
Necrotizing Gingivitis: Microbial Diversity and Quantification of Protein Secretion in Necrotizing Gingivitis

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Patients and Sample Collection
2.2. DNA Extraction
2.3. Amplification and Illumina MiSeq High-Throughput Sequencing
2.4. Analysis of the 16S rRNA Data
2.5. Fluorescence In Situ Hybridization (FISH)
2.6. Image Acquisition and Analysis
2.7. Determination of Cytokines, Chemokines, and Growth Factors
2.8. Statistical Analysis
3. Results
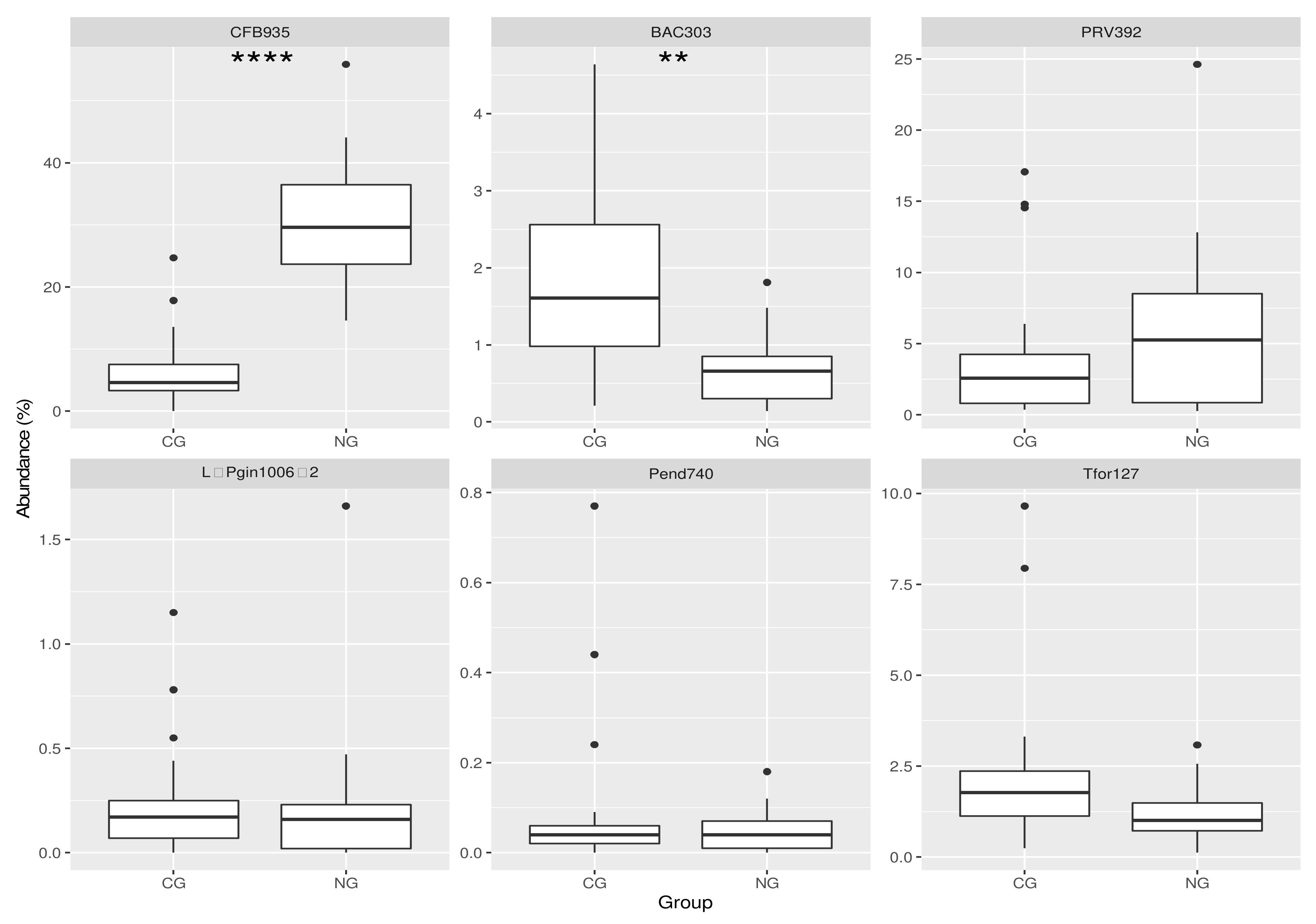
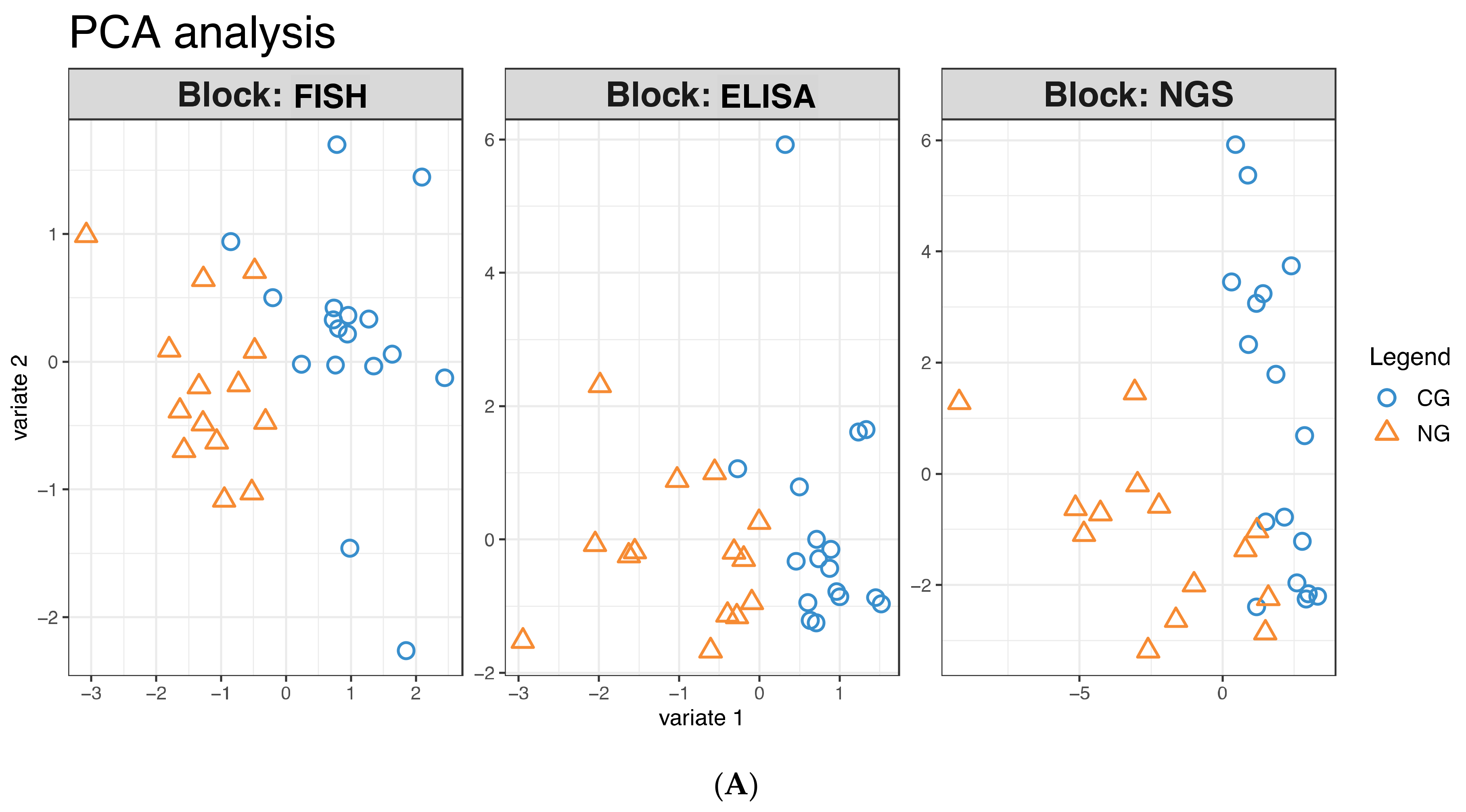
3.1. Analysis of the Bacterial Communities in the Two Cohorts: FISH
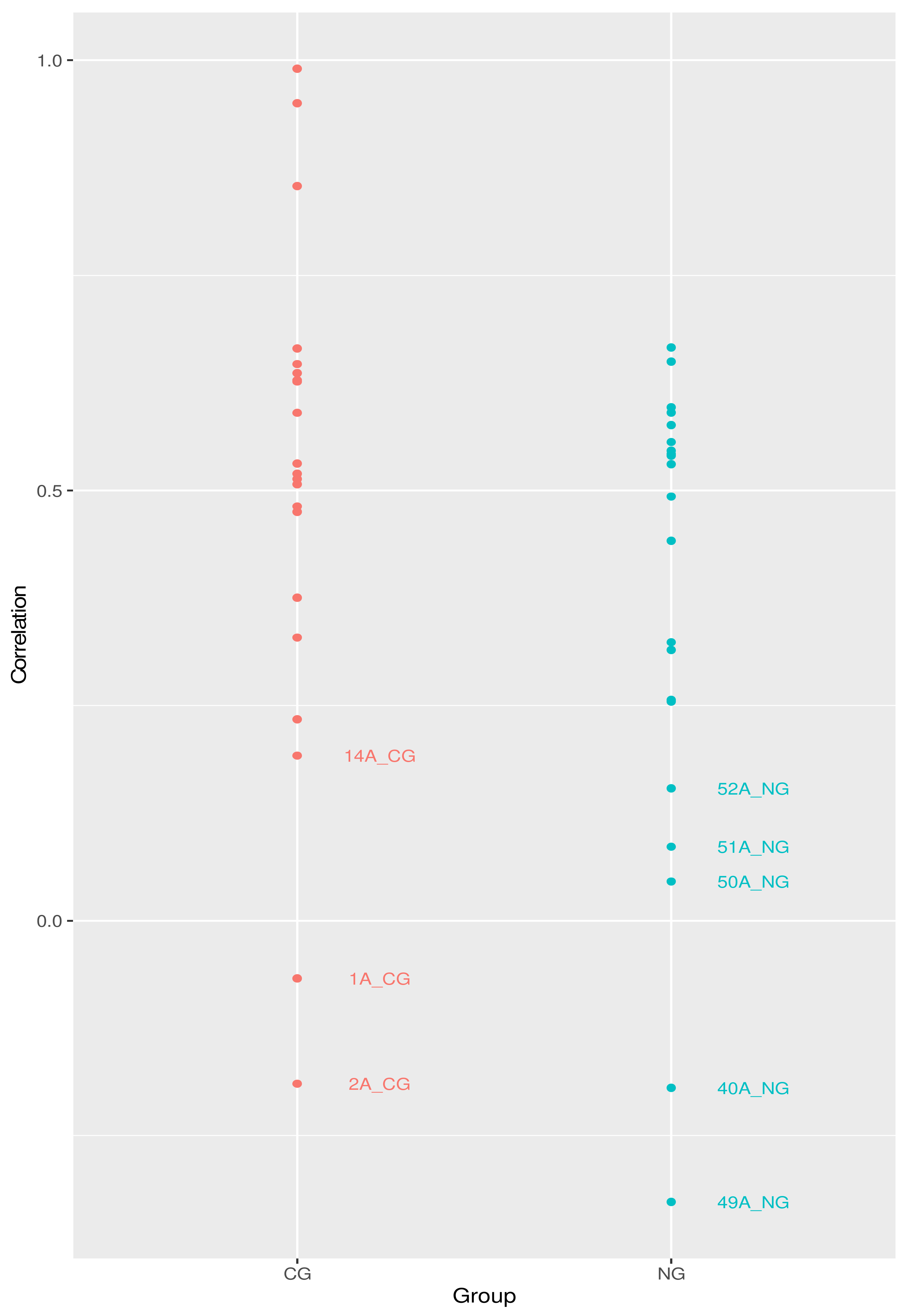
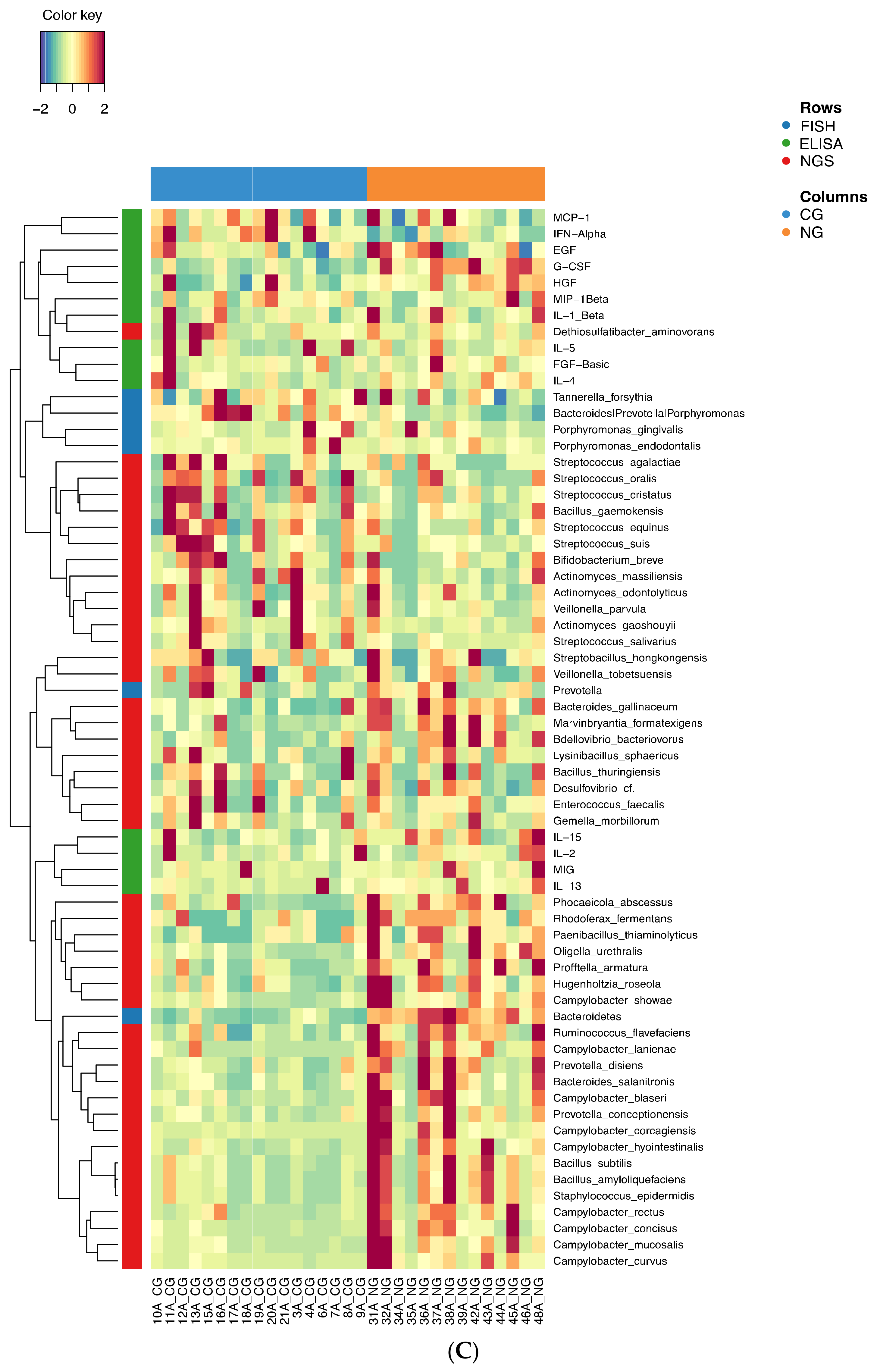
3.2. Analysis of the Bacterial Communities in the Two Cohorts: 16S rRNA NGS Comparison with FISH Data
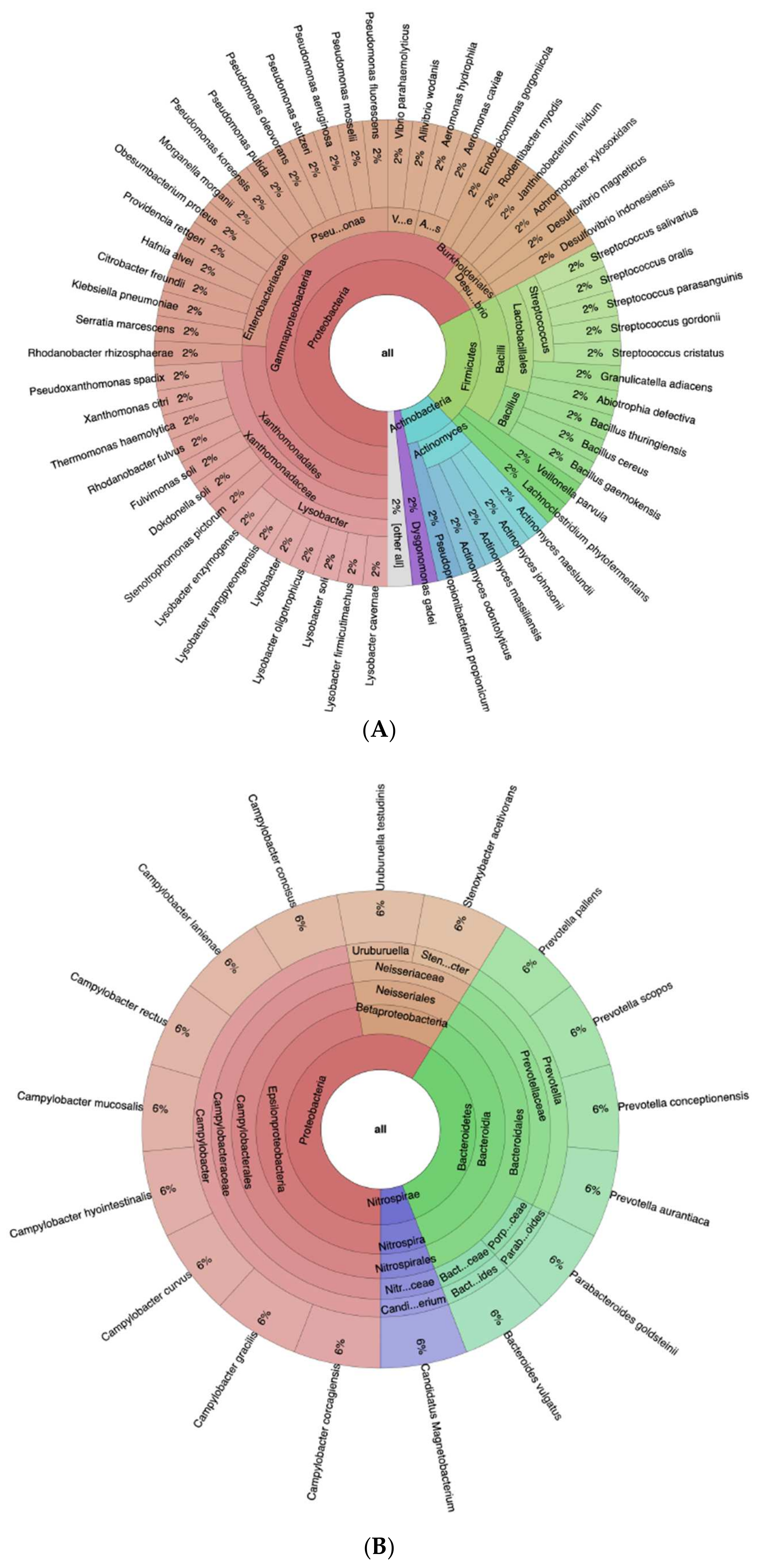
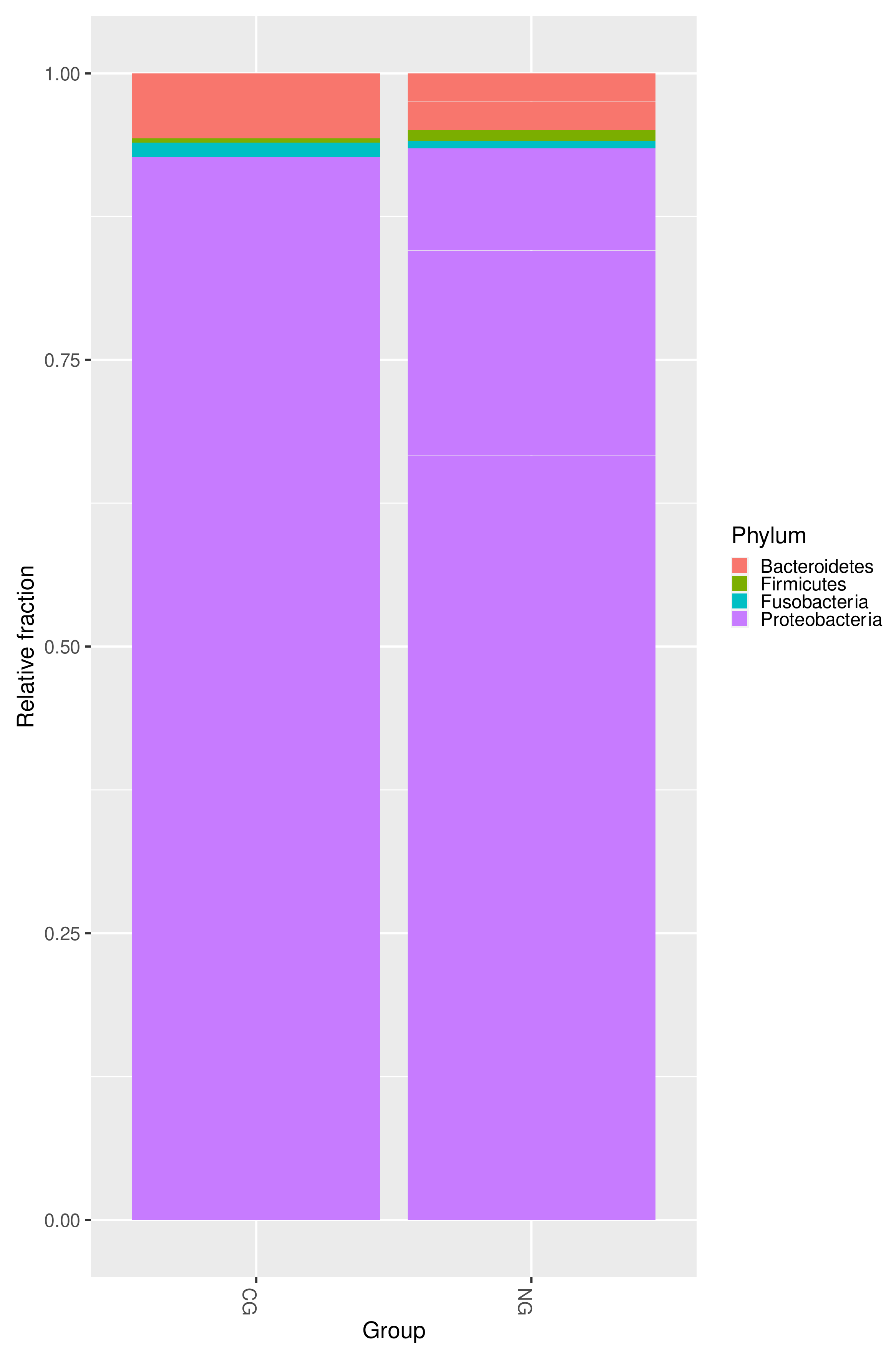
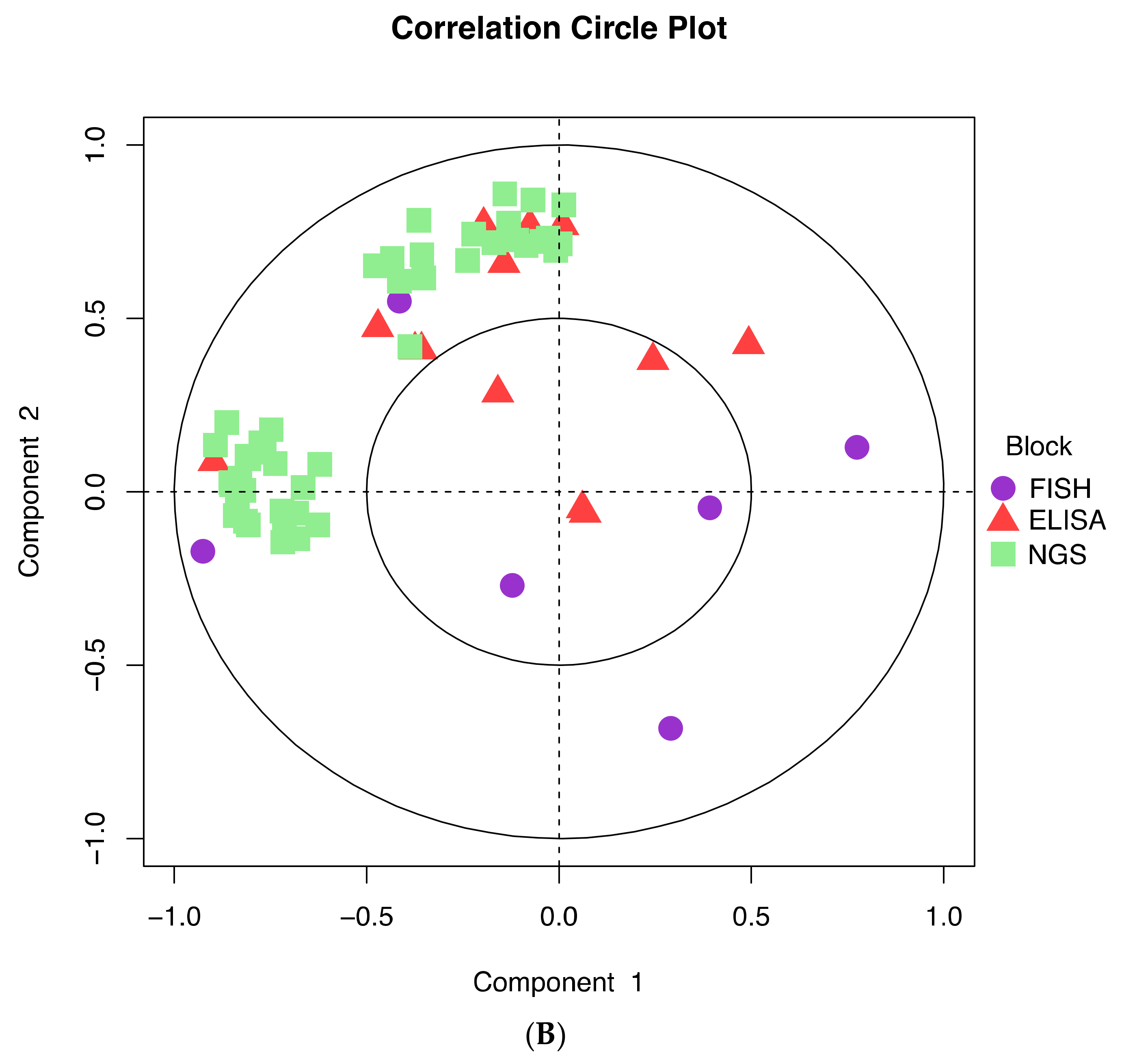
3.3. Further Profiling the Microbial Communities
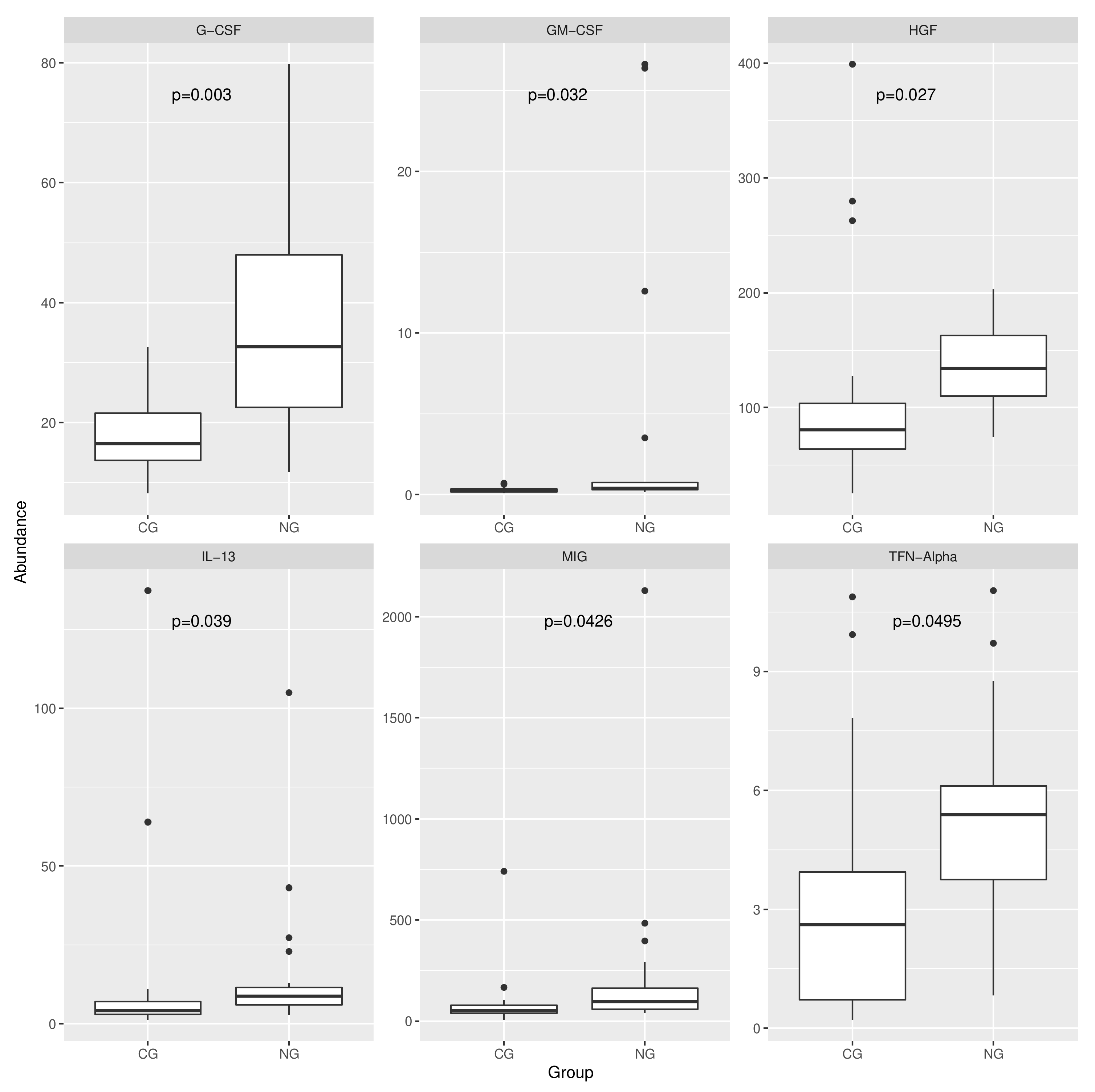
3.4. Analysis of the Expression of the Protein Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrera, D.; Alonso, B.; De Arriba, L.; Cruz, I.S.; Serrano, C.; Sanz, M. Acute Periodontal Lesions. Periodontol. 2000 2014, 65, 149–177. [Google Scholar] [CrossRef] [Green Version]
- Rowland, R.W. Necrotizing Ulcerative Gingivitis. Ann. Periodontol. 1999, 4, 65–73. [Google Scholar] [CrossRef]
- Herrera, D.; Retamal-Valdes, B.; Alonso, B.; Feres, M. Acute Periodontal Lesions (Periodontal Abscesses and Necrotizing Periodontal Diseases) and Endo-Periodontal Lesions. J. Clin. Periodontol. 2018, 45, S78–S94. [Google Scholar] [CrossRef] [Green Version]
- Shangase, L.; Feller, L.; Blignaut, E. Necrotising Ulcerative Gingivitis/Periodontitis as Indicators of HIV-Infection. J. S. Afr. Dent. Assoc. 2004, 59, 105–108. [Google Scholar]
- Smitt, P.A. Some Clinical and Epidemiological Aspects of Vincent’s Gingivitis. Dent. Pract. Dent. Rec. 1965, 15, 281–286. [Google Scholar] [PubMed]
- Hartnett, C.A.; Shiloah, J. The Treatment of Acute Necrotizing Ulcerative Gingivitis. Quintessence Int. 1991, 22, 95–100. [Google Scholar] [PubMed]
- Courtois, G.J., 3rd; Cobb, C.M.; Killoy, W.J. Acute Necrotizing Ulcerative Gingivitis. A Transmission Electron Microscope Study. J. Periodontol. 1983, 54, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Falkler, W.A.; Martin, S.A.; Vincent, J.W.; Tall, B.; Nauman, R.K.; Suzuki, J.B. A Clinical, Demographic and Microbiologic Study of ANUG Patients in an Urban Dental School. J. Clin. Periodontol. 1987, 14, 307–314. [Google Scholar] [CrossRef]
- Listgarten, M.A. Electron Microscopic Observations on the Bacterial Flora of Acute Necrotizing Ulcerative Gingivitis. J. Periodontol. 1965, 36, 328–339. [Google Scholar] [CrossRef]
- Plaut, H. Studien zur Bakteriellen Diagnostik der Diphtherie und der Anginen.4. DMW-Dtsch. Med. Wochenschr. 1894, 20, 920–923. [Google Scholar] [CrossRef] [Green Version]
- Vincent, M. Recherches Bacteriologiques sur l’Angine a Bacilles Fusiformes. Ann. Inst. Pasteur. 1899, 13, 609. [Google Scholar]
- Vincent, H. Sur l’Étiologie et sur les Lésions Anatomo-Pathologiques de la Pourriture d’Hôpital. Ann. Inst. Pasteur. 1896, 10, 488. [Google Scholar]
- Gmür, R.; Wyss, C.; Xue, Y.; Thurnheer, T.; Guggenheim, B. Gingival Crevice Microbiota from Chinese Patients with Gingivitis or Necrotizing Ulcerative Gingivitis. Eur. J. Oral Sci. 2004, 112, 33–41. [Google Scholar] [CrossRef]
- Ranney, R.R. Classification of Periodontal Diseases. Periodontol. 2000 1993, 2, 13–25. [Google Scholar] [CrossRef]
- Dufty, J.; Gkranias, N.; Donos, N. Necrotising Ulcerative Gingivitis: A Literature Review. Oral Health Prev. Dent. 2017, 15, 321–327. [Google Scholar]
- Jiménez, L.M.; Duque, F.L.; Baer, P.N.; Jiménez, S.B. Necrotizing Ulcerative Periodontal Diseases in Children and Young Adults in Medellin, Colombia, 1965--2000. J. Int. Acad. Periodontol. 2005, 7, 55–63. [Google Scholar]
- Cobb, C.M.; Ferguson, B.L.; Keselyak, N.T.; Holt, L.A.; MacNeill, S.R.; Rapley, J.W. A TEM/SEM Study of the Microbial Plaque Overlying the Necrotic Gingival Papillae of HIV-Seropositive, Necrotizing Ulcerative Periodontitis. J. Periodontal Res. 2003, 38, 147–155. [Google Scholar] [CrossRef]
- Horning, G.M.; Hatch, C.L.; Lutskus, J. The Prevalence of Periodontitis in a Military Treatment Population. J. Am. Dent. Assoc. 1990, 121, 616–622. [Google Scholar] [CrossRef]
- Melnick, S.L.; Roseman, J.M.; Engel, D.; Cogen, R.B. Epidemiology of Acute Necrotizing Ulcerative Gingivitis. Epidemiol. Rev. 1988, 10, 191–211. [Google Scholar] [CrossRef]
- Dibart, S. Children, Adolescents and Periodontal Diseases. J. Dent. 1997, 25, 79–89. [Google Scholar] [CrossRef]
- Löe, H.; Theilade, E.; Jensen, S.B. Experimental Gingivitis in Man. J. Periodontol. 1965, 36, 177–187. [Google Scholar] [CrossRef]
- Theilade, E, The Experimental Gingivitis Studies: The Microbiological Perspective. J. Dent. Res. 1996, 75, 1434–1438. [CrossRef]
- Ranney, R.R. Discussion: Pathogenesis of Gingivitis. J. Clin. Periodontol. 1986, 13, 356–359. [Google Scholar] [CrossRef]
- Moore, L.V.; Moore, W.E.; Cato, E.P.; Smibert, R.M.; Burmeister, J.A.; Best, A.M.; Ranney, R.R. Bacteriology of Human Gingivitis. J. Dent. Res. 1987, 66, 989–995. [Google Scholar] [CrossRef]
- Caton, J.G.; Armitage, G.; Berglundh, T.; Chapple, I.L.; Jepsen, S.; Kornman, K.S.; Mealey, B.L.; Papapanou, P.N.; Sanz, M.; Tonetti, M.S. A New Classification Scheme for Periodontal and Peri-implant Diseases and Conditions—Introduction and Key Changes from the 1999 Classification. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S1–S8. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Mealey, B.L.; Van Dyke, T.E.; Bartold, P.M.; Dommisch, H.; Eickholz, P.; Geisinger, M.L.; Genco, R.J.; Glogauer, M.; Goldstein, M.; et al. Periodontal Health and Gingival Diseases and Conditions on an Intact and a Reduced Periodontium: Consensus Report of Workgroup 1 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89 (Suppl. 1), S74–S84. [Google Scholar] [CrossRef]
- Trombelli, L.; Farina, R.; Silva, C.; Tatakis, D.N. Plaque-Induced Gingivitis: Case Definition and Diagnostic Considerations. J. Periodontol. 2018, 89 (Suppl. 1), S46–S73. [Google Scholar] [CrossRef] [Green Version]
- Ramseier, C.A.; Mirra, D.; Schütz, C.; Sculean, A.; Lang, N.P.; Walter, C.; Salvi, G.E. Bleeding on Probing as it Relates to Smoking Status in Patients Enrolled in Supportive Periodontal Therapy for at Least 5 Years. J. Clin. Periodontol. 2015, 42, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Ainamo, J.; Bay, I. Problems and Proposals for Recording Gingivitis and Plaque. Int. Dent. J. 1975, 25, 229–235. [Google Scholar]
- Khocht, A.; Rogers, T.; Janal, M.; Brown, M. Gingival Fluid Inflammatory Biomarkers and Hypertension in African Americans. JDR Clin. Transl. Res. 2017, 2, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Belibasakis, G.N. Gingival Crevicular Fluid and its Immune Mediators in the Proteomic Era. Periodontol. 2000 2018, 76, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Gmür, R.; Munson, M.A.; Wade, W.G. Genotypic and Phenotypic Characterization of Fusobacteria from Chinese and European Patients with Inflammatory Periodontal Diseases. Syst. Appl. Microbiol. 2006, 29, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Züger, J.; Lüthi-Schaller, H.; Gmür, R. Uncultivated Tannerella BU045 and BU063 are Slim Segmented Filamentous Rods of High Prevalence but Low Abundance in Inflammatory Disease-Associated Dental Plaques. Microbiology 2007, 153, 3809–3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, A.; Thurnheer, T.; Lüthi-Schaller, H.; Gmür, R.; Belibasakis, G.N. The Phylum Synergistetes in Gingivitis and Necrotizing Ulcerative Gingivitis. J. Med. Microbiol. 2012, 61, 1600–1609. [Google Scholar] [CrossRef] [Green Version]
- Wyss, C.; Moter, A.; Choi, B.K.; Dewhirst, F.E.; Xue, Y.; Schüpbach, P.; Göbel, U.B.; Paster, B.J.; Guggenheim, B. Treponema putidum sp. nov., a Medium-sized Proteolytic Spirochaete Isolated from Lesions of Human Periodontitis and Acute Necrotizing Ulcerative Gingivitis. Int. J. Syst. Evol. Microbiol. 2004, 54, 1117–1122. [Google Scholar] [CrossRef] [Green Version]
- Wyss, C.; Dewhirst, F.E.; Gmür, R.; Thurnheer, T.; Xue, Y.; Schüpbach, P.; Guggenheim, B.; Paster, B.J. Treponema parvum sp. nov., a Small, Glucoronic or Galacturonic Acid-Dependent Oral Spirochaete from Lesions of Human Periodontitis and Acute Necrotizing Ulcerative Gingivitis. Int. J. Syst. Evol. Microbiol. 2001, 5, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Zinkernagel, A.S.; Gmür, R.; Fenner, L.; Schaffner, A.; Schoedon, G.; Schneemann, M. Marginal and Subgingival Plaque—A Natural Habitat of Tropheryma whipplei? Infection 2003, 31, 86–91. [Google Scholar] [CrossRef]
- Loesche, W.J.; Hockett, R.N.; Syed, S.A. The Predominant Cultivable Flora of Tooth Surface Plaque Removed from Institutionalized Subjects. Arch. Oral Biol. 1972, 17, 1311–1325. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M. Analysis of Actinomycete Communities by Specific Amplification of Genes Encoding 16S rRNA and Gel-Electrophoretic Separation in Denaturing Gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Aronesty, E. Comparison of Sequencing Utility Programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Gmür, R.; Lüthi-Schaller, H. A Combined Immunofluorescence and Fluorescent in Situ Hybridization Assay for Single Cell Analyses of Dental Plaque Microorganisms. J. Microbiol. Methods 2007, 69, 402–405. [Google Scholar] [CrossRef]
- Manz, W.; Amann, R.; Ludwig, W.; Vancanneyt, M.; Schleifer, K.H. Application of a Suite of 16S rRNA-Specific Oligonucleotide Probes Designed to Investigate Bacteria of the Phylum Cytophaga-Flavobacter-Bacteroides in the Natural Environment. Microbiology 1996, 142, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Shirazi-Beechey, S.P.; Daly, K. Design and Evaluation of Group-Specific Oligonucleotide Probes for Quantitative Analysis of Intestinal Ecosystems: Their Application to Assessment of Equine Colonic Microflora. FEMS Microbiol. Ecol. 2003, 44, 243–252. [Google Scholar]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-Targeted Oligonucleotide Probes with Flow Cytometry for Analyzing Mixed Microbial Populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Guggenheim, B.; Gmür, R.; Galicia, J.C.; Stathopoulou, P.G.; Benakanakere, M.R.; Meier, A.; Thurnheer, T.; Kinane, D.F. In Vitro Modeling of Host-Parasite Interactions: The ‘Subgingival‘ Biofilm Challenge of Primary Human Epithelial Cells. BMC Microbiol. 2009, 9, 280. [Google Scholar] [CrossRef] [Green Version]
- Lüdin, N.D. Phylogenetic Characterization of Subgingival Plaque Associated with Aggressive or Chronic Periodontitis. M.D. Dissertation, University of Zurich, Zürich, Switzerland, 2011. [Google Scholar]
- Diaz, P.I.; Chalmers, N.I.; Rickard, A.H.; Kong, C.; Milburn, C.L.; Palmer, R.J.; Kolenbrander, P.E. Molecular Characterization of Subject-Specific Oral Microflora During Initial Colonization of Enamel. Appl. Environ. Microbiol. 2006, 72, 2837–2848. [Google Scholar] [CrossRef] [Green Version]
- Manz, W. In Situ Analysis of Microbial Biofilms by rRNA-Targeted Oligonucleotide Probing. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; pp. 79–91. [Google Scholar]
- Thurnheer, T.; Gmür, R.; Giertsen, E.; Guggenheim, B. Automated Fluorescent in Situ Hybridization for the Specific Detection and Quantification of Oral Streptococci in Dental Plaque. J. Microbiol. Methods 2001, 44, 39–47. [Google Scholar] [CrossRef]
- Dunbar, S.A.; Ritchie, V.B.; Hoffmeyer, M.R.; Rana, G.S.; Zhang, H. Luminex(R) Multiplex Bead Suspension Arrays for the Detection and Serotyping of Salmonella spp. Methods Mol. Biol. 2015, 1225, 1–27. [Google Scholar]
- Gürkan, A.; Tekdal, G.P.; Bostancı, N.; Belibasakis, G.N. Cytokine, Chemokine, and Growth Factor Levels in Peri-Implant Sulcus During Wound Healing and Osseointegration After Piezosurgical Versus Conventional Implant Site Preparation: Randomized, Controlled, Split-Mouth Trial. J. Periodontol. 2018, 90, 616–626. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, M.; Rechenberg, D.-K.; Thurnheer, T.; Lüthi-Schaller, H.; Belibasakis, G. FISHing for Gutta-Percha-Adhered Biofilms in Purulent Post-Treatment Apical Periodontitis. Mol. Oral Microbiol. 2017, 32, 226–235. [Google Scholar] [CrossRef]
- Thurnheer, T.; Gmür, R.; Guggenheim, B. Multiplex FISH Analysis of a Six-Species Bacterial Biofilm. J. Microbiol. Methods 2004, 56, 37–47. [Google Scholar] [CrossRef]
- Paster, B.J.; Boches, S.K.; Galvin, J.L.; Ericson, R.E.; Lau, C.N.; Levanos, V.A.; Sahasrabudhe, A.; Dewhirst, F.E. Bacterial Diversity in Human Subgingival Plaque. J. Bacteriol. 2001, 183, 3770–3783. [Google Scholar] [CrossRef] [Green Version]
- Wade, W.G.; Prosdocimi, E.M. Profiling of Oral Bacterial Communities. J. Dent. Res. 2020, 99, 621–629. [Google Scholar] [CrossRef]
- Bermejo-Fenoll, A.; Sanchez-Perez, A. Necrotising Periodontal Diseases. Med. Oral Patol. Oral Cir. Bucal. 2004, 9 (Suppl. 114–119), 108–114. [Google Scholar] [PubMed]
- Loesche, W.J.; Grossman, N.S. Periodontal Disease as a Specific, Albeit Chronic, Infection: Diagnosis and Treatment. Clin. Microbiol. Rev. 2001, 14, 727–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, R.; Hujoel, P.; Belibasakis, G.N. On Putative Periodontal Pathogens: An Epidemiological Perspective. Virulence 2015, 6, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The Domain-specific Probe EUB338 is Insufficient for the Detection of all Bacteria: Development and Evaluation of a more Comprehensive Probe Set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Contaldo, M.; Lucchese, A.; Romano, A.; Della Vella, F.; Di Stasio, D.; Serpico, R.; Petruzzi, M. Oral Microbiota Features in Subjects with Down Syndrome and Periodontal Diseases: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 9251. [Google Scholar] [CrossRef]
- Amano, A.; Kuboniwa, A.M.; Nakagawa, I.; Akiyama, S.; Morisaki, I.; Hamada, S. Prevalence of Specific Genotypes of Porphyromonas gingivalis fimA and Periodontal Health Status. J. Dent. Res. 2000, 79, 1664–1668. [Google Scholar] [CrossRef]
- Bostanci, N.; Allaker, R.P.; Belibasakis, G.N.; Rangarajan, M.; Curtis, M.A.; Hughes, F.J.; McKay, I.J. Porphyromonas gingivalis Antagonises Campylobacter rectus Induced Cytokine Production by Human Monocytes. Cytokine 2007, 39, 147–156. [Google Scholar] [CrossRef]
- Felix, J.; Savvides, S.N. Mechanisms of Immunomodulation by Mammalian and Viral Decoy Receptors: Insights from Structures. Nat. Rev. Immunol. 2017, 17, 112–129. [Google Scholar] [CrossRef]
- Male, D.; Brostoff, J.; Roth, D.; Roitt, I. Immunology: With STUDENT CONSULT Online Access, 8th ed.; Elsevier: Edinburgh, UK, 2012. [Google Scholar]
- Muñoz-Carrillo, J.L.; Contreras-Cordero, J.F.; Gutiérrez-Coronado, O.; Villalobos-Gutiérrez, P.T.; Ramos-Gracia, L.G.; Hernández-Reyes, V.E. Cytokine Profiling Plays a Crucial Role in Activating Immune System to Clear Infectious Pathogens. In Immune Response Activation and Immunomodulation; IntechOpen: London, UK, 2018. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.H.; Pillai, S. Cellular and Molecular Immunology: With STUDENT CONSULT Online Access; Elsevier: Philadelphia, PA, USA, 2014. [Google Scholar]
- Metcalf, D. The Colony-Stimulating Factors and Cancer. Nat. Rev. Cancer 2010, 10, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.A. Colony-Stimulating Factors in Inflammation and Autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Khajah, M.; Millen, B.; Cara, D.C.; Waterhouse, C.; McCafferty, D.M. Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF): A Chemoattractive Agent for Murine Leukocytes in Vivo. J. Leukoc. Biol. 2011, 89, 945–953. [Google Scholar] [CrossRef]
- Gomez-Cambronero, J.; Horn, J.; Paul, C.C.; Baumann, M.A. Granulocyte-Macrophage Colony-Stimulating Factor Is a Chemoattractant Cytokine for Human Neutrophils: Involvement of the Ribosomal p70 S6 Kinase Signaling Pathway. J. Immunol. 2003, 171, 6846–6855. [Google Scholar] [CrossRef] [Green Version]
- Molineux, G.; Foote, M.; Arvedson, T. Twenty Years of G-CSF Clinical and Nonclinical Discoveries, in Milestones in Drug Therapy; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Carulli, G. Effects of Recombinant Human Granulocyte Colony-Stimulating Factor Administration on Neutrophil Phenotype and Functions. Haematologica 1997, 82, 606–616. [Google Scholar]
- Kawakami, M.; Tsutsumi, H.; Kumakawa, T.; Abe, H.; Hirai, M.; Kurosawa, S.; Mori, M.; Fukushima, M. Levels of Serum Granulocyte Colony-Stimulating Factor in Patients with Infections. Blood 1990, 76, 1962–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Grady, N.P.; Preas, H.L.; Pugin, J.; Fiuza-Marco, C.; Tropea, M.; Reda, D.; Banks, S.M.; Suffredini, A.F. Local Inflammatory Responses Following Bronchial Endotoxin Instillation in Humans. Am. J. Respir. Crit. Care Med. 2001, 163, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, P.; Thiruppathi, M.; Elshabrawy, H.A.; Alharshawi, K.; Kumar, P.; Prabhakar, B.S. GM-CSF: An Immune Modulatory Cytokine that Can Suppress Autoimmunity. Cytokine 2015, 75, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trus, E.; Basta, S.; Gee, K. Who’s in Charge Here? Macrophage Colony Stimulating Factor and Granulocyte Macrophage Colony Stimulating Factor: Competing Factors in Macrophage Polarization. Cytokine 2019, 127, 154939. [Google Scholar] [CrossRef]
- Hamilton, J.A. Rheumatoid Arthritis: Opposing Actions of Haemopoietic Growth Factors and Slow-Acting Anti-Rheumatic Drugs. Lancet 1993, 342, 536–539. [Google Scholar] [CrossRef]
- Yucel-Lindberg, T.; Nilsson, S.; Modéer, T. Signal Transduction Pathways Involved in the Synergistic Stimulation of Prostaglandin Production by Interleukin-1 Beta and Tumor Necrosis Factor Alpha in Human Gingival Fibroblasts. J. Dent. Res. 1999, 78, 61–68. [Google Scholar] [CrossRef]
- Zhang, F.; Engebretson, S.P.; Morton, R.S.; Cavanaugh, P.F.; Subbaramaiah, K.; Dannenberg, A.J. The Overexpression of Cyclo-oxygenase-2 in Chronic Periodontitis. J. Am. Dent. Assoc. 2003, 134, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, K.; Miyauchi, M.; Oka, H.; Komaki, M.; Somerman, M.J.; Takata, T. Cyclooxygenase-2-Dependent Prostaglandin E(2) Upregulates Interleukin (IL)-1alpha-Induced IL-6 Generation in Mouse Cementoblasts. J. Periodontol. 2007, 78, 135–140. [Google Scholar] [CrossRef]
- de Oliveira, R.R.; Schwartz-Filho, H.O.; Novaes, A.B.; Garlet, G.P.; de Souza, R.F.; Taba, M.; Scombatti de Souza, S.L.; Ribeiro, F.J. Antimicrobial Photodynamic Therapy in the Non-Surgical Treatment of Aggressive Periodontitis: Cytokine Profile in Gingival Crevicular Fluid, Preliminary Results. J. Periodontol. 2009, 80, 98–105. [Google Scholar] [CrossRef]
- Tervahartiala, T.; Koski, H.; Xu, J.-W.; Häyrinen-Immonen, R.; Hietanen, J.; Sorsa, T.; Konttinen, Y. Tumor Necrosis Factor-a and its Receptors, p55 and p75, in Gingiva of Adult Periodontitis. J. Dent. Res. 2001, 80, 1535–1539. [Google Scholar] [CrossRef]
- Santos, V.R.; Ribeiro, F.; Lima, J.A.; Napimoga, M.; Bastos, M.F.; Duarte, P.M. Cytokine Levels in Sites of Chronic Periodontitis of Poorly Controlled and Well-Controlled Type 2 Diabetic Subjects. J. Clin. Periodontol. 2010, 37, 1049–1058. [Google Scholar] [CrossRef]
- Bastos, M.; Lima, J.; Vieira, P.; Mestnik, M.; Faveri, M.; Duarte, P. TNF-Alpha and IL-4 Levels in Generalized Aggressive Periodontitis Subjects. Oral Dis. 2009, 15, 82–87. [Google Scholar] [CrossRef]
- Tamura, M.; Arakaki, N.; Tsubouchi, H.; Takada, H.; Daikuhara, Y. Enhancement of Human Hepatocyte Growth Factor Production by Interleukin-1 Alpha and -1 Beta and Tumor Necrosis Factor-Alpha by Fibroblasts in Culture. J. Biol. Chem 1993, 268, 8140–8145. [Google Scholar] [CrossRef]
- Ohnishi, T.; Suwa, M.; Oyama, T.; Arakaki, N.; Torii, M.; Daikuhara, Y. Prostaglandin E2 Predominantly Induces Production of Hepatocyte Growth Factor/Scatter Factor in Human Dental Pulp in Acute Inflammation. J. Dent. Res. 2000, 79, 748–755. [Google Scholar] [CrossRef]
- Sugiyama, A.; Ogawa, T.; Daikuhara, Y.; Takada, H. Enhancement of Hepatocyte Growth Factor (Scatter Factor) Production by Human Gingival Fibroblasts in Culture Stimulated with Porphyromonas gingivalis Fimbriae. J. Med. Microbiol. 2000, 49, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Iki, K.; Kawahara, K.; Sawamura, S.; Arakaki, R.; Sakuta, T.; Sugiyama, A.; Tamura, H.; Sueda, T.; Hamada, S.; Takada, H. A Novel Component Different from Endotoxin Extracted from Prevotella intermedia ATCC 25611 Activates Lymphoid Cells from C3H/HeJ Mice and Gingival Fibroblasts from Humans. Infect. Immun. 1997, 65, 4531–4538. [Google Scholar] [CrossRef] [Green Version]
- Anil, S.; Vellappally, S.; Preethanath, R.S.; Mokeem, S.A.; AlMoharib, H.S.; Patil, S.; Chalisserry, E.; Al Kheraif, A.A. Hepatocyte Growth Factor Levels in the Saliva and Gingival Crevicular Fluid in Smokers with Periodontitis. Dis. Markers 2014, 2014, 146974. [Google Scholar] [CrossRef]
- Ohshima, M.; Sakai, A.; Ito, K.; Otsuka, K. Hepatocyte Growth Factor (HGF) in Periodontal Disease: Detection of HGF in Gingival Crevicular Fluid. J. Periodontal Res. 2002, 37, 8–14. [Google Scholar] [CrossRef]
- Minty, A.; Chalon, P.; Derocq, J.M.; Dumont, X.; Guillemot, J.C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B. Interleukin-13 is a New Human Lymphokine Regulating Inflammatory and Immune Responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef]
- Beklen, A. Effects of IL-13 on TGF-β and MMP-1 in Periodontitis. Biotech. Histochem. 2017, 92, 374–380. [Google Scholar] [CrossRef]
- Gonzales, J.R.; Gröger, S.; Haley, G.; Bödeker, R.-H.; Meyle, J. Production of Interleukin-13 is Influenced by the Interleukin-4 -34TT and -590TT Genotype in Patients with Aggressive Periodontitis. Scand. J. Immunol. 2011, 73, 128–134. [Google Scholar] [CrossRef]
- Wynn, T.A. IL-13 Effector Functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef]
- Liu, C.; Papewalis, C.; Domberg, J.; Scherbaum, W.; Schott, M. Chemokines and Autoimmune Thyroid Diseases. Horm. Metab. Res. 2008, 40, 361–368. [Google Scholar] [CrossRef]
- Antonelli, A.; Ferrari, S.M.; Giuggioli, D.; Ferrannini, E.; Ferri, C.; Fallahi, P. Chemokine (C-X-C motif) Ligand (CXCL)10 in Autoimmune Diseases. Autoimmun. Rev. 2014, 13, 272–280. [Google Scholar] [CrossRef]
- Smit, M.J.; Verdijk, P.; van der Raaij-Helmer, E.M.H.; Navis, M.; Hensbergen, P.J.; Leurs, R.; Tensen, C.P. CXCR3-Mediated Chemotaxis of Human T Cells is Regulated by a Gi- and Phospholipase C-Dependent Pathway and Not Via Activation of MEK/p44/p42 MAPK Nor Akt/PI-3 Kinase. Blood 2003, 102, 1959–1965. [Google Scholar] [CrossRef]
- de Queiroz, A.C.; Taba, M.; O’Connell, P.A.; da Nóbrega, P.B.; Costa, P.P.; Kawata, V.K.; Trevisan, G.L.; Novaes, A.B.; de Souza, S.L.; Palioto, D.B.; et al. Inflammation Markers in Healthy and Periodontitis Patients: A Preliminary Data Screening. Braz. Dent. J. 2008, 19, 3–8. [Google Scholar] [CrossRef]
- Paparo, S.R. The MIG Chemokine in Inflammatory Myopathies. Clin. Ter. 2019, 170, e55–e60. [Google Scholar]
- Tecco, S.; Grusovin, M.G.; Sciara, S.; Bova, F.; Pantaleo, G.; Capparé, P. The Association Between Three Attitude-Related Indexes of Oral Hygiene and Secondary Implant Failures: A Retrospective Longitudinal Study. Int. J. Dent. Hyg. 2018, 16, 372–379. [Google Scholar] [CrossRef]
- Crespi, R.; Capparè, P.; Gherlone, E. Sinus Floor Elevation by Osteotome: Hand Mallet Versus Electric Mallet. A Prospective Clinical Study. Int. J. Oral Maxillofac. Implant. 2012, 27, 1144–1150. [Google Scholar]
- Gherlone, E.; Capparé, P.; Pasciuta, R.; Grusovin, M.G.; Mancini, N.; Burioni, R. Evaluation of Resistance Against Bacterial Microleakage of a New Conical Implant-Abutment Connection Versus Conventional Connections: An in Vitro Study. New Microbiol. 2016, 39, 49–56. [Google Scholar]
- Capparè, P.; Tetè, G.; Sberna, M.T.; Panina-Bordignon, P. The Emerging Role of Stem Cells in Regenerative Dentistry. Curr. Gene Ther. 2020, 20, 259–268. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe 1 | Target | Sequence (5′–3) 2 | 5′ modification | Target Site | F (%) | Source |
|---|---|---|---|---|---|---|
| BAC303 | Most Bacteroidaceae and Prevotellaceae, some Porphyromonadaceae | CCA ATG TGG GGG ACC TT | Cy3 | 303–319 | 50 | [45] |
| CFB935 | Bacteroidetes (very broad) | CCA CAT GTT CCT CCG CTT GT | Cy3 | 935–954 | 50 | [46] |
| EUB338 | Many but not all bacteria/most eubacteria | GCT GCC TCC CGT AGG AGT | Carboxyfluorescein | 338–355 | 40–50 | [47] |
| L-Pgin1006–2 | P. gingivalis | GTT TTC ACC ATC MGT CAT C | Cy3 | 1006–1024 | 45 | [48] |
| Pend740 | P. endodontalis | CAG TGT CAG ACG GAG CCT | Cy3 | 740–757 | 40 | [49] |
| PRV392 | Prevotellaceae (Prevotella, Hallella)/Prevotellaceae (Prevotella spp., Alloprevotella spp., Hallella spp.) | GCA CGC TAC TTG GCT GG | Cy3 | 392–308 | 50 | [50] |
| Tfor127 | T. forsythia | CTC TGT TGC GGG CAG GTT AC | Cy3 | 127–146 | 40 | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerhard, N.; Thurnheer, T.; Kreutzer, S.; Gmür, R.D.; Attin, T.; Russo, G.; Karygianni, L. Necrotizing Gingivitis: Microbial Diversity and Quantification of Protein Secretion in Necrotizing Gingivitis. Antibiotics 2021, 10, 1197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101197
Gerhard N, Thurnheer T, Kreutzer S, Gmür RD, Attin T, Russo G, Karygianni L. Necrotizing Gingivitis: Microbial Diversity and Quantification of Protein Secretion in Necrotizing Gingivitis. Antibiotics. 2021; 10(10):1197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101197
Chicago/Turabian StyleGerhard, Nicolas, Thomas Thurnheer, Susanne Kreutzer, Rudolf Dominik Gmür, Thomas Attin, Giancarlo Russo, and Lamprini Karygianni. 2021. "Necrotizing Gingivitis: Microbial Diversity and Quantification of Protein Secretion in Necrotizing Gingivitis" Antibiotics 10, no. 10: 1197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101197