
Colistin Selection of the Mcr-1 Gene in Broiler Chicken Intestinal Microbiota

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Clinical Signs and Pathological Findings
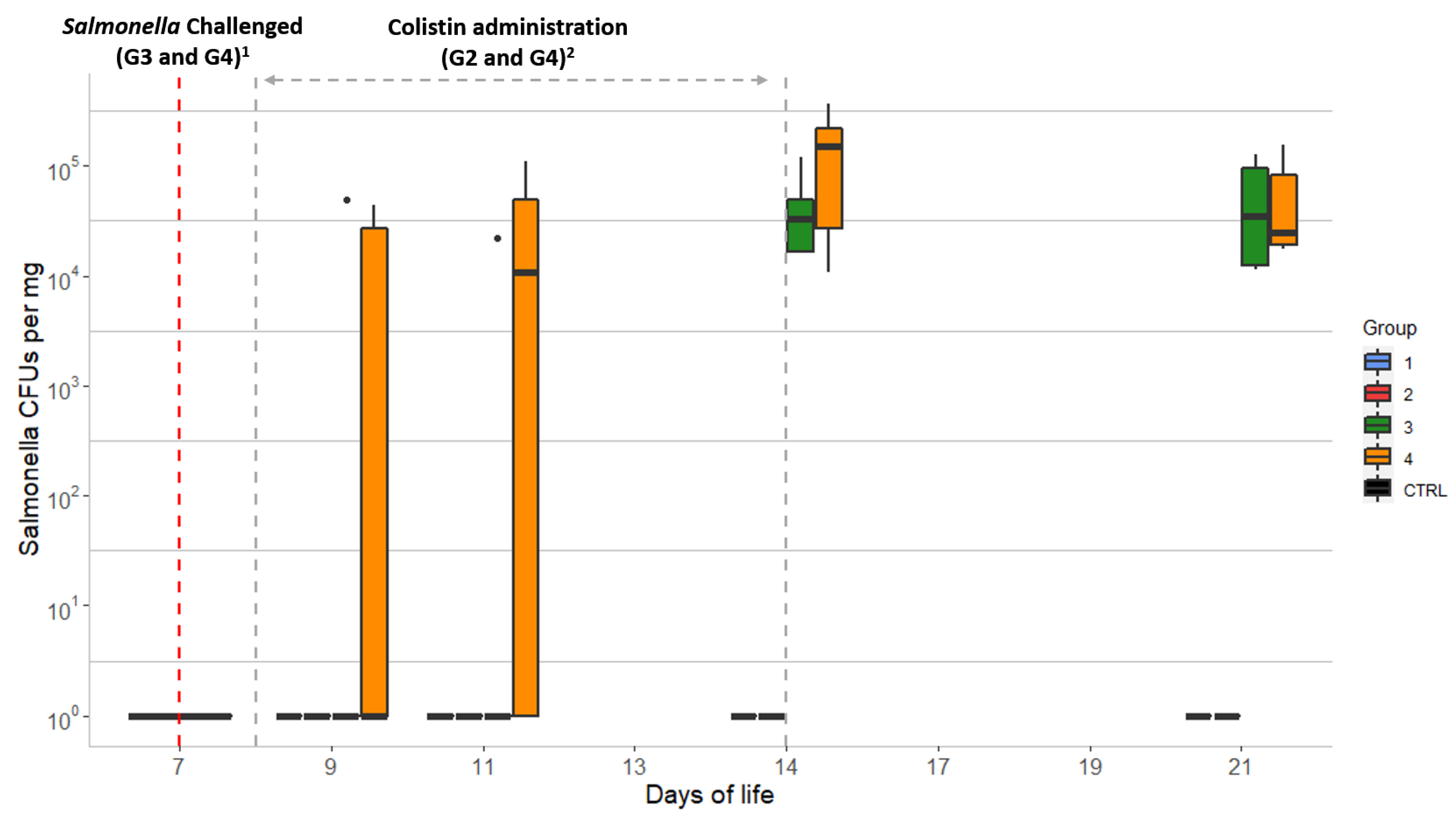
2.2. Salmonella Counts in the Chick Samples
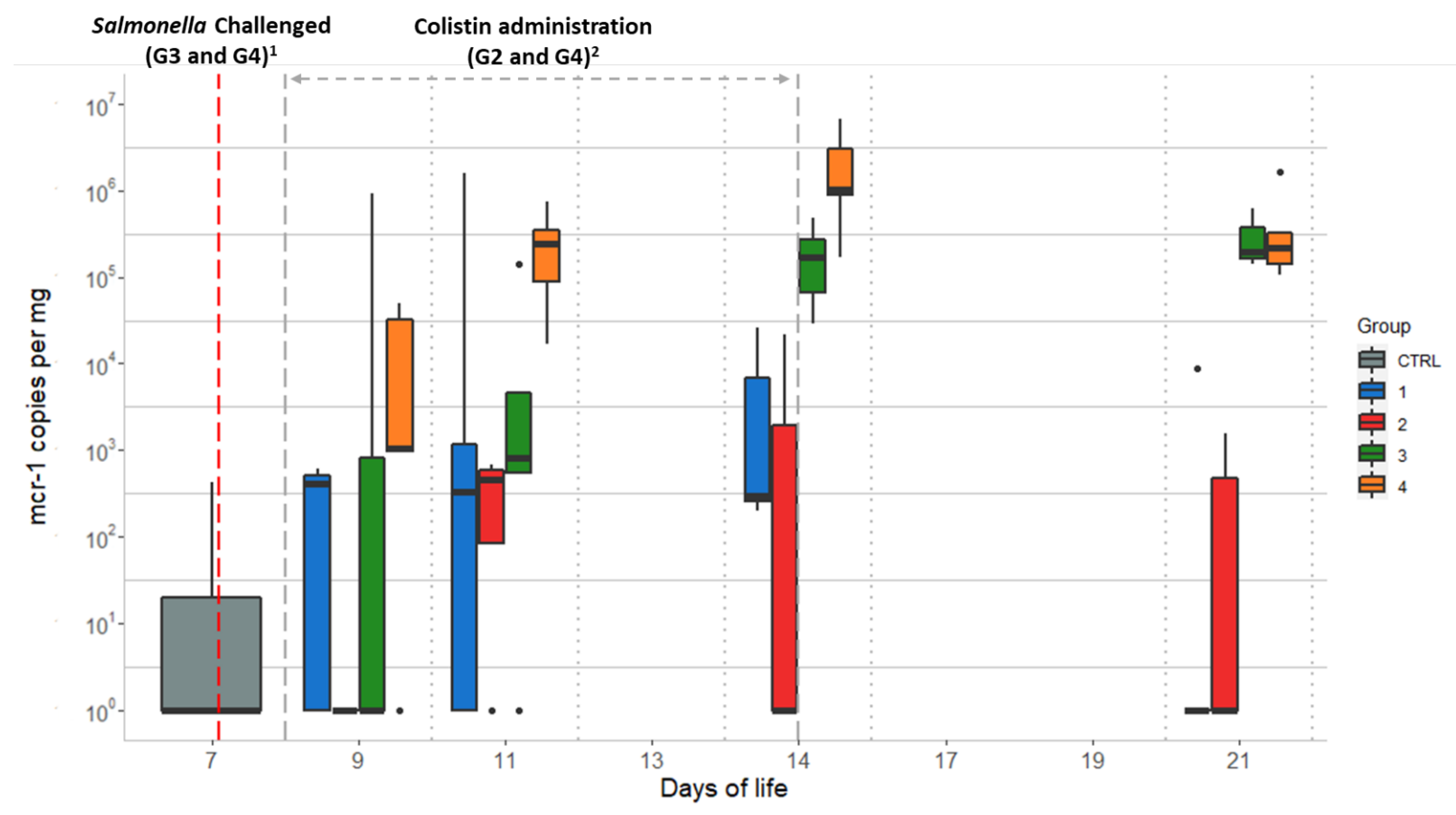
2.3. Quantitative qPCR of the mcr-1 Gene
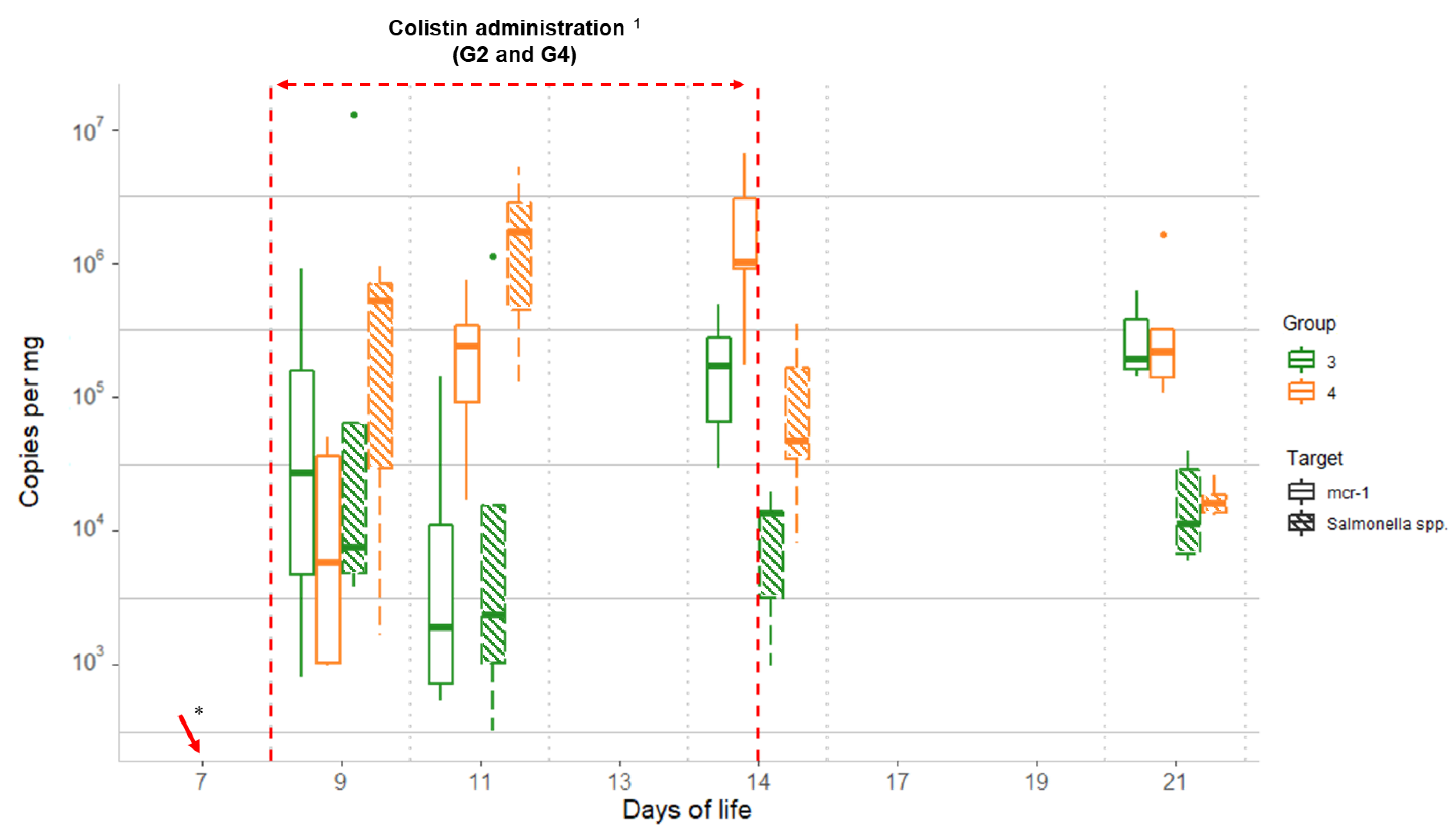
2.4. Comparison of Salmonella Spp. and mcr-1 Quantitative qPCR Data
3. Discussion
4. Methods
4.1. Ethical Approval
4.2. Experimental Design
4.3. Sampling and Sample Preparation
Salmonella Counting Using Selective Media
4.4. Quantitative Assay for Mcr-1
4.5. Quantitative Real-Time PCR Assay for Salmonella
4.6. Statistical Analysis
4.7. Data Visualisation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of Commensal Microflora by Toll-Like Receptors Is Required for Intestinal Homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crhanova, M.; Hradecka, H.; Faldynova, M.; Matulova, M.; Havlickova, H.; Sisak, F.; Rychlik, I. Immune response of chicken gut to natural colonization by gut microflora and to Salmonella enterica serovar enteritidis infection. Infect. Immun. 2011, 79, 2755–2763. [Google Scholar] [CrossRef] [Green Version]
- Litvak, Y.; Mon, K.K.Z.; Nguyen, H.; Chanthavixay, G.; Liou, M.; Velazquez, E.M.; Kutter, L.; Alcantara, M.A.; Byndloss, M.X.; Tiffany, C.R.; et al. Commensal Enterobacteriaceae Protect against Salmonella Colonization through Oxygen Competition. Cell Host Microbe 2019, 25, 128–139.e125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, M.O.A.; Dantas, G.; Church, G.M. Functional Characterization of the Antibiotic Resistance Reservoir in the Human Microflora. Science 2009, 325, 1128–1131. [Google Scholar] [CrossRef] [Green Version]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Lopez Navas, A.; Mackay, D.; et al. Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Wang, Y.; Zhang, S.; Shen, Z.; Wang, S. Emergence of the colistin resistance gene mcr-1 and its variant in several uncommon species of Enterobacteriaceae from commercial poultry farm surrounding environments. Vet. Microbiol. 2018, 219, 161–164. [Google Scholar] [CrossRef]
- Lima, T.; Domingues, S.; Da Silva, G.J. Plasmid-Mediated Colistin Resistance in Salmonella enterica: A Review. Microorganisms 2019, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Model Lists of Essential Medicines. Available online: https://www.who.int/groups/expert-committee-on-selection-and-use-of-essential-medicines/essential-medicines-lists (accessed on 26 April 2021).
- Poirel, L.; Madec, J.Y.; Lupo, A.; Schink, A.K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar] [CrossRef]
- Agency, E.M. Updated Advice on the Use of Colistin Products in Animals within the European Union: Development of Resistance and Possible Impact on Human and Animal Health. Available online: https://www.ema.europa.eu/documents/scientific-guideline/updated-advice-use-colistin-products-animals-within-european-union-development-resistance-possible_en-0.pdf (accessed on 26 April 2021).
- EMA/CVMP. Article 35. Community Interest Referral: Initiated in Cases Involving the Interests of the Community or Concerns Relating to the Protection of Human or Animal Health or the Environment. [European Commission Final Decision]. 2015. Available online: https://www.ema.europa.eu/en/documents/referral/colistin-oral-article-35-referral-annex-i-ii-iii_en.pdf (accessed on 26 April 2021).
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Miguela-Villoldo, P.; Hernández, M.; Moreno, M.A.; Rodríguez-Lázaro, D.; Quesada, A.; Domínguez, L.; Ugarte-Ruiz, M. National colistin sales versus colistin resistance in Spanish pig production. Res. Vet. Sci. 2019, 123, 141–143. [Google Scholar] [CrossRef]
- Poirel, L.; Kieffer, N.; Nordmann, P. In Vitro Study of ISApl1-Mediated Mobilization of the Colistin Resistance Gene mcr-1. Antimicrob. Agents Chemother. 2017, 61, e00127-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zeng, X.; Li, X.-P.; Liao, X.-P.; Liu, Y.-H.; Lin, J. Plasmid-mediated colistin resistance in animals: Current status and future directions. Anim. Health Res. Rev. 2017, 18, 136–152. [Google Scholar] [CrossRef]
- Sadler, W.W.; Brownell, J.R.; Fanelli, M.J. Influence of Age and Inoculum Level on Shed Pattern of Salmonella typhimurium in Chickens. Avian Dis. 1969, 13, 793–803. [Google Scholar] [CrossRef]
- Skov, R.L.; Monnet, D.L. Plasmid-mediated colistin resistance (mcr-1 gene): Three months later, the story unfolds. Euro Surveill. Eur. Commun. Dis. Bull. 2016, 21, 30155. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Hansen, C.; Dahlkilde, A.L.; Herrero-Fresno, A.; Pedersen, K.S.; Nielsen, J.P.; Olsen, J.E. The Effect of Colistin Treatment on the Selection of Colistin-Resistant Escherichia coli in Weaner Pigs. Antibiotics 2021, 10, 465. [Google Scholar] [CrossRef] [PubMed]
- Nang, S.C.; Morris, F.C.; McDonald, M.J.; Han, M.L.; Wang, J.; Strugnell, R.A.; Velkov, T.; Li, J. Fitness cost of mcr-1-mediated polymyxin resistance in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1604–1610. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, J.; Zhang, C.; Li, R.; Wai-Chi Chan, E.; Wu, C.; Wu, C.; Chen, S. Distinct mechanisms of acquisition of mcr-1 -bearing plasmid by Salmonella strains recovered from animals and food samples. Sci. Rep. 2017, 7, 13199. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Encinas, J.; Blanch, M.; Pastor, J.J.; Mereu, A.; Ipharraguerre, I.R.; Menoyo, D. Effects of a bioactive olive pomace extract from Olea europaea on growth performance, gut function, and intestinal microbiota in broiler chickens. Poultry Sci. 2020, 99, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill. Eur. Commun. Dis. Bull. 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, X.; Yin, W.; Wang, Y.; Shen, Z.; Ding, S.; Wang, S. A Multiplex SYBR Green Real-Time PCR Assay for the Detection of Three Colistin Resistance Genes from Cultured Bacteria, Feces, and Environment Samples. Front. Microbiol. 2017, 8, 2078. [Google Scholar] [CrossRef] [PubMed]
- Miguela-Villoldo, P.; Moreno, M.A.; Hernández, M.; Rodríguez-Lázaro, D.; Gallardo, A.; Borge, C.; Quesada, A.; Domínguez, L.; Ugarte-Ruiz, M. Complementarity of Selective Culture and qPCR for Colistin Resistance Screening in Fresh and Frozen Pig Cecum Samples. Front. Microbiol. 2020, 11, 572712. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 26 April 2021).
- Wickham, H. ggplot2; Use R! Springer International Publishing; Available online: https://0-link-springer-com.brum.beds.ac.uk/book/10.1007%2F978-3-319-24277-4 (accessed on 26 April 2021).




{kind=link}
{kind=link}
{kind=link}
| mcr-1 Gene Copies/mg (Expressed in Log10) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SC (D7) | CA (D8) | Day 7 1 | Day 9 | Day 11 | Day 14 | Day 21 | ||||||
| Me | Me | Me | Me | Me | ||||||||
| CTRL 2 | No | No | 2.07 | <2.0 | NA | NA | NA | NA | NA | NA | NA | NA |
| G1 | No | No | NA | NA | 2.49 | 2.60 | 5.51 | 2.52 | 3.83 | 2.47 | 3.24 | <2.0 |
| G2 | No | Yes | NA | NA | <2.0 | <2.0 | 6.43 | 2.75 | 3.67 | <2.0 | 2.60 | <2.0 |
| G3 | Yes | No | NA | NA | 5.26 | <2.0 | 4.47 | 2.90 | 5.32 | 5.24 | 5.47 | 5.29 |
| G4 | Yes | Yes | NA | NA | 4.23 | 3.02 | 5.46 | 5.38 | 6.37 | 6.01 | 5.68 | 5.34 |
| p-Values | ||||||
|---|---|---|---|---|---|---|
| qPCR Data | Group | D7 | D9 | D11 | D14 | D21 |
| mcr-1 | G3 vs. G4 | ND | 0.41 | 0.03 * | 0.02 * | 0.86 |
| Salmonella | G3 vs. G4 | ND | 0.69 | 0.02 * | 0.03 * | 0.69 |
| mcr-1 vs. Salmonella | G3 | ND | 0.06 | 0.44 * | <0.01 * | <0.01 * |
| mcr-1 vs. Salmonella | G4 | ND | 0.94 | <0.01 * | <0.01 * | <0.01 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miguela-Villoldo, P.; Moreno, M.A.; Rebollada-Merino, A.; Rodríguez-Bertos, A.; Hernández, M.; Rodríguez-Lázaro, D.; Gallardo, A.; Quesada, A.; Goyache, J.; Domínguez, L.; et al. Colistin Selection of the Mcr-1 Gene in Broiler Chicken Intestinal Microbiota. Antibiotics 2021, 10, 677. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060677
Miguela-Villoldo P, Moreno MA, Rebollada-Merino A, Rodríguez-Bertos A, Hernández M, Rodríguez-Lázaro D, Gallardo A, Quesada A, Goyache J, Domínguez L, et al. Colistin Selection of the Mcr-1 Gene in Broiler Chicken Intestinal Microbiota. Antibiotics. 2021; 10(6):677. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060677
Chicago/Turabian StyleMiguela-Villoldo, Pedro, Miguel A. Moreno, Agustín Rebollada-Merino, Antonio Rodríguez-Bertos, Marta Hernández, David Rodríguez-Lázaro, Alejandro Gallardo, Alberto Quesada, Joaquín Goyache, Lucas Domínguez, and et al. 2021. "Colistin Selection of the Mcr-1 Gene in Broiler Chicken Intestinal Microbiota" Antibiotics 10, no. 6: 677. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060677