
Phenotypic Adaptation to Antiseptics and Effects on Biofilm Formation Capacity and Antibiotic Resistance in Clinical Isolates of Early Colonizers in Dental Plaque

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Test Substances
2.3. Minimal Inhibitory Concentration (MIC) Passaging and Re-Evaluation of Phenotypic Adaptation
2.4. Biofilm Formation Capacity
2.5. Phenotypic Antibiotic Resistance Testing
2.6. Determination of the Presence of Antibiotic Resistance Genes (ARGs) by PCR
3. Results
3.1. MIC Passaging and Re-Evaluation of Phenotypic Adaptation
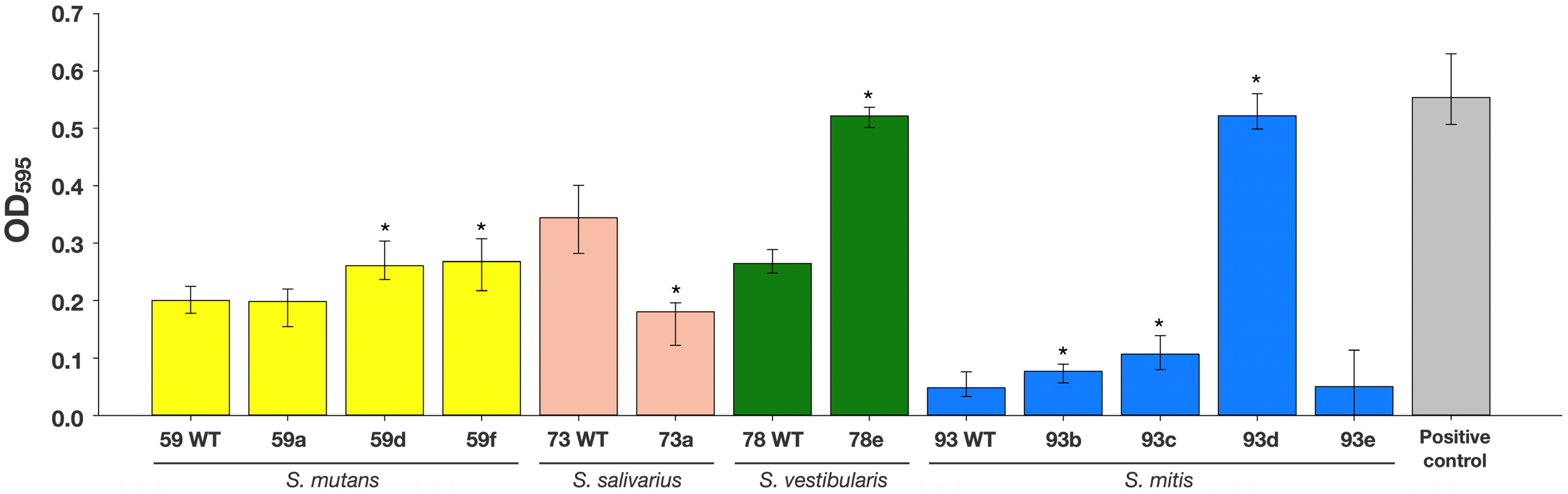
3.2. Biofilm Formation Capacity
3.3. Phenotypic Antibiotic Resistance
3.4. Presence of ARGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tacconelli, E.; Pezzani, M.D. Public health burden of antimicrobial resistance in Europe. Lancet Infect. Dis. 2019, 19, 4–6. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Wellcome Trust: London, UK; HM Government: London, UK, 2016.
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Laxminarayan, R. The overlooked pandemic of antimicrobial resistance. Lancet 2022, 399, 606–607. [Google Scholar] [CrossRef]
- Lu, J.; Guo, J. Disinfection spreads antimicrobial resistance. Science 2021, 371, 474. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Tagkopoulos, I. Benzalkonium Chlorides: Uses, Regulatory Status, and Microbial Resistance. Appl. Environ. Microbiol. 2019, 85, e0037-19. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Auer, D.L.; Buchalla, W.; Hiller, K.-A.; Maisch, T.; Hellwig, E.; Al-Ahmad, A.; Cieplik, F. Cetylpyridinium Chloride: Mechanism of Action, Antimicrobial Efficacy in Biofilms, and Potential Risks of Resistance. Antimicrob. Agents Chemother. 2020, 64, e00576-20. [Google Scholar] [CrossRef]
- Cieplik, F.; Jakubovics, N.S.; Buchalla, W.; Maisch, T.; Hellwig, E.; Al-Ahmad, A. Resistance Toward Chlorhexidine in Oral Bacteria—Is There Cause for Concern? Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Mechanisms of Increased Resistance to Chlorhexidine and Cross-Resistance to Colistin following Exposure of Klebsiella pneumoniae Clinical Isolates to Chlorhexidine. Antimicrob. Agents Chemother. 2017, 61, e01162-16. [Google Scholar] [CrossRef] [Green Version]
- Laumen, J.G.E.; van Dijck, C.; Manoharan-Basil, S.S.; Abdellati, S.; de Baetselier, I.; Cuylaerts, V.; de Block, T.; van den Bossche, D.; Xavier, B.B.; Malhotra-Kumar, S.; et al. Sub-Inhibitory Concentrations of Chlorhexidine Induce Resistance to Chlorhexidine and Decrease Antibiotic Susceptibility in Neisseria gonorrhoeae. Front. Microbiol. 2021, 12, 776909. [Google Scholar] [CrossRef]
- Roberts, A.P.; Mullany, P. Oral biofilms: A reservoir of transferable, bacterial, antimicrobial resistance. Expert Rev. Anti-Infect. Ther. 2010, 8, 1441–1450. [Google Scholar] [CrossRef]
- Jiang, S.; Zeng, J.; Zhou, X.; Li, Y. Drug Resistance and Gene Transfer Mechanisms in Respiratory/Oral Bacteria. J. Dent. Res. 2018, 97, 1092–1099. [Google Scholar] [CrossRef]
- Arredondo, A.; Blanc, V.; Mor, C.; Nart, J.; León, R. Tetracycline and multidrug resistance in the oral microbiota: Differences between healthy subjects and patients with periodontitis in Spain. J. Oral Microbiol. 2020, 13, 1847431. [Google Scholar] [CrossRef]
- Haps, S.; Slot, D.E.; Berchier, C.E.; van der Weijden, G.A. The effect of cetylpyridinium chloride-containing mouth rinses as adjuncts to toothbrushing on plaque and parameters of gingival inflammation: A systematic review. Int. J. Dent. Hyg. 2008, 6, 290–303. [Google Scholar] [CrossRef]
- Figuero, E.; Herrera, D.; Tobías, A.; Serrano, J.; Roldán, S.; Escribano, M.; Martín, C. Efficacy of adjunctive anti-plaque chemical agents in managing gingivitis: A systematic review and network meta-analyses. J. Clin. Periodontol. 2019, 46, 723–739. [Google Scholar] [CrossRef]
- Sanz, M.; Serrano, J.; Iniesta, M.; Santa Cruz, I.; Herrera, D. Antiplaque and antigingivitis toothpastes. Monogr. Oral Sci. 2013, 23, 27–44. [Google Scholar] [CrossRef]
- Cieplik, F.; Kara, E.; Muehler, D.; Enax, J.; Hiller, K.-A.; Maisch, T.; Buchalla, W. Antimicrobial efficacy of alternative compounds for use in oral care toward biofilms from caries-associated bacteria in vitro. Microbiologyopen 2019, 8, e00695. [Google Scholar] [CrossRef] [Green Version]
- Mohd-Said, S.; Mohd-Dom, T.N.; Suhaimi, N.; Rani, H.; McGrath, C. Effectiveness of Pre-procedural Mouth Rinses in Reducing Aerosol Contamination During Periodontal Prophylaxis: A Systematic Review. Front. Med. 2021, 8, 600769. [Google Scholar] [CrossRef]
- Gottsauner, M.J.; Michaelides, I.; Schmidt, B.; Scholz, K.J.; Buchalla, W.; Widbiller, M.; Hitzenbichler, F.; Ettl, T.; Reichert, T.E.; Bohr, C.; et al. A prospective clinical pilot study on the effects of a hydrogen peroxide mouthrinse on the intraoral viral load of SARS-CoV-2. Clin. Oral Investig. 2020, 24, 3707–3713. [Google Scholar] [CrossRef]
- Meister, T.L.; Gottsauner, J.-M.; Schmidt, B.; Heinen, N.; Todt, D.; Audebert, F.; Buder, F.; Lang, H.; Gessner, A.; Steinmann, E.; et al. Mouthrinses against SARS-CoV-2—high antiviral effectivity by membrane disruption in vitro translates to mild effects in a randomized placebo-controlled clinical trial. Virus Res. 2022, 316, 198791. [Google Scholar] [CrossRef]
- Waldron, C.; Nunn, J.; Mac Giolla Phadraig, C.; Comiskey, C.; Guerin, S.; Harten, M.T.; Donnelly-Swift, E.; Clarke, M.J. Oral hygiene interventions for people with intellectual disabilities. Cochrane Database Syst. Rev. 2019, 5, CD012628. [Google Scholar] [CrossRef]
- Pithon, M.M.; Sant’Anna, L.I.D.A.; Baião, F.C.S.; Santos, R.L.d.; Da Coqueiro, R.S.; Maia, L.C. Assessment of the effectiveness of mouthwashes in reducing cariogenic biofilm in orthodontic patients: A systematic review. J. Dent. 2015, 43, 297–308. [Google Scholar] [CrossRef]
- Solderer, A.; Kaufmann, M.; Hofer, D.; Wiedemeier, D.; Attin, T.; Schmidlin, P.R. Efficacy of chlorhexidine rinses after periodontal or implant surgery: A systematic review. Clin. Oral Investig. 2019, 23, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Grönbeck Lindén, I.; Hägglin, C.; van Gahnberg, L.; Andersson, P. Factors Affecting Older Persons’ Ability to Manage Oral Hygiene: A Qualitative Study. JDR Clin. Transl. Res. 2017, 2, 223–232. [Google Scholar] [CrossRef]
- Zhao, T.; Wu, X.; Zhang, Q.; Li, C.; Worthington, H.V.; Hua, F. Oral hygiene care for critically ill patients to prevent ventilator-associated pneumonia. Cochrane Database Syst. Rev. 2020, 12, CD008367. [Google Scholar] [CrossRef]
- Emilson, C.G.; Ericson, T.; Heyden, G.; Lilia, J. Effect of chlorhexidine on human oral streptococci. J. Periodont. Res. 1972, 7, 189–191. [Google Scholar] [CrossRef]
- Schiott, C.R.; Löe, H. The sensitivity of oral streptococci to chlorhexidine. J. Periodont. Res. 1972, 7, 192–194. [Google Scholar] [CrossRef]
- Emilson, C.G.; Fornell, J. Effect of toothbrushing with chlorhexidine gel on salivary microflora, oral hygiene, and caries. Eur. J. Oral Sci. 1976, 84, 308–319. [Google Scholar] [CrossRef]
- Schwarz, S.R.; Hirsch, S.; Hiergeist, A.; Kirschneck, C.; Muehler, D.; Hiller, K.-A.; Maisch, T.; Al-Ahmad, A.; Gessner, A.; Buchalla, W.; et al. Limited antimicrobial efficacy of oral care antiseptics in microcosm biofilms and phenotypic adaptation of bacteria upon repeated exposure. Clin. Oral Investig. 2021, 25, 2939–2950. [Google Scholar] [CrossRef]
- Kitagawa, H.; Izutani, N.; Kitagawa, R.; Maezono, H.; Yamaguchi, M.; Imazato, S. Evolution of resistance to cationic biocides in Streptococcus mutans and Enterococcus faecalis. J. Dent. 2016, 47, 18–22. [Google Scholar] [CrossRef]
- Verspecht, T.; Rodriguez Herrero, E.; Khodaparast, L.; Khodaparast, L.; Boon, N.; Bernaerts, K.; Quirynen, M.; Teughels, W. Development of antiseptic adaptation and cross-adapatation in selected oral pathogens in vitro. Sci. Rep. 2019, 9, 8326. [Google Scholar] [CrossRef]
- Fux, C.A.; Shirtliff, M.; Stoodley, P.; Costerton, J.W. Can laboratory reference strains mirror “real-world” pathogenesis? Trends Microbiol. 2005, 13, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Al-Ahmad, A.; Auschill, T.M.; Braun, G.; Hellwig, E.; Arweiler, N.B. Overestimation of Streptococcus mutans prevalence by nested PCR detection of the 16S rRNA gene. J. Med. Microbiol. 2006, 55, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, S.; Karygianni, L.; Filippi, A.; Anderson, A.C.; Zürcher, A.; Hellwig, E.; Vach, K.; Macchiarelli, G.; Al-Ahmad, A. Combining culture and culture-independent methods reveals new microbial composition of halitosis patients’ tongue biofilm. Microbiologyopen 2020, 9, e958. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Wiedenhofer, A.M.; Pietsch, V.; Hiller, K.-A.; Hiergeist, A.; Wagner, A.; Baldaranov, D.; Linker, R.A.; Jantsch, J.; Buchalla, W.; et al. Oral Health, Oral Microbiota, and Incidence of Stroke-Associated Pneumonia-A Prospective Observational Study. Front. Neurol. 2020, 11, 528056. [Google Scholar] [CrossRef]
- Bernardi, S.; Anderson, A.; Macchiarelli, G.; Hellwig, E.; Cieplik, F.; Vach, K.; Al-Ahmad, A. Subinhibitory Antibiotic Concentrations Enhance Biofilm Formation of Clinical Enterococcus faecalis Isolates. Antibiotics 2021, 10, 874. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Ameen, H.; Pelz, K.; Karygianni, L.; Wittmer, A.; Anderson, A.C.; Spitzmüller, B.; Hellwig, E. Antibiotic Resistance and Capacity for Biofilm Formation of Different Bacteria Isolated from Endodontic Infections Associated with Root-filled Teeth. J. Endod. 2014, 40, 223–230. [Google Scholar] [CrossRef]
- Maekawa, S.; Yoshioka, M.; Kumamoto, Y. Proposal of a new scheme for the serological typing of Enterococcus faecalis strains. Microbiol. Immunol. 1992, 36, 671–681. [Google Scholar] [CrossRef]
- Anderson, A.C.; Rothballer, M.; Altenburger, M.J.; Woelber, J.P.; Karygianni, L.; Vach, K.; Hellwig, E.; Al-Ahmad, A. Long-Term Fluctuation of Oral Biofilm Microbiota following Different Dietary Phases. Appl. Environ. Microbiol. 2020, 86, e01421-20. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance Global Action Plan; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Diaz, P.I.; Hoare, A.; Hong, B.-Y. Subgingival Microbiome Shifts and Community Dynamics in Periodontal Diseases. J. Calif. Dent. Assoc. 2016, 44, 421–435. [Google Scholar]
- Kolenbrander, P.E.; Palmer, R.J.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Jakubovics, N.S.; Goodman, S.D.; Mashburn-Warren, L.; Stafford, G.P.; Cieplik, F. The dental plaque biofilm matrix. Periodontol. 2000 2021, 86, 32–56. [Google Scholar] [CrossRef]
- Järvinen, H.; Pienihäkkinen, K.; Huovinen, P.; Tenovuo, J. Susceptibility of Streptococcus mutans and Streptococcus sobrinus to antimicrobial agents after short-term oral chlorhexidine treatments. Eur. J. Oral Sci. 1995, 103, 32–35. [Google Scholar] [CrossRef]
- Järvinen, H.; Tenovuo, J.; Huovinen, P. In vitro susceptibility of Streptococcus mutans to chlorhexidine and six other antimicrobial agents. Antimicrob. Agents Chemother. 1993, 37, 1158–1159. [Google Scholar] [CrossRef] [Green Version]
- So Yeon, L.; Si Young, L. Susceptibility of Oral Streptococci to Chlorhexidine and Cetylpyridinium Chloride. Biocontrol Sci. 2019, 24, 13–21. [Google Scholar] [CrossRef] [Green Version]
- McBain, A.J.; Bartolo, R.G.; Catrenich, C.E.; Charbonneau, D.; Ledder, R.G.; Gilbert, P. Effects of a chlorhexidine gluconate-containing mouthwash on the vitality and antimicrobial susceptibility of in vitro oral bacterial ecosystems. Appl. Environ. Microbiol. 2003, 69, 4770–4776. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, J.R.; Godwin, M.J.; Velsko, I.M.; Richards, V.P.; Burne, R.A. Spontaneously Arising Streptococcus mutans Variants with Reduced Susceptibility to Chlorhexidine Display Genetic Defects and Diminished Fitness. Antimicrob. Agents Chemother. 2019, 63, e00161-19. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.S. Biocide resistance mechanisms. Int. Biodeterior. Biodegrad. 2003, 51, 133–138. [Google Scholar] [CrossRef]
- Fox, L.J.; Kelly, P.P.; Humphreys, G.J.; Waigh, T.A.; Lu, J.R.; McBain, A.J. Assessing the risk of resistance to cationic biocides incorporating realism-based and biophysical approaches. J. Ind. Microbiol. Biotechnol. 2022, 49, kuab074. [Google Scholar] [CrossRef]
- Stewart, P.S. Diffusion in biofilms. J. Bacteriol. 2003, 185, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Ciofu, O.; Moser, C.; Jensen, P.Ø.; Høiby, N. Tolerance and resistance of microbial biofilms. Nat. Rev. Microbiol. 2022. [Google Scholar] [CrossRef]
- Jin, Y.; Guo, Y.; Zhan, Q.; Shang, Y.; Qu, D.; Yu, F. Subinhibitory Concentrations of Mupirocin Stimulate Staphylococcus aureus Biofilm Formation by Upregulating cidA. Antimicrob. Agents Chemother. 2020, 64, e01912-19. [Google Scholar] [CrossRef]
- Gajdács, M.; Baráth, Z.; Kárpáti, K.; Szabó, D.; Usai, D.; Zanetti, S.; Donadu, M.G. No Correlation between Biofilm Formation, Virulence Factors, and Antibiotic Resistance in Pseudomonas aeruginosa: Results from a Laboratory-Based In Vitro Study. Antibiotics 2021, 10, 1134. [Google Scholar] [CrossRef]
- Nassar, O.; Desouky, S.E.; El-Sherbiny, G.M.; Abu-Elghait, M. Correlation between phenotypic virulence traits and antibiotic resistance in Pseudomonas aeruginosa clinical isolates. Microb. Pathog. 2022, 162, 105339. [Google Scholar] [CrossRef]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef]
- Muehler, D.; Mao, X.; Czemmel, S.; Geißert, J.; Engesser, C.; Hiller, K.-A.; Widbiller, M.; Maisch, T.; Buchalla, W.; Al-Ahmad, A.; et al. Transcriptomic Stress Response in Streptococcus mutans following Treatment with a Sublethal Concentration of Chlorhexidine Digluconate. Microorganisms 2022, 10, 561. [Google Scholar] [CrossRef]
- Kampf, G. Biocidal Agents Used for Disinfection Can Enhance Antibiotic Resistance in Gram-Negative Species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Doern, G.V.; Ferraro, M.J.; Brueggemann, A.B.; Ruoff, K.L. Emergence of high rates of antimicrobial resistance among viridans group streptococci in the United States. Antimicrob. Agents Chemother. 1996, 40, 891–894. [Google Scholar] [CrossRef] [Green Version]
- Reinert, R.R.; Filimonova, O.Y.; Al-Lahham, A.; Grudinina, S.A.; Ilina, E.N.; Weigel, L.M.; Sidorenko, S.V. Mechanisms of Macrolide Resistance among Streptococcus pneumoniae Isolates from Russia. Antimicrob. Agents Chemother. 2008, 52, 2260–2262. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, L.; Ardanuy, C.; Cercenado, E.; Fenoll, A.; Bouza, E.; Pallares, R.; Martín, R.; Liñares, J. Serotypes, Clones, and Mechanisms of Resistance of Erythromycin-Resistant Streptococcus pneumoniae Isolates Collected in Spain. Antimicrob. Agents Chemother. 2007, 51, 3240–3246. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Recchia, G.D.; Scaramuzzi, C.; Collis, C.M.; Stokes, H.W.; Hall, R.M. Definition of the attI1 site of class 1 integrons. Microbiology 2000, 146, 2855–2864. [Google Scholar] [CrossRef] [Green Version]
- Collis, C.M.; Kim, M.-J.; Partridge, S.R.; Stokes, H.W.; Hall, R.M. Characterization of the class 3 integron and the site-specific recombination system it determines. J. Bacteriol. 2002, 184, 3017–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, A.; Takao, A. β-Lactam resistance in Streptococcus mitis isolated from saliva of healthy subjects. J. Infect. Chemother. 2003, 9, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M.I.; Baylay, A.J.; Wong, R.L.; Piddock, L.J.V. Overexpression of patA and patB, which encode ABC transporters, is associated with fluoroquinolone resistance in clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boncoeur, E.; Durmort, C.; Bernay, B.; Ebel, C.; Di Guilmi, A.M.; Croizé, J.; Vernet, T.; Jault, J.-M. PatA and PatB form a functional heterodimeric ABC multidrug efflux transporter responsible for the resistance of Streptococcus pneumoniae to fluoroquinolones. Biochemistry 2012, 51, 7755–7765. [Google Scholar] [CrossRef]
- Feng, J.; Lupien, A.; Gingras, H.; Wasserscheid, J.; Dewar, K.; Légaré, D.; Ouellette, M. Genome sequencing of linezolid-resistant Streptococcus pneumoniae mutants reveals novel mechanisms of resistance. Genome Res. 2009, 19, 1214–1223. [Google Scholar] [CrossRef] [Green Version]
- Garvey, M.I.; Piddock, L.J.V. The efflux pump inhibitor reserpine selects multidrug-resistant Streptococcus pneumoniae strains that overexpress the ABC transporters PatA and PatB. Antimicrob. Agents Chemother. 2008, 52, 1677–1685. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Hatt, J.K.; Weigand, M.R.; Krishnan, R.; Pavlostathis, S.G.; Konstantinidis, K.T. Genomic and Transcriptomic Insights into How Bacteria Withstand High Concentrations of Benzalkonium Chloride Biocides. Appl. Environ. Microbiol. 2018, 84, e00197-18. [Google Scholar] [CrossRef] [Green Version]
- Thomas, L.; Maillard, J.Y.; Lambert, R.J.; Russell, A.D. Development of resistance to chlorhexidine diacetate in Pseudomonas aeruginosa and the effect of a “residual” concentration. J. Hosp. Infect. 2000, 46, 297–303. [Google Scholar] [CrossRef]
- Rozman, U.; Pušnik, M.; Kmetec, S.; Duh, D.; Šostar Turk, S. Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review. Microorganisms 2021, 9, 2550. [Google Scholar] [CrossRef]
- Lins, R.X.; de Oliveira Andrade, A.; Hirata Junior, R.; Wilson, M.J.; Lewis, M.A.; Williams, D.W.; Fidel, R.A.S. Antimicrobial resistance and virulence traits of Enterococcus faecalis from primary endodontic infections. J. Dent. 2013, 41, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Poyart, C.; Jardy, L.; Quesne, G.; Berche, P.; Trieu-Cuot, P. Genetic Basis of Antibiotic Resistance in Streptococcus agalactiae Strains Isolated in a French Hospital. Antimicrob. Agents Chemother 2003, 47, 794–797. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I.; Garrigues-Jeanjean, N.; Mackie, R.I. Molecular Ecology of Tetracycline Resistance: Development and Validation of Primers for Detection of Tetracycline Resistance Genes Encoding Ribosomal Protection Proteins. Appl. Environ. Microbiol. 2001, 67, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Lanz, R.; Kuhnert, P.; Boerlin, P. Antimicrobial resistance and resistance gene determinants in clinical Escherichia coli from different animal species in Switzerland. Vet. Microbiol. 2003, 91, 73–84. [Google Scholar] [CrossRef]
- Call, D.R.; Bakko, M.K.; Krug, M.J.; Roberts, M.C. Identifying Antimicrobial Resistance Genes with DNA Microarrays. Antimicrob. Agents Chemother. 2003, 47, 3290–3295. [Google Scholar] [CrossRef] [Green Version]
- Iwahara, K.; Kuriyama, T.; Shimura, S.; Williams, D.W.; Yanagisawa, M.; Nakagawa, K.; Karasawa, T. Detection of cfxA and cfxA2, the beta-lactamase genes of Prevotella spp., in clinical samples from dentoalveolar infection by real-time PCR. J. Clin. Microbiol. 2006, 44, 172–176. [Google Scholar] [CrossRef] [Green Version]
- Dutour, C.; Bonnet, R.; Marchandin, H.; Boyer, M.; Chanal, C.; Sirot, D.; Sirot, J. CTX-M-1, CTX-M-3, and CTX-M-14 beta-lactamases from Enterobacteriaceae isolated in France. Antimicrob. Agents Chemother. 2002, 46, 534–537. [Google Scholar] [CrossRef] [Green Version]
- Ehrmann, E.; Handal, T.; Tamanai-Shacoori, Z.; Bonnaure-Mallet, M.; Fosse, T. High prevalence of -lactam and macrolide resistance genes in human oral Capnocytophaga species. J. Antimicrob. Chemother. 2014, 69, 381–384. [Google Scholar] [CrossRef]
- Voha, C.; Docquier, J.-D.; Rossolini, G.M.; Fosse, T. Genetic and biochemical characterization of FUS-1 (OXA-85), a narrow-spectrum class D beta-lactamase from Fusobacterium nucleatum subsp. polymorphum. Antimicrob. Agents Chemother. 2006, 50, 2673–2679. [Google Scholar] [CrossRef] [Green Version]
- Böckelmann, U.; Dörries, H.-H.; Ayuso-Gabella, M.N.; Salgot de Marçay, M.; Tandoi, V.; Levantesi, C.; Masciopinto, C.; van Houtte, E.; Szewzyk, U.; Wintgens, T.; et al. Quantitative PCR Monitoring of Antibiotic Resistance Genes and Bacterial Pathogens in Three European Artificial Groundwater Recharge Systems. Appl. Environ. Microbiol. 2009, 75, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Malhotra-Kumar, S.; Lammens, C.; Piessens, J.; Goossens, H. Multiplex PCR for Simultaneous Detection of Macrolide and Tetracycline Resistance Determinants in Streptococci. Antimicrob. Agents Chemother. 2005, 49, 4798–4800. [Google Scholar] [CrossRef] [Green Version]
- Perreten, V.; Vorlet-Fawer, L.; Slickers, P.; Ehricht, R.; Kuhnert, P.; Frey, J. Microarray-Based Detection of 90 Antibiotic Resistance Genes of Gram-Positive Bacteria. J. Clin. Microbiol. 2005, 43, 2291–2302. [Google Scholar] [CrossRef] [Green Version]
- Kangaba, A.A.; Saglam, F.Y.; Tokman, H.B.; Torun, M.; Torun, M.M. The prevalence of enterotoxin and antibiotic resistance genes in clinical and intestinal Bacteroides fragilis group isolates in Turkey. Anaerobe 2015, 35, 72–76. [Google Scholar] [CrossRef]
- Daly, M.M.; Doktor, S.; Flamm, R.; Shortridge, D. Characterization and prevalence of MefA, MefE, and the associated msr(D) gene in Streptococcus pneumoniae clinical isolates. J. Clin. Microbiol. 2004, 42, 3570–3574. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef] [Green Version]
- Dutka-Malen, S.; Evers, S.; Courvalin, P. Detection of glycopeptide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol. 1995, 33, 24–27. [Google Scholar] [CrossRef] [Green Version]
- Depardieu, F.; Perichon, B.; Courvalin, P. Detection of the van Alphabet and Identification of Enterococci and Staphylococci at the Species Level by Multiplex PCR. J. Clin. Microbiol. 2004, 42, 5857–5860. [Google Scholar] [CrossRef] [Green Version]
- Fines, M.; Perichon, B.; Reynolds, P.; Sahm, D.F.; Courvalin, P. VanE, a New Type of Acquired Glycopeptide Resistance in Enterococcus faecalis BM4405. Antimicrob. Agents Chemother. 1999, 43, 2161–2164. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef]
- Malbruny, B.; Werno, A.M.; Murdoch, D.R.; Leclercq, R.; Cattoir, V. Cross-Resistance to Lincosamides, Streptogramins A, and Pleuromutilins Due to the lsa (C) Gene in Streptococcus agalactiae UCN70. Antimicrob. Agents Chemother. 2011, 55, 1470–1474. [Google Scholar] [CrossRef] [Green Version]
- Navas, J.; Fernández-Martínez, M.; Salas, C.; Cano, M.E.; Martínez-Martínez, L. Susceptibility to Aminoglycosides and Distribution of aph and aac(3)-XI Genes among Corynebacterium striatum Clinical Isolates. PLoS ONE 2016, 11, e0167856. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.C.; Sanunu, M.; Schneider, C.; Clad, A.; Karygianni, L.; Hellwig, E.; Al-Ahmad, A. Rapid species-level identification of vaginal and oral lactobacilli using MALDI-TOF MS analysis and 16S rDNA sequencing. BMC Microbiol. 2014, 14, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Taxon | Number of Isolates |
|---|---|
| Streptococcus anginosus | 11 |
| Streptococcus constellatus | 1 |
| Streptococcus oralis | 24 |
| Streptococcus sanguinis | 18 |
| Streptococcus intermedius | 1 |
| Streptococcus gordonii | 9 |
| Streptococcus salivarius | 6 |
| Streptococcus sobrinus | 1 |
| Streptococcus vestibularis | 1 |
| Streptococcus downii | 2 |
| Streptococcus parasanguinis | 6 |
| Streptococcus mitis | 11 |
| Streptococcus mutans | 18 |
| Streptococcus cristatus | 3 |
| Actinomyces naeslundii | 7 |
| Actinomyces oris | 5 |
| Actinomyces odontolyticus | 7 |
| Rothia aeria | 7 |
| Rothia dentocariosa | 6 |
| Rothia mucilaginosa | 7 |
| Veilonella atypica | 10 |
| Veilonella dispar | 2 |
| Veilonella parvula | 14 |
| CHX | CPC | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Strains | MIC [µg/mL] at P1 # | MIC [µg/mL] at P10 # | FC P1–P10 | Numbers of Isolates with | MIC [µg/mL] at P1 # | MIC [µg/mL] at P10 # | FC P1–P10 | Numbers of Isolates with | ||
| 4-Fold | 8-Fold | 4-Fold | 8-Fold | |||||||
| MIC Increase | MIC Increase | |||||||||
| S. anginosus n = 11 | 8 (6; 8) | 16 (16; 16) | 2 (2; 4) | 4 | 0 | 2 (2; 4) | 2 (2; 2) | 1 (0.5; 1) | 0 | 0 |
| S. constellatus n = 1 | 4 | 4 | 1 | 0 | 0 | 1 (1; 1) | 2 (2; 2) | 2 (2; 2) | 0 | 0 |
| S. oralis n = 24 | 8 (4; 16) | 16 (16; 16) | 2 (1; 2.5) | 6 | 0 | 2 (2; 2) | 2 (2; 2,5) | 1 (1; 1.25) | 1 | 0 |
| S. sanguinis n = 18 | 12 (5; 28) | 16 (8; 16) | 1 (0.5; 2) | 1 | 0 | 4 (4; 4) | 2 (2; 4) | 0.5 (0.5; 1.75) | 2 | 0 |
| S. intermedius n = 1 | 4 | 8 | 2 | 0 | 0 | 8 | 4 | 0.5 | 0 | 0 |
| S. gordonii n = 9 | 4 (4; 8) | 16 (8; 16) | 2 (2; 4) | 4 | 0 | 2 (2; 4) | 4 (2; 4) | 1 (0.5; 2) | 0 | 0 |
| S. salivarius n = 6 | 4 (2.5; 4) | 6 (4; 8) | 3 (1.25; 4) | 2 | 1 | 2 (2; 2) | 2 (2; 2) | 1 (1; 1) | 0 | 0 |
| S. sobrinus n = 1 | 4 | 4 | 1 | 0 | 0 | 2 | 2 | 1 | 0 | 0 |
| S. vestibularis n = 1 | 1 | 8 | 8 | 0 | 1 | 2 | 1 | 0.5 | 0 | 0 |
| S. downii n = 2 | 4 (4; 4) ˟ | 16 (16; 16) ˟ | 4 (4; 4) ˟ | 2 | 0 | 3 (2; 4) ˟ | 1 (2; 2) ˟ | 0.38 (0.25; 0:5) ˟ | 0 | 0 |
| S. parasanguinis n = 6 | 4 (4; 7) | 16 (16; 16) | 3 (2; 4) | 3 | 0 | 4 (2.5; 4) | 2 (2; 2) | 0.5 (0.5; 0.88) | 0 | 0 |
| S. mitis n = 11 | 2 (2; 4) | 8 (4; 8) | 2 (2; 3) | 2 | 1 | 1 (1; 1) | 2 (2; 2) | 2 (1.5; 2) | 1 | 0 |
| S. mutans n = 18 | 4 (4; 4) | 4 (4; 4) | 1 (1; 1) | 0 | 1 | 2 (2; 2) | 4 (2; 4) | 2 (1; 2) | 1 | 0 |
| S. cristatus n = 3 | 2 (2; 3) | 4 (4; 6) | 2 (2; 2) | 0 | 0 | 1 (1; 1.5) | 4 (2.5; 4) | 2 (1.5; 3) | 1 | 0 |
| A. naeslundii n = 7 | 1 (0.75; 2) | 2 (1.5; 2) | 2 (1; 2) | 0 | 0 | 2 (1.5; 2) | 2 (2; 2) | 1 (1; 1.5) | 0 | 0 |
| A. oris n = 5 | 2 (1; 2) | 2 (2; 4) | 1 (1; 4) | 2 | 0 | 4 (4; 4) | 2 (2; 2) | 0.5 (0.5; 1) | 0 | 0 |
| A. odontolyticus n = 7 | 4 (3; 4) | 4 (2; 4) | 1 (0.75; 1) | 0 | 0 | 4 (4; 4) | 2 (2; 2) | 0.5 (0.5; 0.5) | 0 | 0 |
| R. aeria n = 7 | 2 (2; 4) | 4 (2; 4) | 1 (1; 2) | 0 | 0 | 2 (2; 4) | 2 (2; 2) | 1 (0.5; 1) | 0 | 0 |
| R. dentocariosa n = 6 | 4 (4; 4) | 4 (4; 4) | 1 (1; 1) | 0 | 0 | 2 (2; 2) | 2 (2; 2) | 1 (1; 1) | 0 | 0 |
| R. mucilaginosa n = 7 | 4 (3; 4) | 4 (4; 6) | 2 (1; 2) | 0 | 0 | 2 (1; 2) | 2 (2; 2) | 1 (1; 1.5) | 0 | 0 |
| V. atypica n = 10 | 1 (1; 1) | 2 (1; 2) | 2 (1; 2) | 0 | 0 | 1 (1; 1.75) | 2 (1.25; 2) | 2 (1; 2) | 0 | 0 |
| V. dispar n = 2 | 2 (2; 2) ˟ | 1 (1; 1) ˟ | 0.5 (0.5; 0.5) ˟ | 0 | 0 | 0.75 (0.625; 0.875) ˟ | 2 (2; 2) ˟ | 3 (2.5; 3.5) ˟ | 1 | 0 |
| V. parvula n = 14 | 2 (1; 2) | 1.5 (1; 2) | 1 (1; 1) | 0 | 0 | 1.5 (1; 2) | 2 (2; 2) | 2 (1; 2) | 0 | 0 |
| Strain | MICCHX (µg/mL) | FC P1–P10 | FC P1–R72 h | FC P10–R72 h | ||||
|---|---|---|---|---|---|---|---|---|
| P1 | P10 | R24 h | R72 h | |||||
| S. mutans | 59a | 0.5 | 4 | 4 | 2 | 8 | 4 | 0.5 |
| 59b | 0.5 | 0.5 | * | * | 1 | * | * | |
| 59c | 0.5 | 2 | * | * | 4 | * | * | |
| 59d | 0.5 | 4 | 4 | 4 | 8 | 8 | 1 | |
| 59e | - | - | - | - | - | - | - | |
| 59f | 0.5 | 4 | 4 | 2 | 8 | 4 | 0.5 | |
| S. salivarius | 73a | 1 | 8 | 16 | 8 | 8 | 8 | 1 |
| 73b | 1 | 2 | * | * | 2 | * | * | |
| 73c | 1 | 4 | * | * | 4 | * | * | |
| 73d | 1 | 4 | * | * | 4 | * | * | |
| 73e | 1 | 4 | * | * | 4 | * | * | |
| 73f | 1 | 4 | * | * | 4 | * | * | |
| S. vestibularis | 78a | 1 | 4 | * | * | 4 | * | * |
| 78b | 1 | 4 | * | * | 4 | * | * | |
| 78c | 2 | 4 | * | * | 2 | * | * | |
| 78d | 2 | 8 | * | * | 4 | * | * | |
| 78e | 1 | 8 | 16 | 8 | 8 | 8 | 1 | |
| 78f | 2 | 4 | * | * | 2 | * | * | |
| S. mitis | 93a | 1 | 4 | * | * | 4 | * | * |
| 93b | 1 | 8 | 4 | 4 | 8 | 4 | 0.5 | |
| 93c | 1 | 8 | 8 | 8 | 8 | 8 | 1 | |
| 93d | 1 | 8 | 8 | 8 | 8 | 8 | 1 | |
| 93e | 1 | 8 | 8 | 8 | 8 | 8 | 1 | |
| 93f | 1 | 4 | * | * | 4 | * | * | |
| S. mutans | S. salivarius | S. vestibularis | S. mitis | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 59 WT | 59a | 59d | 59f | 73 WT | 73a | 78 WT | 78e | 93 WT | 93b | 93c | 93d | 93e | ||
| β-lactams | PenG | 0.023 S | 0.023 S | 0.023 S | 0.023 S | 0.5 I | 0.5–0.75 I | 0.064 S | 0.094 S | 0.19 S | 0.25 S | 0.125 S | 0.016–0.25 S | 0.19 S |
| AMP /AMX | 0.047 S | 0.047 S | 0.047 S | 0.047 S | 0.38–0.75 S/I | 0.5–1.0 S/I | 0.064 S | 0.064 S | 0.38 S | 0.5–0.75 S/I | 0.38 S | 0.38–0.5 I | 0.38 S | |
| CXM | 0.032 S | 0.032 S | 0.047 S | 0.032 S | 0.125–0.19 S | 0.19 S | 0.047 S | 0.032 S | 0.5 S | 0.75–1 R | 0.5 S | 0.032–0.5 S | 0.5–0.75 S/R | |
| MEM | 0.064 S | 0.064 S | 0.064 S | 0.064 S | 0.19 S | 0.25–0.38 S | 0.032 S | 0.047 S | 0.19–0.25 S | 0.38 S | 0.25–0.38 S | 0.032–0.25 S | 0.25–0.38 S | |
| Tetracyclines | TET | 0.19 S | 0.19 S | 0.19 S | 0.19 S | 0.19 S | 0.19–0.25 S | 0.19 S | 0.19 S | 12–24 R | 16–24 R | 12–24 R | 12 R | 12–24 R |
| TGC | 0.064 S | 0.064 S | 0.064 S | 0.064 S | 0.047–0.064 S | 0.047–0.064 S | 0.032 S | 0.047 S | 0.047–0.064 S | 0.047–0.064 S | 0.016–0.064 S | 0.016–0.047 S | 0.032–0.047 S | |
| Lincosamide | CLI | 0.064 S | 0.064 S | 0.064 S | 0.064 S | 0.047 S | 0.047 S | 0.032 S | 0.047 S | 0.094 S | 0.094 S | 0.047–0.094 S | 0.094 S | 0.094 S |
| Macrolide | ERY | 0.047 S | 0.047 S | 0.047 S | 0.047 S | 8–12 R | 4–6 R | 6 R | 3 R | 6–8 R | 8–24 R | 8–24 R | 2 R | 12–24 R |
| Fluoro-quinolone | MXF | 0.25 S | 0.38 S | 0.25 S | 0.25 S | 0.125–0.19 S | 0.125–0.19 S | 0.19 S | 0.19 S | 0.19–0.25 S | 0.19 S | 0.064–0.094 S | 0.125–0.19 S | 0.094–0.125 S |
| Glycopeptide | VAN | 0.75 S | 0.5 S | 0.5 S | 0.75 S | 0.5 S | 0.5–0.75 S | 0.5 S | 0.75 S | 0.25 S | 0.38 S | 0.25–0.38 S | 0.38 S | 0.5–0.38 S |
| ARG | S. mutans | S. salivarius | S. vestibularis | S. mitis | Negative Control | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 59 WT | 59a | 59d | 59f | 73 WT | 73a | 78 WT | 78e | 93 WT | 93b | 93c | 93d | 93e | ||
| tetM | − | − | − | − | − | − | − | − | + | + | + | + | + | − |
| tetO | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetW | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetA-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetB-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetC-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetD-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| tetE-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| blaTEM1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| cfxA | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| blaCTX-M-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| blaCSP-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| blaOXA-85 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| ampC | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| pbpX2 | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| ermA | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| ermB | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| ermC | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| ermF | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| MefI | − | − | − | − | + | + | + | + | + | + | + | + | + | − |
| mefAI | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| mef A II | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| patA | − | − | − | − | − | − | − | − | + | + | + | + | + | − |
| patB | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| vanA | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| vanB | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| vanC1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| vanC2/3 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| vanD | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| vanE | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| mcr-1 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| lsaC | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| aph3 | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| int-II | − | − | − | − | − | − | − | − | + | + | + | + | + | − |
| xis-II | − | − | − | − | − | − | − | − | + | + | + | + | + | − |
| positive control | + | + | + | + | + | + | + | + | + | + | + | + | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auer, D.L.; Mao, X.; Anderson, A.C.; Muehler, D.; Wittmer, A.; von Ohle, C.; Wolff, D.; Frese, C.; Hiller, K.-A.; Maisch, T.; et al. Phenotypic Adaptation to Antiseptics and Effects on Biofilm Formation Capacity and Antibiotic Resistance in Clinical Isolates of Early Colonizers in Dental Plaque. Antibiotics 2022, 11, 688. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050688
Auer DL, Mao X, Anderson AC, Muehler D, Wittmer A, von Ohle C, Wolff D, Frese C, Hiller K-A, Maisch T, et al. Phenotypic Adaptation to Antiseptics and Effects on Biofilm Formation Capacity and Antibiotic Resistance in Clinical Isolates of Early Colonizers in Dental Plaque. Antibiotics. 2022; 11(5):688. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050688
Chicago/Turabian StyleAuer, David L., Xiaojun Mao, Annette Carola Anderson, Denise Muehler, Annette Wittmer, Christiane von Ohle, Diana Wolff, Cornelia Frese, Karl-Anton Hiller, Tim Maisch, and et al. 2022. "Phenotypic Adaptation to Antiseptics and Effects on Biofilm Formation Capacity and Antibiotic Resistance in Clinical Isolates of Early Colonizers in Dental Plaque" Antibiotics 11, no. 5: 688. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050688