
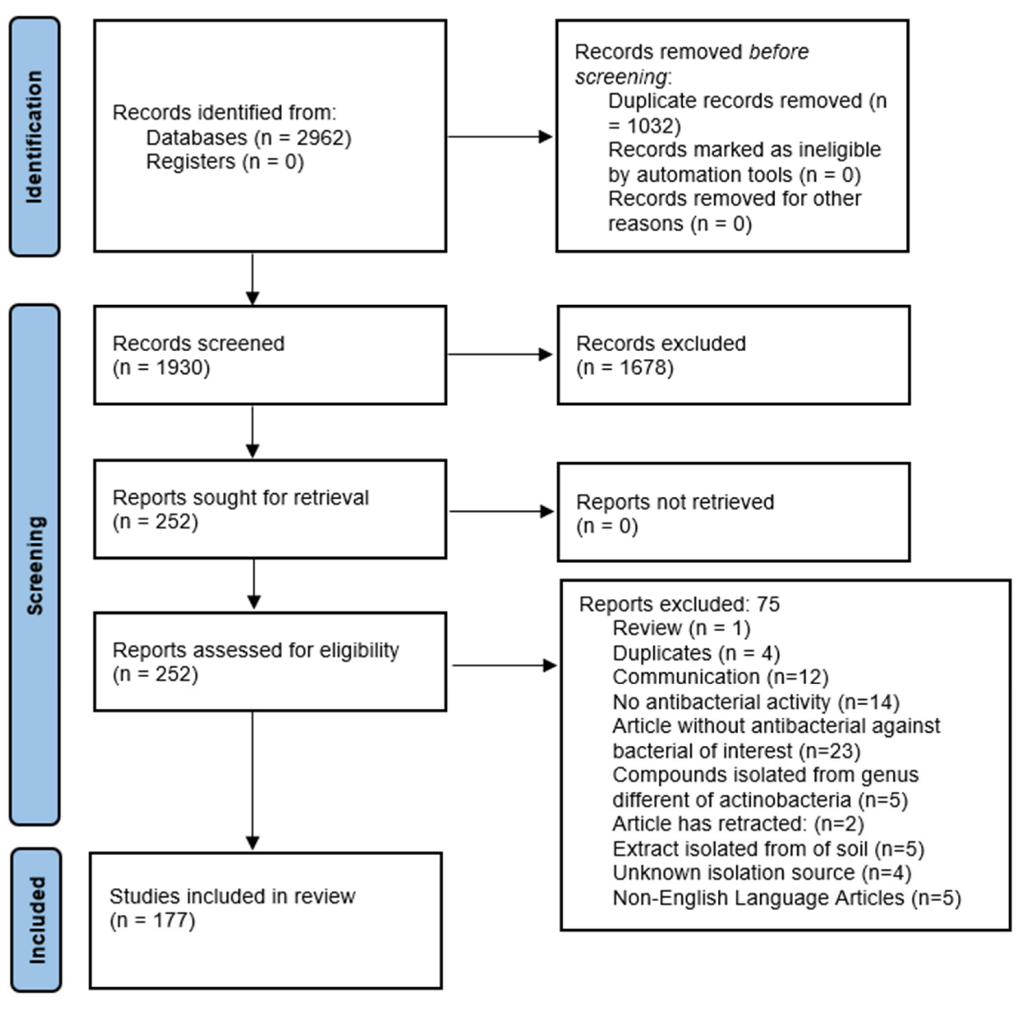
Marine Actinobacteria a New Source of Antibacterial Metabolites to Treat Acne Vulgaris Disease—A Systematic Literature Review


Abstract
:1. Introduction
2. Results
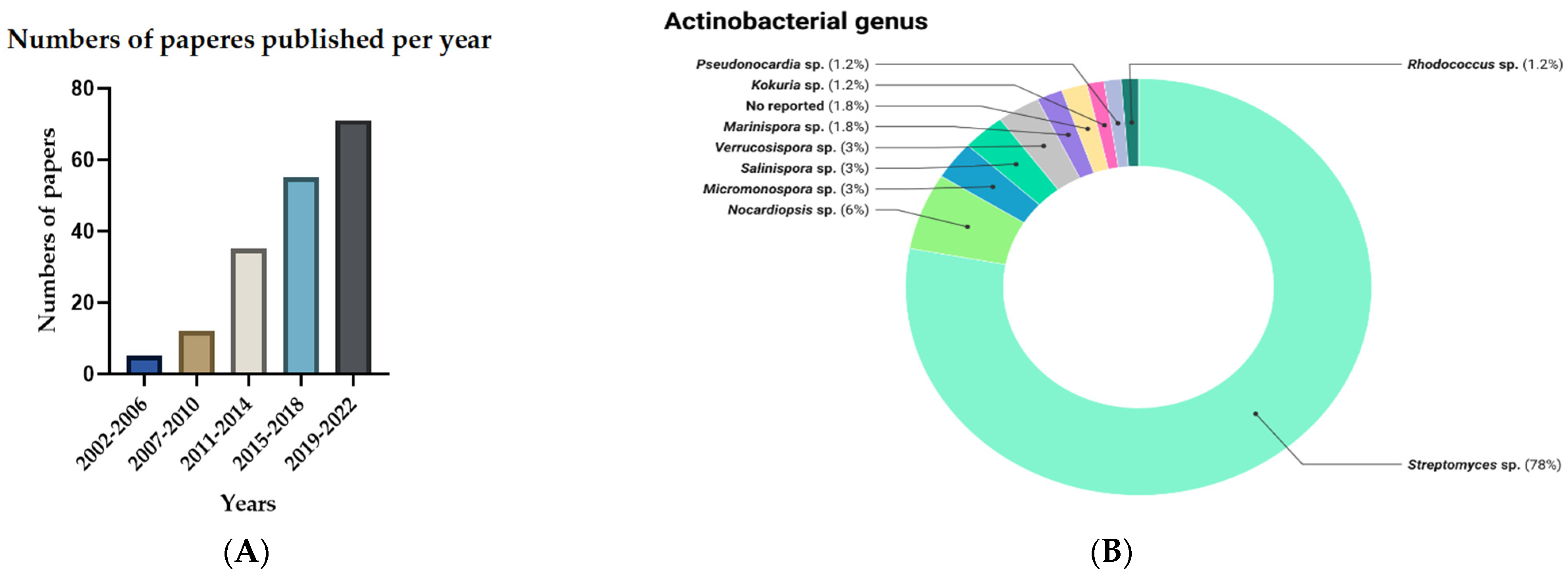
2.1. General Information
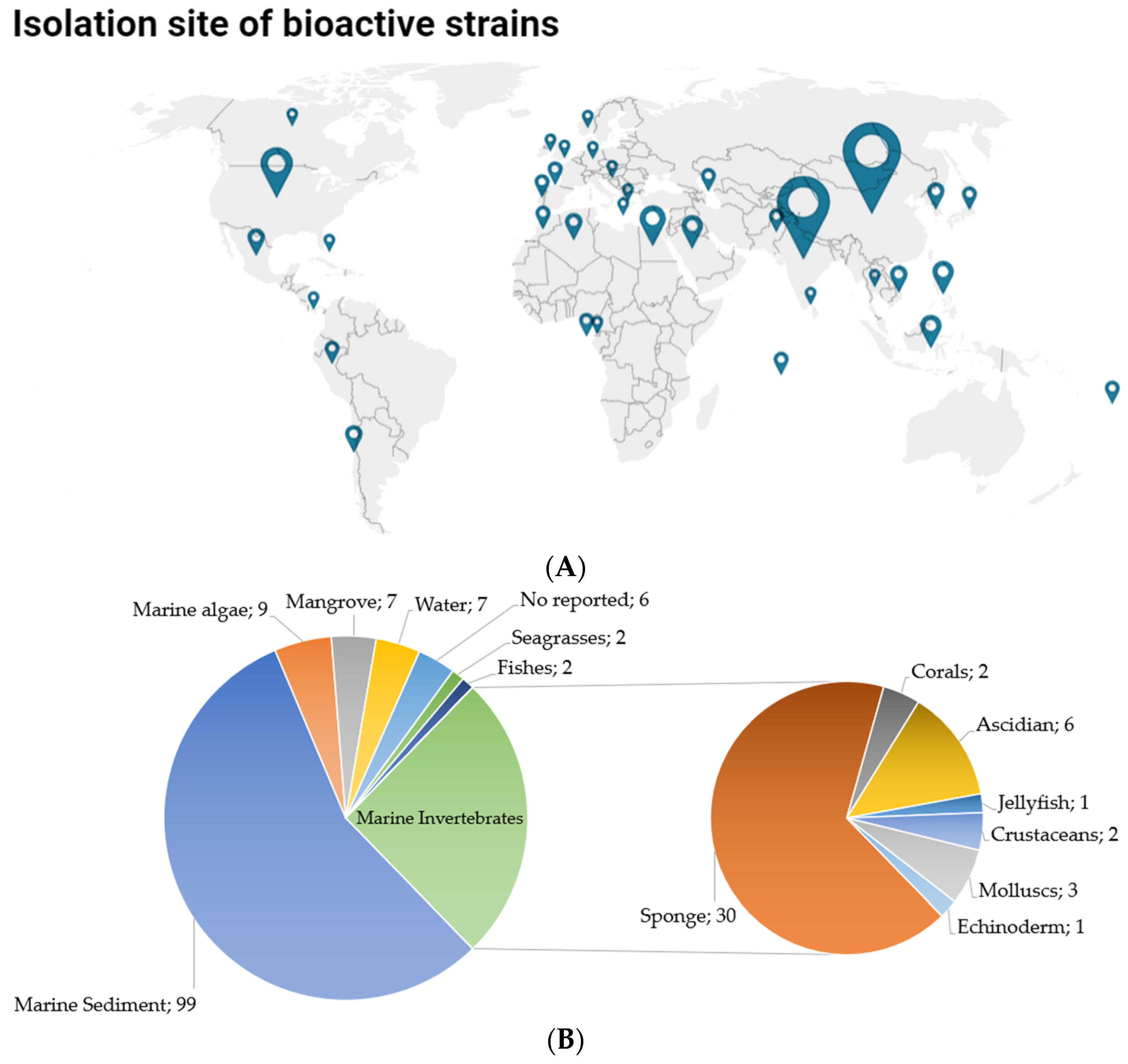
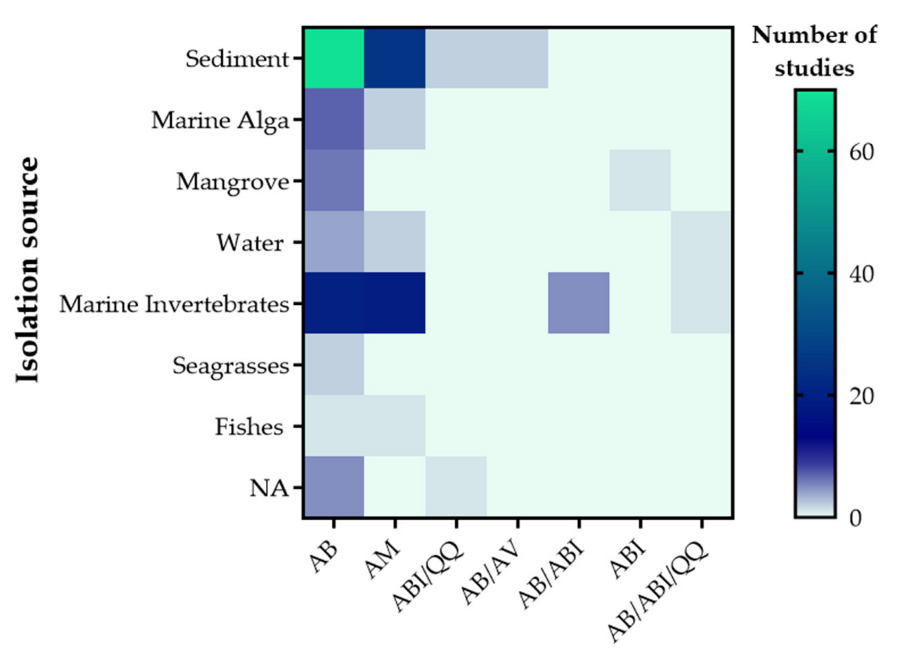
2.2. Isolation Sources
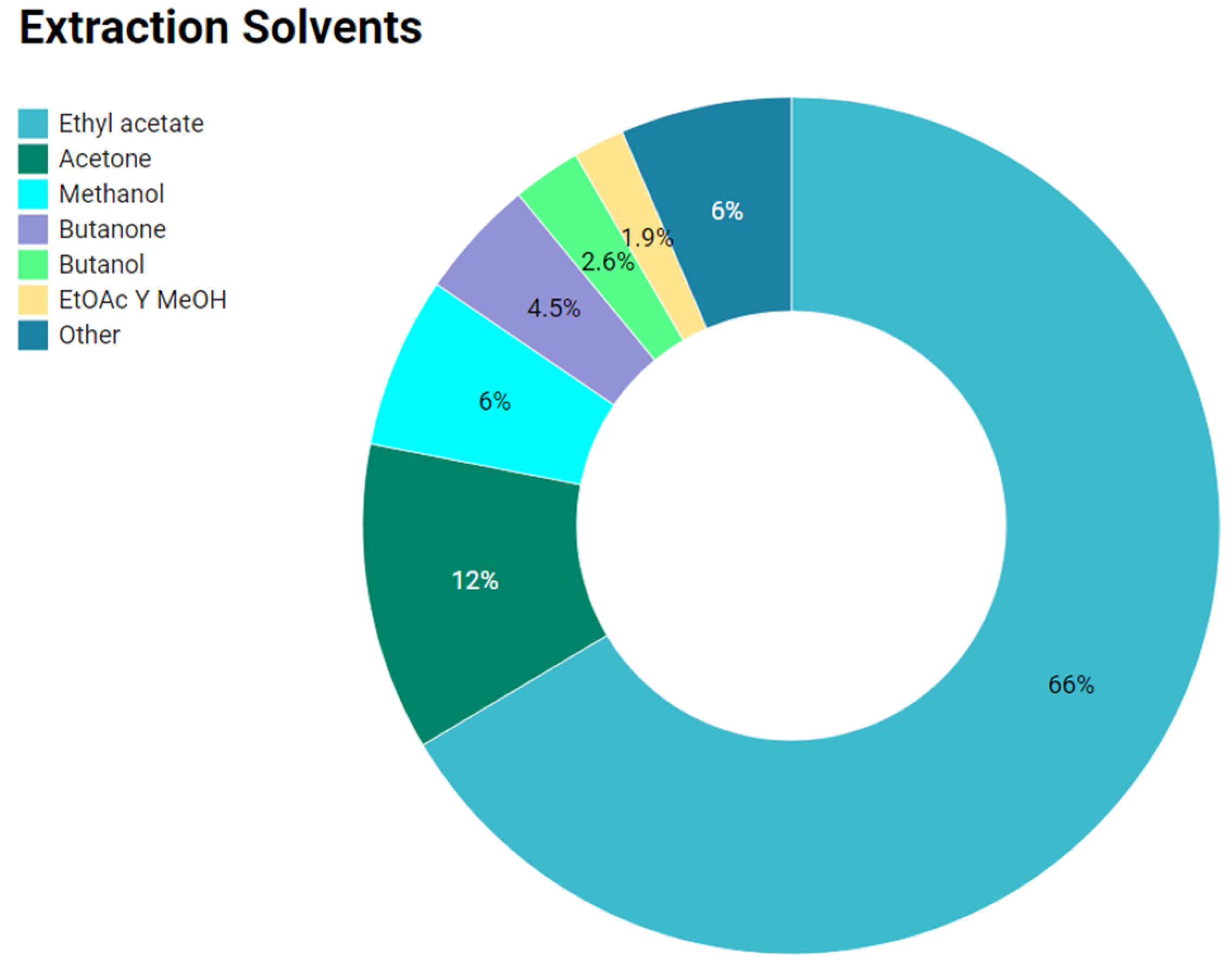
2.3. Organic Solvents Used to Obtain Anti-Infective Extracts
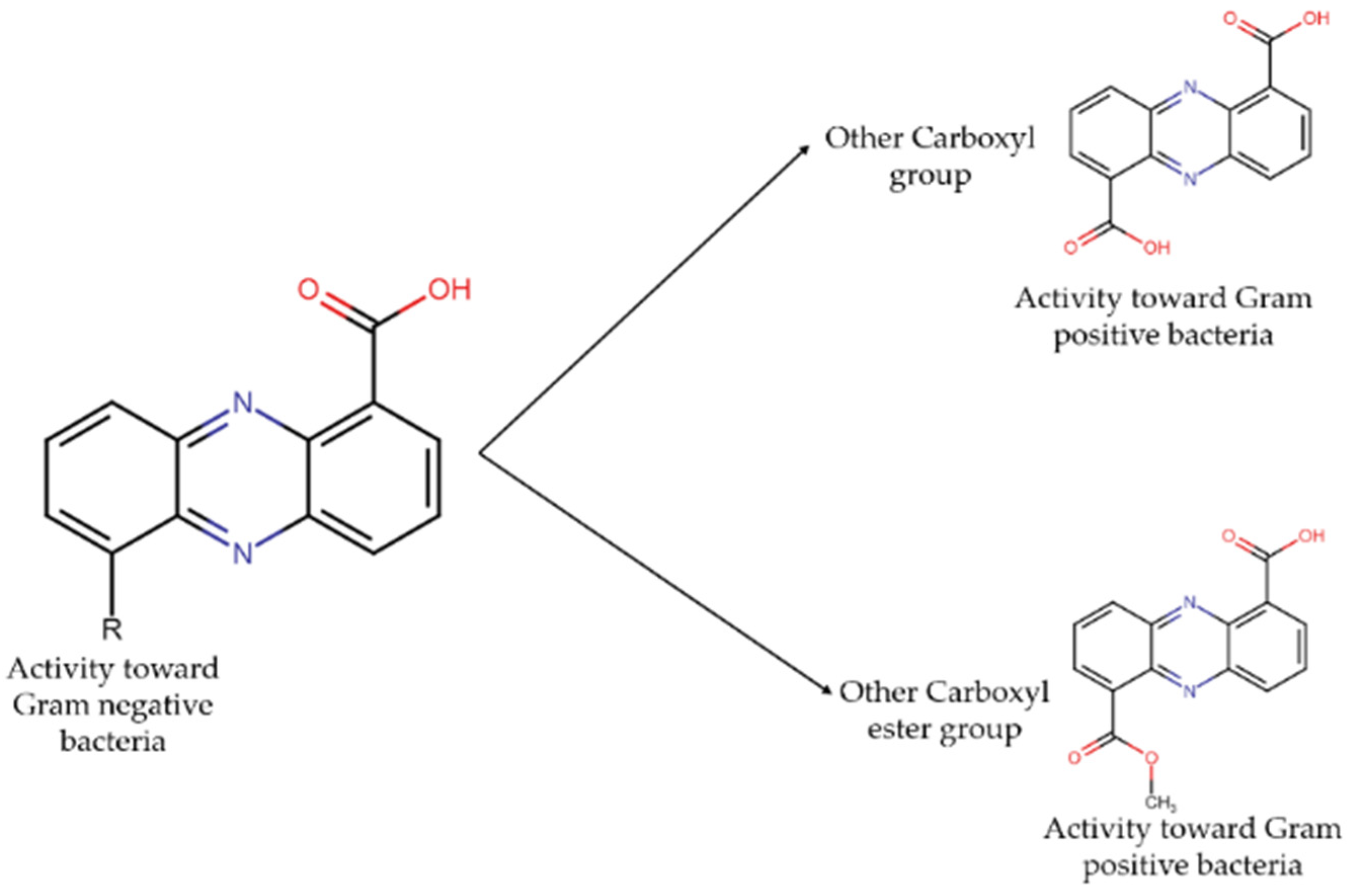
2.4. Anti-Infective Metabolites Derived from Marine Actinobacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Pathogen Target | Compounds/Extracts | MIC (μg/mL) | Ref. |
|---|---|---|---|---|
| Streptomyces sp. | MRSA 1 | Napyradiomycins 1 | 0.016 | [35] |
| Napyradiomycins 8 | 0.002 | [35] | ||
| Streptomyces sp. | MRSA 1 | Marinopyrrole A | 3.24 | [36] |
| Marinopyrrole B | 3.24 | [36] | ||
| Streptomyces sp. | S aureus ATCC NR-46171 | 4-methoxyacetanilide | 32.4 | [18] |
| Streptomyces sp. | S. aureus | Flaviogeranin D | 9.2 | [37] |
| Flaviogeranin C2 | 8.1 | [37] | ||
| Streptomyces sp. | S. aureus | 1-hydroxy-1-norresistomycin | >40 | [38] |
| Streptomyces sp. | MRSA 1 | Fridamycin A | 500 | [23] |
| Fridamycin D | 62.5 | [23] | ||
| Streptomyces sp. | MRSA 1 | Chromomycin A3 | 0.698 | [39] |
| Streptomyces sp. | MRSA 1 | Extract | 2 | [40] |
| Streptomyces sp. | MRSA ATCC 33591 | Actinomycins D1 | 0.125 | [41] |
| Actinomycins D2 | 0.25 | [41] | ||
| Actinomycins D3 | 0.5 | [41] | ||
| Actinomycins D4 | 0.25 | [41] | ||
| Actinomycins D | 0.25 | [41] | ||
| Streptomyces sp. | S. aureus CCARM 3090 | Grincamycin L | 6.25 | [42] |
| Streptomyces sp. | MRSA 1 | Compound 2 | 2 | [43] |
| Streptomyces sp. | S. aureus (ATCC 29213) | 2,4-dichloro-5-sulfamoyl benzoic acid | 0.8–4 | [44] |
| Streptomyces sp. | S. aureus (ATCC 25923). | Dionemycin | 0.5–2 | [45] |
| Streptomyces sp. | S. aureus ATCC 43300 | Extract | 7.9 | [46] |
| Streptomyces sp. | S. aureus ATCC 43300 | Extract | 12.5 | [47] |
| S. epidermidis (ATCC 12228) | Extract | 25 | [47] | |
| Streptomyces sp. | S. aureus | Aborycin | 8.0~64 | [48] |
| MRSA 1 | 16~64 | [48] | ||
| MRSE 3 | 128 | [48] | ||
| Streptomyces sp. | MRSA 1 | Supernatant | 0.78 | [49] |
| Streptomyces sp. | MRSE 3 | Dehydroxyaquayamycin | 16.0 | [50] |
| Streptomyces sp. | MRSA 1 | Medermycin | 2 | [51] |
| G15-F | 4 | [51] | ||
| Streptomyces sp. | MRSA 1 ATCC BAA-44 | Bisanhydroaklavinone | 6.25 | [19] |
| 1-Hydroxybisanhydroaklavinone | 50 | [19] | ||
| Streptomyces sp. | MRSA 1 | 11′,12′-dehydroelaiophylin | 1−4 | [52] |
| MRSA 1, MRSE 3 | Elaiophylin | 1−4 | [52] | |
| 11-monomethoxylated derivative | 2−16 | [52] | ||
| Compound 6 4 | 2−16 | [52] | ||
| Streptomyces sp. | MRSA 1 | Lactoquinomycin A | 0.25–0.5 | [53] |
| MRSA 1 | Stremycin A | 16 | [54] | |
| Stremycin B | 16 | [54] | ||
| Streptomyces sp. | MRSA 1 | Quinomycin G | 16–64 | [55] |
| MRSE 3 | ||||
| MSSE 5 | ||||
| Streptomyces sp. | S. aureus (ATCC 6538) | Actinomycins X2 | 0.394 | [56] |
| MRSA 1 (ATCC 43300) | Actinomycins X2 | 0.190 | [56] | |
| S. aureus (ATCC 6538) | Actinomycins D | 0.389 | [56] | |
| MRSA 1 (ATCC 43300) | Actinomycins D | 0.188 | [56] | |
| Streptomyces sp. | MRSA 1 | Extract 7 | 6.25 | [25] |
| MSSA 6 | Extract 7 | 12.5 | [25] | |
| Streptomyces sp. | MRSA 1 | Extract 8 | 12.5 | [25] |
| MSSA 6 | ||||
| Streptomyces sp. | MRSA 1 | Borrelidins J | 0.195 | [28] |
| Streptomyces sp. | S. aureus | Extract | 256 | [57] |
| Streptomyces sp. | S. epidermidis | Extract | 128 | [57] |
| Streptomyces sp. | MRSA 1 | Streptopertusacin A | 40 | [58] |
| 21,22-en-bafilomycin D | 12.5 | [58] | ||
| 21,22-en-9- | ||||
| hydroxybafilomycin D | 12.5 | [58] | ||
| Streptomyces sp. | S. aureus | Lobophorins E | 32 | [59] |
| ATCC 29213 | Lobophorins F | 8 | [59] | |
| Streptomyces sp. | MRSA 1 | Pyrrole-derivative | 2.8 | [60] |
| Streptomyces sp. | MRSA 1 | Julichromes Q11 | 16–64 | [61] |
| S. aureus ATCC 29213 | Julichromes Q10 | 16–64 | [61] | |
| Julichromes Q6.6 | 16–64 | [61] | ||
| Julichromes Q6 | 16–64 | [61] | ||
| Streptomyces sp. | MRSA 1, S. aureus | Lobophorin-like spirotetronate | 64 | [62] |
| Streptomyces sp. | MRSA 1, S. aureus | Ansamycins | 32 | [63] |
| MRSA 1 | (-)-Streptophenazine B | 4.2 | [63] | |
| Streptomyces sp. | MRSA 1 | Neo-actinomycin A | 16–64 | [64] |
| Streptomyces sp. | S. aureus ATCC 29213 | MarfuraquinocinsA | 8.0 | [65] |
| Streptomyces sp. | MRSE 3 shhs-E1 | Marfuraquinocins C | 8.0 | [65] |
| S. aureus ATCC 29213 | Marfuraquinocins D | 8.0 | [65] | |
| Streptomyces sp. | MRSA 1 ATCC 43300 | 7,8-dideoxygriseorhodin C | 0.08–0.12 | [66] |
| Oxacillin and 7,8-dideoxygriseorhodin C | 0.01–0.02 | [66] | ||
| Streptomyces sp. | MSSA 6 11497 | Desertomycin G | 4.0 | [67] |
| MRSA 1 ATCC 43300 | Desertomycin G | 4.0 | [67] | |
| MRSA 1 ATCC 25923 | Desertomycin G | 4.0 | [67] | |
| Streptomyces sp. | S. aureus ATCC 6518, MTCC 3160 | Aromatic polyketide | 32.40 | [68] |
| MRSA 1 | ||||
| Streptomyces sp. | S. aureus ATCC 29213 | Napyradiomycins 1–8 9 | 0.5 to 32 | [69] |
| MRSA 1 | ||||
| Streptomyces sp. | S. aureus ATCC 29213 | Marinopyrroles A–C | <1 | [70] |
| Marinopyrroles F | 3.1 | [70] | ||
| Streptomyces sp. | MRSA 1-ATCC33591 | A80915A 10 | 1–4 | [71] |
| Streptomyces sp. | MRSA 1 ATCC 43300 | Polyketide 13 11 | 2 | [72] |
| Streptomyces sp. | MRSA 1 | Fijimycins A–C Etamycin A | 4–16 | [73] |
| Streptomyces sp. | S. aureus HA- and CA- | Etamycin | 1–2 | [74] |
| Streptomyces sp. | MRSA 1 | Lydicamycin congeners | 1.56–12.5 | [75] |
| Streptomyces sp. | MRSA 1 | Salinamide F | 100 | [76] |
| S. aureus (ATCC 12600) | ||||
| Streptomyces sp. | S. aureus | Antimycin B1 | 32 | [77] |
| Streptomyces sp. | S. aureus | Merochlorins G | 16 | [78] |
| Merochlorins J | 2 | [78] | ||
| Streptomyces sp. | S. aureus | cyclo(L-Pro-L-Tyr) | 160 | [79] |
| cyclo(L-Pro-L-Phe) | 180 | [79] | ||
| Streptomyces sp. | MRSA 1 | Actinomycin X2 | 3.125–12.5 | [80] |
| Actinomycin D | 12.5–25 | [80] | ||
| Streptomyces sp. | S. aureus | 1,3-Benzodioxole | 256 | [81] |
| Streptomyces sp. | S. aureus ATCC 29213 | Desotamide, Desotamide B | 16 | [82] |
| MRSE | 32 | [82] | ||
| Streptomyces sp. | S. epidermidis | Streptophenazines G | 3.68 | [83] |
| Streptophenazines F | 6.77 | [83] | ||
| Streptomyces sp. | MRSA 1 | Citreamicin θ A | 0.25 | [84] |
| ATCC43300 | Citreamicin θ B | 0.25 | [84] | |
| Citreaglycon A | 8.0 | [84] | ||
| S. aureus UST950701-005 | Dehydrocitreaglycon A | 16 | [84] | |
| Streptomyces sp. | S. aureus DSM346 | Alageninthiocin | 15 | [85] |
| Geninthiocin | 4 | [85] | ||
| Val-geninthiocin | 8 | [85] | ||
| Indolocarbazole staurosporine | 19 | [85] | ||
| Streptomyces sp. | MRSA 1 | Anthraquinone derivatives | 6.25 | [86] |
| Streptomyces sp. | MRSA 1 | Extract | 1000 | [87] |
| Streptomyces sp. | S. aureus | Extracts | 312–2.5 × 102 | [88] |
| Streptomyces sp. | S. aureus | Extract | 400 | [89] |
| Streptomyces sp. | S. aureus | Extract AIA12 | 2.5 × 102 | [90] |
| ATCC 25923 | ||||
| Extract AIA17 | 310 | [90] | ||
| Streptomyces sp. | MRSA 1 | 1-Acetyl-β-Carbonile | 128–256 | [91] |
| MSSA 6 | 1-Acetyl-β-Carbonile | 64 | [91] | |
| Streptomyces sp. | MRSA 1 | Chlororesistoflavins A | 0.25 | [92] |
| MRSA 1 | Chlororesistoflavins B | 2.0 | [92] | |
| Streptomyces sp. | S. aureus | Ligiamycin A | 16 | [26] |
| S. aureus | Ligiamycin B | 64 | [26] | |
| Verrucosispora sp. | S. aureus ATCC 33591 | Active fraction | 16–32 | [93] |
| Verrucosispora sp | S. aureus ATCC29213 | Proximicins B | 16 | [94] |
| MRSA shhs-A1 | ||||
| Verrucosispora sp. | MRSA 1 | 1-hydroxy-2,5-dimethyl benzoate | 12.5 | [95] |
| Verrucosispora sp. | MRSA 1 | Proximicin B | 3.125 | [95] |
| Micromonospora sp. | S. aureus ATCC 29213 | Kendomycins B | 0.5–2 | [96] |
| S. aureus 745524 | Kendomycins C | 0.5–1 | [96] | |
| MRSA 1 shhs-A1 | Kendomycins D | 1–4 | [96] | |
| Micromonospora sp. | MRSA 1 | 2-ethylhexyl 1H-imidazole-4- carboxylate | 16 | [97] |
| Micromonospora sp. | S. aureus ATCC 29213 | Micromonohalimanes B | 40 | [98] |
| Micromonospora sp. | S. aureus ATCC 29213 | Rabelomycin | 1 | [99] |
| Phenanthroviridone | 0.25 | [99] | ||
| Micromonospora sp. | S. aureus ATCC 29213 | homo-dehydrorabelomycin E | 1 | [100] |
| Nocardiopsis sp. | MRSA 1 | Bis (2-ethylhexyl) phthalate | 7.81 | [101] |
| MRSA 1 | 4-bromophenol | 15.62 | [101] | |
| ATCC NR-46071 | ||||
| Nocardiopsis sp. | MRSA 1 | Nocardiopsistin A | 12.5 | [102] |
| Nocardiopsistin B | 3.12 | [102] | ||
| Nocardiopsistin C | 12.5 | [102] | ||
| Nocardiopsis sp. | MRSA 1 | α-Pyrone | 12.5 | [103] |
| Nocardiopsis sp. | MRSA 1 | Extracts | 115–125 | [104] |
| Marinispora sp. | MSSA 6 | Lipoxazolidinone A | 1–2 | [105] |
| MRSA 1 | ||||
| Marinispora sp. | MRSA 1 | Lynamicins A–E | 2.2–45 | [106] |
| MRSE 3 ATCC 700578c | ||||
| Pseudonocardia carboxydivorans | S. aureus ATCC 6538P | Branimycins C | 32 | [107] |
| MRSA 1 MB5393 | Branimycins C | 20–40 | [107] | |
| Kocuria sp. | MRSA 1 ATCC 43300- | Kocurin | 0.25–0.5 | [108] |
| Solwaraspora sp. | MRSA 1 | Solwaric acids A | 32 | [109] |
| Solwaric acids B | 32 | [109] | ||
| MSSA 6 | Solwaric acids A | 64 | [109] | |
| Solwaric acids B | 64 | [109] | ||
| Salinispora sp. | MRSA 1 | Rifamycin W | 15.62 | [110] |
| Genus | Target Bacteria in Antibiofilm Activity | MBIC 1 | Compounds/ Extracts | Percentage Decreased Biofilm | QS System | QQ Activity (IC50) | Biosensor Strain | Ref. |
|---|---|---|---|---|---|---|---|---|
| Streptomyces sp. | MRSA 2 | 200 | Butenolide | >70 | AI-2 up to 70% | NA 3 | Vibrio harveyi BB170 | [138] |
| AHL inhibition up to 97% | C. violaceum | [138] | ||||||
| S. aureus | 100 | Extract | 78.9 | AHL | NA 3 | C. violaceum 12472 | [140] | |
| Nocardiopsis sp. | S. aureus | NA 3 | Melanin JN1M | 64.2 | AHL | NA 3 | C. violaceum MTCC 26563 | [137] |
| Melanin JN2M | 65.9 | AHL | NA 3 | C. violaceum MTCC 26563 | [137] | |||
| Nocardiopsis sp. | S. aureus | 20 vol % 4 | Culture liquid of JS106 | 77.94 | AHL | NA 3 | C. violaceum 12472 | [29] |
| NA 3 | NA 3 | Questiomycin A | NA 3 | AHL | 6.82 | C. violaceum 12472 | [29] | |
| NA 3 | NA 3 | 2-hydroxyacetate-3-hydroxyacetamido-phenoxazine (HHP) | NA 3 | AHL | 23.59 | C. violaceum 12472 | [29] |
2.5. Actinobacteria Producing Quorum Quenching Metabolites
2.6. Strategies to Maximize Anti-Infective Metabolites Activity and Yield
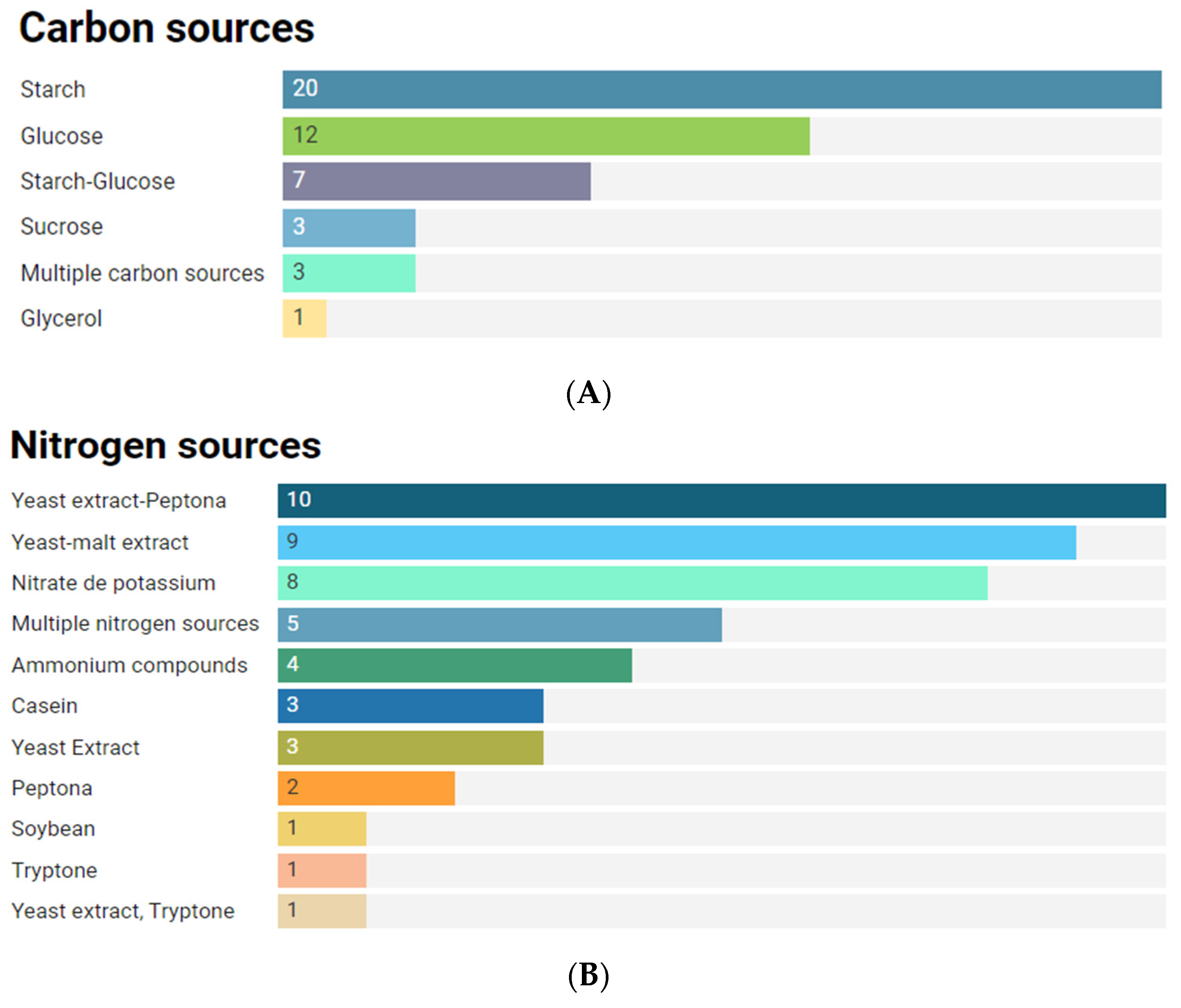
2.6.1. Culture Conditions to Anti-Infective Production Metabolites
2.6.2. Co-Culture Combination as Strategies to Maximize Anti-Infective Metabolites in Marine Actinobacteria
2.7. Main Families of Compounds Found in Marine Actinobacteria with Antibacterial Activity
2.8. Main Family Compounds Found in Marine Actinobacteria with QQ Activity
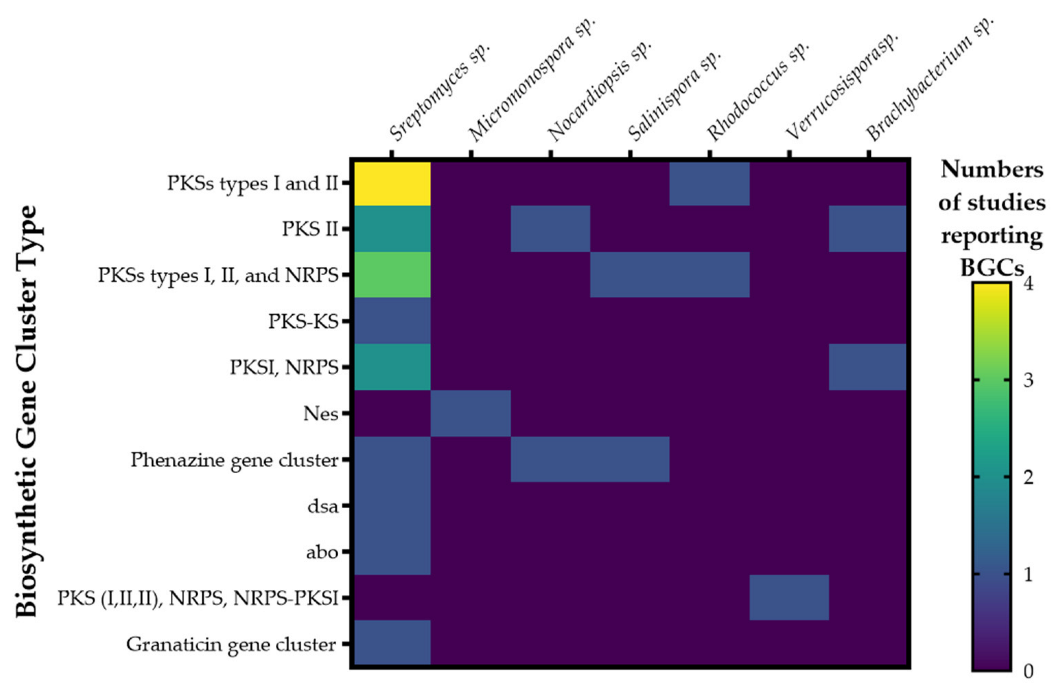
2.9. Biosynthetic Gene Clusters, BGCs
3. Discussion
4. Materials and Methods
4.1. Search Strategy
4.2. Eligibility Criteria
4.3. Data Extraction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuchayi, S.M.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C. Acne vulgaris. Nat. Rev. Dis. Prim. 2015, 1, 15029. [Google Scholar] [CrossRef] [PubMed]
- Heng, A.H.S.; Chew, F.T.; Heng, A.H.S.; Chew, F.T. Systematic review of the epidemiology of acne vulgaris. Sci. Rep. 2020, 10, 5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dréno, B.; Araviiskaia, E.; Kerob, D.; Andriessen, A.; Anfilova, M.; Arenbergerova, M.; Barrios, O.L.F.; Mokos, Z.B.; Haedersdal, M.; Hofmann, M.A.; et al. Nonprescription acne vulgaris treatments: Their role in our treatment armamentarium—An international panel discussion. J. Cosmet. Dermatol. 2020, 19, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Gannesen, A.V.; Zdorovenko, E.L.; Botchkova, E.A.; Hardouin, J.; Massier, S.; Kopitsyn, D.S.; Gorbachevskii, M.V.; Kadykova, A.A.; Shashkov, A.S.; Zhurina, M.V.; et al. Composition of the Biofilm Matrix of Cutibacterium acnes Acneic Strain RT5. Front. Microbiol. 2019, 10, 1284. [Google Scholar] [CrossRef]
- Platsidaki, E.; Dessinioti, C. Recent advances in understanding Propionibacterium acnes (Cutibacterium acnes) in acne. F1000 Res. 2018, 7, 1953. [Google Scholar] [CrossRef] [Green Version]
- Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes: Two Major Sentinels of Skin Microbiota and the Influence of Cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef]
- de Sousa, I.C.V.D. Evaluating FMX-101 as a promising therapeutic for the treatment of acne. Expert Opin. Pharmacother. 2020, 21, 741–746. [Google Scholar] [CrossRef]
- Farrah, G.; Tan, E. The use of oral antibiotics in treating acne vulgaris: A new approach. Dermatol. Ther. 2016, 29, 377–384. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Pimentel-Elardo, S.M.; Hanora, A.; Radwan, M.; Abou-El-Ela, S.H.; Ahmed, S.; Hentschel, U. Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes. Mar. Drugs 2010, 8, 399–412. [Google Scholar] [CrossRef]
- Mary, T.R.J.; Kannan, R.R.; Iniyan, A.M.; Ramachandran, D.; Vincent, S.G.P. Cell wall distraction and biofilm inhibition of marine Streptomyces derived angucycline in methicillin resistant Staphylococcus aureus. Microb. Pathog. 2020, 150, 104712. [Google Scholar] [CrossRef]
- O’Neill, J. The Review on Antimicrobial Resistance. Rev. Laryngol. Otol. Rhinol. 2016, 136, 29–31. [Google Scholar]
- World Health Organization (WHO). La Escasez Mundial de Antibióticos Innovadores Favorece La Aparición y Propagación de La Farmacorresistencia. Available online: https://www.who.int/es/news/item/15-04-2021-global-shortage-of-innovative-antibiotics-fuels-emergence-and-spread-of-drug-resistance (accessed on 28 January 2022).
- World Health Organization. Lack of New Antibiotics Threatens Global Efforts to Contain Drug-Resistant Infections. Available online: https://www.who.int/news-room/detail/17-01-2020-lack-of-new-antibiotics-threatens-global-efforts-to-contain-drug-resistant-infections (accessed on 29 April 2020).
- Szántó, M.; Dózsa, A.; Antal, D.; Szabó, K.; Kemény, L.; Bai, P. Targeting the gut-skin axis—Probiotics as new tools for skin disorder management? Exp. Dermatol. 2019, 28, 1210–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris. Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandwein, M.; Steinberg, D.; Meshner, S. Microbial biofilms and the human skin microbiome. NPJ Biofilms Microb. 2016, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A Review of the Microbial Production of Bioactive Natural Products and Biologics. Front. Microbiol. 2019, 10, 1404. [Google Scholar] [CrossRef] [Green Version]
- Siddharth, S.; Vittal, R.R. Isolation, Characterization, and Structural Elucidation of 4-Methoxyacetanilide from Marine Actinobacteria Streptomyces sp. SCA29 and Evaluation of Its Enzyme Inhibitory, Antibacterial, and Cytotoxic Potential. Arch. Microbiol. 2019, 201, 737–746. [Google Scholar] [CrossRef]
- Paderog, M.J.V.; Suarez, A.F.L.; Sabido, E.M.; Low, Z.J.; Saludes, J.P.; Dalisay, D.S. Anthracycline Shunt Metabolites From Philippine Marine Sediment-Derived Streptomyces Destroy Cell Membrane Integrity of Multidrug-Resistant Staphylococcus aureus. Front. Microbiol. 2020, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- Dholakiya, R.N.; Kumar, R.; Mishra, A.; Mody, K.H.; Jha, B. Antibacterial and Antioxidant Activities of Novel Actinobacteria Strain Isolated from Gulf of Khambhat, Gujarat. Front. Microbiol. 2017, 8, 2420. [Google Scholar] [CrossRef]
- Aljelawi, R.O.; Kadhem, M.F. Production, Purification, and Characterization of Bioactive Metabolites Produced from Rare Actinobacteria Pseudonocardia Alni. Asian J. Pharm. Clin. Res. 2016, 9, 264. [Google Scholar] [CrossRef]
- Gavriilidou, A.; Mackenzie, T.; Sánchez, P.; Tormo, J.; Ingham, C.; Smidt, H.; Sipkema, D. Bioactivity Screening and Gene-Trait Matching across Marine Sponge-Associated Bacteria. Mar. Drugs 2021, 19, 75. [Google Scholar] [CrossRef]
- Sabido, E.M.; Tenebro, C.P.; Suarez, A.F.L.; Ong, S.D.C.; Trono, D.J.V.L.; Amago, D.S.; Evangelista, J.J.E.; Reynoso, A.M.Q.; Villalobos, I.G.M.; Alit, L.D.D.; et al. Marine Sediment-Derived Streptomyces Strain Produces Angucycline Antibiotics against Multidrug-Resistant Staphylococcus aureus Harboring SCCmec Type 1 Gene. J. Mar. Sci. Eng. 2020, 8, 734. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, A.A.; Gromek, S.M.; Balunas, M.J. Upregulation and Identification of Antibiotic Activity of a Marine-Derived Streptomyces sp. via Co-Cultures with Human Pathogens. Mar. Drugs 2017, 15, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.J.; An, J.S.; Bae, E.S.; Cho, E.; Hwang, S.; Nam, S.J.; Oh, D.C. Ligiamycins A and B, Decalin-Amino-Maleimides from the Co-Culture of Streptomyces sp. and Achromobacter sp. Isolated from the Marine Wharf Roach, Ligia Exotica. Mar. Drugs 2022, 20, 93. [Google Scholar] [CrossRef] [PubMed]
- Hifnawy, M.S.; Hassan, H.M.; Mohammed, R.; Fouda, M.M.; Sayed, A.M.; Hamed, A.A.; Abouzid, S.F.; Rateb, M.E.; Alhadrami, H.A.; Abdelmohsen, U.R. Induction of Antibacterial Metabolites by Co-Cultivation of Two Red-Sea-Sponge-Associated Actinomycetes Micromonospora sp. UR56 and Actinokinespora sp. EG49. Mar. Drugs 2020, 18, 243. [Google Scholar] [CrossRef]
- Yu, M.; Li, Y.; Banakar, S.P.; Liu, L.; Shao, C.; Li, Z.; Wang, C. New Metabolites from the Co-culture of Marine-Derived Actinomycete Streptomyces rochei MB037 and Fungus Rhinocladiella similis 35. Front. Microbiol. 2019, 10, 915. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Qian, S.; Qi, S.; Jiang, W.; Dong, K. Culture Medium Optimization and Active Compounds Investigation of an Anti-Quorum Sensing Marine Actinobacterium Nocardiopsis Dassonvillei JS106. Microbiology 2021, 90, 112–123. [Google Scholar] [CrossRef]
- Arasu, M.V.; Duraipandiyan, V.; Ignacimuthu, S. Antibacterial and antifungal activities of polyketide metabolite from marine Streptomyces sp. AP-123 and its cytotoxic effect. Chemosphere 2013, 90, 479–487. [Google Scholar] [CrossRef]
- Zhang, Z.; Sibero, M.T.; Kai, A.; Fukaya, K.; Urabe, D.; Igarashi, Y. TMKS8A, an Antibacterial and Cytotoxic Chlorinated α-Lapachone, from a Sea Slug-Derived Actinomycete of the Genus Streptomyces. J. Antibiot. 2021, 74, 464–469. [Google Scholar] [CrossRef]
- Newaz, A.W.; Yong, K.; Lian, X.Y.; Zhang, Z. Streptoindoles A–D, Novel Antimicrobial Indole Alkaloids from the Marine-Associated Actinomycete Streptomyces Sp. ZZ1118. Tetrahedron 2022, 104, 132598. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Lage, O.M.; Pérez-Moreno, G.; Martinez, I.G.; Tormo, J.R.; et al. Bioactivities and Extract Dereplication of Actinomycetales Isolated from Marine Sponges. Front. Microbiol. 2019, 10, 727. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, Y.; Refaat, J.; Abdelmohsen, U.R.; Othman, E.M.; Stopper, H.; Fouad, M.A. Metabolomic profiling and biological investigation of the marine sponge-derived bacterium Rhodococcus sp. UA13. Phytochem. Anal. 2018, 29, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-B.; Jensen, P.R.; Fenical, W. Cytotoxic and Antimicrobial Napyradiomycins from Two Marine-Derived Streptomyces Strains. Eur. J. Org. Chem. 2013, 2013, 3751–3757. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.C.; Prieto-Davo, A.; Jensen, P.R.; Fenical, W. ChemInform Abstract: The Marinopyrroles, Antibiotics of an Unprecedented Structure Class from a Marine Streptomyces sp. ChemInform 2008, 10, 629–631. [Google Scholar] [CrossRef]
- Shen, X.; Wang, X.; Huang, T.; Deng, Z.; Lin, S. Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173. Biomolecules 2020, 10, 1187. [Google Scholar] [CrossRef]
- Kock, I.; Maskey, R.P.; Biabani, M.A.F.; Helmke, E.; Laatsch, H. 1-Hydroxy-1-norresistomycin and Resistoflavin Methyl Ether: New Antibiotics from Marine-derived Streptomycetes. J. Antibiot. 2005, 58, 530–534. [Google Scholar] [CrossRef]
- Yi, W.; Li, Q.; Song, T.; Chen, L.; Li, X.-C.; Zhang, Z.; Lian, X.-Y. Isolation, structure elucidation, and antibacterial evaluation of the metabolites produced by the marine-sourced Streptomyces sp. ZZ820. Tetrahedron 2019, 75, 1186–1193. [Google Scholar] [CrossRef]
- Nandhagopal, S.; Iniyan, A.M.; Kannan, R.R.; Vincent, S.G.P. In Vivo Evaluation of Anti-MRSA Compound from Streptomyces Collinus ICN1 in Zebrafish Embryos. Indian J. Geo-Marine Sci. 2017, 46, 1155–1161. [Google Scholar]
- Jiao, W.-H.; Yuan, W.; Li, Z.-Y.; Li, J.; Li, L.; Sun, J.-B.; Gui, Y.-H.; Wang, J.; Ye, B.-P.; Lin, H.-W. Anti-MRSA actinomycins D1-D4 from the marine sponge-associated Streptomyces sp. LHW52447. Tetrahedron 2018, 74, 5914–5919. [Google Scholar] [CrossRef]
- Yang, L.; Hou, L.; Li, H.; Li, W. Antibiotic angucycline derivatives from the deepsea-derived Streptomyces lusitanus. Nat. Prod. Res. 2019, 34, 3444–3450. [Google Scholar] [CrossRef]
- Djinni, I.; Defant, A.; Kecha, M.; Mancini, I. Metabolite Profile of Marine-Derived Endophytic Streptomyces Sundarbansensis WR1L1S8 by Liquid Chromatography-Mass Spectrometry and Evaluation of Culture Conditions on Antibacterial Activity and Mycelial Growth. J. Appl. Microbiol. 2014, 116, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Rajan, B.M.; Kannabiran, K. Antibiotic Potency of 2,4-Dichloro-5-Sulfamoyl Benzoic Acid Extracted from Marine Bacterium Streptomyces sp. VITBRK3 against Methicillin Resistant Staphylococcus aureus. Pharm. Lett. 2015, 7, 244–252. [Google Scholar]
- Song, Y.; Yang, J.; Yu, J.; Li, J.; Yuan, J.; Wong, N.-K.; Ju, J. Chlorinated bis-indole alkaloids from deep-sea derived Streptomyces sp. SCSIO 11791 with antibacterial and cytotoxic activities. J. Antibiot. 2020, 73, 542–547. [Google Scholar] [CrossRef] [PubMed]
- León, J.; Aponte, J.J.; Rojas, R.; Cuadra, D.; Ayala, N.; Tomás, G.; Guerrero, M. Estudio de actinomicetos marinos aislados de la costa central del Perú y su actividad antibacteriana frente a Staphylococcus aureus Meticilina Resistentes y Enterococcus faecalis Vancomicina Resistentes. Rev. Peru. Med. Exp. Salud Publ. 2011, 28, 237–246. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Ghilan, A.-K.M.; Esmail, G.A.; Arasu, M.V.; Duraipandiyan, V.; Ponmurugan, K. Bioactivity assessment of the Saudi Arabian Marine Streptomyces sp. Al-Dhabi-90, metabolic profiling and its in vitro inhibitory property against multidrug resistant and extended-spectrum beta-lactamase clinical bacterial pathogens. J. Infect. Public Health 2019, 12, 549–556. [Google Scholar] [CrossRef]
- Shao, M.; Ma, J.; Li, Q.; Ju, J. Identification of the Anti-Infective Aborycin Biosynthetic Gene Cluster from Deep-Sea-Derived Streptomyces sp. SCSIO ZS0098 Enables Production in a Heterologous Host. Mar. Drugs 2019, 17, 127. [Google Scholar] [CrossRef] [Green Version]
- Padmanaban, V.P.; Verma, P.; Venkatabaskaran, S.; Keppayan, T.; Gopal, D.; Sekar, A.K.; Ramalingam, K. Antimicrobial potential and taxonomic investigation of piezotolerant Streptomyces sp. NIOT-Ch-40 isolated from deep-sea sediment. World J. Microbiol. Biotechnol. 2017, 33, 27. [Google Scholar] [CrossRef]
- Song, Y.; Liu, G.; Li, J.; Huang, H.; Zhang, X.; Zhang, H.; Ju, J. Cytotoxic and Antibacterial Angucycline- and Prodigiosin- Analogues from the Deep-Sea Derived Streptomyces sp. SCSIO 11594. Mar. Drugs 2015, 13, 1304–1316. [Google Scholar] [CrossRef]
- Lacret, R.; Oves-Costales, D.; Pérez-Victoria, I.; de la Cruz, M.; Díaz, C.; Vicente, F.; Genilloud, O.; Reyes, F. MDN-0171, a new medermycin analogue from Streptomyces albolongus CA-186053. Nat. Prod. Res. 2018, 33, 66–73. [Google Scholar] [CrossRef]
- Wu, C.; Tan, Y.; Gan, M.; Wang, Y.; Guan, Y.; Hu, X.; Zhou, H.; Shang, X.; You, X.; Yang, Z.; et al. Identification of Elaiophylin Derivatives from the Marine-Derived Actinomycete Streptomyces sp. 7-145 Using PCR-Based Screening. J. Nat. Prod. 2013, 76, 2153–2157. [Google Scholar] [CrossRef]
- Chung, B.; Kwon, O.-S.; Shin, J.; Oh, K.-B. Antibacterial Activity and Mode of Action of Lactoquinomycin A from Streptomyces bacillaris. Mar. Drugs 2020, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Akhter, N.; Liu, Y.; Auckloo, B.N.; Shi, Y.; Wang, K.; Chen, J.; Wu, X.; Wu, B. Stress-Driven Discovery of New Angucycline-Type Antibiotics from a Marine Streptomyces pratensis NA-ZhouS1. Mar. Drugs 2018, 16, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, X.; Gong, T.; Liu, F.; Zhang, P.C.; Zhou, W.Q.; Li, Y.; Zhu, P. A New Analogue of Echinomycin and a New Cyclic Dipeptide from a Marine-Derived Streptomyces Sp. LS298. Mar. Drugs 2015, 13, 6947–6961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Wang, C.; Gui, P.; Liu, H.; Khalaf, S.M.H.; Elsayed, E.A.; Wadaan, M.A.M.; Hozzein, W.N.; Zhu, W. Identification, Bioactivity, and Productivity of Actinomycins from the Marine-Derived Streptomyces heliomycini. Front. Microbiol. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Ravikumar, S.; Gnanadesigan, M.; Saravanan, A.; Monisha, N.; Brindha, V.; Muthumari, S. Antagonistic properties of seagrass associated Streptomyces sp. RAUACT-1: A source for anthraquinone rich compound. Asian Pac. J. Trop. Med. 2012, 5, 887–890. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, L.; Chai, W.; Lian, X.-Y.; Zhang, Z. A unique indolizinium alkaloid streptopertusacin A and bioactive bafilomycins from marine-derived Streptomyces sp. HZP-2216E. Phytochemistry 2017, 144, 119–126. [Google Scholar] [CrossRef]
- Niu, S.; Li, S.; Chen, Y.; Tian, X.; Zhang, H.; Zhang, G.; Zhang, W.; Yang, X.; Zhang, S.; Ju, J.; et al. Lobophorins E and F, new spirotetronate antibiotics from a South China Sea-derived Streptomyces sp. SCSIO 01127. J. Antibiot. 2011, 64, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Norouzi, H.; Khorasgani, M.R.; Danesh, A. Anti-MRSA activity of a bioactive compound produced by a marine Streptomyces and its optimization using statistical experimental design. Iran. J. Basic Med. Sci. 2019, 22, 1073–1084. [Google Scholar] [CrossRef]
- Dong, Y.; Ding, W.; Sun, C.; Ji, X.; Ling, C.; Zhou, Z.; Chen, Z.; Chen, X.; Ju, J. Julichrome Monomers from Marine Gastropod Mollusk-Associated Streptomyces and Stereochemical Revision of Julichromes Q(3.5) and Q(3.3). Chem. Biodivers. 2020, 17, e2000057. [Google Scholar] [CrossRef]
- Luo, M.; Tang, L.; Dong, Y.; Huang, H.; Deng, Z.; Sun, Y. Antibacterial natural products lobophorin L and M from the marine-derived Streptomyces sp. 4506. Nat. Prod. Res. 2020, 35, 5581–5587. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, L.; Ye, X.; Anjum, K.; Lian, X.Y.; Zhang, Z. New streptophenazines from marine Streptomyces sp. 182SMLY. Nat. Prod. Res. 2017, 31, 411–417. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wang, M.; Tan, Y.; Chunling, X.; He, H.; Xiao, C.; You, X.; Wang, Y.; Gan, M. Neo-actinomycins A and B, natural actinomycins bearing the 5H-oxazolo[4,5-b]phenoxazine chromophore, from the marine-derived Streptomyces sp. IMB094. Sci. Rep. 2017, 7, 3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Huang, H.; Chen, Y.; Ding, J.; Zhang, Y.; Sun, A.; Zhang, W.; Ju, J. Cytotoxic and Antibacterial Marfuraquinocins from the Deep South China Sea-Derived Streptomyces niveus SCSIO 3406. J. Nat. Prod. 2013, 76, 2263–2268. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.W.; Torres, J.P.; Tun, J.O.; Flores, M.S.; Forteza, I.; Rosenberg, G.; Haygood, M.G.; Schmidt, E.W.; Concepcion, G.P. Synergistic anti-methicillin-resistant Staphylococcus aureus (MRSA) activity and absolute stereochemistry of 7,8-dideoxygriseorhodin C. J. Antibiot. 2020, 73, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Pérez-Victoria, I.; Martín, J.; Otero, L.; Palacios-Gutiérrez, J.J.; Fernández, J.; Mohamedi, Y.; Fontanil, T.; Salmón, M.; et al. Desertomycin G, a New Antibiotic with Activity against Mycobacterium tuberculosis and Human Breast Tumor Cell Lines Produced by Streptomyces althioticus MSM3, Isolated from the Cantabrian Sea Intertidal Macroalgae Ulva sp. Mar. Drugs 2019, 17, 114. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, G.; Santhi, V.S.; Jebakumar, S.R.D. Antimicrobial potential of phylogenetically unique actinomycete, Streptomyces sp. JRG-04 from marine origin. Biologicals 2014, 42, 305–311. [Google Scholar] [CrossRef]
- Wu, Z.; Li, S.; Li, J.; Chen, Y.; Saurav, K.; Zhang, Q.; Zhang, H.; Zhang, W.; Zhang, W.; Zhang, S.; et al. Antibacterial and Cytotoxic New Napyradiomycins from the Marine-Derived Streptomyces sp. SCSIO 10428. Mar. Drugs 2013, 11, 2113–2125. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.C.; Kauffman, C.; Jensen, P.; Fenical, W. Structures, Reactivities, and Antibiotic Properties of the Marinopyrroles A−F. J. Org. Chem. 2010, 75, 3240–3250. [Google Scholar] [CrossRef] [Green Version]
- Haste, N.M.; Farnaes, L.; Perera, V.R.; Fenical, W.; Nizet, V.; Hensler, M.E. Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus. Mar. Drugs 2011, 9, 680–689. [Google Scholar] [CrossRef]
- Djinni, I.; Defant, A.; Kecha, M.; Mancini, I. Antibacterial Polyketides from the Marine Alga-Derived Endophitic Streptomyces sundarbansensis: A Study on Hydroxypyrone Tautomerism. Mar. Drugs 2013, 11, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Maloney, K.N.; Nam, S.-J.; Haste, N.M.; Raju, R.; Aalbersberg, W.; Jensen, P.R.; Nizet, V.; Hensler, M.E.; Fenical, W. Fijimycins A–C, three antibacterial etamycin-class depsipeptides from a marine-derived Streptomyces sp. Bioorganic Med. Chem. 2011, 19, 6557–6562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haste, N.M.; Perera, V.R.; Maloney, K.N.; Tran, D.N.; Jensen, P.; Fenical, W.; Nizet, V.; Hensler, E.M. Activity of the streptogramin antibiotic etamycin against methicillin-resistant Staphylococcus aureus. J. Antibiot. 2010, 63, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Furumai, T.; Eto, K.; Sasaki, T.; Higuchi, H.; Onaka, H.; Saito, N.; Fujita, T.; Naoki, H.; Igarashi, Y. TPU-0037-A, B, C and D, Novel Lydicamycin Congeners with Anti-MRSA Activity from Streptomyces platensis TP-A0598. J. Antibiot. 2002, 55, 873–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, H.M.; Degen, D.; Jang, K.H.; Ebright, R.H.; Fenical, W. Salinamide F, new depsipeptide antibiotic and inhibitor of bacterial RNA polymerase from a marine-derived streptomyces sp. J. Antibiot. 2015, 68, 206–209. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Xu, Y.; McConnell, O.; Liu, L.; Li, Y.; Qi, S.; Huang, X.; Qian, P. Two Antimycin A Analogues from Marine-Derived Actinomycete Streptomyces lusitanus. Mar. Drugs 2012, 10, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, M.-J.; Hillman, P.F.; Lee, J.; Hwang, S.; Lee, E.-Y.; Cha, S.-S.; Yang, I.; Oh, D.-C.; Nam, S.-J.; Fenical, W. Antibacterial Meroterpenoids, Merochlorins G-J from the Marine Bacterium Streptomyces sp. Mar. Drugs 2021, 19, 618. [Google Scholar] [CrossRef]
- Yang, W.Z.; Liang, G.J.; Sun, Y.; Gong, Z.J. Bioactive Secondary Metabolites from Marine Streptomyces Griseorubens F8: Isolation, Identification and Biological Activity Assay. J. Mar. Sci. Eng. 2021, 9, 978. [Google Scholar] [CrossRef]
- Qureshi, K.A.; Bholay, A.D.; Rai, P.K.; Mohammed, H.A.; Khan, R.A.; Azam, F.; Jaremko, M.; Emwas, A.-H.; Stefanowicz, P.; Waliczek, M.; et al. Isolation, Characterization, Anti-MRSA Evaluation, and in-Silico Multi-Target Anti-Microbial Validations of Actinomycin X(2) and Actinomycin D Produced by Novel Streptomyces Smyrnaeus UKAQ_23. Sci. Rep. 2021, 11, 14539. [Google Scholar] [CrossRef]
- Thi, D.P.; Mai, H.D.T.; Cao, D.D.; Thi, Q.V.; Nguyen, M.A.; Le Thi, H.M.; Tran, D.T.; Chau, V.M.; Pham, V.C. Novel 1,3-Benzodioxole From Marine-Derived Actinomycete in East Vietnam Sea. Nat. Prod. Commun. 2020, 15, 1934578X20920042. [Google Scholar] [CrossRef]
- Song, Y.; Li, Q.; Liu, X.; Chen, Y.; Zhang, Y.; Sun, A.; Zhang, W.; Zhang, J.; Ju, J. Cyclic Hexapeptides from the Deep South China Sea-Derived Streptomyces scopuliridis SCSIO ZJ46 Active Against Pathogenic Gram-Positive Bacteria. J. Nat. Prod. 2014, 77, 1937–1941. [Google Scholar] [CrossRef]
- Kunz, A.L.; Labes, A.; Wiese, J.; Bruhn, T.; Bringmann, G.; Imhoff, J.F. Nature’s Lab for Derivatization: New and Revised Structures of a Variety of Streptophenazines Produced by a Sponge-Derived Streptomyces Strain. Mar. Drugs 2014, 12, 1699–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.-L.; Xu, Y.; Han, Z.; Li, Y.-X.; Lu, L.; Lai, P.-Y.; Zhong, J.-L.; Guo, X.-R.; Zhang, X.-X.; Qian, P.-Y. Four New Antibacterial Xanthones from the Marine-Derived Actinomycetes Streptomyces caelestis. Mar. Drugs 2012, 10, 2571–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iniyan, A.M.; Sudarman, E.; Wink, J.; Kannan, R.R.; Vincent, S.G.P. Ala-geninthiocin, a new broad spectrum thiopeptide antibiotic, produced by a marine Streptomyces sp. ICN19. J. Antibiot. 2018, 72, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, H.S.; Shahnavaz, B.; Makhdoumi, A.; Iranshahy, M. Evaluating the effect of various bacterial consortia on antibacterial activity of marine Streptomyces sp. AC117. Biocontrol Sci. Technol. 2021, 31, 1248–1266. [Google Scholar] [CrossRef]
- Kumar, S.S.; Rao, K.V.B. In–vitro antimicrobial activity of marine actinobacteria against multidrug resistance Staphylococcus aureus. Asian Pac. J. Trop. Biomed. 2012, 2, S1802–S1807. [Google Scholar] [CrossRef] [Green Version]
- Sabido, E.; Tenebro, C.; Trono, D.; Vicera, C.; Leonida, S.; Maybay, J.; Reyes-Salarda, R.; Amago, D.; Aguadera, A.; Octaviano, M.; et al. Insights into the Variation in Bioactivities of Closely Related Streptomyces Strains from Marine Sediments of the Visayan Sea against ESKAPE and Ovarian Cancer. Mar. Drugs 2021, 19, 441. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Applications of Prodigiosin Extracted from Marine Red Pigmented Bacteria Zooshikella sp. and Actinomycete Streptomyces sp. Microorganisms 2020, 8, 556. [Google Scholar] [CrossRef]
- Kurnianto, M.A.; Kusumaningrum, H.D.; Lioe, H.N.; Chasanah, E. Antibacterial and Antioxidant Potential of Ethyl Acetate Extract from Streptomyces AIA12 and AIA17 Isolated from Gut of Chanos chanos. J. Biol. Divers. 2021, 22, 813. [Google Scholar] [CrossRef]
- Shin, H.J.; Lee, H.-S.; Lee, D.-S. The synergistic antibacterial activity of 1-acetyl-beta-carboline and beta-lactams against methicillin-resistant Staphylococcus aureus (MRSA). J. Microbiol. Biotechnol. 2010, 20, 501–505. [Google Scholar]
- Kim, M.C.; Li, Z.; Cullum, R.; Molinski, T.F.; Eid, M.A.G.; Hebishy, A.M.S.; Faraag, A.H.I.; Abdel Moneim, A.E.; Abdelfattah, M.S.; Fenical, W. Chlororesistoflavins A and B, Chlorinated Benzopyrene Antibiotics Produced by the Marine-Derived Actinomycete Streptomyces Sp. Strain EG32. J. Nat. Prod. 2022, 85, 270–275. [Google Scholar] [CrossRef]
- Shanthi, J.; Senthil, A.; Gopikrishnan, V.; Balagurunathan, R. Characterization of a Potential β-Lactamase Inhibitory Metabolite from a Marine Streptomyces sp. PM49 Active Against Multidrug-Resistant Pathogens. Appl. Biochem. Biotechnol. 2015, 175, 3696–3708. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhang, Q.; Zhu, Y.; Zhang, L.; Zhang, W.; Ma, L.; Zhang, H.; Zhang, C. Proximicins F and G and Diproximicin A: Aminofurans from the Marine-Derived Verrucosispora sp. SCSIO 40062 by Overexpression of PPtase Genes. J. Nat. Prod. 2020, 83, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Xie, F.; Ren, B.; Wang, Q.; Wang, J.; Wang, Q.; Abdel-Mageed, W.M.; Liu, M.; Han, J.; Oyeleye, A.; et al. Anti-MRSA and anti-TB metabolites from marine-derived Verrucosispora sp. MS100047. Appl. Microbiol. Biotechnol. 2016, 100, 7437–7447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xie, Q.; Sun, C.; Tian, X.P.; Gui, C.; Qin, X.; Ju, J. Cytotoxic Kendomycins Containing the Carbacylic Ansa Scaffold from the Marine-Derived Verrucosispora Sp. SCSIO 07399. Nat. Prod. 2019, 82, 3366–3371. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-H.; Lian, Y.-Y.; Fang, D.-S.; Chen, L.; Jia, J.; Zhang, W.-L.; Lin, R.; Xie, Y.; Bi, H.-K.; Jiang, H. Identification and antimicrobial properties of a new alkaloid produced by marine-derived Verrucosispora sp. FIM06-0036. Nat. Prod. Res. 2019, 35, 4211–4217. [Google Scholar] [CrossRef]
- Zhang, Y.; Adnani, N.; Braun, D.R.; Ellis, G.A.; Barns, K.J.; Parker-Nance, S.; Guzei, I.A.; Bugni, T.S. Micromonohalimanes A and B: Antibacterial Halimane-Type Diterpenoids from a Marine Micromonospora Species. J. Nat. Prod. 2016, 79, 2968–2972. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, Z.; Li, S.; Lu, Y.; Chen, Y.; Zhang, H.; Zhang, G.; Zhu, Y.; Zhang, G.; Zhang, W.; et al. Fluostatins I–K from the South China Sea-Derived Micromonospora rosaria SCSIO N160. J. Nat. Prod. 2012, 75, 1937–1943. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, Q.; Zhu, Y.; Nie, F.; Wu, Z.; Yang, C.; Zhang, L.; Tian, X.; Zhang, C. Isolation, structure elucidation and biosynthesis of benzo[b]fluorene nenestatin A from deep-sea derived Micromonospora echinospora SCSIO 04089. Tetrahedron 2017, 73, 3585–3590. [Google Scholar] [CrossRef]
- Siddharth, S.; Rai, V.R. Isolation and characterization of bioactive compounds with antibacterial, antioxidant and enzyme inhibitory activities from marine-derived rare actinobacteria, Nocardiopsis sp. SCA21. Microb. Pathog. 2019, 137, 103775. [Google Scholar] [CrossRef]
- Xu, D.; Nepal, K.K.; Chen, J.; Harmody, D.; Zhu, H.; McCarthy, P.J.; Wright, A.E.; Wang, G. Nocardiopsistins A-C: New angucyclines with anti-MRSA activity isolated from a marine sponge-derived Nocardiopsis sp. HB-J378. Synth. Syst. Biotechnol. 2018, 3, 246–251. [Google Scholar] [CrossRef]
- Yang, N.; Song, F. Bioprospecting of Novel and Bioactive Compounds from Marine Actinomycetes Isolated from South China Sea Sediments. Curr. Microbiol. 2017, 75, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Rajivgandhi, G.; Vijayan, R.; Kannan, M.; Santhanakrishnan, M.; Manoharan, N. Molecular characterization and antibacterial effect of endophytic actinomycetes Nocardiopsis sp. GRG1 (KT235640) from brown algae against MDR strains of uropathogens. Bioact. Mater. 2016, 1, 140–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunga, M.J.; Teisan, S.; Tsueng, G.; Macherla, V.R.; Lam, K.S. Seawater requirement for the production of lipoxazolidinones by marine actinomycete strain NPS8920. J. Ind. Microbiol. Biotechnol. 2008, 35, 761–765. [Google Scholar] [CrossRef] [PubMed]
- McArthur, K.A.; Mitchell, S.S.; Tsueng, G.; Rheingold, A.; White, D.J.; Grodberg, J.; Lam, K.S.; Potts, B.C.M. Lynamicins A−E, Chlorinated Bisindole Pyrrole Antibiotics from a Novel Marine Actinomycete. J. Nat. Prod. 2008, 71, 1732–1737. [Google Scholar] [CrossRef]
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Pérez-Victoria, I.; Otero, L.; Fernández, J.; Palacios, J.J.; Martín, J.; de la Cruz, M.; Díaz, C.; Vicente, F.; et al. Branimycins B and C, Antibiotics Produced by the Abyssal Actinobacterium Pseudonocardia Carboxydivorans M-227. J. Nat. Prod. 2017, 80, 569–573. [Google Scholar] [CrossRef]
- Palomo, S.; González, I.; de la Cruz, M.; Martín, J.; Tormo, J.R.; Anderson, M.; Hill, R.T.; Vicente, F.; Reyes, F.; Genilloud, O. Sponge-Derived Kocuria and Micrococcus spp. as Sources of the New Thiazolyl Peptide Antibiotic Kocurin. Mar. Drugs 2013, 11, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Ellis, G.; Wyche, T.P.; Fry, C.G.; Braun, D.R.; Bugni, T.S. Solwaric Acids A and B, Antibacterial Aromatic Acids from a Marine Solwaraspora sp. Mar. Drugs 2014, 12, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Prasad, P.; Subramani, R.; Aalbersberg, W. Production and Purification of a Bioactive Substance against Multi-Drug Resistant Human Pathogens from the Marine-Sponge-Derived Salinispora Sp. Asian Pac. J. Trop. Biomed. 2014, 4, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Yi, W.; Ge, H.; Zhang, Z.; Wu, B. Bioactive Streptoglutarimides A–J from the Marine-Derived Streptomyces Sp. ZZ741. J. Nat. Prod. 2019, 82, 2800–2808. [Google Scholar] [CrossRef]
- Zhou, B.; Huang, Y.; Zhang, H.-J.; Li, J.-Q.; Ding, W.-J. Nitricquinomycins A-C, uncommon naphthopyrrolediones from the Streptomyces sp. ZS-A45. Tetrahedron 2019, 75, 3958–3961. [Google Scholar] [CrossRef]
- Carretero-Molina, D.; Ortiz-López, F.J.; Martín, J.; Oves-Costales, D.; Díaz, C.; de la Cruz, M.; Cautain, B.; Vicente, F.; Genilloud, O.; Reyes, F. New Napyradiomycin Analogues from Streptomyces sp. Strain CA-271078. Mar. Drugs 2020, 18, 22–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sujatha, P.; Raju, K.B.; Ramana, T. Studies on a new marine streptomycete BT-408 producing polyketide antibiotic SBR-22 effective against methicillin resistant Staphylococcus aureus. Microbiol. Res. 2005, 160, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.-Q.; Zhang, S.-Y.; Wang, N.; Li, Z.-L.; Hua, H.-M.; Hu, J.-C.; Wang, S.-J. New Spirotetronate Antibiotics, Lobophorins H and I, from a South China Sea-Derived Streptomyces sp. 12A35. Mar. Drugs 2013, 11, 3891–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ye, X.; Chai, W.; Lian, X.-Y.; Zhang, Z. New Metabolites and Bioactive Actinomycins from Marine-Derived Streptomyces sp. ZZ338. Mar. Drugs 2016, 14, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Wang, M.; Wu, C.; Tan, Y.; Li, J.; Hao, X.; Duan, Y.; Guan, Y.; Shang, X.; Wang, Y.; et al. Identification and Proposed Relative and Absolute Configurations of Niphimycins C–E from the Marine-Derived Streptomyces sp. IMB7-145 by Genomic Analysis. J. Nat. Prod. 2018, 81, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.D.; Do, T.Q.; Doan Thi Mai, H.; Vu Thi, Q.; Nguyen, M.A.; Le Thi, H.M.; Tran, D.T.; Chau, V.M.; Cong Thung, D.; Pham, V.C. Antimicrobial lavandulylated flavonoids from a sponge-derived actinomycete. Nat. Prod. Res. 2020, 34, 413–420. [Google Scholar] [CrossRef]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Wang, J. Cytotoxic Anthracycline and Antibacterial Tirandamycin Analogues from a Marine-Derived Streptomyces Sp. SCSIO 41399. J. Antibiot. 2019, 72, 45–49. [Google Scholar] [CrossRef]
- Saurav, K.; Kannabiran, K. In vitro activity of 5-(2,4-dimethylbenzyl) pyrrolidin-2-one extracted from marine Streptomyces VITSVK5 spp. against fungal and bacterial human pathogens. Rev. Iberoam. Micol. 2012, 29, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Kwon, O.-S.; Chung, B.; Lee, J.; Sun, J.; Shin, J.; Oh, K.-B. Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus. Mar. Drugs 2020, 18, 522. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chai, W.; Zhu, R.; Song, T.; Zhang, Z.; Lian, X.-Y. Streptopyrazinones A−D, rare metabolites from marine-derived Streptomyces sp. ZZ446. Tetrahedron 2018, 74, 2100–2106. [Google Scholar] [CrossRef]
- Lacret, R.; Pérez-Victoria, I.; Oves-Costales, D.; de la Cruz, M.; Domingo, E.; Martín, J.; Díaz, C.; Vicente, F.; Genilloud, O.; Reyes, F. MDN-0170, a New Napyradiomycin from Streptomyces sp. Strain CA-271078. Mar. Drugs 2016, 14, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaala, L.A.; Youssef, D.T.A.; Alzughaibi, T.A.; Elhady, S.S. Antimicrobial Chlorinated 3-Phenylpropanoic Acid Derivatives from the Red Sea Marine Actinomycete Streptomyces coelicolor LY001. Mar. Drugs 2020, 18, 450. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, P.; Ren, B.; Song, Z.; Zhu, G.; He, W.; Zhang, J.; Oyeleye, A.; Dai, H.; Zhang, L.; et al. Antibacterial polyene-polyol macrolides and cyclic peptides from the marine-derived Streptomyces sp. MS110128. Appl. Microbiol. Biotechnol. 2021, 105, 4975–4986. [Google Scholar] [CrossRef] [PubMed]
- Setiawati, S.; Nuryastuti, T.; Sholikhah, E.N.; Lisdiyanti, P.; Pratiwi, S.U.T.; Sulistiyani, T.R.; Mustofa, M. The Potency of Actinomycetes Extracts Isolated from Pramuka Island, Jakarta, Indonesia as Antimicrobial Agents. Biodivers. J. Biol. Divers. 2021, 22, 150933. [Google Scholar] [CrossRef]
- Dalisay, D.; Williams, D.E.; Wang, X.L.; Centko, R.; Chen, J.; Andersen, R.J. Marine Sediment-Derived Streptomyces Bacteria from British Columbia, Canada Are a Promising Microbiota Resource for the Discovery of Antimicrobial Natural Products. PLoS ONE 2013, 8, e77078. [Google Scholar] [CrossRef]
- Uzair, B.; Menaa, F.; Khan, B.A.; Mohammad, F.V.; Ahmad, V.U.; Djeribi, R.; Menaa, B. Isolation, purification, structural elucidation and antimicrobial activities of kocumarin, a novel antibiotic isolated from actinobacterium Kocuria marina CMG S2 associated with the brown seaweed Pelvetia canaliculata. Microbiol. Res. 2018, 206, 186–197. [Google Scholar] [CrossRef]
- Kwon, H.C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Marinomycins A-D, Antitumor-Antibiotics of a New Structure Class from a Marine Actinomycete of the Recently Discovered Genus “Marinispora”. J. Am. Chem. Soc. 2006, 128, 1622–1632. [Google Scholar] [CrossRef]
- Chen, M.H.; Zhang, W.L.; Chen, L.; Lin, R.; Xie, Y.; Fang, D.S.; Jiang, H.; Lian, Y.-Y. Isolation, Purification and Identification of Two New Alkaloids Metabolites from Marine-Derived Verrucosispora Sp. FIM06025. Nat. Prod. Res. 2019, 33, 2897–2903. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Iniyan, A.M.; Mary, T.R.J.; Joseph, F.-J.R.S.; Kannan, R.R.; Vincent, S.G.P. Cell wall distracting anti-Methicillin-resistant Staphylococcus aureus compound PVI331 from a marine sponge associated Streptomyces. J. Appl. Biomed. 2016, 14, 273–283. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Skaf, J.; Holzgrabe, U.; Bharti, R.; Förstner, K.U.; Ziebuhr, W.; Humeida, U.H.; Abdelmohsen, U.R.; Oelschlaeger, T.A. A New Bioactive Compound From the Marine Sponge-Derived Streptomyces sp. SBT348 Inhibits Staphylococcal Growth and Biofilm Formation. Front. Microbiol. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, F.-J.R.S.; Iniyan, A.M.; Vincent, S.G.P. HR-LC-MS based analysis of two antibacterial metabolites from a marine sponge symbiont Streptomyces pharmamarensis ICN40. Microb. Pathog. 2017, 111, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Devescovi, G.; Kojic, M.; Covaceuszach, S.; Camara, M.; Williams, P.; Bertani, I.; Subramoni, S.; Venturi, V. Negative Regulation of Violacein Biosynthesis in Chromobacterium violaceum. Front. Microbiol. 2017, 8, 349. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Li, M.; Yi, G.; Liao, L.; Cheng, Q.; Zhu, J.; Zhang, B.; Wang, Y.; Chen, Y.; Zeng, M. Screening strategies for quorum sensing inhibitors in combating bacterial infections. J. Pharm. Anal. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kamarudheen, N.; Naushad, T.; Rao, K.V.B. Biosynthesis, Characterization and Antagonistic Applications of Extracellular Melanin Pigment from Marine Nocardiopsis Sps. Indian J. Pharm. Educ. Res. 2019, 53, s112–s120. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.-L.; Zhang, Y.; Xu, Y. Butenolide, a Marine-Derived Broad-Spectrum Antibiofilm Agent Against Both Gram-Positive and Gram-Negative Pathogenic Bacteria. Mar. Biotechnol. 2019, 21, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Le, K.Y.; Otto, M. Quorum-Sensing Regulation in Staphylococci-an Overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [Green Version]
- Kamarudheen, N.; Rao, K.B. Fatty acyl compounds from marine Streptomyces griseoincarnatus strain HK12 against two major bio-film forming nosocomial pathogens; an in vitro and in silico approach. Microb. Pathog. 2018, 127, 121–130. [Google Scholar] [CrossRef]
- Vignesh, A.; Ayswarya, S.; Gopikrishnan, V.; Radhakrishnan, M. Bioactive Potential of Actinobacteria Isolated from the Gut of Marine Fishes. Indian J. Geo-Marine Sci. 2019, 48, 1280–1285. [Google Scholar]
- Abd-Elnaby, H.; Abo-Elala, G.; Abdel-Raouf, U.; Abd-Elwahab, A.; Hamed, M. Antibacterial and anticancer activity of marine Streptomyces parvus: Optimization and application. Biotechnol. Biotechnol. Equip. 2015, 30, 180–191. [Google Scholar] [CrossRef]
- Mohamedin, A.H.; El-Naggar, N.E.-A.; Sherief, A.E.-D.A.; Hussien, S.M. Optimization of Bioactive Metabolites production by a Newly Isolated Marine Streptomyces sp. Using Statistical Approach. Biotechnology 2015, 14, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Katif, C.; Chilczuk, T.; Sabour, B.; Belattmania, Z.; Hilmi, A.; Niedermeyer, T.H.J.; Barakate, M. Isolation and Structure Elucidation of Desferrioxamine B and the New Desferrioxamine B2 Antibiotics from a Brown Marine Macroalga Carpodesmia Tamariscifolia Associated Streptomyces Isolate. Biointerface Res. Appl. Chem. 2022, 12, 5647–5662. [Google Scholar]
- Contreras-Castro, L.; Martínez-García, S.; Cancino-Diaz, J.C.; Maldonado, L.A.; Hernández-Guerrero, C.J.; Martínez-Díaz, S.F.; González-Acosta, B.; Quintana, E.T. Marine Sediment Recovered Salinispora sp. Inhibits the Growth of Emerging Bacterial Pathogens and other Multi-Drug-Resistant Bacteria. Pol. J. Microbiol. 2020, 69, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Ji, Y.-Y.; Zhang, H.-J.; Shen, L. Gephyyamycin and cysrabelomycin, two new angucyclinone derivatives from the Streptomyces sp. HN-A124. Nat. Prod. Res. 2019, 35, 2117–2122. [Google Scholar] [CrossRef] [PubMed]
- Iniyan, A.M.; Joseph, F.R.S.; Kannan, R.R.; Vincent, S.G.P. Anti-MRSA Potential of Phenolic Compound Isolated from a Marine Derived Actinomycete Micromonospora Sp. ICN36. Indian J. Geo-Marine Sci. 2016, 45, 1279–1287. [Google Scholar]
- Manikandan, M.; Gowdaman, V.; Duraimurugan, K.; Prabagaran, S.R. Taxonomic characterization and antimicrobial compound production from Streptomyces chumphonensis BDK01 isolated from marine sediment. 3 Biotech 2019, 9, 167. [Google Scholar] [CrossRef]
- Suthindhiran, K.; Kannabiran, K. Diversity and exploration of bioactive marine actinomycetes in the Bay of Bengal of the Puducherry coast of India. Indian J. Microbiol. 2010, 50, 76–82. [Google Scholar] [CrossRef]
- Ibrahimi, M.; Korichi, W.; Hafidi, M.; Lemee, L.; Ouhdouch, Y.; Loqman, S. Marine Actinobacteria: Screening for Predation Leads to the Discovery of Potential New Drugs against Multidrug-Resistant Bacteria. Antibiotics 2020, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Nazir, M.; Tousif, M.I.; Saleem, M.; Mustafa, R.; Khatoon, T. A New Polyketide Antibiotic from the Marine Bacterium Streptomyces sp. PGC 32. Chem. Nat. Compd. 2019, 55, 1–4. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, L.; Zhang, X.; Liang, Y.; Anjum, K.; Chen, L.; Lian, X.-Y. Bioactive Bafilomycins and a New N-Arylpyrazinone Derivative from Marine-derived Streptomyces sp. HZP-2216E. Planta Med. 2017, 83, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Song, Y.; Li, X.; Wang, X.; Ling, C.; Qin, X.; Zhou, Z.; Li, Q.; Wei, X.; Ju, J. Abyssomicin Monomers and Dimers from the Marine-Derived Streptomyces koyangensis SCSIO 5802. J. Nat. Prod. 2018, 81, 1892–1898. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Dong, Y.; Ju, J.; Li, Q. The roles of genes associated with regulation, transportation, and macrocyclization in desotamide biosynthesis in Streptomyces scopuliridis SCSIO ZJ46. Appl. Microbiol. Biotechnol. 2020, 104, 2603–2610. [Google Scholar] [CrossRef] [PubMed]
- Karuppiah, V.; Li, Y.; Sun, W.; Feng, G.; Li, Z. Functional gene-based discovery of phenazines from the actinobacteria associated with marine sponges in the South China Sea. Appl. Microbiol. Biotechnol. 2015, 99, 5939–5950. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, J.; Hu, Y.; Li, L.; Long, L.; Wang, F.; Zhang, S. Characterization of a thermophilic hemoglobin-degrading protease from Streptomyces rutgersensis SCSIO 11720 and its application in antibacterial peptides production. Biotechnol. Bioprocess Eng. 2015, 20, 79–90. [Google Scholar] [CrossRef]
- Abdulkhair, W.M.; Alghuthaymi, M.A. Double Inhibitory Effect of Extracellular Protein of Marine Streptomyces Tendae against Different Strains of MRSA. Der Pharm. Lett. 2016, 8, 11–20. [Google Scholar]
- Jiang, Y.-J.; Zhang, D.-S.; Zhang, H.-J.; Li, J.-Q.; Ding, W.-J.; Xu, C.-D.; Ma, Z.-J. Medermycin-Type Naphthoquinones from the Marine-Derived Streptomyces sp. XMA39. J. Nat. Prod. 2018, 81, 2120–2124. [Google Scholar] [CrossRef]
- Kurata, A.; Sugiura, M.; Kokoda, K.; Tsujimoto, H.; Numata, T.; Kato, C.; Nakasone, K.; Kishimoto, N. Taxonomy of actinomycetes in the deep-sea Calyptogena communities and characterization of the antibacterial compound produced by Actinomadura sp. DS-MS-114. Biotechnol. Biotechnol. Equip. 2017, 31, 1000–1006. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tian, X.; Niu, S.; Zhang, W.; Chen, Y.; Zhang, H.; Yang, X.; Zhang, W.; Li, W.; Zhang, S.; et al. Pseudonocardians A–C, New Diazaanthraquinone Derivatives from a Deap-Sea Actinomycete Pseudonocardia sp. SCSIO 01299. Mar. Drugs 2011, 9, 1428–1439. [Google Scholar] [CrossRef] [Green Version]
- Arslan, I. Trends in Antimicrobial Resistance in Healthcare-Associated Infections: A Global Concern. Ref. Modul. Biomed. Sci. 2022, 4, 652–661. [Google Scholar] [CrossRef]
- Kirst, H.A. Macrolide Antibiotics. In Antimicrobials; Springer: Berlin/Heidelberg, Germany, 2013; pp. 211–230. ISBN 978-3-642-39968-8. [Google Scholar] [CrossRef]
- Xu, H.; Li, H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am. J. Clin. Dermatol. 2019, 20, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Stahl, J.-P. Lincosamidas. EMC Tratado Med. 2017, 21, 1–4. [Google Scholar] [CrossRef]
- Ma, M.; Rateb, M.E.; Teng, Q.; Yang, D.; Rudolf, J.D.; Zhu, X.; Huang, Y.; Zhao, L.-X.; Jiang, Y.; Li, X.; et al. Angucyclines and Angucyclinones from Streptomyces sp. CB01913 Featuring C-Ring Cleavage and Expansion. J. Nat. Prod. 2015, 78, 2471–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2017, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Han, L.; Li, C.; Cao, Q.; Zhu, D.; Barrett, N.H.; Harmody, D.; Chen, J.; Zhu, H.; McCarthy, P.J.; et al. Bioprospecting Deep-Sea Actinobacteria for Novel Anti-infective Natural Products. Front. Microbiol. 2018, 9, 787. [Google Scholar] [CrossRef] [Green Version]
- Cumsille, A.; Undabarrena, A.; González, V.; Claverías, F.; Rojas, C.; Cámara, B. Biodiversity of Actinobacteria from the South Pacific and the Assessment of Streptomyces Chemical Diversity with Metabolic Profiling. Mar. Drugs 2017, 15, 286. [Google Scholar] [CrossRef] [Green Version]
- Su, P.; Wang, D.-X.; Ding, S.-X.; Zhao, J. Isolation and diversity of natural product biosynthetic genes of cultivable bacteria associated with marine sponge Mycale sp. from the coast of Fujian, China. Can. J. Microbiol. 2014, 60, 217–225. [Google Scholar] [CrossRef]
- Meena, B.; Anburajan, L.; Vinithkumar, N.V.; Kirubagaran, R.; Dharani, G. Biodiversity and antibacterial potential of cultivable halophilic actinobacteria from the deep sea sediments of active volcanic Barren Island. Microb. Pathog. 2019, 132, 129–136. [Google Scholar] [CrossRef]
- Bauermeister, A.; Pereira, F.; Grilo, I.R.; Godinho, C.C.; Paulino, M.; Almeida, V.; Gobbo-Neto, L.; Prieto-Davó, A.; Sobral, R.G.; Lopes, N.P.; et al. Intra-clade metabolomic profiling of MAR4 Streptomyces from the Macaronesia Atlantic region reveals a source of anti-biofilm metabolites. Environ. Microbiol. 2019, 21, 1099–1112. [Google Scholar] [CrossRef]
- Anggelina, A.C.; Pringgenies, D.; Setyati, W.A. Presence of Biosynthetic Gene Clusters (NRPS/PKS) in Actinomycetes of Mangrove Sediment in Semarang and Karimunjawa, Indonesia. Environ. Nat. Res. J. 2021, 19, 391–401. [Google Scholar] [CrossRef]
- Íñiguez-Martínez, A.M.; Cardoso-Martínez, F.; De La Rosa, J.; Cueto, M.; Díaz-Marrero, A.; Darias, J.; Becerril-Espinosa, A.; Rosas, L.J.P.; Soria-Mercado, E.I. Compounds isolated from Salinispora arenicola of the Gulf of California, México. Rev. Biol. Mar. Oceanogr. 2016, 51, 161–170. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, M.Y.; Barakat, K.M.; Aly, N.S. Physiological Response, Antibacterial Activity and Cinnamaldehyde Production by a Marine Streptomyces Chartreusis. J. Pure Appl. Microbiol. 2016, 10, 1797–1808. [Google Scholar]
- Rajasabapathy, R.; Ghadi, S.C.; Manikandan, B.; Mohandass, C.; Surendran, A.; Dastager, S.G.; Meena, R.M.; James, R.A. Antimicrobial profiling of coral reef and sponge associated bacteria from southeast coast of India. Microb. Pathog. 2020, 141, 103972. [Google Scholar] [CrossRef] [PubMed]
- Gandhimathi, R.; Arunkumar, M.; Selvin, J.; Thangavelu, T.; Sivaramakrishnan, S.; Kiran, G.; Shanmughapriya, S.; Natarajaseenivasan, K. Antimicrobial potential of sponge associated marine actinomycetes. J. Mycol. Med. 2008, 18, 16–22. [Google Scholar] [CrossRef]
- Wang, C.; Du, W.; Lu, H.; Lan, J.; Liang, K.; Cao, S. A Review: Halogenated Compounds from Marine Actinomycetes. Molecules 2021, 26, 2754. [Google Scholar] [CrossRef] [PubMed]
- Machmudah, S.; Kanda, H.; Goto, M. Hydrolysis of Biopolymers in Near-Critical and Subcritical Water; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128093801. [Google Scholar]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New antibacterial xanthone from the marine sponge-derived Micrococcus sp. EG45. Bioorganic Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef]
- Kurnianto, M.A.; Kusumaningrum, H.D.; Lioe, H.N.; Chasanah, E. Partial Purification and Characterization of Bacteriocin-Like Inhibitory Substances Produced by Streptomyces Sp. Isolated from the Gut of Chanos chanos. Biomed Res. Int. 2021, 2021, 7190152. [Google Scholar] [CrossRef]
- Nivina, A.; Yuet, K.P.; Hsu, J.; Khosla, C. Evolution and Diversity of Assembly-Line Polyketide Synthases. Chem. Rev. 2019, 119, 12524–12547. [Google Scholar] [CrossRef]
- Wink, J.; Mohammadipanah, F.; Hamedi, J. Biology and Biotechnology of Actinobacteria; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; ISBN 9783319603391. [Google Scholar]
- Luti, K.J.K. Mixture Design of Experiments for the Optimization of Carbon Source for Promoting Undecylprodigiosin and Actinorhodin Production. J. Pure Appl. Microbiol. 2018, 12, 1783–1793. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Ballav, S.; Kerkar, S.; Thomas, S.; Augustine, N. Halophilic and halotolerant actinomycetes from a marine saltern of Goa, India producing anti-bacterial metabolites. J. Biosci. Bioeng. 2015, 119, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, S.; Jayashree, M.; Shivani, R.; Anwesha, S.; Rao, K.B. Characterisation and identification of antibacterial compound from marine actinobacteria: In vitro and in silico analysis. J. Infect. Public Health 2018, 12, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; MacIntyre, L.; Abdelmohsen, U.R.; Horn, H.; Polymenakou, P.N.; Edrada-Ebel, R.; Hentschel, U. Biodiversity, Anti-Trypanosomal Activity Screening, and Metabolomic Profiling of Actinomycetes Isolated from Mediterranean Sponges. PLoS ONE 2015, 10, e0138528. [Google Scholar] [CrossRef] [PubMed]
- Rajan, B.M.; Kannabiran, K. Extraction and Identification of Antibacterial Secondary Metabolites from Marine Streptomyces sp. VITBRK2. Int. J. Mol. Cell. Med. 2014, 3, 130–137. [Google Scholar]
- Thosar, A.; Satpathy, P.; Devi, C.S. Marine Streptomyces sp. VITASP as a Source of New Bioactive Secondary Metabolites. Curr. Bioact. Compd. 2020, 16, 611–617. [Google Scholar] [CrossRef]
- Mercy, R.B.; Kannabiran, K. Identification of Antibacterial Secondary Metabolite from Marine Streptomyces Sp. VITBRK4 and Its Activity against Drug Resistant Gram Positive Bacteria. Int. J. Drug Dev. Res. 2013, 5, 224. [Google Scholar]
- Mohseni, M.; Norouzi, H.; Hamedi, J.; Roohi, A. Screening of Antibacterial Producing Actinomycetes from Sediments of the Caspian Sea. Int. J. Mol. Cell. Med. 2013, 2, 64–71. [Google Scholar]
- Yuan, X.W.; Yang, R.L.; Cao, X.; Gao, J.J. Taxonomic identification of a novel strain of Streptomyces cavourensis subsp. washingtonensis, ACMA006, exhibiting antitumor and antibacteria activity. Drug Discov. Ther. 2010, 4, 405–411. [Google Scholar]
- Wahaab, F.; Subramaniam, K. Bioprospecting marine actinomycetes for multidrug-resistant pathogen control from Rameswaram coastal area, Tamil Nadu, India. Arch. Microbiol. 2017, 200, 57–71. [Google Scholar] [CrossRef]
- Rajan, B.M.; Kannabiran, K. Antagonistic Activity of Marine Streptomyces Sp. VITBRK1 on Drug Resistant Gram Positive Cocci. Der Pharm. Lett. 2013, 5, 185–191. [Google Scholar]
- Kokare, C.R.; Mahadik, K.R.; Kadam, S.S.; Chopade, B.A. Isolation, Characterization and Antimicrobial Activity of Marine Halophilic Actinopolyspora Species AH1 from the West Coast of India. Curr. Sci. 2004, 86, 593–597. [Google Scholar]
- Tenebro, C.P.; Trono, D.J.V.L.; Vicera, C.V.B.; Sabido, E.M.; Ysulat, J.J.A.; Macaspac, A.J.M.; Tampus, K.A.; Fabrigar, T.A.P.; Saludes, J.P.; Dalisay, D.S. Multiple Strain Analysis of Streptomyces Species from Philippine Marine Sediments Reveals Intraspecies Heterogeneity in Antibiotic Activities. Sci. Rep. 2021, 11, 17544. [Google Scholar] [CrossRef] [PubMed]
- Mane, M.; Mahadik, K.; Kokare, C. Purification, Characterization and Applications of Thermostable Alkaline Protease from Marine Streptomyces Sp. D1. Int. J. Pharma Bio. Sci. 2013, 4, 572–582. [Google Scholar]
- Ouchene, R.; Intertaglia, L.; Zaatout, N.; Kecha, M.; Suzuki, M.T. Selective isolation, antimicrobial screening and phylogenetic diversity of marine actinomycetes derived from the Coast of Bejaia City (Algeria), a polluted and microbiologically unexplored environment. J. Appl. Microbiol. 2021, 132, 2870–2882. [Google Scholar] [CrossRef]
- Mani, A.; Ravi, L.; Krishnan, K. Antibacterial and antifungal potential of marine Streptomyces sp. VITAK1 derived novel compound Pyrrolidinyl-Hexadeca-Heptaenone by in Silico docking analysis. Res. J. Pharm. Technol. 2018, 11, 1901. [Google Scholar] [CrossRef]
- Undabarrena, A.; Beltrametti, F.; Claverías, F.P.; González, M.; Moore, E.R.B.; Seeger, M.; Cámara, B. Exploring the Diversity and Antimicrobial Potential of Marine Actinobacteria from the Comau Fjord in Northern Patagonia, Chile. Front. Microbiol. 2016, 7, 1135. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, X.; Chen, L.; Yan, S.; Ye, X.; Anjum, K.; Huang, H.; Lian, X.; Zhang, Z. Bioactive Polycyclic Quinones from Marine Streptomyces sp. 182SMLY. Mar. Drugs 2016, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Attimarad, S.L.; Gaviraj, E.N.; Nagesh, C.; Kugaji, M.S.; Sutar, R.S. Screening, Isolation and Purification of Antibiotic(s) from Marine Actinomycetes. Int. J. Res. Ayurveda Pharm. 2012, 3, 447–453. [Google Scholar]
- León, J.; Liza, L.; Soto, I.; Cuadra, D.; Patiño, L.; Zerpa, R. Bioactives Actinomycetes of Marine Sediment from the Central Coast of Peru [Actinomycetes Bioactivos de Sedimento Marino de La Costa Central Del Perú]. Rev. Peru. Biol. 2007, 14, 259–270. [Google Scholar]
- Jagan Mohan, Y.S.Y.V.; Sirisha, B.; Haritha, R.; Ramana, T. Selective Screening, Isolation and Characterization of Antimicrobial Agents from Marine Actinomycetes. Int. J. Pharm. Pharm. Sci. 2013, 5, 443–449. [Google Scholar]
- Song, Y.; Li, Q.; Qin, F.; Sun, C.; Liang, H.; Wei, X.; Wong, N.-K.; Ye, L.; Zhang, Y.; Ju, J.; et al. Neoabyssomicins A–C, Polycyclic Macrolactones from the Deep-Sea Derived Streptomyces Koyangensis SCSIO 5802. Tetrahedron 2017, 73, 5366–5372. [Google Scholar] [CrossRef]
- Eliwa, E.M.; Abdel-Razek, A.S.; Frese, M.; Halawa, A.H.; El-Agrody, A.M.; Bedair, A.H.; Sewald, N.; Shaaban, M. New naturally occurring phenolic derivatives from marine Nocardiopsis sp. AS23C: Structural elucidation and in silico computational studies. Vietnam J. Chem. 2019, 57, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Tangjitjaroenkun, J.; Pluempanupat, W.; Tangchitcharoenkhul, R.; Yahayo, W.; Supabphol, R. Antibacterial, antioxidant, cytotoxic effects and GC-MS analysis of mangrove-derived Streptomyces achromogenes TCH4 extract. Arch. Biol. Sci. 2021, 73, 223–235. [Google Scholar] [CrossRef]
- Asnani, A.; Purwanti, A.; Bakrudin, W.A.; Anjarwati, D.U. The Production of Streptomyces W-5B Extract for Antibiofilm against Methicillin-resistant Staphylococcus aureus. J. Pure Appl. Microbiol. 2022, 16, 337–346. [Google Scholar] [CrossRef]
- Devi, N.A. Isolation and Identification of Marine Actinomycetes and their Potential in Antimicrobial Activity. Pak. J. Biol. Sci. 2006, 9, 470–472. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.T.; Tran, V.H.; Vu, V.N.; Mai, H.D.T.; Le, T.H.M.; Vu, T.Q.; Nguyen, H.H.; Chau, V.M.; Pham, V.C. Antimicrobial metabolites from a marine-derived Actinomycete Streptomyces sp. G278. Nat. Prod. Res. 2018, 33, 3223–3230. [Google Scholar] [CrossRef]
- Asolkar, R.N.; Kirkland, T.N.; Jensen, P.; Fenical, W. Arenimycin, an antibiotic effective against rifampin- and methicillin-resistant Staphylococcus aureus from the marine actinomycete Salinispora arenicola. J. Antibiot. 2009, 63, 37–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Powers, M.; Králová, S.; Nguyen, G.-S.; Fawwal, D.V.; Degnes, K.; Lewin, A.S.; Klinkenberg, G.; Wentzel, A.; Liles, M.R. Streptomyces Poriferorum Sp. Nov., a Novel Marine Sponge-Derived Actinobacteria Species Expressing Anti-MRSA Activity. Syst. Appl. Microbiol. 2021, 44, 126244. [Google Scholar] [CrossRef]
- Dharmaraj, S.; Sumantha, A. Bioactive Potential of Streptomyces Associated with Marine Sponges. World J. Microbiol. Biotechnol. 2009, 25, 1971–1979. [Google Scholar] [CrossRef]
- Cristianawati, O.; Sibero, M.T.; Ayuningrum, D.; Nuryadi, H.; Syafitri, E.; Radjasa, O.K.; Riniarsih, I. Screening of Antibacterial Activity of Seagrass-Associated Bacteria from the North Java Sea, Indonesia against Multidrug-Resistant Bacteria. AACL Bioflux 2019, 12, 1054–1064. [Google Scholar]
- Antoniraj, A.; Anandan, V.; Ganesan, T.; Gunasingh, A. Isolation of Marine Actinomycetes Associated with the Carangid Fish Alepes Melanoptera, (Swainson, 1839) and an Evaluation of Their Antimicrobial Activity. J. Microbiol. 2018, 20, 235–247. [Google Scholar]








| Genus | Pathogen Target | Compounds/Extracts | MIC (μg/mL) | Ref. |
|---|---|---|---|---|
| Streptomyces sp. | S. aureus FDA209P JC-1 | Chlorinated α-lapachone | 12.5 | [31] |
| Streptomyces sp. | MRSA 1 | Streptoindoles A | 25 | [32] |
| Streptoindoles B | 7 | [32] | ||
| Streptoindoles D | 25 | [32] | ||
| Streptomyces sp. | MRSA 1 | Streptoglutarimides A−J | 9–11 | [111] |
| Streptomyces sp. | S. aureus | Nitricquinomycin C | 17 | [112] |
| Streptomyces sp. | MRSA 1 | Napyradiomycin D1 | 12–24 | [113] |
| Streptomyces sp. | S. aureus ATCC 33591 | Polyketide antibiotic SBR-22 | 64 | [114] |
| Streptomyces sp. | S. aureus ATCC 29213 | Lobophorins F | 6.25 | [115] |
| Streptomyces sp. | S. aureus | Polyketide related antibiotic | 37.5 | [30] |
| Streptomyces sp. | MRSA 1 | Actinomycin D | 0.08 | [116] |
| Actinomycin V | 0.08 | [116] | ||
| Actinomycin X0β | 0.61 | [116] | ||
| Streptomyces sp. | MRSA 1 | Niphimycins C | 4–32 | [117] |
| MRSE 2 | Niphimycin Iα | 4–32 | [117] | |
| Streptomyces sp. | S. aureus ATCC 25923 | Trihydroxylflavanone 3 | 32 | [118] |
| Tetrahydroxylchalcone 4 | 1 | [118] | ||
| Streptomyces sp. | S. aureus | Anthracycline analogues | 20 | [119] |
| β-rhodomycin-II | 40 | [119] | ||
| Streptomyces sp. | S. aureus | DMBPO 5 | >1000 | [120] |
| Streptomyces sp. | S. aureus ATCC 25923 | Chromomycin A9 | 0.03 | [121] |
| Chromomycin Ap | 0.13 | [121] | ||
| Chromomycin A2 | 0.06 | [121] | ||
| Chromomycin A3 | 0.13 | [121] | ||
| Streptomyces sp. | MRSA 1 | Streptopyrazinones A–D | 58–65 | [122] |
| N-acetyl-L-isoleucine-L-leucinamide | 65 | [122] | ||
| Streptomyces sp. | MRSA 1 | 4-dehydro-4a-dechloronapyradiomycin A1 | 4–8 | [123] |
| Napyradiomycin A1 | 0.5–1 | [123] | ||
| Streptomyces sp. | S. aureus | 3-propanoic acid 6 | 32 | [124] |
| Propanoic acid methyl ester 7 | 64 | [124] | ||
| 3-(3-chloro-4-hydroxyphenyl) propanoic acid | 32 | [124] | ||
| Streptomyces sp. | S. aureus (ATCC 6538) | Natural cyclic peptide | 1.25 | [125] |
| MRSA 1 | 12.5 | [125] | ||
| S. aureus (ATCC 6538) | Cyclic peptides | 0.025–0.156 | [125] | |
| MRSA 1 | Cyclic peptides | 0.1–0.78 | [125] | |
| Streptomyces sp. | S. aureus | Extracts A758 | 6.25 | [126] |
| Extracts A759 | 500 | [126] | ||
| Extracts A760 | 100 | [126] | ||
| Extracts A765 | 3.125 | [126] | ||
| Streptomyces sp. | MRSA 1 | Novobiocin | 0.25 | [127] |
| Desmethylnovobiocin | 16 | [127] | ||
| 5-Hydroxynovobiocin | 8 | [127] | ||
| Kocuria marina | S. aureus | Kocumarin | 10 | [128] |
| MRSA 1 | Kocumarin | 10 | [128] | |
| Rhodococcus sp. | S. aureus | n-butanol | 9.3 | [34] |
| fraction | ||||
| EtOAc fraction | 12.6 | [34] | ||
| Marinispora sp. | MRSA 1 | Marinomycin A | 0.130 | [129] |
| Marinomycin B–C | 0.49 | [129] | ||
| Marinomycin D | 2.43 | [129] | ||
| Verrucosispora sp. | S. aureus | (2-(hydroxymethyl)-3-(2-(hydroxymethyl)-3-methylaziridin-1-yl) (2-hydroxyphenyl) methanone | 3.4 | [130] |
| Genus | Pathogen Target | Compounds/Extracts | MIC (μg/mL) | Ref. |
|---|---|---|---|---|
| Streptomyces sp. | MRSA 1 | Compound PVI331 | 1 | [132] |
| Streptomyces sp. | MRSA 1 | 8-O-metyltetrangomycin | 2 | [10] |
| Streptomyces sp. | MRSE 2 RP62A | Compound (SKC3) | 31.25 | [133] |
| Streptomyces sp. | MRSA 1 | PVI401 | 0.5 | [134] |
| PVI402 | 2 |
| Source | Genus | Disrupter QS System | Biosensor Strains | Ref. |
|---|---|---|---|---|
| Gut of marine fishes | Streptomyces sp. | AI-1: AHL | C. violaceum and Serratia marcescens. | [141] |
| NA 1 | Streptomyces sp. | AI-1: AHL, AI-2: LuxS | C. violaceum CV026 and Vibrio harveyi BB170 | [138] |
| Marine Sponge | Streptomyces sp. | AI-1: AHL: LasI | Pseudomona- Molecular docking. | [140] |
| Marine sediment | Nocardiopsis sp. | AI-1: AHL | C. violaceum 12472 | [29] |
| Seawater | Nocardiopsis sp. | AI-1: AHL | C. violaceum (MTCC 2656) | [137] |
| Compound | Frequency | Constituents | Ref. |
|---|---|---|---|
| Polyketide | 19 | Naphthoquinone-based meroterpenoids Naphthoquinone Derivatives | [37] [25] |
| Chlorinated Meroterpenoids (Merochlorins G–J) | [78] | ||
| Angucycline | [23,42,50,54,100,102] | ||
| Aromatic Polyketides | [61,68,151] | ||
| Polyketide 1 | [72] | ||
| Compound 1 2 | [43] | ||
| Macrolides 3 | [52,57,67,96,107,132,152] | ||
| Phenolic compound | 1 | Bromophenol derivative | [101] |
| Phthalate | 1 | Bis (2-ethylhexyl) | [101] |
| Acetamide | 2 | 4-methoxyacetanilide | [18] |
| Alkaloids | 3 | 2-ethylhexyl 1H-imidazole-4-carboxylate | [97] |
| butyl 1Himidazole-4-carboxylate | [97] | ||
| Chlorinated bis-indole alkaloids | [45] | ||
| Indolizinium alkaloid | [58] | ||
| Pyrrole | 3 | Chlorinated Bisindole Pyrrole | [106] |
| Pyrrole-derivative | [41,60] | ||
| Chromopeptides | 6 | Actinomycins (X0β, X2, D, D1–D4, A) | [56] |
| Neo-actinomycin A, B, actinomycins D and C4, X2,) | [64,77,80,153] | ||
| Cyclo peptides | 3 | Desotamides A–D | [154] |
| cyclo(l-Val-l-Pro), | [79] | ||
| cyclo-(l-Pro-4-OH-l-Leu) | [55] | ||
| Antracycline | 1 | Bisanhydroaklavi-none 1-Hydroxybisanhydroaklavinone | [19] |
| Marinopyrroles | 1 | (−)-marinopyrroles A | [70] |
| (−)-marinopyrroles B | |||
| Phenazines | 5 | phenazine-1,6-dicarboxylate, phencomycin, tubermycin | [27] [63] |
| Streptophenazines G | |||
| 1,6-Dihydroxy phenazine, dimethoxy phenazine Actinomycins D1 and D2 | [155] [41] | ||
| Spirotetronate antibiotics | 2 | Lobophorins L and M | [62] |
| Lobophorins E | |||
| Proteins | 2 | Enzyme PA720 (Thermophilic Hemoglobin-degrading Protease) | [156] |
| β-lactamase inhibitory protein | [157] | ||
| Pyranonaphthoquinones | 3 | Medermycin-type naphthoquinones | [158] |
| Medermycin derivative | [51] | ||
| Lactoquinomycin A (LQM-A) | [53] | ||
| Quinomycin family antibiotics | 1 | Quinomycin G | [55] |
| Quinona | 1 | 1- hydroxy-1-norresistomycin | [38] |
| Siderophore native | 3 | S1, S2, S3 4 | [57,144,159] |
| Thiazolyl Peptide Antibiotic Family | 1 | Kocurin | [108] |
| Pigment | 1 | Melanin pigment | [137] |
| Aminofuran natural products | 1 | Proximicin F and G | [94] |
| Type I lasso peptide natural products | 1 | Aborycin | [48] |
| Natural product class diazaanthraquinone | 1 | Diazaanthraquinone | [160] |
| Benzoic acid | 1 | 2,4-dichloro-5-sulfamoyl benzoic acid | [44] |
| 4-oxazolidinone antibiotics | 1 | Lipoxazolidinone A, B and C. | [105] |
| Cyslabdan-like compound | 1 | Cyslabdan-like compound | [93] |
| Benzene Derivative | 1 | 1,3-Benzodioxole | [81] |
| Flavonoids | 3 | Citreamicin θ A | [84] |
| Citreamicin θ B | |||
| Citreaglycon A | |||
| Dehydrocitreaglycon A |
| Compound | Frequency | Constituents | Ref. |
|---|---|---|---|
| Fatty acyl compounds | 1 | 13Z-Octadecenal. | [140] |
| Phenoxazines | 1 | Questiomycin A | [29] |
| 2-hydroxyacetate-3-hydroxyacetamido-phenoxazine (HHP) | [29] | ||
| Lactones | 1 | Butenolide | [138] |
| Pigment | 1 | Melanin | [137] |
| Strain IM20 1 | 1 | NA 2 | [141] |
| Genus | BGS | Genes | Metabolites Production | Ref. |
|---|---|---|---|---|
| Streptomyces sp. | PKS gene cluster | PKS-I and PKS-II Genes | Polyketide | [20,30] |
| Angucycline | [23] | |||
| PKS-II Genes | Angucyclinone derivatives | [146] | ||
| PKS-KS | NA 1 | [169] | ||
| PKS | Niphimycins | [117] | ||
| PKS/NRPS | PKS II | Analogue of paulomenol | [103] | |
| NA 1 | Antimycin A analogues | [77] | ||
| NA 1 | NA 1 | [168,170] | ||
| NRPS, PKS Type I, II, and III | Naphthoquinone antibiotics | [25] | ||
| NRPS gene cluster | NRPS-A | NA 1 | [169] | |
| NA 1 | NA 1 | [49] | ||
| Aborycin biosynthetic gene cluster (abo) | NA 1 | Aborycin | [48] | |
| Lassopeptide cluster | NA 1 | Lasso peptide family | [48] | |
| Phenazine cluster | phzE and phzF | Streptophenazines (Phenazines) | [83] | |
| dsa cluster | DsaA y DsaN, dsaB y dsaJ | Desotamides | [154] | |
| PKS/terpenoid biosynthetic pathways | NA 1 | Napyradiomycin derivatives (Terpenoids) | [71,113,171] | |
| Micromonospora sp. | nes gene cluster | NA 1 | nenestatin A (Benzofluorene angucyclines) | [100] |
| Co-culture of Actinokineospora sp. and Micromonospora sp. | NA | NA 1 | Phenazine | [27] |
| Nocardiopsis sp. | PKS/NRPS | NA 1 | Polyketide | [170] |
| PKS gene cluster | PKS-II | α-pyrone compound | [103] | |
| ACP synthase α-subunit (KSα), β-subunit (KSβ) and acyl carrier protein (ACP) | Angucyclines | [102] | ||
| PKS-II | Angucycline | [102] | ||
| phzE | Phenazines | [155] | ||
| Rhodococcus sp. | NRPS/NRPS | NA 1 | NA 1 | [22] |
| PKS/NRPS | NA 1 | Polyketide | [170] | |
| Salinispora sp. | PKS gene cluster | PKS I, II | Rifamycin B | [103] |
| PKS/NRPS | NA 1 | Polyketide | [170] | |
| Verrucosispora sp. | PKS gene cluster | PKSI (pks1 and pks2), two PKSII (pks3 and pks4), PKSIII (pks5); | New salicylic derivative, brevianamide F, abyssomicin B | [95] |
| NRPS gene cluster | NA 1 | |||
| Terpene clusters | terp1, terp2, terp3 and terp4 | |||
| NRPS-PKSI hybrid clusters | np1 and np2 | |||
| Lanthipeptide clusters | lant1 and lant2 | |||
| Siderophore cluster | sid | |||
| Brachybacterium paraconglomeratum | NRPS/PKS | NRPS genes, PKS type I genes, and PKS type II gene | NA 1 | [172] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De La Hoz-Romo, M.C.; Díaz, L.; Villamil, L. Marine Actinobacteria a New Source of Antibacterial Metabolites to Treat Acne Vulgaris Disease—A Systematic Literature Review. Antibiotics 2022, 11, 965. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11070965
De La Hoz-Romo MC, Díaz L, Villamil L. Marine Actinobacteria a New Source of Antibacterial Metabolites to Treat Acne Vulgaris Disease—A Systematic Literature Review. Antibiotics. 2022; 11(7):965. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11070965
Chicago/Turabian StyleDe La Hoz-Romo, Maria Clara, Luis Díaz, and Luisa Villamil. 2022. "Marine Actinobacteria a New Source of Antibacterial Metabolites to Treat Acne Vulgaris Disease—A Systematic Literature Review" Antibiotics 11, no. 7: 965. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11070965