
The Class A β-Lactamase Produced by Burkholderia Species Compromises the Potency of Tebipenem against a Panel of Isolates from the United States


Abstract
:1. Introduction
2. Results
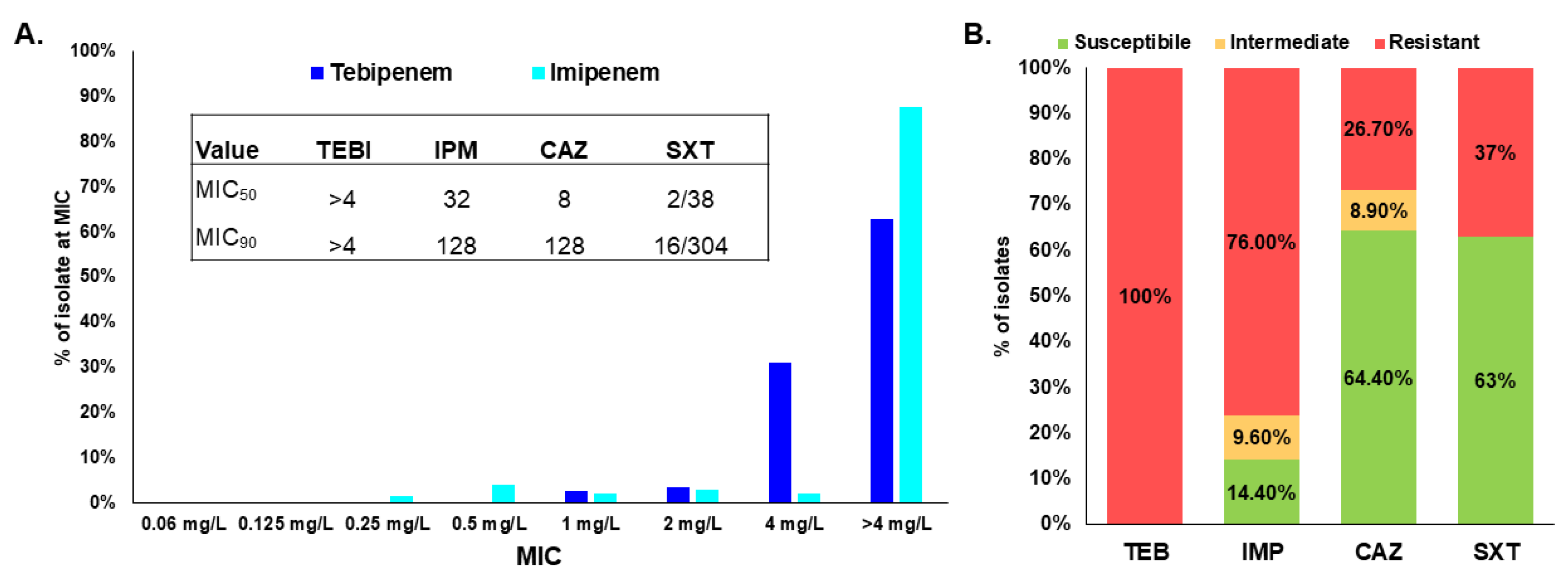
2.1. Tebipenem Does Not Demonstrate Clinically Relevant Antimicrobial Activity against Bcc and B. gladioli
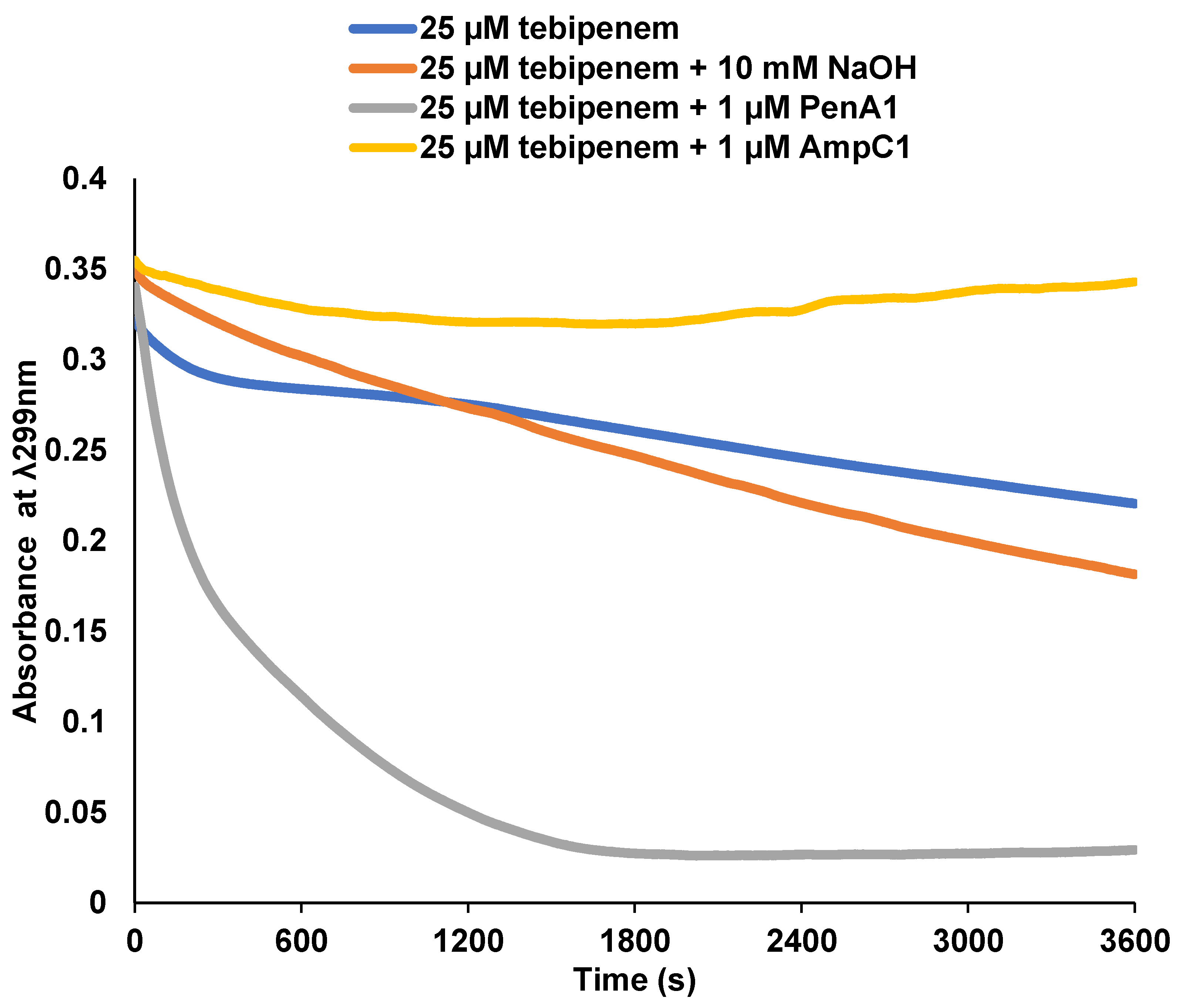
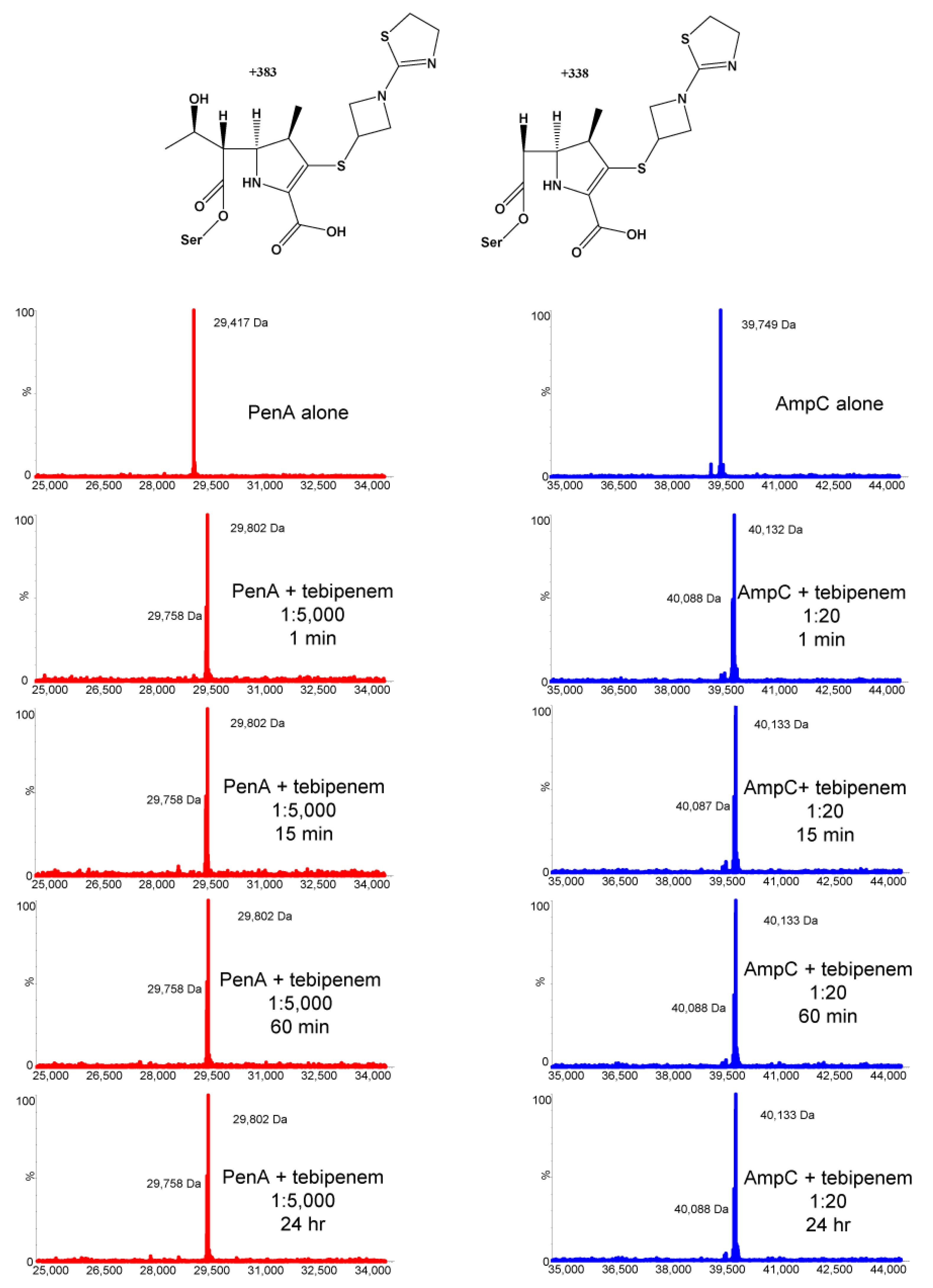
2.2. Tebipenem Is Slowly Hydrolyzed by PenA1 but Inhibits AmpC1, but Eventually Forms a Stable Complex with Both β-lactamases
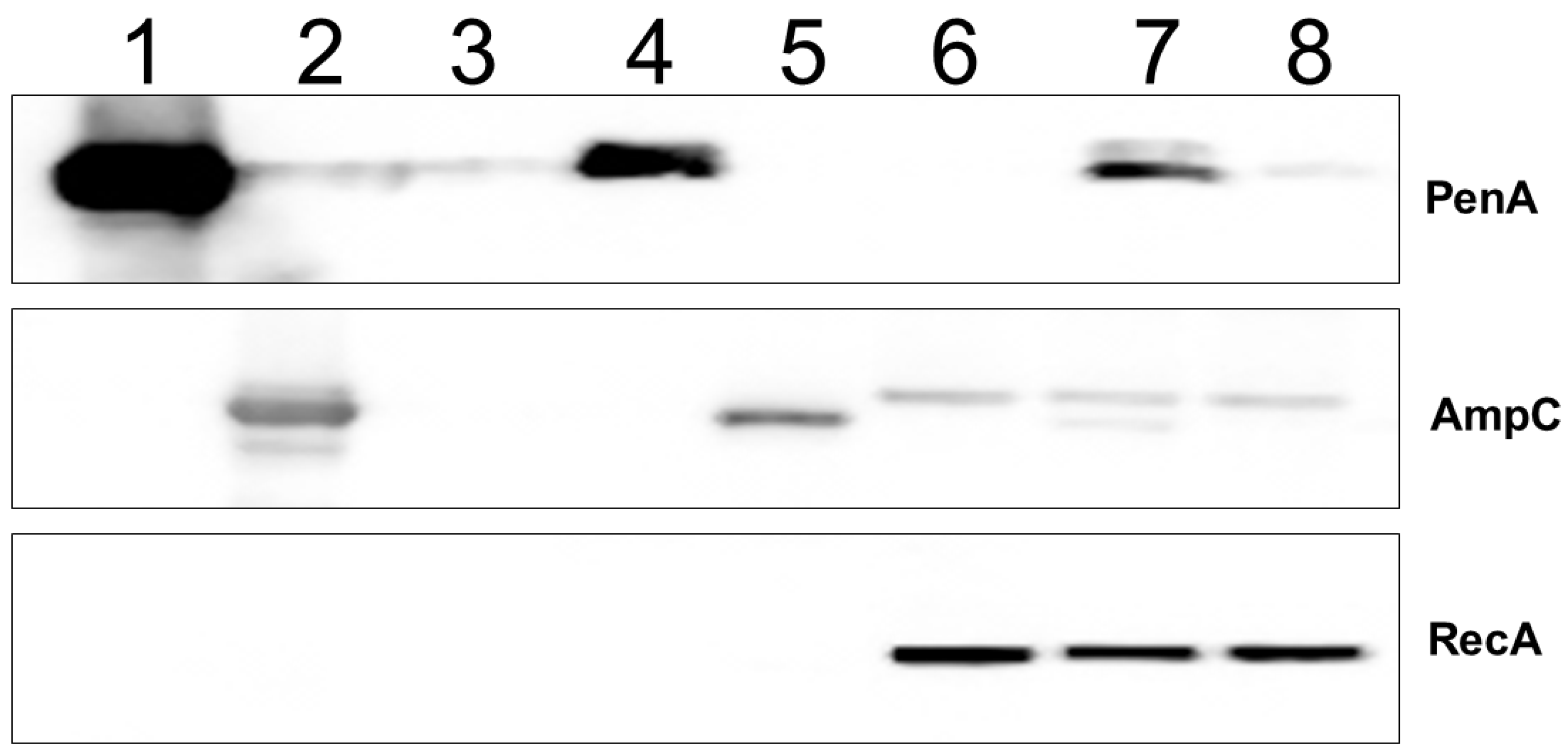
2.3. Tebipenem Is a Minor Inducer of blaPenA1 Expression
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hikida, M.; Itahashi, K.; Igarashi, A.; Shiba, T.; Kitamura, M. In vitro antibacterial activity of LJC 11,036, an active metabolite of L-084, a new oral carbapenem antibiotic with potent antipneumococcal activity. Antimicrob. Agents Chemother. 1999, 43, 2010–2016. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, R.; Konomi, M.; Hasegawa, K.; Morozumi, M.; Sunakawa, K.; Ubukata, K. In vitro activity of tebipenem, a new oral carbapenem antibiotic, against penicillin-nonsusceptible Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2005, 49, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Utley, L.; Parr, T.R.; Zabawa, T.; Pucci, M.J. Tebipenem, the first oral carbapenem antibiotic. Expert Rev. Anti Infect. Ther. 2018, 16, 513–522. [Google Scholar] [CrossRef]
- Hasegawa, K.; Chiba, N.; Kobayashi, R.; Murayama, S.Y.; Iwata, S.; Sunakawa, K.; Ubukata, K. Rapidly increasing prevalence of β-lactamase-nonproducing, ampicillin-resistant Haemophilus influenzae type b in patients with meningitis. Antimicrob. Agents Chemother. 2004, 48, 1509–1514. [Google Scholar] [CrossRef] [Green Version]
- Kuroki, H.; Tateno, N.; Ikeda, H.; Saito, N. Investigation of pneumonia-causing pathogenic organisms in children and the usefulness of tebipenem pivoxil for their treatment. J. Infect. Chemother. 2010, 16, 280–287. [Google Scholar] [CrossRef]
- McEntee, L.; Johnson, A.; Farrington, N.; Unsworth, J.; Dane, A.; Jain, A.; Cotroneo, N.; Critchley, I.; Melnick, D.; Parr, T.; et al. Pharmacodynamics of tebipenem: New options for oral treatment of multidrug-resistant Gram-negative infections. Antimicrob. Agents Chemother. 2019, 63, e00603-19. [Google Scholar] [CrossRef] [Green Version]
- Thamlikitkul, V.; Lorchirachoonkul, N.; Tiengrim, S. In vitro and in vivo activity of tebipenem against ESBL-producing E. coli. J. Med. Assoc. Thail. 2014, 97, 1259–1268. [Google Scholar]
- Yao, Q.; Wang, J.; Cui, T.; Yang, Z.; Su, M.; Zhao, P.; Yan, H.; Zhan, Y.; Yang, H. Antibacterial properties of tebipenem pivoxil tablet, a new rral carbapenem preparation against a variety of pathogenic bacteria in vitro and in vivo. Molecules 2016, 21, 62. [Google Scholar] [CrossRef] [Green Version]
- Rubio, A.; Pucci, M.J.; Jain, A. Characterization of SPR994, an orally available carbapenem, with activity comparable to intravenously administered carbapenems. ACS Infect. Dis. 2018, 4, 1436–1438. [Google Scholar] [CrossRef]
- Arends, S.J.R.; Rhomberg, P.R.; Cotroneo, N.; Rubio, A.; Flamm, R.K.; Mendes, R.E. Antimicrobial activity evaluation of tebipenem (SPR859), an orally available carbapenem, against a global set of Enterobacteriaceae isolates, including a challenge set of organisms. Antimicrob. Agents Chemother. 2019, 63, e02618-18. [Google Scholar] [CrossRef] [Green Version]
- Cotroneo, N.; Rubio, A.; Critchley, I.A.; Pillar, C.; Pucci, M.J. In vitro and in vivo characterization of tebipenem, an oral carbapenem. Antimicrob. Agents Chemother. 2020, 64, e02240-19. [Google Scholar] [CrossRef]
- Lacasse, E.; Brouillette, E.; Larose, A.; Parr, T.R.; Rubio, A., Jr.; Malouin, F. In vitro activity of tebipenem (SPR859) against penicillin-binding proteins of Gram-negative and Gram-positive bacteria. Antimicrob. Agents Chemother. 2019, 63, e02181-18. [Google Scholar] [CrossRef] [Green Version]
- Sodhi, V.; Kronsberg, K.A.; Clark, M.; Cho, J.C. Tebipenem pivoxil hydrobromide-No PICC, no problem! Pharmacotherapy 2021, 41, 748–761. [Google Scholar] [CrossRef]
- Seenama, C.; Tiengrim, S.; Thamlikitkul, V. In vitro activity of tebipenem against Burkholderia pseudomallei. Int. J. Antimicrob. Agents 2013, 42, 375. [Google Scholar] [CrossRef]
- Clayton, N.P.; Jain, A.; Halasohoris, S.A.; Pysz, L.M.; Lembirik, S.; Zumbrun, S.D.; Kane, C.D.; Hackett, M.J.; Pfefferle, D.; Smiley, M.A.; et al. In vitro and in vivo characterization of tebipenem (TBP), an orally active carbapenem, against biothreat pathogens. Antimicrob. Agents Chemother. 2021, 65, e02385-20. [Google Scholar] [CrossRef]
- Marson, F.A.; Hortencio, T.D.; Aguiar, K.C.; Ribeiro, J.D. Demographic, clinical, and laboratory parameters of cystic fibrosis during the last two decades: A comparative analysis. BMC Pulm. Med. 2015, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Abbott, I.J.; Peleg, A.Y. Stenotrophomonas, Achromobacter, and nonmelioid Burkholderia species: Antimicrobial resistance and therapeutic strategies. Semin. Respir. Crit. Care Med. 2015, 36, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Hanulik, V.; Webber, M.A.; Chroma, M.; Uvizl, R.; Holy, O.; Whitehead, R.N.; Baugh, S.; Matouskova, I.; Kolar, M. An outbreak of Burkholderia multivorans beyond cystic fibrosis patients. J. Hosp. Infect. 2013, 84, 248–251. [Google Scholar] [CrossRef]
- Chiappini, E.; Taccetti, G.; de Martino, M. Bacterial lung infections in cystic fibrosis patients: An update. Pediatr. Infect. Dis. J. 2014, 33, 653–654. [Google Scholar] [CrossRef]
- Gautam, V.; Singhal, L.; Ray, P. Burkholderia cepacia complex: Beyond Pseudomonas and Acinetobacter. Indian J. Med. Microbiol. 2011, 29, 4–12. [Google Scholar] [CrossRef]
- Avgeri, S.G.; Matthaiou, D.K.; Dimopoulos, G.; Grammatikos, A.P.; Falagas, M.E. Therapeutic options for Burkholderia cepacia infections beyond co-trimoxazole: A systematic review of the clinical evidence. Int. J. Antimicrob. Agents 2009, 33, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Wuthiekanun, V.; Peacock, S.J. Management of melioidosis. Expert Rev. Anti Infect. Ther. 2006, 4, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Becka, S.A.; Zeiser, E.T.; Ohuchi, N.; Mojica, M.F.; Gatta, J.A.; Falleni, M.; Tosi, D.; Borghi, E.; Winkler, M.L.; et al. Overcoming an extremely drug resistant (XDR) pathogen: Avibactam restores susceptibility to ceftazidime for Burkholderia cepacia complex isolates from Cystic Fibrosis patients. ACS Infect. Dis. 2017, 3, 502–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dalem, A.; Herpol, M.; Echahidi, F.; Peeters, C.; Wybo, I.; De Wachter, E.; Vandamme, P.; Pierard, D. In vitro susceptibility of Burkholderia cepacia complex isolated from Cystic Fibrosis patients to ceftazidime-avibactam and ceftolozane-tazobactam. Antimicrob. Agents Chemother. 2018, 62, e00590-18. [Google Scholar] [CrossRef] [Green Version]
- Cheung, T.K.; Ho, P.L.; Woo, P.C.; Yuen, K.Y.; Chau, P.Y. Cloning and expression of class A β-lactamase gene blaA(BPS) in Burkholderia pseudomallei. Antimicrob. Agents Chemother. 2002, 46, 1132–1135. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, A.J.; Wong, S.; Dance, D.A.; Chaowagul, W.; Bryan, L.E. Pseudomonas pseudomallei resistance to β-lactam antibiotics due to alterations in the chromosomally encoded β-lactamase. Antimicrob. Agents Chemother. 1991, 35, 1635–1640. [Google Scholar] [CrossRef] [Green Version]
- Tribuddharat, C.; Moore, R.A.; Baker, P.; Woods, D.E. Burkholderia pseudomallei class A β-lactamase mutations that confer selective resistance against ceftazidime or clavulanic acid inhibition. Antimicrob. Agents Chemother. 2003, 47, 2082–2087. [Google Scholar] [CrossRef] [Green Version]
- Trepanier, S.; Prince, A.; Huletsky, A. Characterization of the penA and penR genes of Burkholderia cepacia 249 which encode the chromosomal class A penicillinase and its LysR-type transcriptional regulator. Antimicrob. Agents Chemother. 1997, 41, 2399–2405. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Rodriguez-Martinez, J.M.; Plesiat, P.; Nordmann, P. Naturally occurring class A β-lactamases from the Burkholderia cepacia complex. Antimicrob. Agents Chemother. 2009, 53, 876–882. [Google Scholar] [CrossRef] [Green Version]
- Becka, S.A.; Zeiser, E.T.; Marshall, S.H.; Gatta, J.A.; Nguyen, K.; Singh, I.; Greco, C.; Sutton, G.G.; Fouts, D.E.; LiPuma, J.J.; et al. Sequence heterogeneity of the PenA carbapenemase in clinical isolates of Burkholderia multivorans. Diagn. Microbiol. Infect. Dis. 2018, 92, 253–258. [Google Scholar] [CrossRef]
- Becka, S.A.; Zeiser, E.T.; Barnes, M.D.; Taracila, M.A.; Nguyen, K.; Singh, I.; Sutton, G.G.; LiPuma, J.J.; Fouts, D.E.; Papp-Wallace, K.M. Characterization of the AmpC β-lactamase from Burkholderia multivorans. Antimicrob. Agents Chemother. 2018, 62, e01140-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp-Wallace, K.M.; Taracila, M.A.; Gatta, J.A.; Ohuchi, N.; Bonomo, R.A.; Nukaga, M. Insights into β-lactamases from Burkholderia species, two phylogenetically related yet distinct resistance determinants. J. Biol. Chem. 2013, 288, 19090–19102. [Google Scholar] [CrossRef] [Green Version]
- Dhar, S.; Kumari, H.; Balasubramanian, D.; Mathee, K. Cell-wall recycling and synthesis in Escherichia coli and Pseudomonas aeruginosa-their role in the development of resistance. J. Med. Microbiol. 2018, 67, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Shapiro, A.B.; Becka, S.A.; Zeiser, E.T.; LiPuma, J.J.; Lane, D.J.; Panchal, R.G.; Mueller, J.P.; O’Donnell, J.P.; Miller, A.A. In vitro antibacterial activity and in vivo efficacy of sulbactam-durlobactam against pathogenic Burkholderia species. Antimicrob. Agents Chemother. 2021, 65, e01930-20. [Google Scholar] [CrossRef] [PubMed]
- Zeiser, E.T.; Becka, S.A.; Barnes, M.D.; Taracila, M.A.; LiPuma, J.J.; Papp-Wallace, K.M. Resurrecting old β-lactams: Potent inhibitory activity of temocillin against multidrug-resistant Burkholderia species isolates from the United States. Antimicrob. Agents Chemother. 2019, 63, e02315-18. [Google Scholar] [CrossRef] [Green Version]
- Zeiser, E.T.; Becka, S.A.; Wilson, B.M.; Barnes, M.D.; LiPuma, J.J.; Papp-Wallace, K.M. “Switching partners”: Piperacillin-avibactam is a highly potent xombination against multidrug-eesistant Burkholderia cepacia complex and Burkholderia gladioli Cystic Fibrosis isolates. J. Clin. Microbiol. 2019, 57, e00181-19. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptiblity Testing. CLSI Supplement M100 , 31st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Becka, S.A.; Zeiser, E.T.; LiPuma, J.J.; Papp-Wallace, K.M. Activity of imipenem-relebactam against multidrug- and extensively drug-resistant Burkholderia cepacia complex and Burkholderia gladioli. Antimicrob. Agents Chemother. 2021, 65, e0133221. [Google Scholar] [CrossRef]
- Tremblay, L.W.; Fan, F.; Blanchard, J.S. Biochemical and structural characterization of Mycobacterium tuberculosis beta-lactamase with the carbapenems ertapenem and doripenem. Biochemistry 2010, 49, 3766–3773. [Google Scholar] [CrossRef] [Green Version]
- Endimiani, A.; Doi, Y.; Bethel, C.R.; Taracila, M.; Adams-Haduch, J.M.; O’Keefe, A.; Hujer, A.M.; Paterson, D.L.; Skalweit, M.J.; Page, M.G.; et al. Enhancing resistance to cephalosporins in class C beta-lactamases: Impact of Gly214Glu in CMY-2. Biochemistry 2010, 49, 1014–1023. [Google Scholar] [CrossRef] [Green Version]
- Drawz, S.M.; Babic, M.; Bethel, C.R.; Taracila, M.; Distler, A.M.; Ori, C.; Caselli, E.; Prati, F.; Bonomo, R.A. Inhibition of the class C beta-lactamase from Acinetobacter spp.: Insights into effective inhibitor design. Biochemistry 2010, 49, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Hugonnet, J.E.; Tremblay, L.W.; Boshoff, H.I.; Barry, C.E.; Blanchard, J.S., 3rd. Meropenem-clavulanate is effective against extensively drug-resistant Mycobacterium tuberculosis. Science 2009, 323, 1215–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; Spilker, T.; Martin, A.; LiPuma, J.J. Comparative assessment of genotyping methods for epidemiologic study of Burkholderia cepacia genomovar III. J. Clin. Microbiol. 2002, 40, 3300–3307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehmann, D.E.; Jahic, H.; Ross, P.L.; Gu, R.F.; Hu, J.; Durand-Reville, T.F.; Lahiri, S.; Thresher, J.; Livchak, S.; Gao, N.; et al. Kinetics of avibactam inhibition against Class A, C, and D β-lactamases. J. Biol. Chem. 2013, 288, 27960–27971. [Google Scholar] [CrossRef] [Green Version]
- Papp-Wallace, K.M.; Winkler, M.L.; Gatta, J.A.; Taracila, M.A.; Chilakala, S.; Xu, Y.; Johnson, J.K.; Bonomo, R.A. Reclaiming the efficacy of β-lactam-β-lactamase inhibitor combinations: Avibactam restores the susceptibility of CMY-2-producing Escherichia coli to ceftazidime. Antimicrob. Agents Chemother. 2014, 58, 4290–4297. [Google Scholar] [CrossRef] [Green Version]
- Papp-Wallace, K.M.; Becka, S.A.; Taracila, M.A.; Winkler, M.L.; Gatta, J.A.; Rholl, D.A.; Schweizer, H.P.; Bonomo, R.A. Exposing a β-lactamase “twist”: The mechanistic basis for the high level of ceftazidime resistance in the C69F variant of the Burkholderia pseudomallei PenI β-lactamase. Antimicrob. Agents Chemother. 2016, 60, 777–788. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Strain | Tebipenem |
|---|---|
| B. multivorans ATCC 17616 | >4 |
| E. coli ATCC 25922 | ≤0.06 |
| P. aeruginosa ATCC 27853 | >4 |
| Parameter | PenA1 | AmpC1 |
|---|---|---|
| Ki app (µM) | 4.7 ± 0.5 | 22 ± 2 |
| k2/K (M−1s−1) | N/D | 1.9 ± 0.1 × 103 |
| koff (s−1) | N/D | 3 ± 1 × 10−4 |
| tn at 15 min | 4000 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becka, S.A.; Zeiser, E.T.; LiPuma, J.J.; Papp-Wallace, K.M. The Class A β-Lactamase Produced by Burkholderia Species Compromises the Potency of Tebipenem against a Panel of Isolates from the United States. Antibiotics 2022, 11, 674. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050674
Becka SA, Zeiser ET, LiPuma JJ, Papp-Wallace KM. The Class A β-Lactamase Produced by Burkholderia Species Compromises the Potency of Tebipenem against a Panel of Isolates from the United States. Antibiotics. 2022; 11(5):674. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050674
Chicago/Turabian StyleBecka, Scott A., Elise T. Zeiser, John J. LiPuma, and Krisztina M. Papp-Wallace. 2022. "The Class A β-Lactamase Produced by Burkholderia Species Compromises the Potency of Tebipenem against a Panel of Isolates from the United States" Antibiotics 11, no. 5: 674. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050674