem. Photobiol. Sci. 2020, 19, 1750–1764. [Google Scholar] [CrossRef]
- Viviani, V.R.; Gabriel, G.V.; Bevilaqua, V.R.; Simões, A.F.; Hirano, T.; Lopes-de-Oliveira, P.S. The proton and metal binding sites responsible for the pH-dependent green-red bioluminescence color tuning in firefly luciferases. Sci. Rep. 2018, 8, 17594. [Google Scholar] [CrossRef]
- Gabriel, G.V.; Viviani, V.R. Novel application of pH-sensitive firefly luciferases as dual reporter genes for simultaneous ratiometric analysis of intracellular pH and gene expression/location. Photochem. Photobiol. Sci. 2014, 13, 1661–1670. [Google Scholar] [CrossRef]
- Gabriel, G.V.; Viviani, V.R. Engineering the metal sensitive sites in Macrolampis sp2 firefly luciferase and use as a novel bioluminescent ratiometric biosensor for heavy metals. Anal. Bioanal. Chem. 2016, 408, 8881–8893. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, G.V.; Yasuno, R.; Mitani, Y.; Ohmiya, Y.; Viviani, V.R. Novel application of Macrolampis sp2 firefly luciferase for intracellular pH-biosensing in mammalian cells. Photochem. Photobiol. Sci. 2019, 18, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Arosio, D.; Ratto, G.M. Twenty years of fluorescence imaging of intracellular chloride. Front. Cell. Neurosci. 2014, 8, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, J.; Barber, D.L.; Jacobson, M.P. Intracellular pH sensors: Design principles and functional significance. Physiology 2007, 22, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J.W.; Brul, S. Compartment-specific pH monitoring in Bacillus subtilis using fluorescent sensor proteins: A tool to analyze the antibacterial effect of weak organic acids. Front. Microbiol. 2013, 4, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llopis, J.; McCaffery, J.M.; Miyawaki, A.; Farquhar, M.G.; Tsien, R.Y. Measurement of cytosolic, mitochondrial, and Golgi pH in single living cells with green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 6803–6808. [Google Scholar] [CrossRef] [Green Version]
- Miesenböck, G.; De Angelis, D.A.; Rothman, J.E. Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature 1998, 394, 192–195. [Google Scholar] [CrossRef]
- Hao, Z.; Zhu, R.; Chen, P.R. Genetically encoded fluorescent sensors for measuring transition and heavy metals in biological systems. Curr. Opin. Chem. Biol. 2018, 43, 87–96. [Google Scholar] [CrossRef]
- Arosio, D.; Ricci, F.; Marchetti, L.; Gualdani, R.; Albertazzi, L.; Beltram, F. Simultaneous intracellular chloride and pH measurements using a GFP-based sensor. Nat. Methods 2010, 7, 516–518. [Google Scholar] [CrossRef]
- Chen, M.; Hombrebueno, J.R.; Luo, C.; Penalva, R.; Zhao, J.; Colhoun, L.; Pandi, S.P.S.; Forrester, J.V.; Xu, H. Age-and light-dependent development of localised retinal atrophy in CCL2−/− CX3CR1GFP/GFP mice. PLoS ONE 2013, 8, e61381. [Google Scholar] [CrossRef] [Green Version]
- Martynov, V.I.; Pakhomov, A.A.; Deyev, I.E.; Petrenko, A.G. Genetically encoded fluorescent indicators for live cell pH imaging. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2924–2939. [Google Scholar] [CrossRef] [PubMed]
- Benčina, M. Illumination of the spatial order of intracellular pH by genetically encoded pH-sensitive sensors. Sensors 2013, 13, 16736–16758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, X.; Tian, X.; Liu, A.; Jia, L. Carboxamidoquinoline–coumarin derivative: A ratiometric fluorescent sensor for Cu (II) in a dual fluorophore hybrid. Sens. Actuators B Chem. 2015, 218, 37–41. [Google Scholar] [CrossRef]
- Xu, W.; Ren, C.; Teoh, C.L.; Peng, J.; Gadre, S.H.; Rhee, H.W.; Lee, C.L.K.; Chang, Y.T. An artificial tongue fluorescent sensor array for identification and quantitation of various heavy metal ions. Anal. Chem. 2014, 86, 8763–8769. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Venkatesan, P.; Wu, S.P. A sensitive and selective fluorescent sensor for Zinc (II) and its application to living cell imaging. Sens. Actuators B Chem. 2014, 203, 719–725. [Google Scholar] [CrossRef]
- Hosseini, M.; Ghafarloo, A.; Ganjali, M.R.; Faridbod, F.; Norouzi, P.; Niasari, M.S. A turn-on fluorescent sensor for Zn2+ based on new Schiff’s base derivative in aqueous media. Sens. Actuators B Chem. 2014, 198, 411–415. [Google Scholar] [CrossRef]
- Tang, L.; Dai, X.; Zhong, K.; Wen, X.; Wu, D. A phenylbenzothiazole derived fluorescent sensor for Zn (II) recognition in aqueous solution through “Turn-On” excited-state intramolecular proton transfer emission. J. Fluoresc. 2014, 24, 1487–1493. [Google Scholar] [CrossRef]
- Song, E.J.; Park, G.J.; Lee, J.J.; Lee, S.; Noh, I.; Kim, Y.; Kim, S.J.; Kim, C.; Harrison, R.G. A fluorescence sensor for Zn2+ that also acts as a visible sensor for Co2+ and Cu2+. Sens. Actuators B Chem. 2015, 213, 268–275. [Google Scholar] [CrossRef]
- Tian, X.; Guo, X.; Jia, L.; Yang, R.; Cao, G.; Liu, C. A fluorescent sensor based on bicarboxamidoquinoline for highly selective relay recognition of Zn2+ and citrate with ratiometric response. Sens. Actuators B Chem. 2015, 221, 923–929. [Google Scholar] [CrossRef]
- Hosseini, M.; Khabbaz, H.; Dezfoli, A.S.; Ganjali, M.R.; Dadmehr, M. Selective recognition of Glutamate based on fluorescence enhancement of graphene quantum dot. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 136, 1962–1966. [Google Scholar] [CrossRef]
- Aydin, Z.; Wei, Y.; Guo, M. An “off–on” optical sensor for mercury ion detection in aqueous solution and living cells. Inorg. Chem. Commun. 2014, 50, 84–87. [Google Scholar] [CrossRef]
- Han, A.; Liu, X.; Prestwich, G.D.; Zang, L. Fluorescent sensor for Hg2+ detection in aqueous solution. Sens. Actuators B Chem. 2014, 198, 274–277. [Google Scholar] [CrossRef]
- Erdemir, S.; Kocyigit, O.; Malkondu, S. Detection of Hg2+ ion in aqueous media by new fluorometric and colorimetric sensor based on triazole–rhodamine. J. Photochem. Photobiol. A Chem. 2015, 309, 15–21. [Google Scholar] [CrossRef]
- Maity, S.B.; Banerjee, S.; Sunwoo, K.; Kim, J.S.; Bharadwaj, P.K. A fluorescent chemosensor for Hg2+ and Cd2+ ions in aqueous medium under physiological pH and its applications in imaging living cells. Inorg. Chem. 2015, 54, 3929–3936. [Google Scholar] [CrossRef]
- Wu, B.; Xu, L.; Wang, S.; Wang, Y.; Zhang, W. A PEGylated colorimetric and turn-on fluorescent sensor based on BODIPY for Hg (II) detection in water. Polym. Chem. 2015, 6, 4279–4289. [Google Scholar] [CrossRef]
- Hui, C.Y.; Guo, Y.; Liu, L.; Yi, J. Recent advances in bacterial biosensing and bioremediation of cadmium pollution: A mini-review. World J. Microbiol. Biotechnol. 2022, 38, 9. [Google Scholar] [CrossRef]
- Kumar, S.; Verma, N.; Singh, A.K. Development of cadmium specific recombinant biosensor and its application in milk samples. Sens. Actuators B Chem. 2017, 240, 248–254. [Google Scholar] [CrossRef]
- Chiu, T.Y.; Chen, P.H.; Chang, C.L.; Yang, D.M. Live-cell dynamic sensing of Cd2+ with a FRET-based indicator. PLoS ONE 2013, 8, e65853. [Google Scholar] [CrossRef] [Green Version]
- Taki, M.; Desaki, M.; Ojida, A.; Iyoshi, S.; Hirayama, T.; Hamachi, I.; Yamamoto, Y. Fluorescence imaging of intracellular cadmium using a dual-excitation ratiometricchemosensor. J. Am. Chem. Soc. 2008, 130, 12564–12565. [Google Scholar] [CrossRef]
- Michelini, E.; Cevenini, L.; Mezzanotte, L.; Roda, B.; Dolci, L.S.; Roda, A. Bioluminescent reporter proteins for multicolor assays. Minerva Biotecnol. 2009, 21, 87–96. [Google Scholar]
- Girotti, S.; Ferri, E.N.; Fumo, M.G.; Maiolini, E. Monitoring of environmental pollutants by bioluminescent bacteria. Anal. Chim. Acta 2008, 608, 2–29. [Google Scholar] [CrossRef]
- Su, L.; Jia, W.; Hou, C.; Lei, Y. Microbial biosensors: A review. Biosens. Bioelectron. 2011, 26, 1788–1799. [Google Scholar] [CrossRef]
- Tauriainen, S.; Karp, M.; Chang, W.; Virta, M. Recombinant luminescent bacteria for measuring bioavailable arsenite and antimonite. Appl. Environ. Microbiol. 1997, 63, 4456–4461. [Google Scholar] [CrossRef] [Green Version]
- Tauriainen, S.; Karp, M.; Chang, W.; Virta, M. Luminescent bacterial sensor for cadmium and lead. Biosens. Bioelectron. 1998, 13, 931–938. [Google Scholar] [CrossRef]
- Nakajima, Y.; Ikeda, M.; Kimura, T.; Honma, S.; Ohmiya, Y.; Honma, K.I. Bidirectional role of orphan nuclear receptor RORα in clock gene transcriptions demonstrated by a novel reporter assay system. FEBS Lett. 2004, 565, 122–126. [Google Scholar] [CrossRef]
- Hattori, M.; Ozawa, T. Split luciferase complementation for analysis of intracellular signaling. Anal. Sci. 2014, 30, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Agulhon, C.; Platel, J.C.; Kolomiets, B.; Forster, V.; Picaud, S.; Brocard, J.; Faure, P.; Brulet, P. Bioluminescent imaging of Ca2+ activity reveals spatiotemporal dynamics in glial networks of dark-adapted mouse retina. J. Physiol. 2007, 583, 945–958. [Google Scholar] [CrossRef]
- Rogers, K.L.; Stinnakre, J.; Agulhon, C.; Jublot, D.; Shorte, S.L.; Kremer, E.J.; Brûlet, P. Visualization of local Ca2+ dynamics with genetically encoded bioluminescent reporters. Eur. J. Neurosci. 2005, 21, 597–610. [Google Scholar] [CrossRef]
- Saito, K.; Chang, Y.F.; Horikawa, K.; Hatsugai, N.; Higuchi, Y.; Hashida, M.; Yoshida, Y.; Matsuda, T.; Arai, Y.; Nagai, T. Luminescent proteins for high-speed single-cell and whole-body imaging. Nat. Commun. 2012, 3, 1262. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef]
- Ghose, A.; Maltsev, O.V.; Humbert, N.; Hintermann, L.; Arntz, Y.; Naumov, P.; Mély, Y.; Didier, P. Oxyluciferin Derivatives: A Toolbox of Environment-Sensitive Fluorescence Probes for Molecular and Cellular Applications. J. Phys. Chem. B 2017, 121, 1566–1575. [Google Scholar] [CrossRef]
- Ohmiya, Y. Basic and Applied aspects of color tuning of BL systems. Jpn. J. Appl. Phys. 2005, 44, 6368–6379. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, Q.; Robertson, J.B.; Johnson, C.H. pHlash: A New Genetically Encoded and Ratiometric Luminescence Sensor of Intracellular pH. PLoS ONE 2012, 7, e43072. [Google Scholar] [CrossRef]
- Zhang, Y.; Robertson, J.B.; Xie, Q.; Johnson, C.H. Monitoring Intracellular pH Change with a Genetically Encoded and Ratiometric Luminescence Sensor in Yeast and Mammalian Cells. Methods Mol. Biol. (Clifton N.J.) 2016, 1461, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kubota, H.; Yamada, N.; Irie, T.; Akiyama, H. Quantum yields and quantitative spectra of firefly bioluminescence with various bivalent metal ions. Photochem. Photobiol. 2011, 87, 846–852. [Google Scholar] [CrossRef]
- Yang, J.; Johnson, C.H. Bioluminescent Sensors for Ca++ Flux Imaging and the Introduction of a New Intensity-Based Ca++ Sensor. Front. Bioeng. Biotechnol. 2021, 1036, 773353. [Google Scholar] [CrossRef]
- Yang, J.; Cumberbatch, D.; Centanni, S.; Shi, S.Q.; Winder, D.; Webb, D.; Johnson, C.H. Coupling optogenetic stimulation with NanoLuc-based luminescence (BRET) Ca++ sensing. Nat. Commun. 2016, 7, 13268. [Google Scholar] [CrossRef] [Green Version]
- Merkx, M.; Golynskiy, M.V.; Lindenburg, L.H.; Vinkenborg, J.L. Rational design of FRET sensor proteins based on mutually exclusive domain interactions. Biochem. Soc. Trans. 2013, 41, 1201–1205. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Niwa, K.; Yamada, N.; Enomoto, T.; Irie, T.; Kubota, H.; Ohmiya, Y.; Akiyama, H. Firefly bioluminescence quantum yield and colour change by pH-sensitive green emission. Nat. Photonics 2008, 2, 44–47. [Google Scholar] [CrossRef]
- Viviani, V.R.; Arnoldi, F.G.C.; Neto, A.S.; Oehlmeyer, T.L.; Bechara, E.J.H.; Ohmiya, Y. The structural origin and biological function of pH-sensitivity in firefly luciferases. Photochem. Photobiol. Sci. 2008, 7, 159–169. [Google Scholar] [CrossRef]
- Oliveira, G.; Viviani, V.R. Comparison of the thermostability of recombinant luciferases from Brazilian bioluminescent beetles: Relationship with kinetics and bioluminescence colours. Luminescence 2018, 33, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Pelentir, G.F.; Bevilaqua, V.R.; Viviani, V.R. A highly efficient, thermostable and cadmium selective firefly luciferase suitable for ratiometric metal and pH biosensing and for sensitive ATP assays. Photochem. Photobiol. Sci. 2019, 18, 2061–2070. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; Viviani, V.R. Temperature effect on the bioluminescence spectra of firefly luciferases: Potential applicability for ratiometric biosensing of temperature and pH. Photochem. Photobiol. Sci. 2019, 18, 2682–2687. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Wang, Y.; Hiyama, M.; Akiyama, H. Robust Red-Emission Spectra and Yields in Firefly Bioluminescence against Temperature Changes. Appl. Phys. Lett. 2014, 104, 213704. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; Murtiashaw, M.H.; Magyar, R.A.; Gonzalez, S.A.; Ruggiero, M.C.; Stroh, J.G. An alternative mechanism of bioluminescence color determination in firefly luciferase. Biochemistry 2004, 43, 7255–7262. [Google Scholar] [CrossRef]
- Ugarova, N.N.; Brovko, L.Y. Protein structure and bioluminescent spectra for firefly bioluminescence. Lumin. J. Biol. Chem. Lumin. 2002, 17, 321–330. [Google Scholar] [CrossRef]
- Viviani, V.R.; Simões, A.; Bevilaqua, V.R.; Gabriel GV, M.; Arnoldi FG, C.; Hirano, T. Glu311 and Arg337 stabilize a closed active-site conformation and provide a critical catalytic base and countercation for green bioluminescence in beetle luciferases. Biochemistry 2016, 55, 4764–4776. [Google Scholar] [CrossRef]
- Granier, T.; Comberton, G.; Gallois, B.; d’Estaintot, B.L.; Dautant, A.; Crichton, R.R.; Précigoux, G. Evidence of new cadmium binding sites in recombinant horse L-chain ferritin by anomalous Fourier difference map calculation. Proteins 1998, 31, 477–485. [Google Scholar] [CrossRef]
- Friedman, R. Structural and computational insights into the versatility of cadmium binding to proteins. Dalton Trans. 2014, 43, 2878–2887. [Google Scholar] [CrossRef]
- Hirano, T.; Hasumi, Y.; Ohtsuka, K.; Maki, S.; Niwa, H.; Yamaji, M.; Hashizume, D. Spectroscopic studies of the light-color modulation mechanism of firefly (beetle) bioluminescence. J. Am. Chem. Soc. 2009, 131, 2385–2396. [Google Scholar] [CrossRef]
- Carrasco-Lopez, C.; Lui, N.M.; Schramm, S.; Naumov, P. The elusive relationship between structure and color emission in beetle luciferases. Nat. Rev. Chem. 2021, 5, 4–20. [Google Scholar] [CrossRef]
- Regan, L. Protein Design: Novel metal-binding sites. Trends Biochem. Sci. 1995, 20, 280–285. [Google Scholar] [CrossRef]
- Sciortino, G.; Garribba, E.; Rodríguez-Guerra Pedregal, J.; Maréchal, J.D. Simple Coordination Geometry Descriptors Allow to Accurately Predict Metal-Binding Sites in Proteins. ACS Omega 2019, 4, 3726–3731. [Google Scholar] [CrossRef] [PubMed]
- Niwa, K.; Ichino, Y.; Kumata, S.; Nakajima, Y.; Hiraishi, Y.; Kato, D.I.; Viviani, V.R.; Ohmiya, Y. Quantum yields and kinetics of the firefly bioluminescence reaction of beetle luciferases. Photochem. Photobiol. 2010, 86, 1046–1049. [Google Scholar] [CrossRef]
- Enomoto, T.; Kubota, H.; Mori, K.; Shimogawara, M.; Yoshita, M.; Ohmiya, Y.; Akiyama, H. Absolute bioluminescence imaging at the single-cell level with a light signal at the Attowatt level. Biotechniques 2018, 64, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Koop, A.; Cobbold, P.H. Continuous bioluminescent monitoring of cytoplasmic ATP in single isolated rat hepatocytes during metabolic poisoning. Biochem. J. 1993, 295, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Viviani, V.R.; Bechara, E.J.; Ohmiya, Y. Cloning, sequence analysis, and expression of active Phrixothrix railroad-worms luciferases: Relationship between bioluminescence spectra and primary structures. Biochemistry 1999, 38, 8271–8279. [Google Scholar] [CrossRef]
- Ohmiya, Y.; Sumiya, M.; Viviani, V.R.; Ohba, N. Comparative aspects of a luciferase molecule from the Japanese luminous beetle, Rhagophthalmus ohbai. Sci. Rep. Yokosuka City Mus 2000, 47, 31–38. [Google Scholar]
- Jathoul, A.P.; Grounds, H.; Anderson, J.C.; Pule, M.A. A dual-color far-red to near-infrared firefly luciferin analogue designed for multiparametric bioluminescence imaging. Angew. Chem. (Int. Ed. Engl.) 2014, 53, 13059–13063. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Luciferase | RA (%) | pH Sensitivity (nm) | RA ZnSO4 1 mM (%) | ZnSO4 Detection Limit * (µM) | ZnSO4 Spectral Shift (nm) ** | RA CdSO4 1 mM (%) | CdSO4 Detection Limit * (µM) | CdSO4 Spectral Shift (nm) ** | RA HgCl2 1 mM (%) | HgCl2 Detection Limit * (µM) | HgCl2 Spectral Shift (nm) ** | Applicability |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Macrolampis sp2 | 100 | 569–616 | 77 | 500 | 9 | 36 | 890 | 19 | 18 | 170 | 34 | pH-indicator |
| N354H | 5.7 | 568–615 | 20 | 110 | 30 | 21 | 2000 | 5 | 7 | 150 | 37 | pH-indicator |
| N354C | 75 | 564–606 | 8 | 20 | 47 | 13 | 15 | 50 | 6 | 15 | 37 | Zinc, Cadmium and Mercury enzymatic sensor |
| H310C | 62 | 573–613 | 20 | 100 | 15 | - | 3500 | 0 | 11 | 90 | 34 | pH-indicator |
| H310C/N354 | - | - | 15 | 15 | 34 | 11 | 260 | 41 | 4 | 30 | 36 | Zinc and Mercury enzymatic sensors |
| N354E | - | - | 90 | 225 | 23 | 14 | 4000 | 0 | 7 | 130 | 21 | |
| Cratomorphus distinctus | 100 | 554–614 | - | - | 36 | - | - | - | - | - | 46 | pH-indicator |
| Amydetes viviani | 100 | 549–596 | 2 | 2000 | 4 | 4 | 100 | 29 | 1,5 | 60 | 33 | Cadmium and Mercury enzymatic sensors; thermostable pH indicator |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viviani, V.R.; Pelentir, G.F.; Bevilaqua, V.R. Bioluminescence Color-Tuning Firefly Luciferases: Engineering and Prospects for Real-Time Intracellular pH Imaging and Heavy Metal Biosensing. Biosensors 2022, 12, 400. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12060400
Viviani VR, Pelentir GF, Bevilaqua VR. Bioluminescence Color-Tuning Firefly Luciferases: Engineering and Prospects for Real-Time Intracellular pH Imaging and Heavy Metal Biosensing. Biosensors. 2022; 12(6):400. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12060400
Chicago/Turabian StyleViviani, Vadim R., Gabriel F. Pelentir, and Vanessa R. Bevilaqua. 2022. "Bioluminescence Color-Tuning Firefly Luciferases: Engineering and Prospects for Real-Time Intracellular pH Imaging and Heavy Metal Biosensing" Biosensors 12, no. 6: 400. https://0-doi-org.brum.beds.ac.uk/10.3390/bios12060400