

Antiparasitic Activities of Compounds Isolated from Aspergillus fumigatus Strain Discovered in Northcentral Nigeria

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterisation and Identification of Strain UIAU-3F
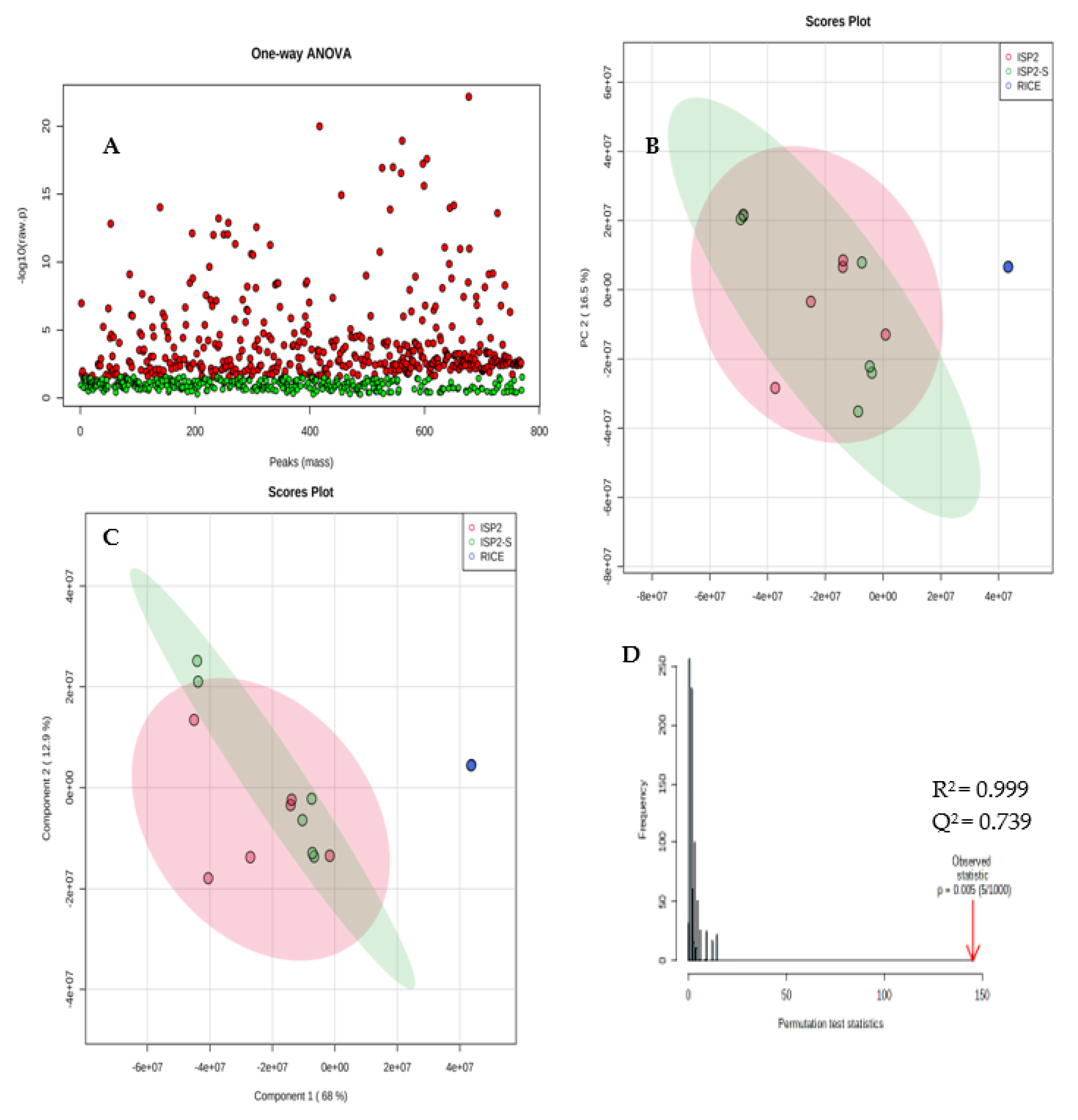
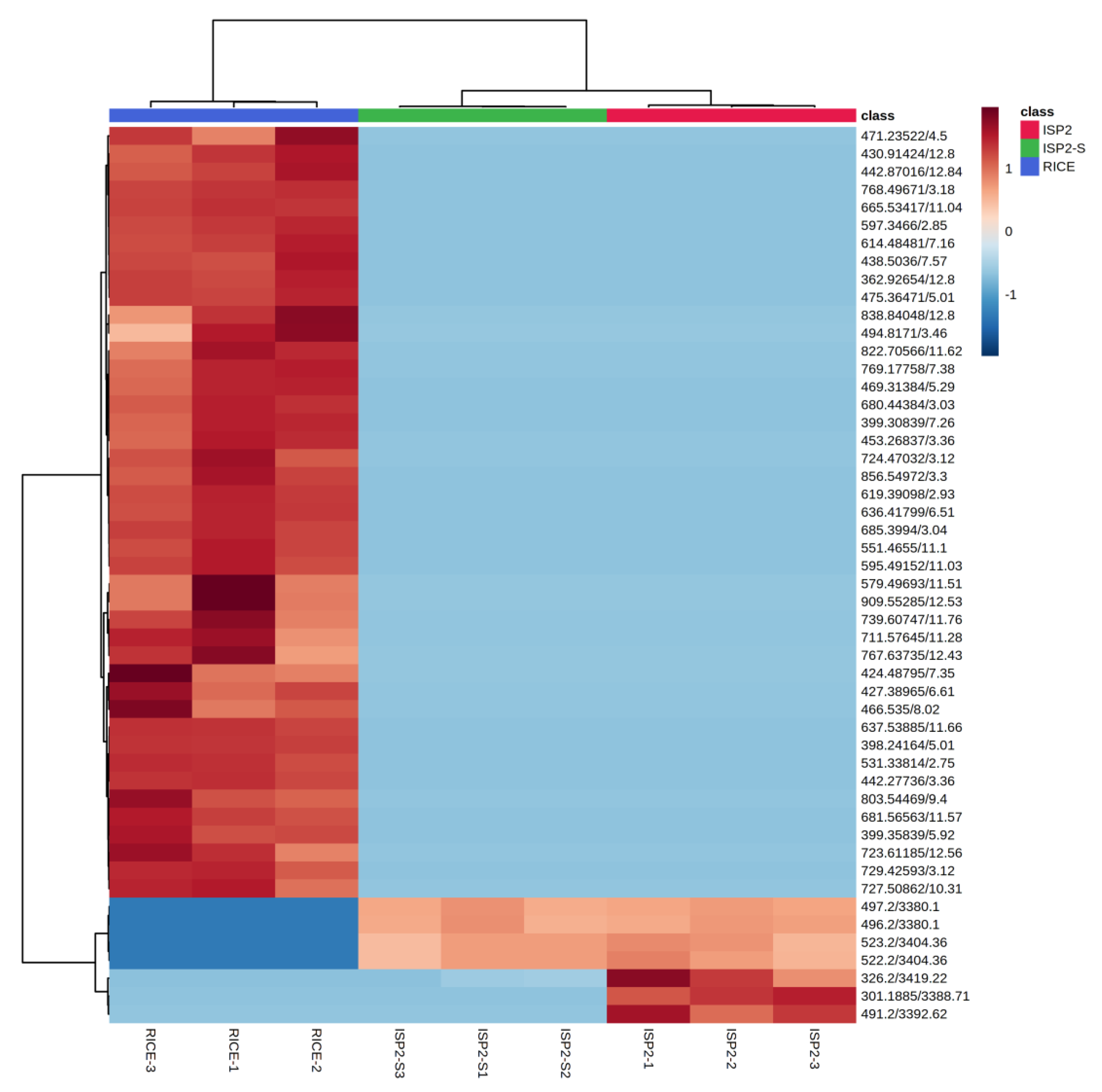
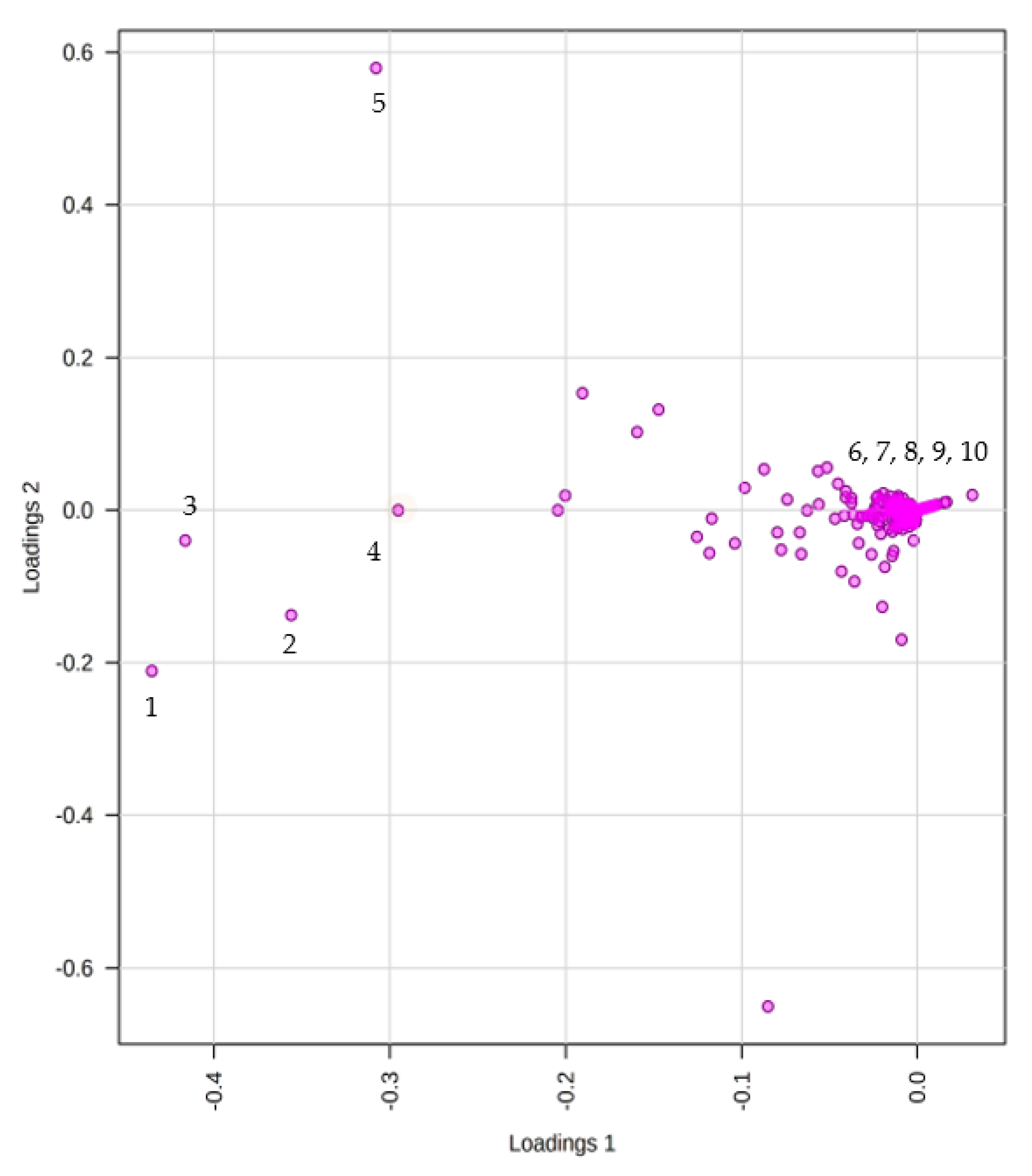
2.2. Non-Targeted Multivariate Analysis
Metabolites Identification
2.3. Structural Elucidation of Isolated Compounds
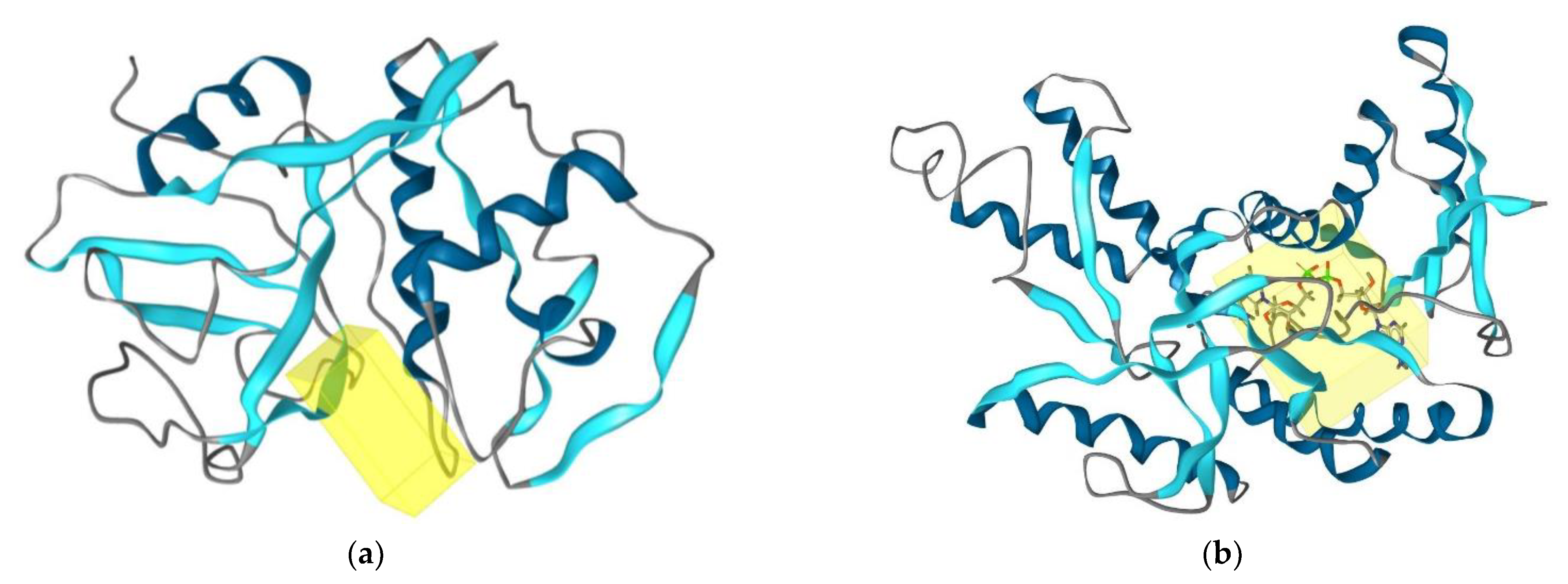
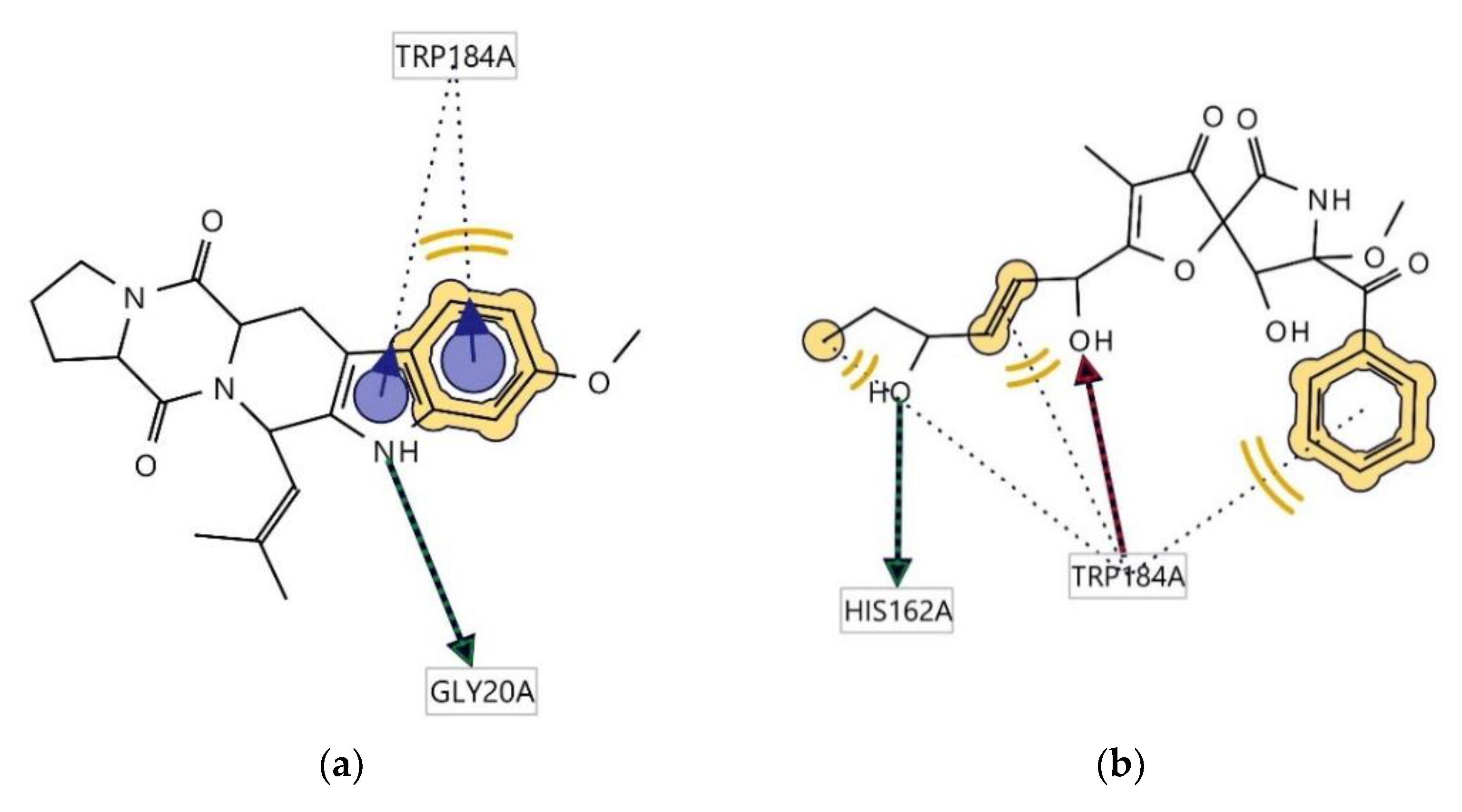
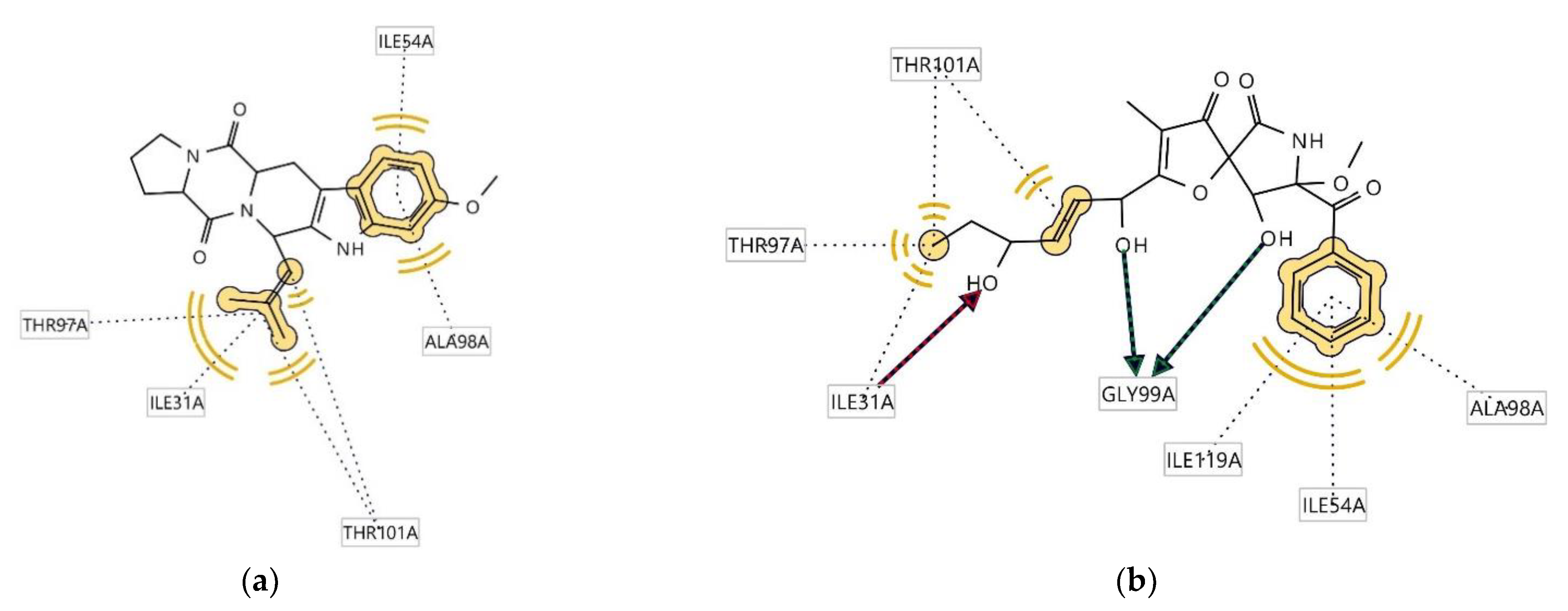
2.4. Determination of Antitrypanosomal and Antiplasmodium Activity Using Molecular Docking
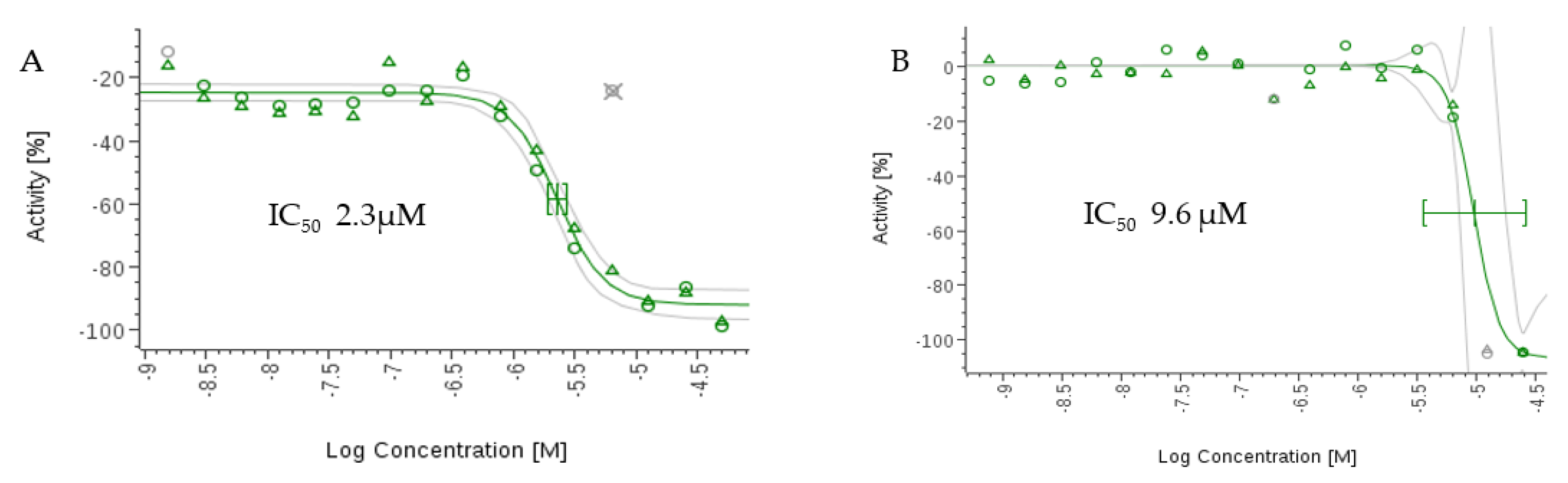
2.5. In Vitro Antiparasitic Activity
3. Material and Methods
3.1. General Procedure
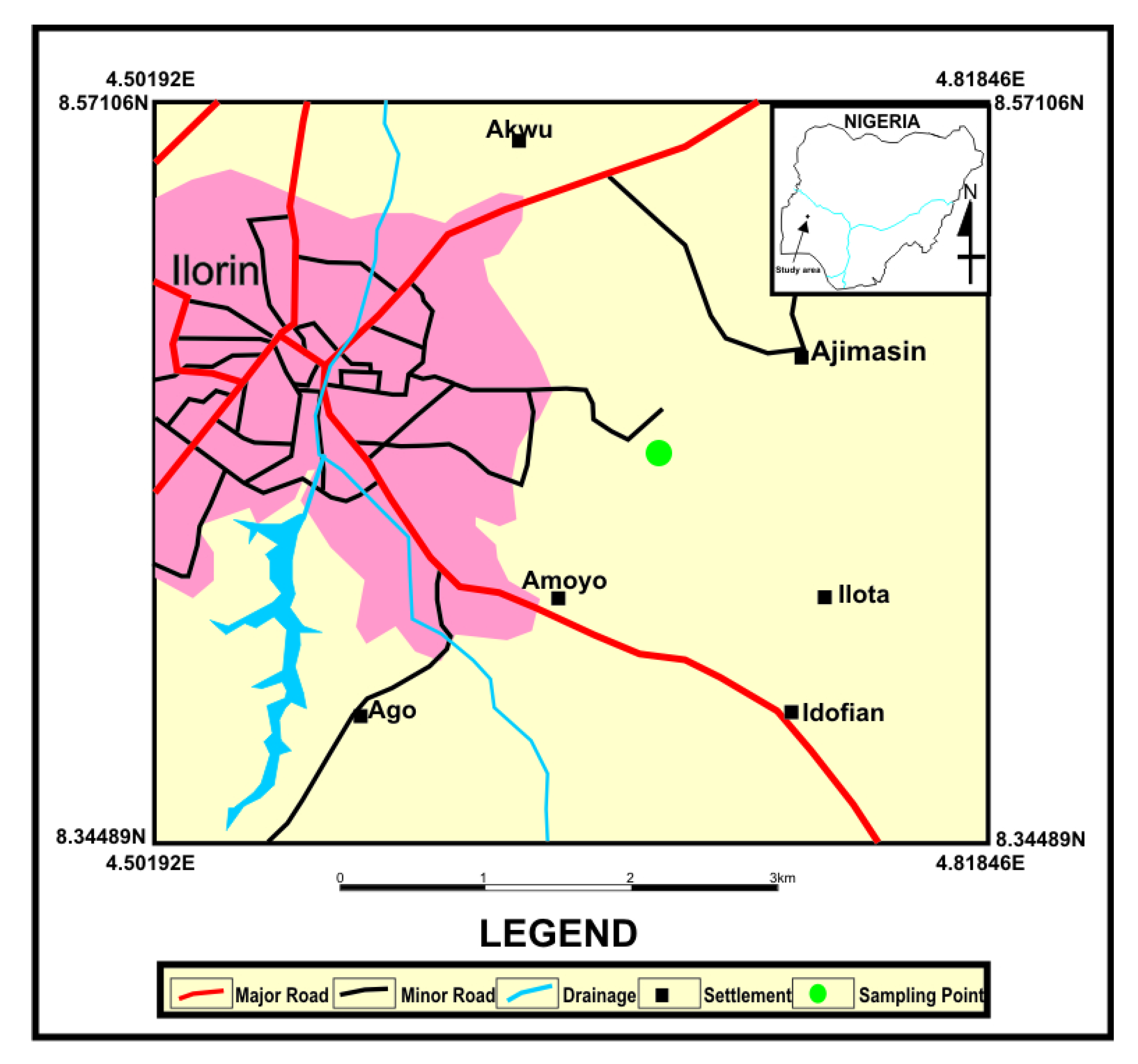
3.2. Aspergillus fumigatus Collection
3.3. Chemical Reagents
3.4. DNA Isolation and Molecular Identification
3.5. Fermentation and Extraction
3.6. Metabolic Profiling and Multivariate Statistical Analysis
3.7. Isolation of Metabolites
3.8. Molecular Docking
3.9. In Vitro Antitrypanosomal Activity
3.10. Plasmodium falciparum 3D7 Lactase Dehydrogenase In-Vitro Assay
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klich, M.A. Biogeography of Aspergillus Species in Soil and Litter. Mycologia 2002, 94, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Lević, J.; Gošič-dondo, S.N.; Ivanović, D.; Stanković, S.; Krnjaja, V.; Bočarov-Stančić, A.; Stepanić, A. An outbreak of Aspergillus species in response to environmental conditions in Serbia. Pestic. Phytomed./Pestic. Fitomed. 2013, 7, 28. [Google Scholar]
- Frisvad, J.C.; Rank, C.; Nielsen, K.F.; Larsen, T.O. Metabolomics of Aspergillus fumigatus. Med. Mycol. 2009, 47 (Suppl. S1), S53–S71. [Google Scholar] [CrossRef] [PubMed]
- Vadlapudi, V.; Borah, N.; Yellusani, K.R.; Gade, S.; Reddy, P.; Rajamanikyam, M.; Vempati, L.N.S.; Gubbala, S.P.; Chopra, P.; Upadhyayula, S.M.; et al. Aspergillus Secondary Metabolite Database, a resource to understand the Secondary metabolome of Aspergillus genus. Sci. Rep. 2017, 7, 7325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-Y.; Song, Y.; Zhang, Z.; Wang, L.; Guo, Z.; Zou, W.; Tan, R. Aspergillus fumigatus CY018, an endophytic fungus in Cynodon dactylon as a versatile producer of new and bioactive metabolites. J. Biotechnol. 2004, 114, 279–287. [Google Scholar] [CrossRef]
- Waksman, S.A.; Geiger, W.B. The Nature of the Antibiotic Substances Produced by Aspergillus fumigatus. J. Bacteriol. 1944, 47, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.; Fujioka, S.; Nukina, M.; Kawano, T.; Shimada, A.; Kimura, Y. Fumiquinones A and B, nematicidal quinones produced by Aspergillus fumigatus. Biosci. Biotechnol. Biochem. 2007, 71, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.F.; Jenner, P.C. Distribution of trypacidin in cultures of Aspergillus fumigatus. Appl. Microbiol. 1968, 16, 1251–1252. [Google Scholar] [CrossRef]
- Balan, J.; Ebringer, L.; Němec, P. Trypacidin a new antiprotozoal antibiotic. Die Nat. 1964, 51, 227. [Google Scholar] [CrossRef]
- Van Middlesworth, F.; Dufresne, C.; Wincott, F.E.; Mosley, R.T.; Wilson, K.E. Determination of the relative and absolute stereochemistry of sphingofungins A, B, C and D. Tetrahedron Lett. 1992, 33, 297–300. [Google Scholar]
- Lindequist, U.; Lesnau, A.; Teuscher, E.; Pilgrim, H. The antiviral action of ergosterol peroxide. Die Pharm. 1989, 44, 579–580. [Google Scholar]
- Tomoda, H.; Kim, Y.K.; Nishida, H.; Masuma, R.; Omura, S. Pyripyropenes, novel inhibitors of acyl-CoA: Cholesterol acyltransferase produced by Aspergillus fumigatus I. Production, isolation, and biological properties. J. Antibiot. 1994, 47, 148–153. [Google Scholar] [CrossRef]
- Tomoda, H.; Ōmura, S. Pyripyropenes, highly potent inhibitors of acyl-CoA: Cholesterol acyltransferase produced by Aspergillus fumigatus. J. Antibiot. 1993, 46, 1168–1169. [Google Scholar]
- Bhatti, A.B.; Usman, M.; Kandi, V. Current Scenario of HIV/AIDS, Treatment Options, and Major Challenges with Compliance to Antiretroviral Therapy. Cureus 2016, 8, e515. [Google Scholar] [CrossRef]
- May, J.F. World Population Policies: Their Origin, Evolution, and Impact. Can. Stud. Popul. 2012, 39, 125. [Google Scholar]
- Roberts, D.; Matthews, G. Risk factors of malaria in children under the age of five years old in Uganda. Malar. J. 2016, 15, 246. [Google Scholar] [CrossRef] [Green Version]
- Afoakwah, C.; Deng, X.; Onur, I. Malaria infection among children under-five: The use of large-scale interventions in Ghana. BMC Public Health 2018, 18, 536. [Google Scholar] [CrossRef] [Green Version]
- Iskander, D. The Power of Parasites: Malaria as (un) Conscious Strategy; Springer Nature: Singapore, 2021. [Google Scholar]
- Crompton, D.W.; Savioli, L. Intestinal parasitic infections and urbanization. Bull. World Health Organ. 1993, 71, 1–7. [Google Scholar]
- Peniche, A.G.; Renslo, A.R.; Melby, P.C.; Travi, B.L. Antileishmanial Activity of Disulfiram and Thiuram Disulfide Analogs in an Ex Vivo Model System Is Selectively Enhanced by the Addition of Divalent Metal Ions. Antimicrob. Agents Chemother. 2015, 59, 6463–6470. [Google Scholar] [CrossRef] [Green Version]
- Watts, K.R.; Ratnam, J.; Ang, K.H.; Tenney, K.; Compton, J.E.; McKerrow, J.; Crews, P. Assessing the trypanocidal potential of natural and semi-synthetic diketopiperazines from two deep water marine-derived fungi. Bioorganic Med. Chem. 2010, 18, 2566–2574. [Google Scholar] [CrossRef] [Green Version]
- Tagboto, S.; Townson, S. Antiparasitic properties of medicinal plants and other naturally occurring products. Adv. Parasitol. 2001, 50, 199–295. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Medicinal Plants: A Source of Anti-Parasitic Secondary Metabolites. Molecules 2012, 17, 12771–12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simarro, P.P.; Cecchi, G.; Paone, M.; Franco, J.R.; Diarra, A.; Ruiz, A.J.; Fèvre, E.M.; Courtin, F.; Mattioli, R.C.; Jannin, J.G. The Atlas of human African trypanosomiasis: A contribution to global mapping of neglected tropical diseases. Int. J. Health Geogr. 2010, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An overview on target-based drug design against kinetoplastid protozoan infections: Human African trypanosomiasis, Chagas disease and leishmaniases. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Gou, X.; Jia, J.; Wei, J.; Zheng, M.; Ding, W.; Bi, H.; Wu, B.; Tang, J. New diketopiperazine alkaloid and polyketides from marine-derived fungus Penicillium sp. TW58-16 with antibacterial activity against Helicobacter pylori. Fitoterapia 2022, 156, 105095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, H.; Chen, B.; Dai, H.; Sun, J.; Han, J.; Liu, H. Discovery of Anti-MRSA Secondary Metabolites from a Marine-Derived Fungus Aspergillus fumigatus. Mar. Drugs 2022, 20, 302. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, R.; Yang, J.; Li, H.; Zhou, F. Bioactive Alkaloids of Aspergillus fumigatus, an Endophytic Fungus from Astragalus membranaceus. Chem. Nat. Compd. 2017, 53, 802–805. [Google Scholar] [CrossRef]
- Nakanishi, K.; Doi, M.; Usami, Y.; Amagata, T.; Minoura, K.; Tanaka, R.; Numata, A.; Yamada, T. Anthcolorins A–F, novel cytotoxic metabolites from a sea urchin-derived Aspergillus versicolor. Tetrahedron 2013, 69, 4617–4623. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R.; Rabie, C.J. Alkaloids from Aspergillus caespitosus. Phytochemistry 1981, 20, 538–539. [Google Scholar] [CrossRef]
- Yan, L.-H.; Li, X.-M.; Chi, L.-P.; Li, X.; Wang, B.-G. Six New Antimicrobial Metabolites from the Deep-Sea Sediment-Derived Fungus Aspergillus fumigatus SD-406. Mar. Drugs 2021, 20, 4. [Google Scholar] [CrossRef]
- Ma, Y.-M.; Liang, X.-A.; Zhang, H.-C.; Liu, R. Cytotoxic and Antibiotic Cyclic Pentapeptide from an Endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016, 64, 3789–3793. [Google Scholar] [CrossRef]
- Sajid, M.; Robertson, S.A.; Brinen, L.S.; McKerrow, J.H. Cruzain—The path from target validation to the clinic. In Cysteine Proteases of Pathogenic Organisms; Robinson, M.W., Dalton, J.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Egan, T.J.; Ncokazi, K.K. Quinoline antimalarials decrease the rate of β-hematin formation. J. Inorg. Biochem. 2005, 99, 1532–1539. [Google Scholar] [CrossRef]
- Wolber, G.; Langer, T. LigandScout: 3-D pharmacophores derived from protein-bound ligands and their use as virtual screening filters. J. Chem. Inf. Model. 2005, 45, 160–169. [Google Scholar] [CrossRef]
- Wilson, G.; Bryan, J.; Cranston, K.; Kitzes, J.; Nederbragt, L.; Teal, T.K. Good enough practices in scientific computing. PLoS Comput. Biol. 2017, 13, e1005510. [Google Scholar] [CrossRef] [Green Version]
- Ogunkunle, C.O.; Mustapha, K.; Oyedeji, S.; Fatoba, P.O. Assessment of metallic pollution status of surface water and aquatic macrophytes of earthen dams in Ilorin, north-central of Nigeria as indicators of environmental health. J. King Saud Univ.-Sci. 2016, 28, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Okoro, H.K.; Jimoh, H.A. Speciation and determination of priority metals in sediments of Oyun River, Ilorin, Kwara, Nigeria. Bull. Chem. Soc. Ethiop. 2016, 30, 199–208. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Adusumilli, R.; Mallick, P. Data conversion with ProteoWizard msConvert. In Proteomics; Humana Press: New York, NY, USA, 2017; pp. 339–368. [Google Scholar]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.S., Jr.; Kupchan, S.M. A convenient separation of alkaloid mixtures by partition chromatography, using an indicator in the stationary phase. J. Chromatogr. A 1962, 9, 71–80. [Google Scholar] [CrossRef]
- Mott, B.T.; Ferreira, R.S.; Simeonov, A.; Jadhav, A.; Ang, K.K.; Leister, W.; Shen, M.; Silveira, J.T.; Doyle, P.S.; Arkin, M.R.; et al. Identification and optimization of inhibitors of trypanosomal cysteine proteases: Cruzain, rhodesain, and TbCatB. J. Med. Chem. 2010, 53, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.R.; Banfield, M.J.; Barker, J.J.; Higham, C.W.; Moreton, K.M.; Turgut-Balik, D.; Brady, R.L.; Holbrook, J.J. The structure of lactate dehydrogenase from Plasmodium falciparum reveals a new target for anti-malarial design. Nat. Struct. Biol. 1996, 3. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2. 0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annang, F.; Pérez-Moreno, G.; García-Hernández, R.; Cordon-Obras, C.; Martín, J.; Tormo, J.R.; Rodríguez, L.; De Pedro, N.; Gómez-Pérez, V.; Valente, M.; et al. High-throughput screening platform for natural product–based drug discovery against 3 neglected tropical diseases: Human african trypanosomiasis, leishmaniasis, and chagas disease. J. Biomol. Screen. 2015, 20, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Annang, F.; Perez-Victoria, I.; Perez-Moreno, G.; Domingo, E.; Gonzalez, I.; Tormo, J.R.; Martin, J.; Ruiz-Perez, L.M.; Genilloud, O.; Gonzalez-Pacanowska, D.; et al. MDN-0185, an antiplasmodial polycyclic xanthone isolated from Micromonospora sp. CA-256353. J. Nat. Prod. 2018, 81, 1687–1691. [Google Scholar] [CrossRef]
- Ignatushchenko, M.V.; Riscoe, M.; Winter, R.W. Xanthones as antimalarial agents: Stage specificity. Am. J. Trop. Med. Hyg. 2000, 62, 77–81. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | [M+H]+ | tR (min) | MF | Compound | Source | Ref |
|---|---|---|---|---|---|---|
| 1 | 480.1595 | 4.5 | C27H33N3O5 | Fumitremorgen B | ISP2, ISP2-S | [26] |
| 2 | 496.2338 | 3.8 | C27H33N3O6 | Spirotryprostatin C | ISP2, ISP2-S | [27] |
| 3 | 528.5353 | 7.7 | C27H33N3O8 | Spirotryprostatin E | ISP2, ISP2-S | [28] |
| 4 | 524.3704 | 5.6 | C33H49NO4 | Anthcolorin E | ISP2, ISP2-S | [29] |
| 5 | 522.3404 | 3.3 | C33H47NO4 | Anthcolorin H | ISP2, ISP2-S | [29] |
| 6 | 478.5882 | 7.1 | C28H34N2O5 | Fumitremorgen B derivative | RICE | [30] |
| 7 | 431.4423 | 6.9 | C22H25NO8 | Pseurotin D | RICE | [31] |
| 8 | 381.4316 | 6.2 | C21H23N3O4 | Cyclotryprostatin C | RICE | [28] |
| 9 | 379.4151 | 6.4 | C21H21N3O4 | Cyclotryprostatin D | RICE | [32] |
| 10 | 511.6752 | 4.5 | C27H33N3O7 | Verruculogen | RICE | [32] |
| Compounds | Docking Score (-) (kcal mol−1) | Docking Score (-) (kcal mol−1) |
|---|---|---|
| (PDB: 3I06) | (PDB: 1LDG) | |
| Cruzain from Trypanosoma cruzi | l-lactate Dehydrogenase from Plasmodium falciparum | |
| 1 | 7.1 | 7.5 |
| 2 | 6.8 | 8.9 |
| Benznidazole (standard) a | 5.6 | - |
| Chloroquine (standard) b | - | 6.3 |
| Compounds | IC50 (μM) | |
|---|---|---|
| Antitrypanosomal Activity Trypanosoma cruzi C2C4 Strain | Antimalarial Activity Plasmodium falciparum 3D7 Strain | |
| 1 | 9.6 | 2.3 |
| 2 | >50 | >50 |
| Benznidazole [a] | 2.6 | NA |
| Chloroquine [b] | NA | 0.017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diyaolu, O.A.; Preet, G.; Fagbemi, A.A.; Annang, F.; Pérez-Moreno, G.; Bosch-Navarrete, C.; Adebisi, O.O.; Oluwabusola, E.T.; Milne, B.F.; Jaspars, M.; et al. Antiparasitic Activities of Compounds Isolated from Aspergillus fumigatus Strain Discovered in Northcentral Nigeria. Antibiotics 2023, 12, 109. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12010109
Diyaolu OA, Preet G, Fagbemi AA, Annang F, Pérez-Moreno G, Bosch-Navarrete C, Adebisi OO, Oluwabusola ET, Milne BF, Jaspars M, et al. Antiparasitic Activities of Compounds Isolated from Aspergillus fumigatus Strain Discovered in Northcentral Nigeria. Antibiotics. 2023; 12(1):109. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12010109
Chicago/Turabian StyleDiyaolu, Oluwatofunmilayo A., Gagan Preet, Adeshola A. Fagbemi, Frederick Annang, Guiomar Pérez-Moreno, Cristina Bosch-Navarrete, Olusoji O. Adebisi, Emmanuel T. Oluwabusola, Bruce F. Milne, Marcel Jaspars, and et al. 2023. "Antiparasitic Activities of Compounds Isolated from Aspergillus fumigatus Strain Discovered in Northcentral Nigeria" Antibiotics 12, no. 1: 109. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12010109