
Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin



Abstract
:1. Introduction
2. Results
2.1. A Systematic Review on Plant-Based Natural Compounds (PBCs) as Antibacterial Agents
2.2. Antibacterial Potency of PBCs in Planktonic Form
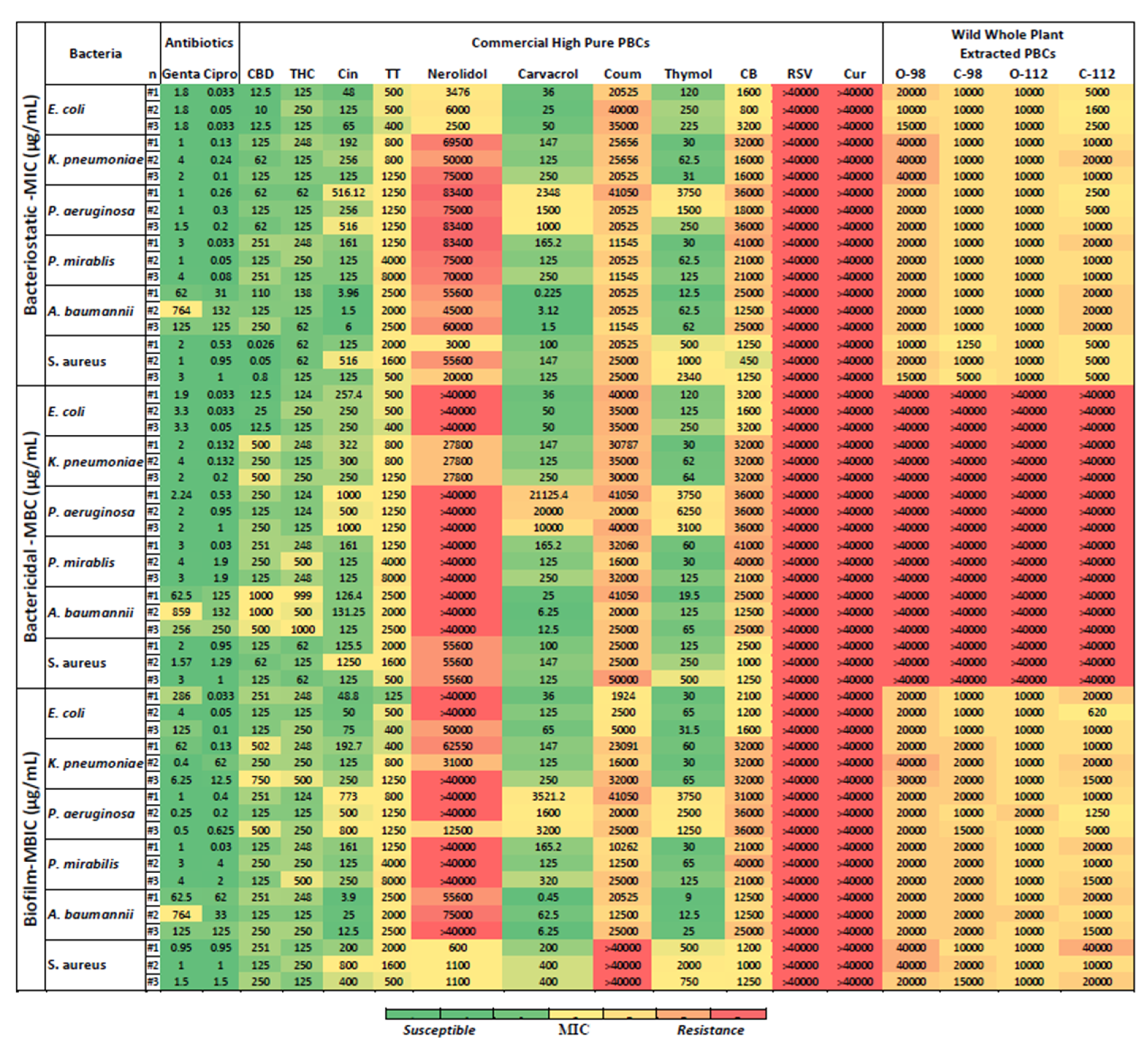
2.3. Bacteriostatic Potency of PBCs
2.4. Bactericidal Potency of PBCs
2.5. Antibiofilm Potency of PBCs
2.6. Targeted Treatment of P. aeruginosa, S. aureus, E. coli, K. pneumoniae, P. mirabilis, and A. baumannii with PBCs
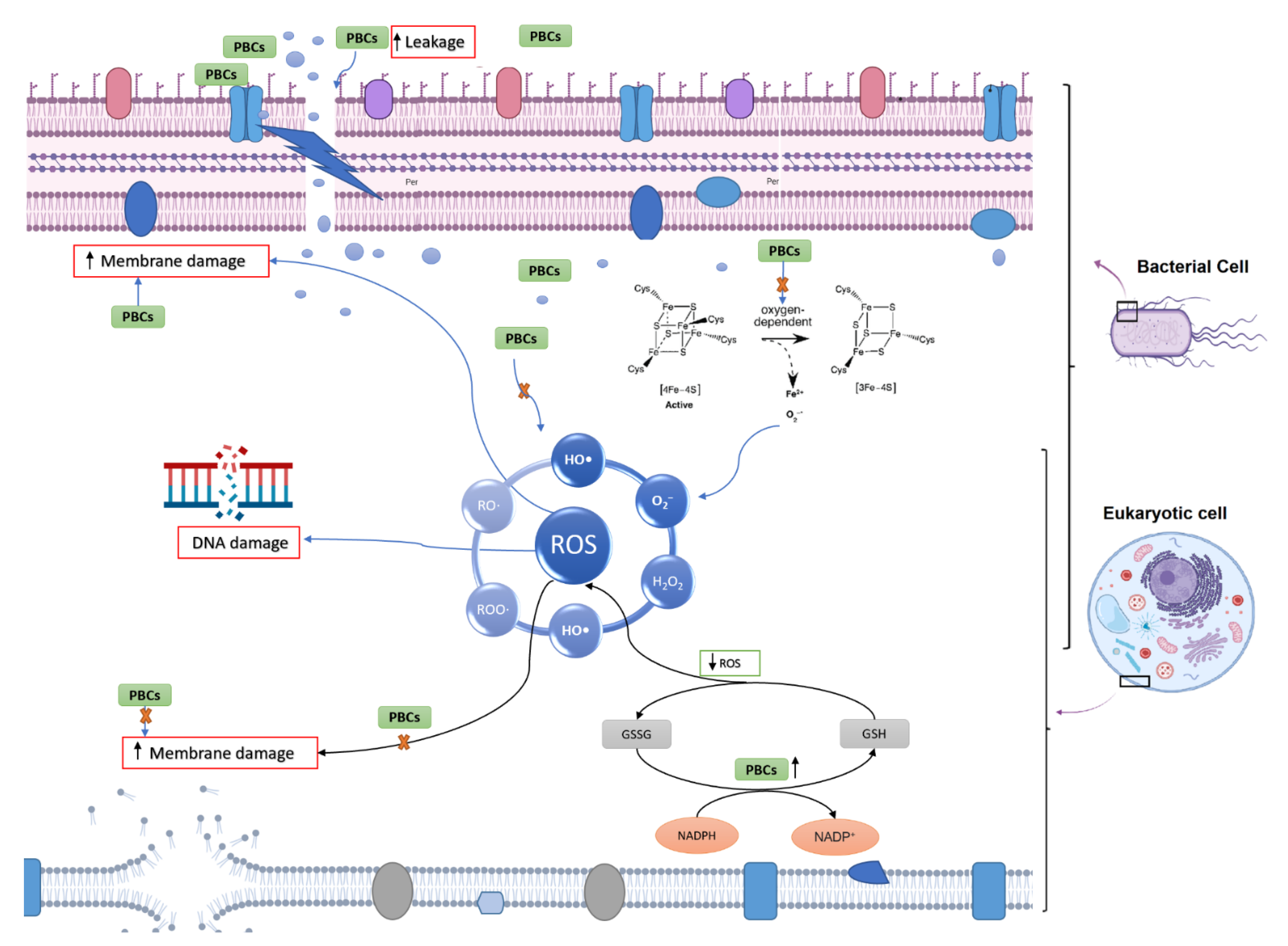
2.7. Potential Antibacterial Mechanism of Selected PBCs
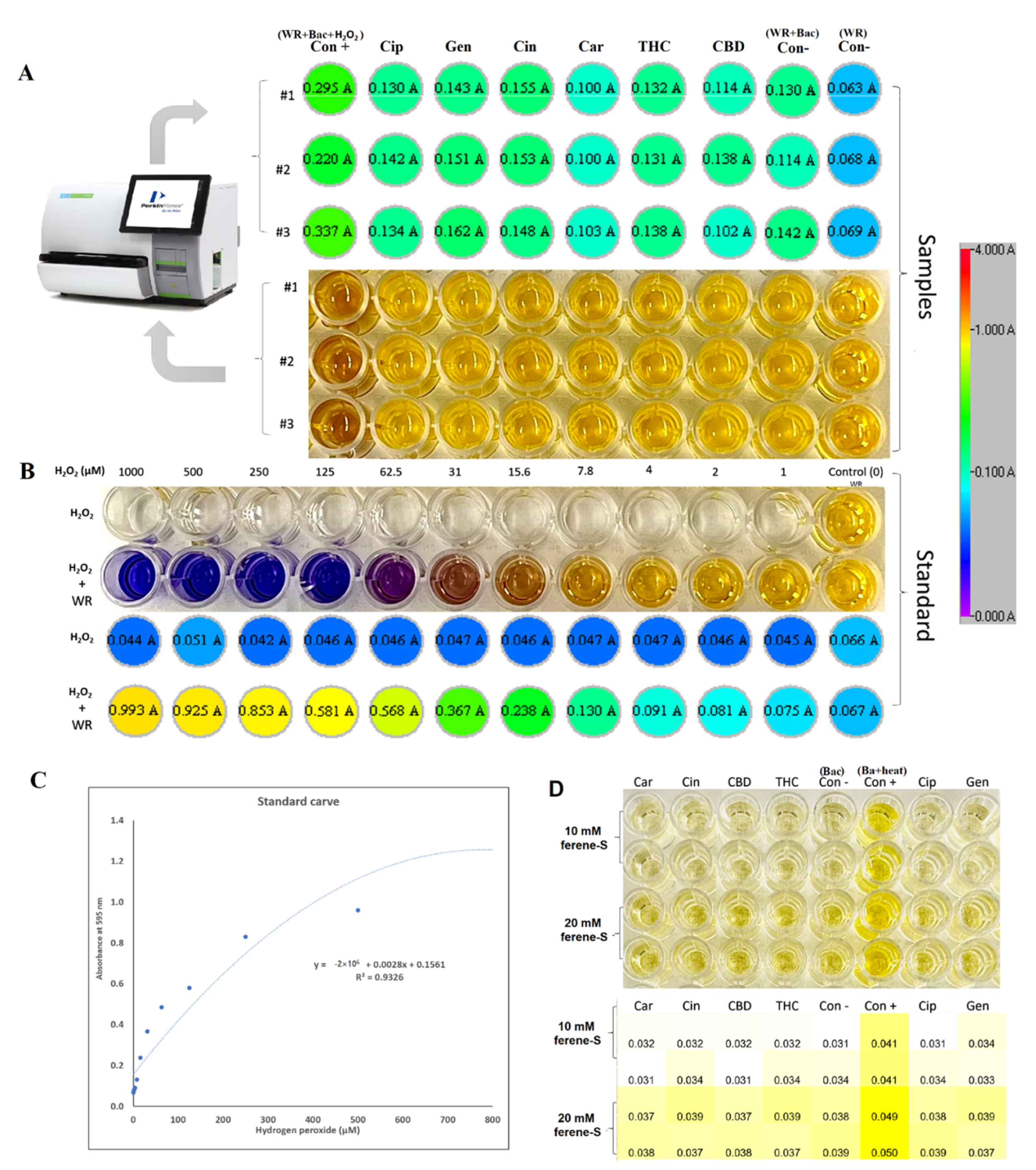
2.7.1. Oxidative Stress and Fe-S Complex
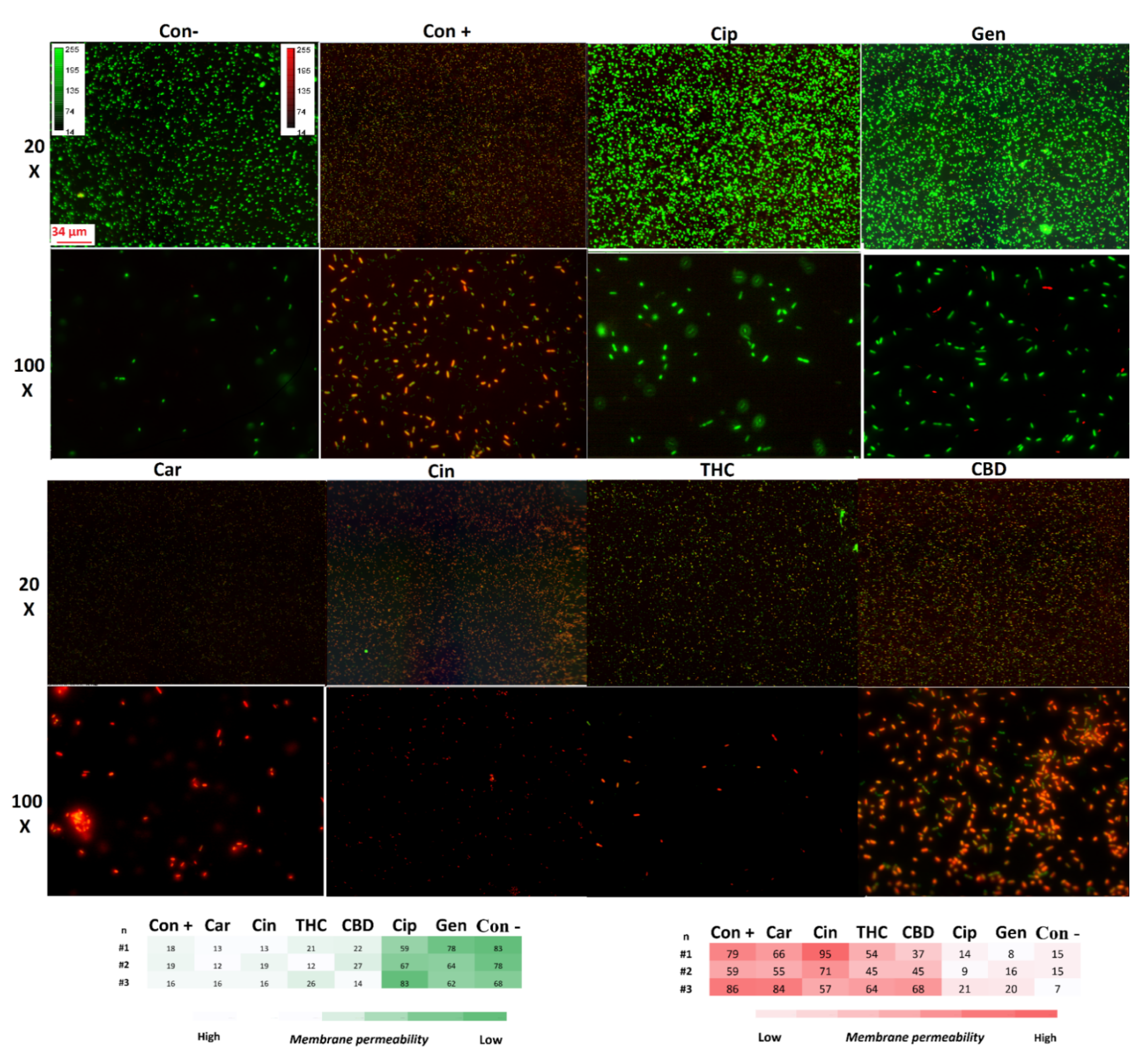
2.7.2. Membrane Permeability
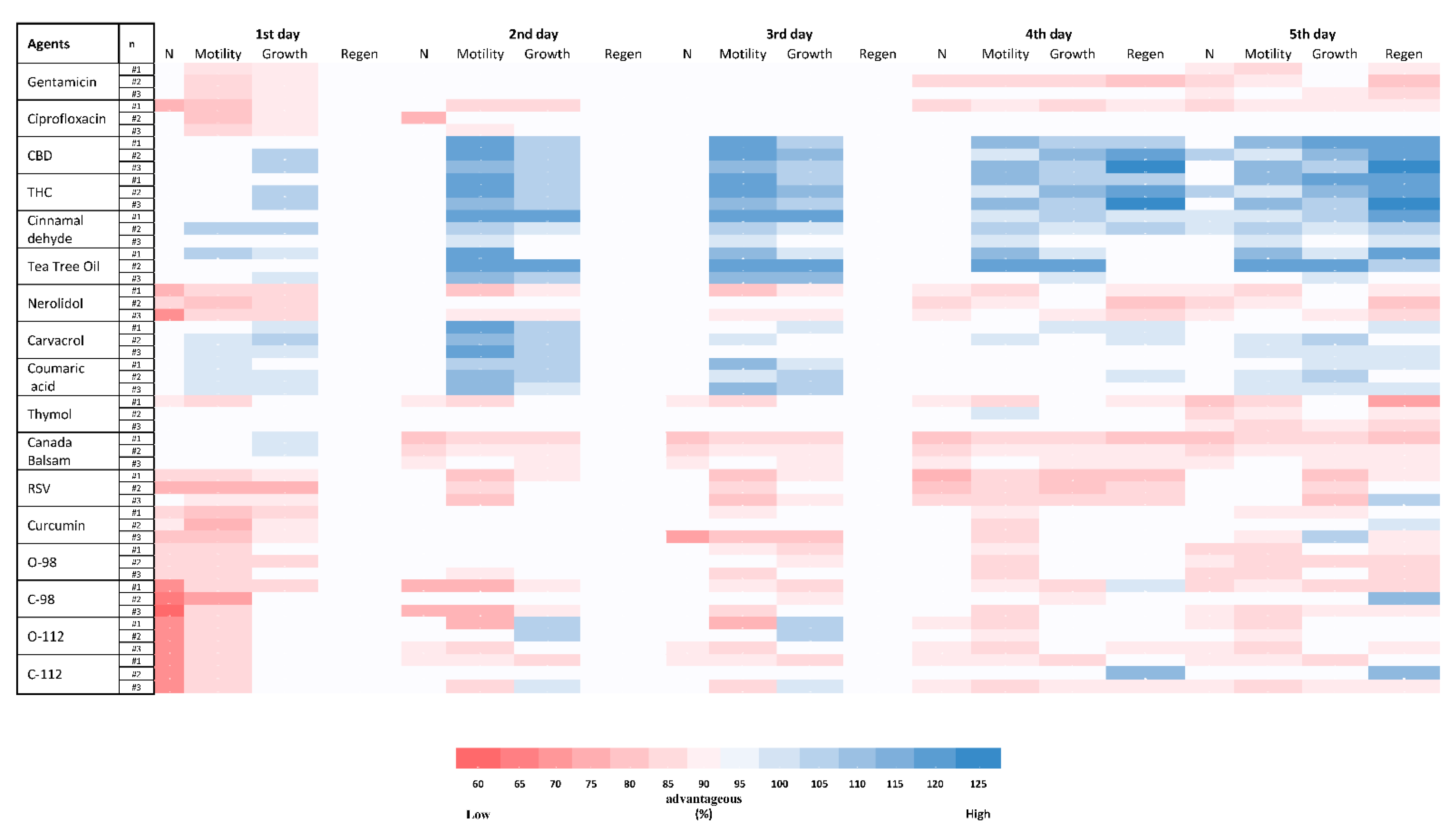
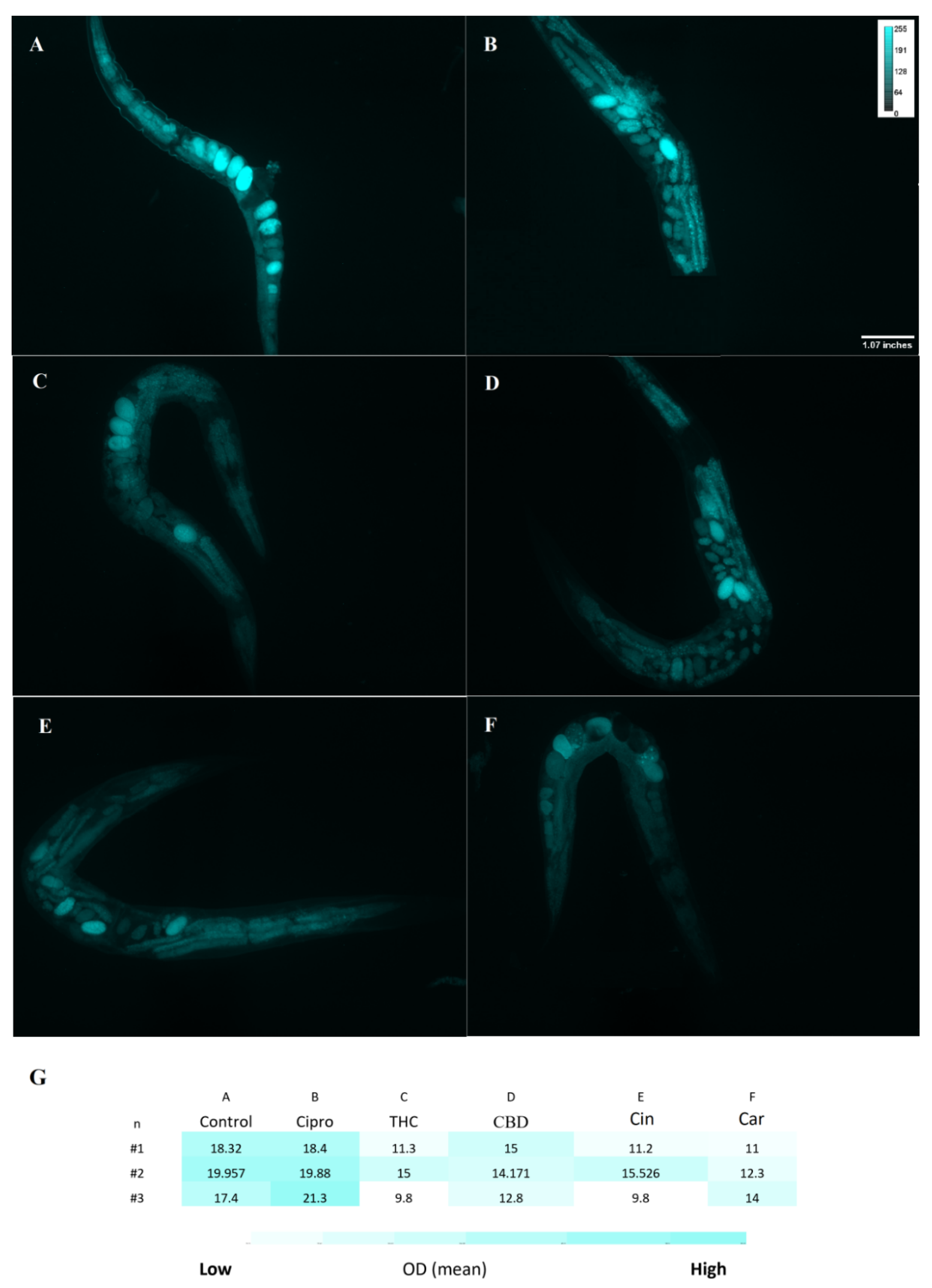
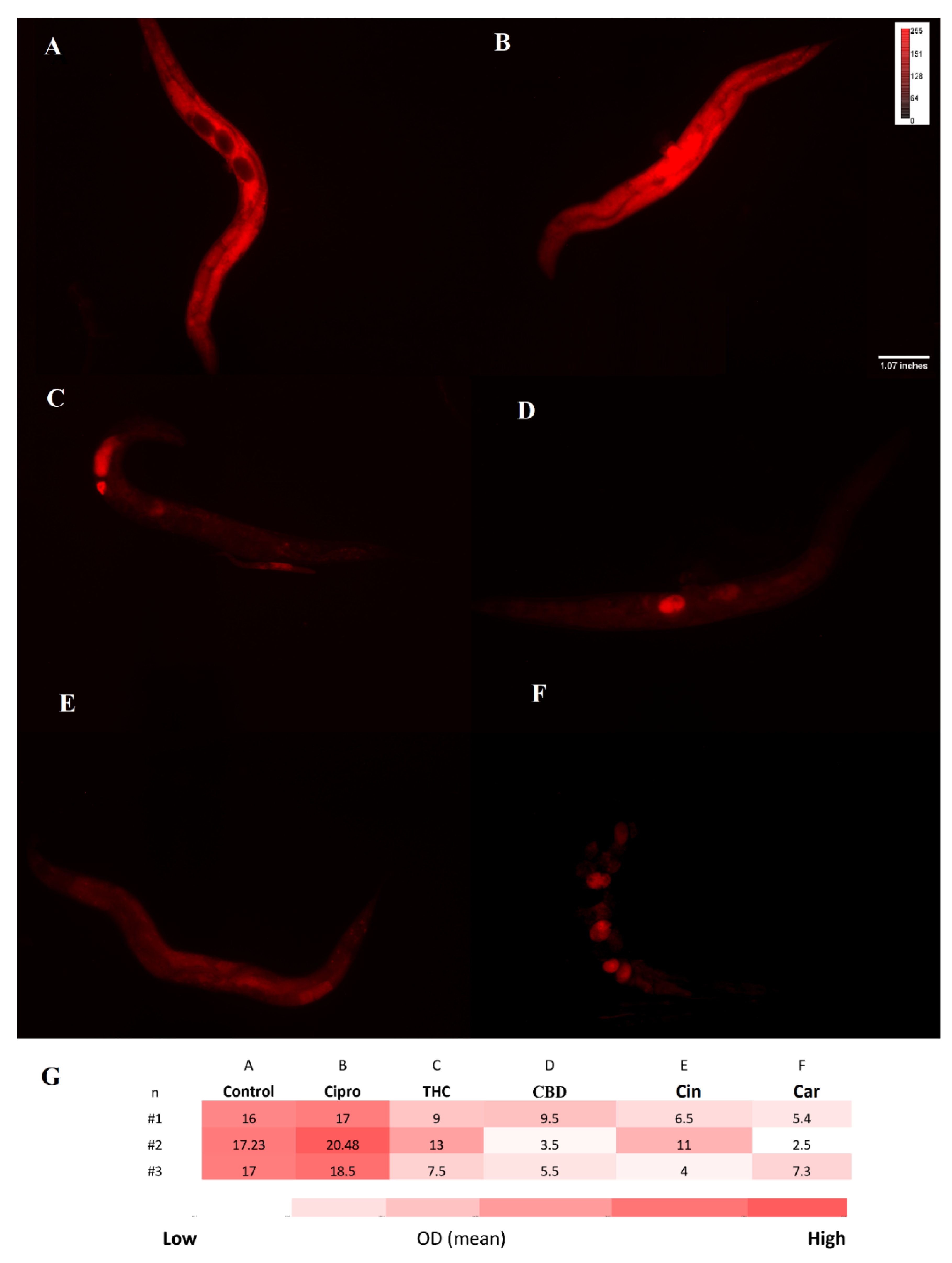
2.8. Toxicity of PBCs towards Caenorhabditis elegans
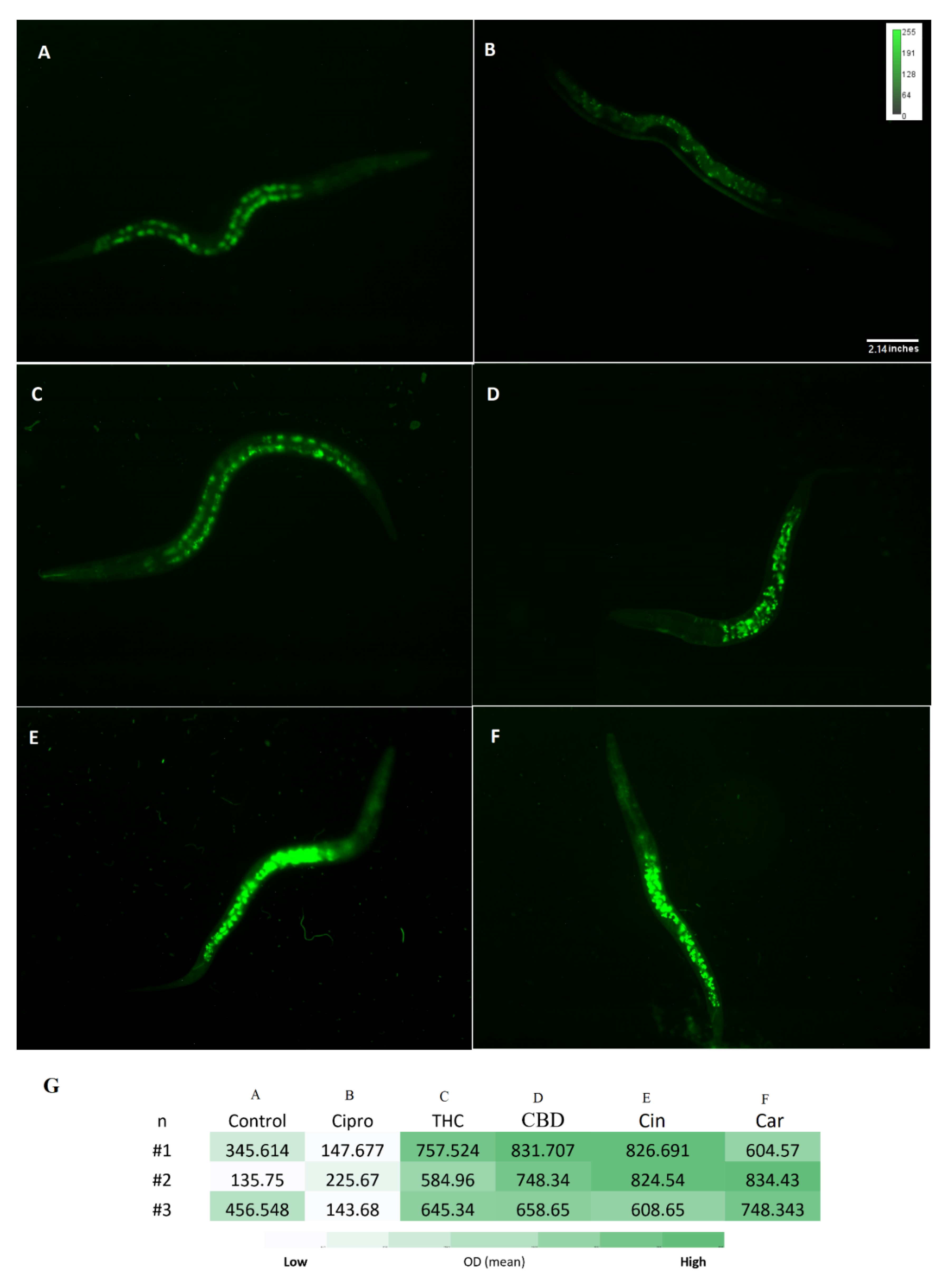
2.9. The Antioxidant Potency of PBCs in Caenorhabditis elegans Model
3. Discussion
4. Materials and Methods
4.1. Systematic Review for Selecting Highly Effective Plant-Based Natural Compounds (PBCs) as Antibacterial Agents
4.2. Bacterial Strains, Culture Media, Antibiotics, and Plant-Based Natural Compounds (PBCs)
4.3. Antibacterial and Antibiofilm Assays
4.4. Cannabis Sativa Oil Extraction
4.5. Hydrogen Peroxide Assay
4.6. Iron Detection Ferene-S Assay
4.7. Membrane Disruption and Permeability Measurements
4.8. Toxicity Test by the Caenorhabditis elegans Model
4.9. Antioxidant Test by the Caenorhabditis elegans Model
4.10. Statistical Tests and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug resistance: An emerging crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 541340. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; Gijón, D.; Ruiz-Garbajosa, P. Antimicrobial resistance in ICUs: An update in the light of the COVID-19 pandemic. Curr. Opin. Crit. Care 2020, 26, 433–441. [Google Scholar] [CrossRef] [PubMed]
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 27 February 2017).
- Shriram, V.; Khare, T.; Bhagwat, R.; Shukla, R.; Kumar, V. Inhibiting bacterial drug efflux pumps via phyto-therapeutics to combat threatening antimicrobial resistance. Front. Microbiol. 2018, 9, 2990. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, S.; Viljoen, A. Plant-based antimicrobial studies–methods and approaches to study the interaction between natural products. Planta Med. 2011, 77, 1168–1182. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Buss, A.D. Natural products—The future scaffolds for novel antibiotics? Biochem. Pharmacol. 2006, 71, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Debnath, T.; Kim, D.H.; Lim, B.O. Natural products as a source of anti-inflammatory agents associated with inflammatory bowel disease. Molecules 2013, 18, 7253–7270. [Google Scholar] [CrossRef]
- Witaicenis, A.; Seito, L.N.; Chagas, A.d.; Junior, L.D.d.; Luchini, A.C.; Rodrigues-Orsi, P.; Cestari, S.H.; di Stasi, L.C. Antioxidant and intestinal anti-inflammatory effects of plant-derived coumarin derivatives. Phytomedicine 2014, 21, 240–246. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-inflammatory activity of natural products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Caelli, M.; Porteous, J.; Carson, C.F.; Heller, R.; Riley, T.V. Tea tree oil as an alternative topical decolonization agent for methicillin-resistant Staphylococcus aureus. Int. J. Aromather. 2001, 11, 97–99. [Google Scholar] [CrossRef]
- Kwieciński, J.; Eick, S.; Wójcik, K. Effects of tea tree (Melaleuca alternifolia) oil on Staphylococcus aureus in biofilms and stationary growth phase. Int. J. Antimicrob. Agents 2009, 33, 343–347. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J. Resveratrol alleviates Staphylococcus aureus pneumonia by inhibition of the NLRP3 inflammasome. Exp. Ther. Med. 2017, 14, 6099–6104. [Google Scholar] [PubMed]
- Mun, S.-H.; Kim, S.; Kong, R.; Choi, J.; Kim, Y.; Shin, D.; Kang, O.; Kwon, D. Curcumin reverse methicillin resistance in Staphylococcus aureus. Molecules 2014, 19, 18283–18295. [Google Scholar] [CrossRef] [PubMed]
- Beya, M.M.; Netzel, M.E.; Sultanbawa, Y.; Smyth, H.; Hoffman, L.C. Plant-based phenolic molecules as natural preservatives in comminuted meats: A review. Antioxidants 2021, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Mahady, G.B.; Huang, Y.; Doyle, B.J.; Locklear, T. Natural products as antibacterial agents. Stud. Nat. Prod. Chem. 2008, 35, 423–444. [Google Scholar]
- Vu, T.T.; Kim, H.; Tran, V.K.; le Dang, Q.; Nguyen, H.T.; Kim, H.; Kim, I.S.; Choi, G.J.; Kim, J. In vitro antibacterial activity of selected medicinal plants traditionally used in Vietnam against human pathogenic bacteria. BMC Complementary Altern. Med. 2015, 16, 32. [Google Scholar] [CrossRef]
- Lelario, F.; Scrano, L.; de Franchi, S.; Bonomo, M.G.; Salzano, G.; Milan, S.; Milella, L.; Bufo, S.A. Identification and antimicrobial activity of most representative secondary metabolites from different plant species. Chem. Biol. Technol. Agric. 2018, 5, 13. [Google Scholar] [CrossRef]
- Martinenghi, L.D.; Jønsson, R.; Lund, T.; Jenssen, H. Isolation, Purification, and antimicrobial characterization of cannabidiolic acid and cannabidiol from Cannabis sativa L. Biomolecules 2020, 10, 900. [Google Scholar] [CrossRef]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical characterization and evaluation of the antibacterial activity of essential oils from fibre-type Cannabis sativa L. (Hemp). Molecules 2019, 24, 2302. [Google Scholar]
- Gonzalez, L.S., III; Spencer, J.P. Aminoglycosides: A practical review. Am. Fam. Physician 1998, 58, 1811. [Google Scholar]
- Chen, C.; Chen, Y.; Wu, P.; Chen, B. Update on new medicinal applications of gentamicin: Evidence-based review. J. Formos. Med. Assoc. 2014, 113, 72–82. [Google Scholar] [CrossRef] [PubMed]
- LeBel, M. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials, and adverse reactions. Pharmacother. J. Hum. Pharmacol. Drug Ther. 1988, 8, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Tangy, F.; Moukkadem, M.; Vindimian, E.; Capmau, M.L.; Le Goffic, F. Mechanism of action of gentamicin components: Characteristics of their binding to Escherichia coli ribosomes. Eur. J. Biochem. 1985, 147, 381–386. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 30, 2437. [Google Scholar] [CrossRef] [PubMed]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef]
- Rizvi, M.; Ahmed, J.; Khan, F.; Shukla, I.; Malik, A. Assessment of combination therapy by time kill curve analysis and chequerboard assay for treatment of multi-drug resistant Pseudomonas aeruginosa isolates. J. Glob. Antimicrob. Resist. 2013, 1, 103–108. [Google Scholar] [CrossRef]
- Schnaider, L.; Brahmachari, S.; Schmidt, N.W.; Mensa, B.; Shaham-Niv, S.; Bychenko, D.; Adler-Abramovich, L.; Shimon, L.J.W.; Kolusheva, S.; DeGrado, W.F. Self-assembling dipeptide antibacterial nanostructures with membrane disrupting activity. Nat. Commun. 2017, 8, 1365. [Google Scholar] [CrossRef]
- Brynildsen, M.P.; Winkler, J.A.; Spina, C.S.; MacDonald, I.C.; Collins, J.J. Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production. Nat. Biotechnol. 2013, 31, 160–165. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, C.; Jang, H.; Kim, B.; Bae, H.; Chung, I.; Kim, E.S.; Cho, Y. Antibacterial strategies inspired by the oxidative stress and response networks. J. Microbiol. 2019, 57, 203–212. [Google Scholar] [CrossRef]
- Calderon, I.L.; Elías, A.O.; Fuentes, E.L.; Pradenas, G.A.; Castro, M.E.; Arenas, F.A.; Pérez, J.M.; Vásquez, C.C. Tellurite-mediated disabling of [4Fe–4S] clusters of Escherichia coli dehydratases. Microbiology 2009, 155, 1840–1846. [Google Scholar] [CrossRef]
- Chillappagari, S.; Seubert, A.; Trip, H.; Kuipers, O.P.; Marahiel, M.A.; Miethke, M. Copper stress affects iron homeostasis by destabilizing iron-sulfur cluster formation in Bacillus subtilis. J. Bacteriol. 2010, 192, 2512–2524. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.F.; Imlay, J.A. Silver (I), mercury (II), cadmium (II), and zinc (II) target exposed enzymic iron-sulfur clusters when they toxify Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 3614–3621. [Google Scholar] [CrossRef] [PubMed]
- Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Maestri, D.; Nepote, V.; Lamarque, A.; Zygadlo, J. Natural products as antioxidants. Phytochem. Adv. Res. 2006, 37, 105–135. [Google Scholar]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Gill, R.; Tsung, A.; Billiar, T. Linking oxidative stress to inflammation: Toll-like receptors. Free Radic. Biol. Med. 2010, 48, 1121–1132. [Google Scholar] [CrossRef]
- Rahman, I.; MacNee, W. Oxidative stress and regulation of glutathione in lung inflammation. Eur. Respir. J. 2000, 16, 534–554. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Deans, S. Bioactivity of selected plant essential oils against Listeria monocytogenes. J. Appl. Microbiol. 1997, 82, 759–762. [Google Scholar] [CrossRef]
- Liu, J.; Yang, L.; Dong, Y.; Zhang, B.; Ma, X. Echinacoside, an inestimable natural product in treatment of neurological and other disorders. Molecules 2018, 23, 1213. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A.; et al. Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 5–23. [Google Scholar]
- Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef]
- Draenert, R.; Seybold, U.; Grützner, E.; Bogner, J. Novel antibiotics: Are we still in the pre–post-antibiotic era? Infection 2015, 43, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kmietowicz, Z. Few novel antibiotics in the pipeline, WHO warns. Br. Med. J. 2017, 358, j4339. [Google Scholar] [CrossRef] [PubMed]
- Cacciatore, I.; di Giulio, M.; Fornasari, E.; di Stefano, A.; Cerasa, L.S.; Marinelli, L.; Turkez, H.; di Campli, E.; di Bartolomeo, S.; Robuffo, I.; et al. Carvacrol codrugs: A new approach in the antimicrobial plan. PLoS ONE 2015, 10, e0120937. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, P.; García-Armesto, M.R.; Arriaga, D.; González-Donquiles, C.; Rodríguez-Fernández, P.; Rúa, J. Antimicrobial activity of kaempferol and resveratrol in binary combinations with parabens or propyl gallate against Enterococcus faecalis. Food Control 2016, 61, 213–220. [Google Scholar] [CrossRef]
- Brun, P.; Bernabè, G.; Filippini, R.; Piovan, A. In vitro antimicrobial activities of commercially available tea tree (Melaleuca alternifolia) essential oils. Curr. Microbiol. 2019, 76, 108–116. [Google Scholar] [CrossRef]
- Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Tenth Edition. Available online: https://clsi.org/media/1632/m07a10_sample.pdf (accessed on 1 January 2015).
- Khaki, P.; Sharma, A.; Bhalla, P. Comparison of two disc diffusion methods with minimum inhibitory concentration for antimicrobial susceptibility testing of Neisseria gonorrhoeae isolates. Ann. Med. Health Sci. Res. 2014, 4, 453–456. [Google Scholar]
- Maalej, S.; Meziou, M.; Rhimi, F.; Hammami, A. Comparison of disc diffusion, Etest and agar dilution for susceptibility testing of colistin against Enterobacteriaceae. Lett. Appl. Microbiol. 2011, 53, 546–551. [Google Scholar] [CrossRef]
- Van Klingeren, B.; Ten Ham, M. Antibacterial activity of Δ9-tetrahydrocannabinol and cannabidiol. Antonie Van Leeuwenhoek 1976, 42, 9–12. [Google Scholar] [CrossRef]
- Klement, E.; Chaffer, M.; Leitner, G.; Shwimmer, A.; Friedman, S.; Saran, A.; Shpigel, N. Assessment of accuracy of disk diffusion tests for the determination of antimicrobial susceptibility of common bovine mastitis pathogens: A novel approach. Microb. Drug Resist. 2005, 11, 342–350. [Google Scholar] [CrossRef]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for discovery of novel antibiotics. In How to Overcome the Antibiotic Crisis; Springer: Cham, Switzerland, 2016; Volume 398, pp. 303–338. [Google Scholar]
- Auckloo, B.N.; Pan, C.; Akhter, N.; Wu, B.; Wu, X.; He, S. Stress-driven discovery of novel cryptic antibiotics from a marine fungus Penicillium sp. BB1122. Front. Microbiol. 2017, 8, 1450. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Proksch, P. Fifty years of drug discovery from fungi. Fungal Divers. 2011, 50, 3–19. [Google Scholar] [CrossRef]
- Loferer, H. Mining bacterial genomes for antimicrobial targets. Mol. Med. Today 2000, 6, 470–474. [Google Scholar] [CrossRef]
- Dougherty, T.J.; Barrett, J.F.; Pucci, M.J. Microbial genomics and novel antibiotic discovery: New technology to search for new drugs. Curr. Pharm. Des. 2002, 8, 1119–1135. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, D.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel anti-infective compounds from marine bacteria. Mar. Drugs 2010, 8, 498–518. [Google Scholar] [CrossRef] [PubMed]
- Stincone, P.; Brandelli, A. Marine bacteria as source of antimicrobial compounds. Crit. Rev. Biotechnol. 2020, 40, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Schinke, C.; Martins, T.; Queiroz, S.C.; Melo, I.S.; Reyes, F.G. Antibacterial compounds from marine bacteria, 2010–2015. J. Nat. 2017, 80, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M.; Hartzell, W.O.; Robbins, C.S.; Gomis-Rüth, F.X.; Shapiro, S.D. Macrophage elastase kills bacteria within murine macrophages. Nature 2009, 460, 637–641. [Google Scholar] [CrossRef]
- Ymele-Leki, P.; Cao, S.; Sharp, J.; Lambert, K.G.; McAdam, A.J.; Husson, R.N.; Tamayo, G.; Clardy, J.; Watnick, P.I. A high-throughput screen identifies a new natural product with broad-spectrum antibacterial activity. PLoS ONE 2012, 7, e31307. [Google Scholar] [CrossRef]
- Jensen, P.R.; Fenical, W. Marine microorganisms and drug discovery: Current status and future potential. In Drugs from the Sea; Karger: Basel, Switzerland, 2000; pp. 6–29. [Google Scholar]
- Doshi, G.; Aggarwal, G.; Martis, E.; Shanbhag, P. Novel antibiotics from marine sources. Int. J. Pharm. Sci. Nanotechnol. 2011, 4, 1446–1461. [Google Scholar]
- Ellis, M.J.; Cao, S.; Sharp, J.; Lambert, K.G.; McAdam, A.J.; Husson, R.N.; Tamayo, G.; Clardy, J.; Watnick, P.I. A macrophage-based screen identifies antibacterial compounds selective for intracellular Salmonella typhimurium. Nat. Commun. 2019, 10, 197. [Google Scholar] [CrossRef]
- Christophe, T.; Ewann, F.; Jeon, H.K.; Cechetto, J.; Brodin, P. High-content imaging of Mycobacterium tuberculosis-infected macrophages: An in vitro model for tuberculosis drug discovery. Future Med. Chem. 2010, 2, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Antimicrobial Peptides: Discovery, Design and Novel Therapeutic Strategies; CABI: Wallingford, UK, 2010. [Google Scholar]
- Hancock, R.; Patrzykat, A. Clinical development of cationic antimicrobial peptides: From natural to novel antibiotics. Curr. Drug Targets-Infect. Disord. 2002, 2, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Sierra, J.M.; Fusté, E.; Rabanal, F.; Vinuesa, T.; Viñas, M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, T.; Venkatanagaraju, E.; Goli, D.; Basha, S.J. Evaluation of antimicrobial activity of different herbal plant extracts. Int. J. Pharm. Sci. Res. 2014, 5, 1460. [Google Scholar]
- Murugan, T.; Saranraj, P. Antibacterial activity of various solvent extracts of the Indian herbal plant Acalypha indica against human pathogens causing nosocomial infection. Int. J. Pharm. Biol. Arch. 2011, 2, 1498–1503. [Google Scholar]
- Hemeg, H.A.; Moussa, I.M.; Ibrahim, S.; Dawoud, T.M.; Alhaji, J.H.; Mubarak, A.S.; Kabli, S.A.; Alsubki, R.A.; Tawfik, A.M.; Marouf, S.A. Antimicrobial effect of different herbal plant extracts against different microbial population. Saudi J. Biol. Sci. 2020, 27, 3221–3227. [Google Scholar] [CrossRef]
- Farzaneh, V.; Carvalho, I.S. A review of the health benefit potentials of herbal plant infusions and their mechanism of actions. Ind. Crops Prod. 2015, 65, 247–258. [Google Scholar] [CrossRef]
- Talib, W.H. Anticancer and Antimicrobial Potential of Plant-Derived Natural Products. In Phytochemicals–Bioactivities and Impact on Health; Intech Open: London, UK, 2011; pp. 141–158. [Google Scholar]
- Abdi, R.D.; Dego, O.K. Antimicrobial activity of Persicaria pensylvanica extract against Staphylococcus aureus. Eur. J. Integr. Med. 2019, 29, 100921. [Google Scholar] [CrossRef]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial cannabinoids from Cannabis sativa: A structure− activity study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Molnar, J.; Csiszar, K.; Nishioka, I.; Shoyama, Y. The effects of cannabispiro compounds and tetrahydrocannabidiolic acid on the plasmid transfer and maintenance in Escherichia coli. Acta Microbiol. Hung. 1986, 33, 221–231. [Google Scholar] [PubMed]
- Baquero, F.; Levin, B.R. Proximate and ultimate causes of the bactericidal action of antibiotics. Nat. Rev. Microbiol. 2021, 19, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Abdi-Ali, A.; Mohammadi-Mehr, M.; Alaei, Y.A. Bactericidal activity of various antibiotics against biofilm-producing Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2006, 27, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Moser, C.; Jensen, P.Ø.; Høiby, N. Tolerance and resistance of microbial biofilms. Nat. Rev. Microbiol. 2022, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pormohammad, A.; Turner, R.J. Silver antibacterial synergism activities with eight other metal (loid)-based antimicrobials against Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus. Antibiotics 2020, 9, 853. [Google Scholar] [CrossRef]
- Eckert, R.; Sullivan, R.; Shi, W. Targeted antimicrobial treatment to re-establish a healthy microbial flora for long-term protection. Adv. Dent. Res. 2012, 24, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C. Mechanisms of antimicrobial resistance in bacteria. Am. J. Med. 2006, 119, S3–S10. [Google Scholar] [CrossRef]
- Martinez, J.L.; Sánchez, M.B.; Martínez-Solano, L.; Hernandez, A.; Garmendia, L.; Fajardo, A.; Alvarez-Ortega, C. Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol. Rev. 2009, 33, 430–449. [Google Scholar] [CrossRef]
- Eckhardt, S.; Brunetto, P.S.; Gagnon, J.; Priebe, M.; Giese, B.; Fromm, K.M. Nanobio silver: Its interactions with peptides and bacteria, and its uses in medicine. Chem. Rev. 2013, 113, 4708–4754. [Google Scholar] [CrossRef] [PubMed]
- Wijesundara, N.M.; Lee, S.F.; Cheng, Z.; Davidson, R.; Rupasinghe, H. Carvacrol exhibits rapid bactericidal activity against Streptococcus pyogenes through cell membrane damage. Sci. Rep. 2021, 11, 1487. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, F.; Ji, B.P.; Pei, R.S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Amos, L.A.; van den Ent, F.; Löwe, J. Structural/functional homology between the bacterial and eukaryotic cytoskeletons. Curr. Opin. Cell Biol. 2004, 16, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xiong, T.; Meng, X.; Yu, D.; Xiao, Z.; Song, L. Different influences on mitochondrial function, oxidative stress and cytotoxicity of antibiotics on primary human neuron and cell lines. J. Biochem. Mol. Toxicol. 2019, 33, e22277. [Google Scholar] [CrossRef] [PubMed]
- Pals, J.; Attene-Ramos, M.S.; Xia, M.; Wagner, E.D.; Plewa, M.J. Human cell toxicogenomic analysis linking reactive oxygen species to the toxicity of monohaloacetic acid drinking water disinfection byproducts. Environ. Sci. Technol. 2013, 47, 12514–12523. [Google Scholar] [CrossRef] [PubMed]
- Land, M.H.; Toth, M.L.; MacNair, L.; Vanapalli, S.A.; Lefever, T.W.; Peters, E.N.; Bonn-Miller, M.O. Effect of cannabidiol on the long-term toxicity and lifespan in the preclinical model Caenorhabditis elegans. Cannabis Cannabinoid Res. 2020, 6, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-C.; Yen, P.; Chang, S.; Cheng, P.; Lo, Y.; Liao, V.H. Antioxidative activities of both oleic acid and Camellia tenuifolia seed oil are regulated by the transcription factor DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE 2016, 11, e0157195. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Shen, P.; Xu, Y.; Park, Y. p-Coumaric acid improves oxidative and osmosis stress responses in Caenorhabditis elegans. J. Sci. Food Agric. 2019, 99, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Ergen, N.; Hoşbaş, S.; Orhan, D.D.; Aslan, M.; Sezik, E.; Atalay, A. Evaluation of the lifespan extension effects of several Turkish medicinal plants in Caenorhabditis elegans. Turk. J. Biol. 2018, 42, 163–173. [Google Scholar] [CrossRef]
- Liu, L.; Guo, P.; Wang, P.; Zheng, S.; Qu, Z.; Liu, N. The review of anti-aging mechanism of polyphenols on Caenorhabditis elegans. Front. Bioeng. Biotechnol. 2021, 9, 635768. [Google Scholar] [CrossRef] [PubMed]
- Pierce™ Quantitative Peroxide Assay Kits. Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https%3A%2F%2Fassets.thermofisher.com%2FTFS-Assets%2FLSG%2Fmanuals%2FMAN0011275_Pierce_Quant_Peroxide_Asy_UG.pdf (accessed on 13 October 2018).
- Hennessy, D.J.; Reid, G.R.; Smith, F.E.; Thompson, S.L. Ferene—A new spectrophotometric reagent for iron. Can. J. Chem. 1984, 62, 721–724. [Google Scholar] [CrossRef]
- Morones-Ramirez, J.R.; Winkler, J.A.; Spina, C.S.; Collins, J.J. Silver enhances antibiotic activity against gram-negative bacteria. Sci. Transl. Med. 2013, 5, 190ra181. [Google Scholar] [CrossRef] [PubMed]
- Novo, D.J.; Perlmutter, N.G.; Hunt, R.H.; Shapiro, H.M. Multiparameter flow cytometric analysis of antibiotic effects on membrane potential, membrane permeability, and bacterial counts of Staphylococcus aureus and Micrococcus luteus. Antimicrob. Agents Chemother. 2000, 44, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, J.A.; Beceiro, A.; Rumbo-Feal, S.; Rodríguez-Palero, M.J.; Russo, T.A.; Bou, G. Optimisation of the Caenorhabditis elegans model for studying the pathogenesis of opportunistic Acinetobacter baumannii. Int. J. Antimicrob. Agents 2015, 43, 203–220. [Google Scholar] [CrossRef]
- Cnubben, N.H.; Rietjens, I.M.; Wortelboer, H.; van Zanden, J.; van Bladeren, P.J. The interplay of glutathione-related processes in antioxidant defense. Environ. Toxicol. Pharmacol. 2001, 10, 141–152. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- De Santos, W.M.; Secoli, S.R.; de Püschel, V.A.A. The Joanna Briggs Institute approach for systematic reviews. Rev. Lat. Am. Enferm. 2018, 26, e3074. [Google Scholar] [CrossRef]
- AR Bank # 0033 Acinetobacter baumannii. Available online: https://wwwn.cdc.gov/ARIsolateBank/Panel/IsolateDetail?IsolateID=33 (accessed on 31 March 2021).
- Lemire, J.A.; Kalan, L.; Bradu, A.; Turner, R.J. Silver oxynitrate, an unexplored silver compound with antimicrobial and antibiofilm activity. Antimicrob. Agents Chemother. 2015, 59, 4031–4039. [Google Scholar] [CrossRef]
- Monych, N.K.; Turner, R.J. Multiple Compounds Secreted by Pseudomonas aeruginosa Increase the Tolerance of Staphylococcus aureus to the Antimicrobial Metals Copper and Silver. mSystems 2020, 5, e00746-20. [Google Scholar] [CrossRef]
- Agarwal, G.; Kapil, A.; Kabra, S.; Das, B.K.; Dwivedi, S. In vitro efficacy of ciprofloxacin and gentamicin against a biofilm of Pseudomonas aeruginosa and its free-living forms. Natl. Med. J. India 2005, 18, 184. [Google Scholar] [PubMed]
- Diepoltová, A.; Konečná, K.; Janďourek, O.; Nachtigal, P. Study of the impact of cultivation conditions and peg surface modification on the in vitro biofilm formation of Staphylococcus aureus and Staphylococcus epidermidis in a system analogous to the Calgary biofilm device. J. Med. Microbiol. 2021, 70, 001371. [Google Scholar] [CrossRef] [PubMed]
- Indicators for Highly Reactive Oxygen Species. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/mp36003.pdf (accessed on 9 May 2005).
- Jung, H.J.; Lee, D.G. Synergistic antibacterial effect between silybin and N, N′-dicyclohexylcarbodiimide in clinical Pseudomonas aeruginosa isolates. J. Microbiol. 2008, 46, 462–467. [Google Scholar] [CrossRef]
- Linnett, P.; Beechey, R. Inhibitors of the ATP synthetase systems. Methods Enzymol. 1979, 55, 472–518. [Google Scholar]
- Stiernagle, T. Maintenance of C. elegans (11 February 2006); The C. elegans Research Community, WormBook: Pasadena, CA, USA, 2006. [Google Scholar] [CrossRef]
- Karas, J.A.; Wong, L.J.M.; Paulin, O.K.A.; Mazeh, A.C.; Hussein, M.H.; Li, J.; Velkov, T. The antimicrobial activity of cannabinoids. Antibiotics 2020, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.M.; Elsohly, M.A.; Slade, D.; Ahmed, S.A.; Khan, I.A.; Ross, S.A. Biologically active cannabinoids from high-potency Cannabis sativa. J. Nat. Prod. 2009, 72, 906–911. [Google Scholar] [CrossRef]
- Sarmadyan, H.; Solhi, H.; Najarian-Araghi, N.; Ghaznavi-Rad, E. Determination of the Antimicrobial Effects of Hydro-Alcoholic Extract of Cannabis Sativa on Multiple Drug Resistant Bacteria Isolated from Nosocomial Infections. Iran. J. Toxicol. 2014, 7, 967–972. [Google Scholar]
- Zengin, G.; Menghini, L.; di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A. Chromatographic analyses, in vitro biological activities, and cytotoxicity of Cannabis sativa L. essential oil: A multidisciplinary study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef]
- Palmieri, S.; Maggio, F.; Pellegrini, M.; Ricci, A.; Serio, A.; Paparella, A.; Sterzo, C.L. Effect of the Distillation Time on the Chemical Composition, Antioxidant Potential and Antimicrobial Activity of Essential Oils from Different Cannabis sativa L. Cultivars. Molecules 2021, 26, 4770. [Google Scholar] [CrossRef]
- Muscarà, C.; Smeriglio, A.; Trombetta, D.; Mandalari, G.; la Camera, E.; Grassi, G.; Circosta, C. Antioxidant and antimicrobial activity of two standardized extracts from a new Chinese accession of non-psychotropic Cannabis sativa L. Phytother. Res. 2021, 35, 1099–1112. [Google Scholar] [CrossRef]
- Blaskovich, M.A.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J. The antimicrobial potential of cannabidiol. Commun. Biol. 2021, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Paulo, L.; Ferreira, S.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Antimicrobial activity and effects of resveratrol on human pathogenic bacteria. World J. Microbiol. Biotechnol. 2010, 26, 1533–1538. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Zetterström, C.E.; Hasselgren, J.; Salin, O.; Davis, R.A.; Quinn, R.J.; Sundin, C.; Elofsson, M. The resveratrol tetramer (-)-hopeaphenol inhibits type III secretion in the gram-negative pathogens Yersinia pseudotuberculosis and Pseudomonas aeruginosa. PLoS ONE 2013, 8, e81969. [Google Scholar]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Encinar, J.A.; Rodríguez-Díaz, J.C.; Micol, V. Antimicrobial capacity of plant polyphenols against gram-positive bacteria: A comprehensive review. Curr. Med. Chem. 2020, 27, 2576–2606. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Hurdle, J.G.; Lee, R.; Lee, R.; Cushman, M.; Pezzuto, J.M. Evaluation of flavonoid and resveratrol chemical libraries reveals abyssinone II as a promising antibacterial lead. ChemMedChem 2012, 7, 1541. [Google Scholar] [CrossRef] [PubMed]
- Makobongo, M.O.; Gilbreath, J.J.; Merrell, D.S. Nontraditional therapies to treat Helicobacter pylori infection. J. Microbiol. 2014, 52, 259–272. [Google Scholar] [CrossRef]
- Lemos, M.F.; Lemos, M.F.; Pacheco, H.P.; Guimarães, A.C.; Fronza, M.; Endringer, D.C.; Scherer, R. Seasonal variation affects the composition and antibacterial and antioxidant activities of Thymus vulgaris. Ind. Crops Prod. 2017, 95, 543–548. [Google Scholar] [CrossRef]
- Imelouane, B.; Amhamdi, H.; Wathelet, J.P.; Ankit, M.; Khedid, K.; El Bachiri, A. Chemical composition and antimicrobial activity of essential oil of thyme (Thymus vulgaris) from Eastern Morocco. Int. J. Agric. Biol. 2009, 11, 205–208. [Google Scholar]
- Ahmad, A.; Van Vuuren, S.; Viljoen, A. Unravelling the complex antimicrobial interactions of essential oils—The case of Thymus vulgaris (Thyme). Molecules 2014, 19, 2896–2910. [Google Scholar] [CrossRef]
- Fournomiti, M.; Kimbaris, A.; Mantzourani, I.; Plessas, S.; Theodoridou, I.; Papaemmanouil, V.; Kapsiotis, I.; Panopoulou, M.; Stavropoulou, E.; Bezirtzoglou, E.E.; et al. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microb. Ecol. Health Dis. 2015, 26, 23289. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, E.J.; Tjeerdsma-van Bokhoven, J.L.; Zweijtzer, C.; Burt, S.A.; Haagsman, H.P. Structural requirements for the antimicrobial activity of carvacrol. J. Agric. Food Chem. 2006, 54, 1874–1879. [Google Scholar] [CrossRef]
- Guarda, A.; Rubilar, J.F.; Miltz, J.; Galotto, M.J. The antimicrobial activity of microencapsulated thymol and carvacrol. Int. J. Food Microbiol. 2011, 146, 144–150. [Google Scholar] [CrossRef]
- Rúa, J.; Del Valle, P.; de Arriaga, D.; Fernández-Álvarez, L.; García-Armesto, M.R. Combination of carvacrol and thymol: Antimicrobial activity against Staphylococcus aureus and antioxidant activity. Foodborne Pathog. Dis. 2019, 16, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Issam, A.-A.; Zimmermann, S.; Reichling, J.; Wink, M. Pharmacological synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi-drug resistant microbial pathogens. Phytomedicine 2015, 22, 245–255. [Google Scholar]
- Al-Mariri, A.; Swied, G.; Oda, A.; Al Hallab, L. Antibacterial activity of thymus syriacus boiss essential oil and its components against some Syrian gram-negative bacteria isolates. Iran. J. Med. Sci. 2013, 38, 180. [Google Scholar] [PubMed]
- Du, E.; Gan, L.; Li, Z.; Wang, W.; Liu, D.; Guo, Y. In vitro antibacterial activity of thymol and carvacrol and their effects on broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2015, 6, 58. [Google Scholar] [CrossRef]
- Song, Y.-M.; Zhou, H.; Wu, Y.; Wang, J.; Liu, Q.; Mei, Y. In vitro evaluation of the antibacterial properties of tea tree oil on planktonic and biofilm–forming Streptococcus mutans. AAPS PharmSciTech 2020, 21, 227. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhang, X.; Guo, N. The antimicrobial activities and action-mechanism of tea tree oil against food-borne bacteria in fresh cucumber juice. Microb. Pathog. 2018, 125, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Karpanen, T.J.; Worthington, T.; Hendry, E.; Conway, B.R.; Lambert, P.A. Antimicrobial efficacy of chlorhexidine digluconate alone and in combination with eucalyptus oil, tea tree oil and thymol against planktonic and biofilm cultures of Staphylococcus epidermidis. J. Antimicrob. Chemother. 2008, 62, 1031–1036. [Google Scholar] [CrossRef]
- Low, W.; Martin, C.; Hill, D.; Kenward, M. Antimicrobial efficacy of silver ions in combination with tea tree oil against Pseudomonas aeruginosa, Staphylococcus aureus and Candida albicans. Int. J. Antimicrob. Agents 2011, 37, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Firmino, D.F.; Cavalcante, T.T.A.; Gomes, G.A.; Firmino, N.C.S.; Rosa, L.D.; de Carvalho, M.G.; Catunda, F.E.A., Jr. Antibacterial and antibiofilm activities of Cinnamomum sp. essential oil and cinnamaldehyde: Antimicrobial activities. Sci. World J. 2018, 2018, 7405736. [Google Scholar] [CrossRef]
- Ali, S.M.; Khan, A.A.; Ahmed, I.; Musaddiq, M.; Ahmed, K.S.; Polasa, H.; Rao, L.V.; Habibullah, C.M.; Sechi, L.A.; Ahmed, N. Antimicrobial activities of Eugenol and Cinnamaldehyde against the human gastric pathogen Helicobacter pylori. Ann. Clin. Microbiol. Antimicrob. 2005, 4, 20. [Google Scholar] [CrossRef]
- He, Z.; Huang, Z.; Jiang, W.; Zhou, W. Antimicrobial activity of cinnamaldehyde on Streptococcus mutans biofilms. Front. Microbiol. 2019, 10, 2241. [Google Scholar] [CrossRef]
- Krist, S.; Banovac, D.; Tabanca, N.; Wedge, D.E.; Gochev, V.K.; Wanner, J.; Schmidt, E.; Jirovetz, L. Antimicrobial activity of nerolidol and its derivatives against airborne microbes and further biological activities. Nat. Prod. Commun. 2015, 10, 1934578X1501000133. [Google Scholar] [CrossRef]
- Khatkar, A.; Nanda, A.; Kumar, P.; Narasimhan, B. Synthesis, antimicrobial evaluation and QSAR studies of p-coumaric acid derivatives. Arab. J. Chem. 2017, 10, S3804–S3815. [Google Scholar] [CrossRef]
- Forero-Doria, O.; Araya-Maturana, R.; Barrientos-Retamal, A.; Morales-Quintana, L.; Guzmán, L. N-alkylimidazolium salts functionalized with p-coumaric and cinnamic acid: A study of their antimicrobial and antibiofilm effects. Molecules 2019, 24, 3484. [Google Scholar] [CrossRef] [PubMed]
- Jorge, R.; Furtado, N.A.J.C.; Sousa, J.P.B.D.; da Silva Filho, A.A.; Gregório Junior, L.E.; Martins, C.H.G.; Soares, A.E.E.; Bastos, J.K.; Cunha, W.R.; Silva, M.L.A. Brazilian propolis: Seasonal variation of the prenylated p-coumaric acids and antimicrobial activity. Pharm. Biol. 2008, 46, 889–893. [Google Scholar] [CrossRef]
- Mandroli, P.S.; Bhat, K. An in-vitro evaluation of antibacterial activity of curcumin against common endodontic bacteria. J. Appl. Pharm. Sci. 2013, 3, 16. [Google Scholar]
- Izui, S.; Sekine, S.; Maeda, K.; Kuboniwa, M.; Takada, A.; Amano, A.; Nagata, H. Antibacterial activity of curcumin against periodontopathic bacteria. J. Periodontol. 2016, 87, 83–90. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | The Most Effective Antibiofilm and Antibacterial PBCs | |||
|---|---|---|---|---|
| PBCs | MIC (μg/mL) | MBC (μg/mL) | MBIC (μg/mL) | |
| S. aureus | CBD | 0.026–0.8 | 62–125 | 125–150 |
| THC | 62–125 | 62–125 | 125–150 | |
| A. baumannii | Cinnamaldehyde | 1.5–3.96 | 126.5–131.25 | 4–12.5 |
| Carvacrol | 0.225–3.12 | 6.25–25 | 1–62.5 | |
| Thymol | 12.5–62.5 | 19.5–125 | 9–25 | |
| K. pneumoniae | Thymol | 30–62.5 | 30–64 | 30–65 |
| E. coli | CBD | 10–12.5 | 12.5–25 | 125–250 |
| Carvacrol | 25–50 | 36–50 | 36–125 | |
| P. aeruginosa | CBD | 62–125 | 125–250 | 125–500 |
| THC | 62–125 | 125 | 125–250 | |
| P. mirabilis | Thymol | 30–125 | 60–125 | 30–125 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pormohammad, A.; Hansen, D.; Turner, R.J. Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin. Antibiotics 2022, 11, 1099. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081099
Pormohammad A, Hansen D, Turner RJ. Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin. Antibiotics. 2022; 11(8):1099. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081099
Chicago/Turabian StylePormohammad, Ali, Dave Hansen, and Raymond J. Turner. 2022. "Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin" Antibiotics 11, no. 8: 1099. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081099