
Essential Oils: A Natural Weapon against Antibiotic-Resistant Bacteria Responsible for Nosocomial Infections



Abstract
:1. Introduction
2. Results
2.1. Antibacterial Susceptibility and Synergy Testing
2.2. Time–Kill Studies
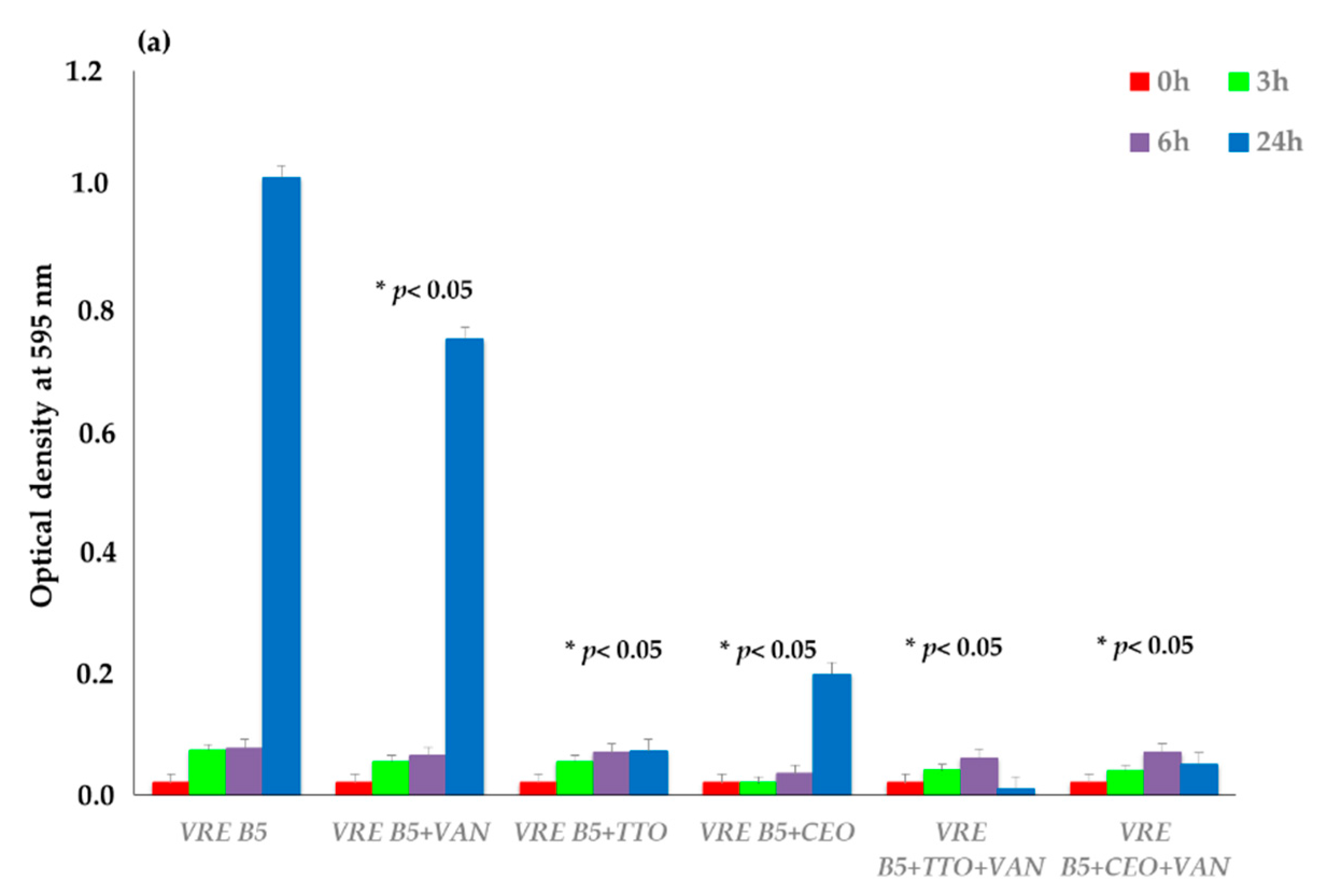
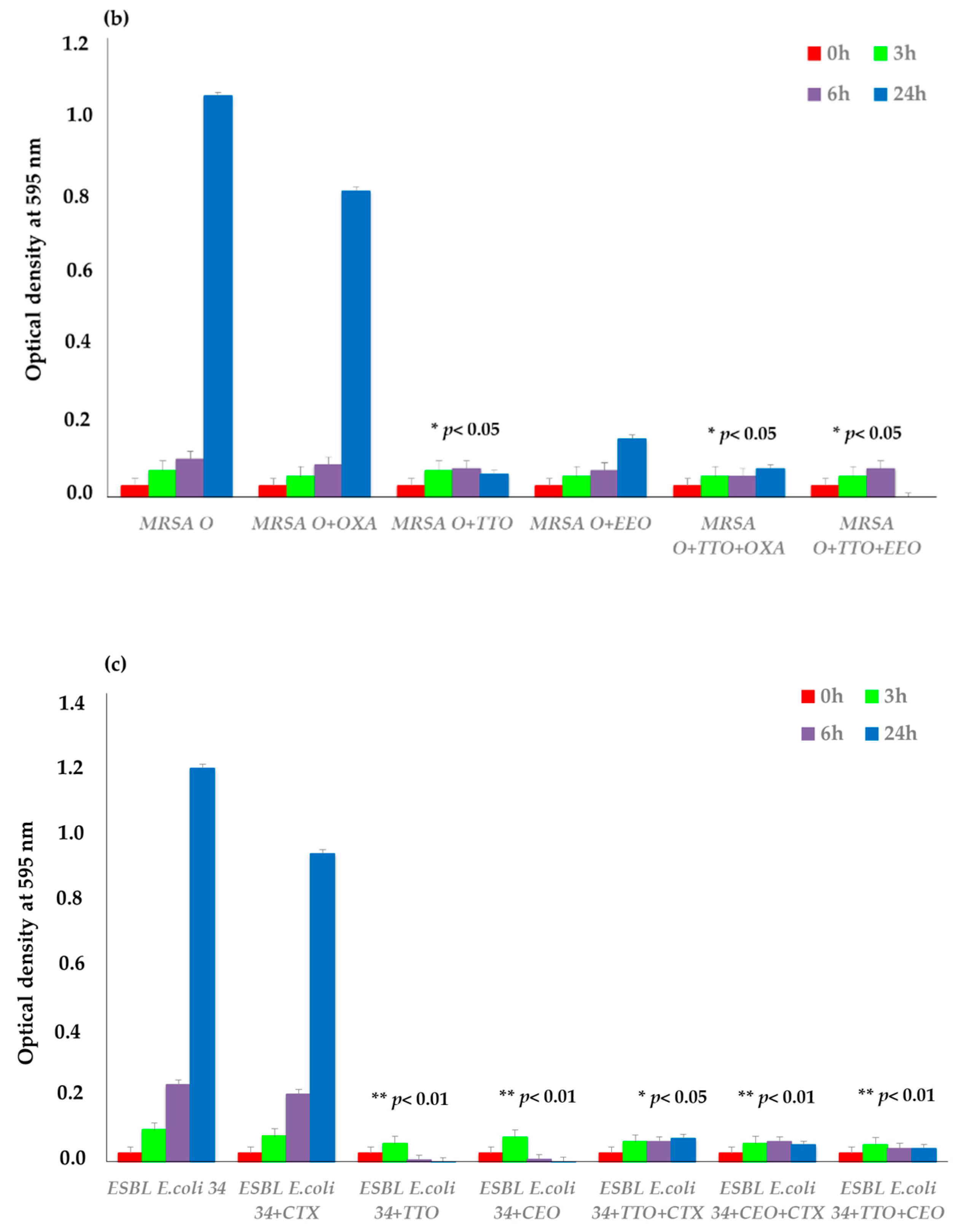
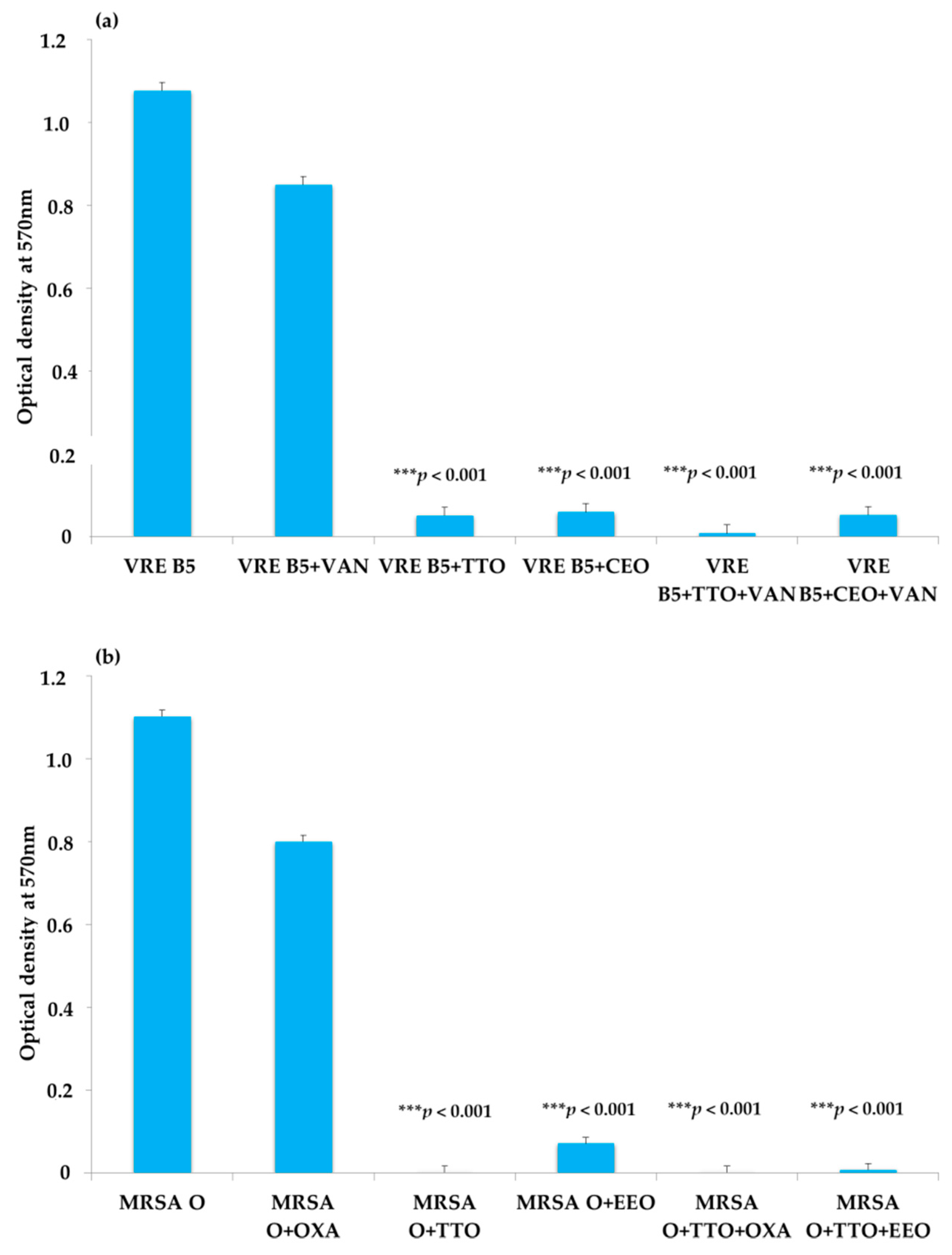
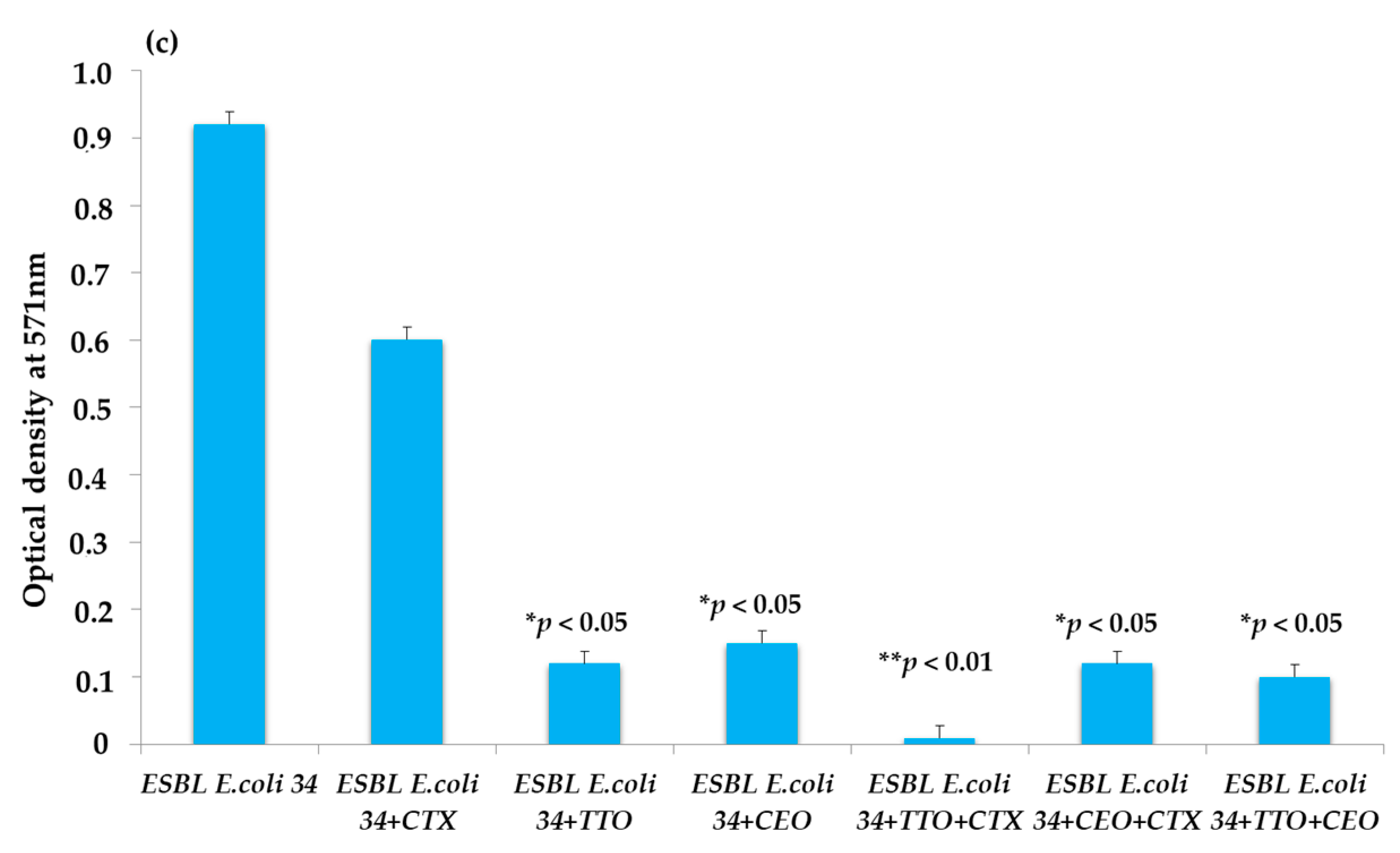
2.3. EO Activity on Mature Biofilm
3. Discussion
4. Materials and Methods
4.1. Essential Oils
4.2. Microbial Strains
4.3. Antibacterial Susceptibility Testing
4.4. Determination of the Fractional Inhibitory (FIC) Index—Synergistic Testing
4.5. Time–Kill Studies
4.6. EO Activity on Mature Biofilm
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64 (Suppl. 1), i29–i36. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Antimicrobial Awareness Week-WAAW from 18 to 24 November 2020. Available online: https://www.who.int/campaigns/world-antimicrobial-awareness-week/2020 (accessed on 9 April 2021).
- Bereket, W.; Hemalatha, K.; Getenet, B.; Wondwossen, T.; Solomon, A.; Zeynudin, A.; Kannan, S. Update on bacterial nosocomial infections. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1039–1044. [Google Scholar]
- Cassini, A.; Högberg, L.D.; Plachoura, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Kalan, L.; Wright, G.D. Antibiotic adjuvants: Multicomponent anti-infective strategies. Expert Rev. Mol. Med. 2011, 13, e5. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, H.; Gousia, P.; Economou, V.; Sakkas, V.; Petsios, S.; Papadopoulou, C. In vitro antimicrobial activity of five essential oils on multidrug resistant Gram-negative clinical isolates. J. Intercult. Ethnopharmacol. 2016, 5, 212–218. [Google Scholar] [CrossRef]
- Borges, A.; Lopez-Romero, J.C.; Oliveira, D.; Giaouris, E.; Simões, M. Prevention, removal and inactivation of Escherichia coli and Staphylococcus aureus biofilms using selected monoterpenes of essential oils. J. Appl. Microbiol. 2017, 123, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, M.J.; Ilanko, A.; Blonk, B.; Cock, I.E. Developing New Antimicrobial Therapies: Are Synergistic Combinations of Plant Extracts/Compounds with Conventional Antibiotics the Solution? Pharmacogn. Rev. 2017, 11, 57–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical characterization and evaluation of the antibacterial activity of essential oils from fibre-type Cannabis sativa L. (Hemp). Molecules 2019, 24, 2302. [Google Scholar] [CrossRef] [Green Version]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. Antimicrobial, Antioxidant, and Immunomodulatory Properties of Essential Oils: A Systematic Review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef] [Green Version]
- Condò, C.; Anacarso, I.; Sabia, C.; Iseppi, R.; Anfelli, I.; Forti, L.; de Niederhäusern, S.; Bondi, M.; Messi, P. Antimicrobial activity of spices essential oils and its effectiveness on mature biofilms of human pathogens. Nat. Prod. Res. 2020, 34, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Okla, M.K.; Alamri, S.A.; Salem, M.Z.M.; Ali, H.M.; Behiry, S.I.; Nasser, R.A.; Alaraidh, I.A.; Al-Ghtani, S.M.; Soufan, W. Yield, Phytochemical Constituents, and Antibacterial Activity of Essential Oils from the Leaves/Twigs, Branches, Branch Wood, and Branch Bark of Sour Orange (Citrus aurantium L.). Processes 2019, 7, 363. [Google Scholar] [CrossRef] [Green Version]
- Rguez, S.; Djébali, N.; Ben Slimene, I.; Abid, G.; Hammemi, M.; Chenenaoui, S.; Bachkouel, S.; Daami-Remadi, M.; Ksouri, R.; Hamrouni Sellami, I. Cupressus sempervirens essential oils and their major compounds successfully control postharvest grey mould disease of tomato. Ind. Crop. Prod. 2018, 123, 135–141. [Google Scholar] [CrossRef]
- Papadopoulos, C.J.; Carson, C.F.; Chang, B.J.; Riley, T.V. Role of the MexAB-OprM efflux pump of Pseudomonas aeruginosa in tolerance to tea tree (Melaleuca alternifolia) oil and its monoterpene components terpinen-4-ol, 1,8-cineole, and α-terpineol. Appl. Environ. Microbiol. 2008, 74, 1932–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.O.; Holley, R.A. Disruption of Escherichia coli, Listeria monocytogenes and Lactobacillus sakei cellular membranes by plant oil aromatics. Int. J. Food Microbiol. 2006, 108, 1–9. [Google Scholar] [CrossRef]
- Gibbons, S. Phytochemicals for bacterial resistance—Strengths, weaknesses and opportunities. Planta Med. 2008, 74, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemaiswarya, S.; Doble, M. Synergistic interaction of eugenol with antibiotics against Gram negative bacteria. Phytomedicine 2009, 16, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Langeveld, W.T.; Veldhuizen, E.J.; Burt, S.A. Synergy between essential oil components and antibiotics: A review. Crit. Rev. Microbiol. 2014, 40, 76–94. [Google Scholar] [CrossRef]
- Kuok, C.F.; Hoi, S.O.; Hoi, C.F.; Chan, C.H.; Fong, I.H.; Ngok, C.K.; Meng, L.R.; Fong, P. Synergistic antibacterial effects of herbal extracts and antibiotics on methicillin-resistant Staphylococcus aureus: A computational and experimental study. Exp. Biol. Med. 2017, 242, 731–743. [Google Scholar] [CrossRef] [Green Version]
- Ait Said, L.; Zahlane, K.; Ghalbane, I.; El Messoussi, S.; Romane, A.; Cavaleiro, C.; Salgueiro, L. Chemical composition and antibacterial activity of Lavandula coronopifolia essential oil against antibiotic-resistant bacteria. Nat. Prod. Res. 2015, 29, 582–585. [Google Scholar] [CrossRef]
- Benameur, Q.; Gervasi, T.; Pellizzeri, V.; Pľuchtová, M.; Tali-Maama, H.; Assaous, F.; Guettou, B.; Rahal, K.; Gruľov, D.; Dugo, G.; et al. Antibacterial activity of Thymus vulgaris essential oil alone and in combination with cefotaxime against blaESBL producing multidrug resistant Enterobacteriaceae isolates. Nat Prod Res. 2019, 33, 2647–2654. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.S.; Albuquerque Azevedo, Á.M.; Gomes Pereira, A.M.; Rocha, R.R.; Bastos Cavalcante, R.M.; Carneiro Matos, M.N.; Ribeiro Lopes, P.H.; Gomes, G.A.; Soares Rodrigues, T.H.; Santos, H.S.D.; et al. Chemical Composition and Antimicrobial Effectiveness of Ocimum gratissimum L. Essential Oil against Multidrug-Resistant Isolates of Staphylococcus aureus and Escherichia coli. Molecules 2019, 24, 3864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, P.R.; de Araújo, A.C.J.; Dos Santos Barbosa, C.R.; Muniz, D.F.; Rocha, J.E.; de Araújo Neto, J.B.; da Silva, M.M.C.; Silva Pereir, R.L.; da Silva, L.E.; do Amaral, W.; et al. Characterization and antibacterial activity of the essential oil obtained from the leaves of Baccharis coridifolia DC against multiresistant strains. Microb. Pathog. 2020, 145, 104223. [Google Scholar] [CrossRef] [PubMed]
- Naeim, H.; El-Hawiet, A.; Abdel Rahman, R.A.; Hussein, A.; El Demellawy, M.A.; Embaby, A.M. Antibacterial activity of Centaurea pumilio L. root and aerial part extracts against some multidrug resistant bacteria. BMC Complement. Med. Ther. 2020, 20, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Ferreira, S.; Silva, F.; Domingues, F.C. Synergistic activity of coriander oil and conventional antibiotics against Acinetobacter baumannii. Phytomedicine 2012, 19, 236–238. [Google Scholar] [CrossRef]
- Naveed, R.; Hussain, I.; Tawab, A.; Tariq, M.; Rahman, M.; Hameed, S.; Mahmood, M.S.; Siddique, A.B.; Iqbal, M. Antimicrobial activity of the bioactive components of essential oils from Pakistani spices against Salmonella and other multi-drug resistant bacteria. BMC Complement. Altern. Med. 2013, 13, 265. [Google Scholar] [CrossRef] [Green Version]
- Yap, P.S.; Lim, S.H.; Hu, C.P.; Yiap, B.C. Combination of essential oils and antibiotics reduce antibiotic resistance in plasmid-conferred multidrug resistant bacteria. Phytomedicine 2013, 20, 710–713. [Google Scholar] [CrossRef]
- Zharkova, M.S.; Orlov, D.S.; Golubeva, O.Y.; Chakchir, O.B.; Eliseev, I.E.; Grinchuk, T.M.; Shamova, O.V. Application of Antimicrobial Peptides of the Innate Immune System in Combination With Conventional Antibiotics-A Novel Way to Combat Antibiotic Resistance? Front. Cell Infect. Microbiol. 2019, 30, 128. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowski, P.; Pruss, A.; Grygorcewicz, B.; Wojciuk, B.; Dołęgowska, B.; Giedrys-Kalemba, S.; Kochan, E.; Sienkiewicz, M. Preliminary Study on the Antibacterial Activity of Essential Oils Alone and in Combination with Gentamicin against Extended-Spectrum β-Lactamase-Producing and New Delhi Metallo-β-Lactamase-1-Producing Klebsiella pneumoniae Isolates. Microb. Drug Resist. 2018, 24, 1368–1375. [Google Scholar] [CrossRef]
- Alexopoulos, A.; Kimbaris, A.C.; Plessas, S.; Mantzouran, I.; Voidarou, C.; Pagonopoulou, O.; Tsigalou, C.; Fournomiti, M.; Bontsidis, C.; Stavropoulou, E.; et al. Combined Action of Piperitenone Epoxide and Antibiotics against Clinical Isolates of Staphylococcus aureus and Escherichia coli. Front. Microbiol. 2019, 10, 2607. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, S.H.; Lai, K.S. Lavender essential oil induces oxidative stress which modifies the bacterial membrane permeability of carbapenemase producing Klebsiella pneumoniae. Sci. Rep. 2020, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Owen, L.; Webb, J.P.; Green, J.; Smith, L.J.; Laird, K. From formulation to in vivo model: A comprehensive study of a synergistic relationship between vancomycin, carvacrol, and cuminaldehyde against Enterococcus faecium. Phytother. Res. 2020, 34, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Veeregowda, B.M.; Krishnappa, G. Biofilms: A survival strategy of bacteria. Curr. Sci. 2003, 85, 1299–1307. [Google Scholar]
- Messi, P. Biofilm formation, development and relevance. In Biofilm in Bioengineering; Nova Science Publishers, Inc.: New York, NY, USA, 2013; Volume 268, pp. 1–26. [Google Scholar]
- Bassolé, I.H.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Ramirez, L.A.; Gutiérrez-Pacheco, M.M.; Vargas-Arispuro, I.; González-Aguilar, G.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Inhibition of Glucosyltransferase Activity and Glucan Production as an Antibiofilm Mechanism of Lemongrass Essential Oil against Escherichia coli O157:H7. Antibiotics 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quave, C.L.; Horswill, A.R. Flipping the switch: Tools for detecting small molecule inhibitors of staphylococcal virulence. Front. Microbiol. 2014, 5, 706. [Google Scholar] [CrossRef] [Green Version]
- Oliva, A.; Costantini, S.; De Angelis, M.; Garzoli, S.; Božović, M.; Mascellino, M.T.; Vullo, V.; Ragno, R. High Potency of Melaleuca alternifolia Essential Oil against Multi-Drug Resistant Gram-Negative Bacteria and Methicillin-Resistant Staphylococcus aureus. Molecules 2018, 23, 2584. [Google Scholar] [CrossRef] [Green Version]
- Kon, K.V.; Rai, M.K. Plant essential oils and their constituents in coping with multidrug-resistant bacteria. Expert Rev. Anti. Infect. Ther. 2012, 10, 775–790. [Google Scholar] [CrossRef]
- Yap, P.S.; Krishnan, T.; Chan, K.G.; Lim, S.H. Antibacterial Mode of Action of Cinnamomum verum Bark Essential Oil, Alone and in Combination with Piperacillin, against a Multi-Drug-Resistant Escherichia coli Strain. J. Microbiol. Biotechnol. 2015, 25, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- El Atk, Y.; Aouam, I.; El Kamari, F.; Taroq, A.; Nayme, K.; Timinouni, M.; Lyoussi, B.; Abdellaoui, A. Antibacterial activity of cinnamon essential oils and their synergistic potential with antibiotics. J. Adv. Pharm. Technol. Res. 2019, 10, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, N.G.; Queiroz, J.H.F.S.; Silva, K.E.D.; Vasconcelos, P.C.P.; Croda, J.; Simionatto, S. Synergistic effects of Cinnamomum cassia L. essential oil in combination with polymyxin B against carbapenemase-producing Klebsiella pneumoniae and Serratia marcescens. PLoS ONE 2020, 15, e0236505. [Google Scholar] [CrossRef] [PubMed]
- Cuaron, J.A.; Dulal, S.; Song, Y.; Singh, A.K.; Montelongo, C.E.; Yu, W.; Nagarajan, V.; Jayaswal, R.K.; Wilkinson, B.J.; Gustafson, J.E. Tea tree oil-induced transcriptional alterations in Staphylococcus aureus. Phytother. Res. 2013, 27, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Faleiro, M.L. The mode of antibacterial action of essential oils. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Formatex Research Center: Norristown, PA, USA, 2011; Volume 3, pp. 1143–1156. [Google Scholar]
- Nazzaro, F.; Fratiann, F.; De Martino, L.; Coppol, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Romero, J.C.; González-Río, H.; Borges, A.; Simões, M. Antibacterial Effects and Mode of Action of Selected Essential Oils Components against Escherichia coli and Staphylococcus aureus. Evid. Based Complement. Altern. Med. 2015, 2015, 795435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cáceres, M.; Hidalgo, W.; Stashenko, E.; Torres, R.; Ortiz, C. Essential Oils of Aromatic Plants with Antibacterial, Anti-Biofilm and Anti-Quorum Sensing Activities against Pathogenic Bacteria. Antibiotics 2020, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burt, S. Essential oils: Their antimicrobial properties and potential applications in foods: A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Ouedrhiri, W.; Balouiri, M.; Harki, E.; Moja, S.; Greche, H. Synergistic antimicrobial activity of two binary combinations of marjoram, lavender, and wild thyme essential oils. Int. J. Food Prop. 2017, 20, 3149–3158. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019; Volume 39. [Google Scholar]
- Klancnik, A.; Piskernik, S.; Jersek, B.; Mozina, S.S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Methods 2010, 81, 121–126. [Google Scholar] [CrossRef]
- Hemaiswaryaa, S.; Kruthiventib, A.K.; Doblea, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Kwieciński, J.; Eick, S.; Wójcik, K. Effects of tea tree (Melaleuca alternifolia) oil on Staphylococcus aureus in biofilms and stationary growth phase. Int. J. Antimicrob. Agents 2009, 33, 343–347. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | VAN | Strains | OXA | ||
|---|---|---|---|---|---|
| E. faecium A29 | 512 | R | S. aureus 6 | 512 | R |
| E. faecium A30 | 512 | R | S. aureus 12A | 512 | R |
| E. faecium B5 | 512 | R | S. aureus 12B | 512 | R |
| E. faecium VAN 2 | 16 | R | S. aureus C1 | 512 | R |
| E. faecalis VAN 3 | 128 | R | S. aureus C3 | 512 | R |
| E. faecalis VAN 4 | 512 | R | S. aureus MRSA | 512 | R |
| E. faecium VAN 5 | 8 | R | S. aureus MRSA1 | 8 | R |
| E. faecalis VAN 19 | 8 | R | S. aureus MRSA2 | 8 | R |
| S. aureus O | 512 | R | |||
| Strains | CTX | Strains | CTX | ||
|---|---|---|---|---|---|
| E. coli 34 | 32 | R | E. coli 23CT | 64 | R |
| E. coli 39 | 64 | R | E. coli 40CT | 32 | R |
| E. coli 22A | 64 | R | E. coli 23DT | 64 | R |
| E. coli 26A | 32 | R | E. coli 24DT | 64 | R |
| E. coli 28A | 32 | R | E. coli 31DT | 32 | R |
| E. coli 38A | 32 | R | E. coli 45DT | 32 | R |
| E. coli 41A | 32 | R | E. coli 22F | 64 | R |
| E. coli 24AT | 64 | R | E. coli 23F | 64 | R |
| E. coli 36AT | 32 | R | E. coli 31FT | 64 | R |
| E. coli 24B | 64 | R | E. coli 27G | 64 | R |
| E. coli 22BT | 64 | R | E. coli 4 CL | 64 | R |
| E. coli 36BT | 64 | R | E. coli 9 CL | 16 | R |
| E. coli 24C | 8 | R | E. coli 11 CL | 64 | R |
| E. coli 22CT | 64 | R |
| Strains | AEO | LEO | EEO | TTO | CEO | Strains | AEO | LEO | EEO | TTO | CEO |
|---|---|---|---|---|---|---|---|---|---|---|---|
| E. faecium A29 | 256 | 64 | 8 | 1 | 32 | S. aureus 6 | >512 | >512 | 128 | 32 | >512 |
| E. faecium A30 | >512 | >512 | 8 | 16 | 128 | S. aureus 12A | >512 | 32 | 64 | 32 | >512 |
| E. faecium B5 | >512 | >512 | >512 | 8 | 64 | S. aureus 12B | >512 | 256 | >512 | 64 | >512 |
| E. faecium VAN2 | >512 | >512 | >512 | 128 | >512 | S. aureus C1 | >512 | >512 | 8 | 4 | >512 |
| E. faecalis VAN3 | >512 | 256 | 16 | 64 | 64 | S. aureus C3 | >512 | >512 | 32 | 8 | >512 |
| E. faecalis VAN4 | >512 | >512 | 512 | 8 | 128 | S. aureus MRSA | 128 | 32 | 32 | 8 | >512 |
| E. faecium VAN5 | >512 | >512 | >512 | 32 | >512 | S. aureus MRSA1 | >512 | >512 | >512 | 32 | 512 |
| E. faecalis VAN 19 | >512 | >512 | >512 | 64 | 256 | S. aureus MRSA2 | >512 | 32 | 8 | 16 | >512 |
| S. aureus O | >512 | >512 | 32 | 8 | >512 |
| Strains | AEO | LEO | EEO | TTO | CEO | Strains | AEO | LEO | EEO | TTO | CEO |
|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli 34 | >512 | 256 | >512 | 8 | 64 | E. coli 23CT | >512 | >512 | >512 | 2 | >512 |
| E. coli 39 | >512 | >512 | >512 | >512 | 32 | E. coli 40CT | >512 | 256 | >512 | 4 | >512 |
| E. coli 22A | >512 | 512 | >512 | 8 | 32 | E. coli 23DT | >512 | >512 | >512 | 2 | >512 |
| E. coli 26A | >512 | >512. | >512 | 1 | >512 | E. coli 24DT | >512 | >512 | >512 | 0.5 | >512 |
| E. coli 28A | >512 | 256 | 64 | 4 | 64 | E. coli 31DT | >512 | 512 | >512 | 1 | 64 |
| E. coli 38A | >512 | 256 | >512 | 4 | 128 | E. coli 45DT | >512 | >512 | 128 | 1 | 128 |
| E. coli 41A | >512 | 256 | >512 | 4 | >512 | E. coli 22F | >512 | 512 | >512 | 2 | 64 |
| E. coli 24AT | >512 | >512 | >512 | 4 | >512 | E. coli 23F | >512 | 128 | >512 | 1 | >512 |
| E. coli 36AT | >512 | >512 | >512 | 2 | >512 | E. coli 31FT | >512 | 512 | 256 | 16 | >512 |
| E. coli 24B | >512 | >512 | >512 | 0.5 | >512 | E. coli 27G | >512 | >512 | >512 | 2 | 64 |
| E. coli 22BT | >512 | 256 | 32 | 128 | 128 | E. coli 4 CL | >512 | >512 | >512 | 2 | >512 |
| E. coli 36BT | >512 | >512 | >512 | 1 | >512 | E. coli 9 CL | >512 | >512 | >512 | 4 | >512 |
| E. coli 24C | >512 | >512 | >512 | 256 | >512 | E. coli 11 CL | 512 | >512 | >512 | >512 | >512 |
| E. coli 22CT | >512 | 256 | >512 | 4 | 64 |
| Strains | TTO/CTX | CEO/CTX | TTO/CEO | Strains | TTO/VAN | CEO/VAN | TTO/CEO |
|---|---|---|---|---|---|---|---|
| E. coli 34 | 0.04 | 0.5 | 0.5 | E. faecium A29 | 0.25 | 0.5 | 0.5 |
| E. coli 22A | 0.27 | 0.5 | 0.5 | E. faecium A30 | 0.02 | 0.5 | 1.5 |
| E. coli 28A | 0.07 | 0.5 | 0.5 | E. faecium B5 | 0.03 | 0.5 | 1.5 |
| E. coli 38A | 0.07 | 0.5 | 0.5 | E. faecalis VAN4 | 0.5 | 0.5 | 1.5 |
| E. coli 22BT | 0.5 | 0.5 | 0.5 | ||||
| E. coli 23DT | 0.27 | 0.5 | 0.5 | Strains | TTO/OXA | EEO/OXA | TTO/EEO |
| E. coli 31DT | 0.26 | 0.5 | 1.5 | S. aureus 12A | 0.5 | 0.5 | 0.5 |
| E. coli 22F | 0.13 | 0.5 | 0.5 | S. aureus C3 | 0.03 | 0.5 | 0.5 |
| E. coli 27G | 0.14 | 0.5 | 2.0 | S. aureus MRSA | 0.04 | 2.0 | 0.5 |
| E. coli 9 CL | 0.27 | 0.5 | 0.5 | S. aureus O | 0.03 | 1.5 | 0.375 |
| Strains | MIC Alone | MIC Combined | FIC (A) | MIC Alone | MIC Combined | FIC (B) | FICI |
|---|---|---|---|---|---|---|---|
| VRE B5 | TTO | TTO | TTO | VAN | VAN | VAN | 0.032 |
| 8 | 0.25 | 0.03 | 256 | 0.5 | 0.002 | ||
| CEO | CEO | CEO | VAN | VAN | VAN | 0.5 | |
| 64 | 16 | 0.25 | 256 | 64 | 0.25 | ||
| MRSA O | TTO | TTO | TTO | OXA | OXA | OXA | 0.032 |
| 8 | 0.25 | 0.03 | 256 | 0.5 | 0.002 | ||
| EEO | EEO | EEO | TTO | TTO | TTO | 0.375 | |
| 32 | 8 | 0.25 | 8 | 1 | 0.125 | ||
| ESBL E. coli 34 | TTO | TTO | TTO | CTX | CTX | CTX | 0.038 |
| 8 | 0.25 | 0.03 | 32 | 0.25 | 0.008 | ||
| CEO | CEO | CEO | CTX | CTX | CTX | 0.5 | |
| 64 | 16 | 0.25 | 32 | 8 | 0.25 | ||
| TTO | TTO | TTO | CEO | CEO | CEO | 0.5 | |
| 8 | 2 | 0.25 | 64 | 16 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iseppi, R.; Mariani, M.; Condò, C.; Sabia, C.; Messi, P. Essential Oils: A Natural Weapon against Antibiotic-Resistant Bacteria Responsible for Nosocomial Infections. Antibiotics 2021, 10, 417. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040417
Iseppi R, Mariani M, Condò C, Sabia C, Messi P. Essential Oils: A Natural Weapon against Antibiotic-Resistant Bacteria Responsible for Nosocomial Infections. Antibiotics. 2021; 10(4):417. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040417
Chicago/Turabian StyleIseppi, Ramona, Martina Mariani, Carla Condò, Carla Sabia, and Patrizia Messi. 2021. "Essential Oils: A Natural Weapon against Antibiotic-Resistant Bacteria Responsible for Nosocomial Infections" Antibiotics 10, no. 4: 417. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10040417