
Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens

 ,
, 
 ,
,  , and
, and
Abstract
:1. Introduction
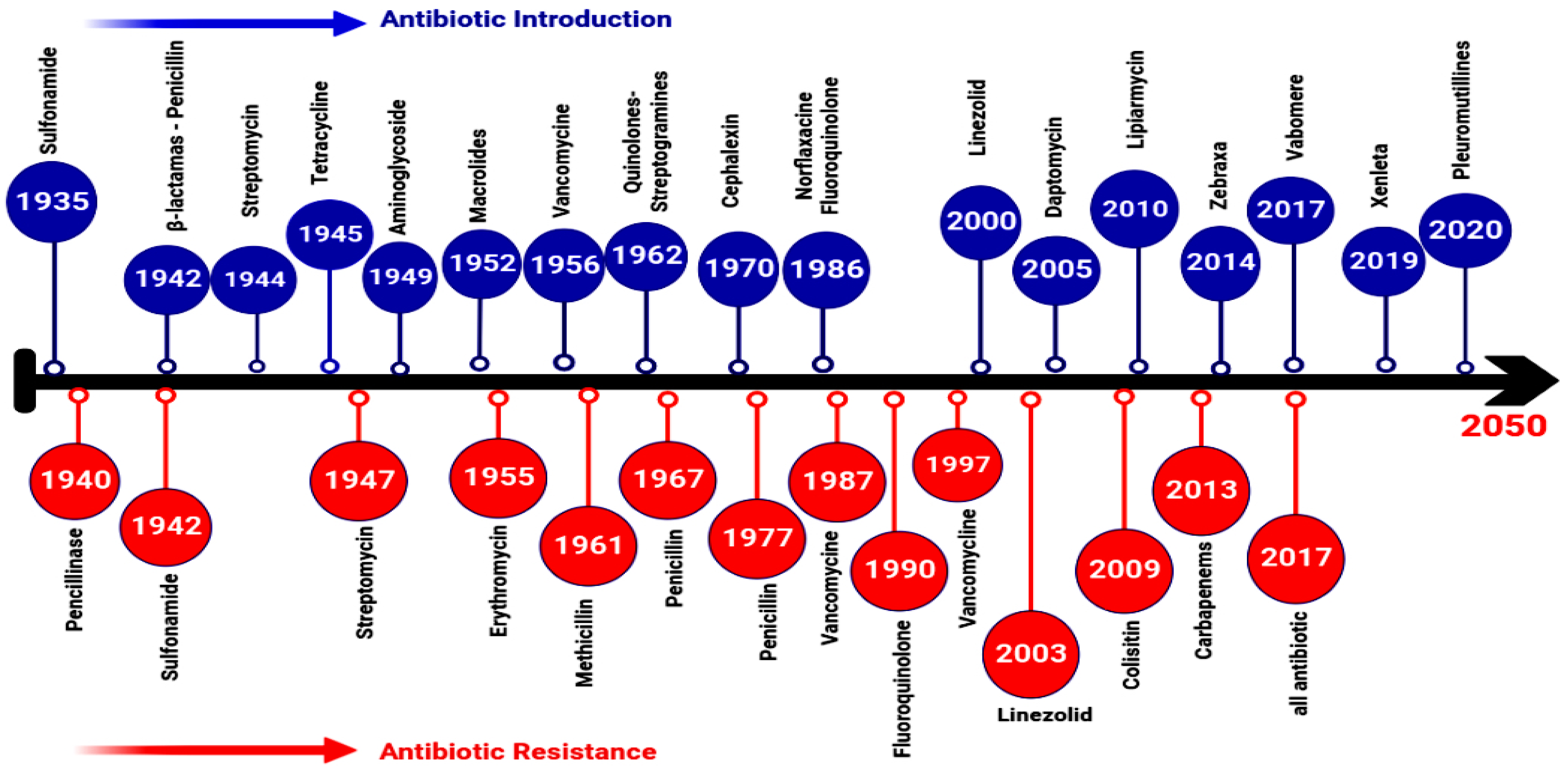
2. Evolution, Source, and Transmission of AMR
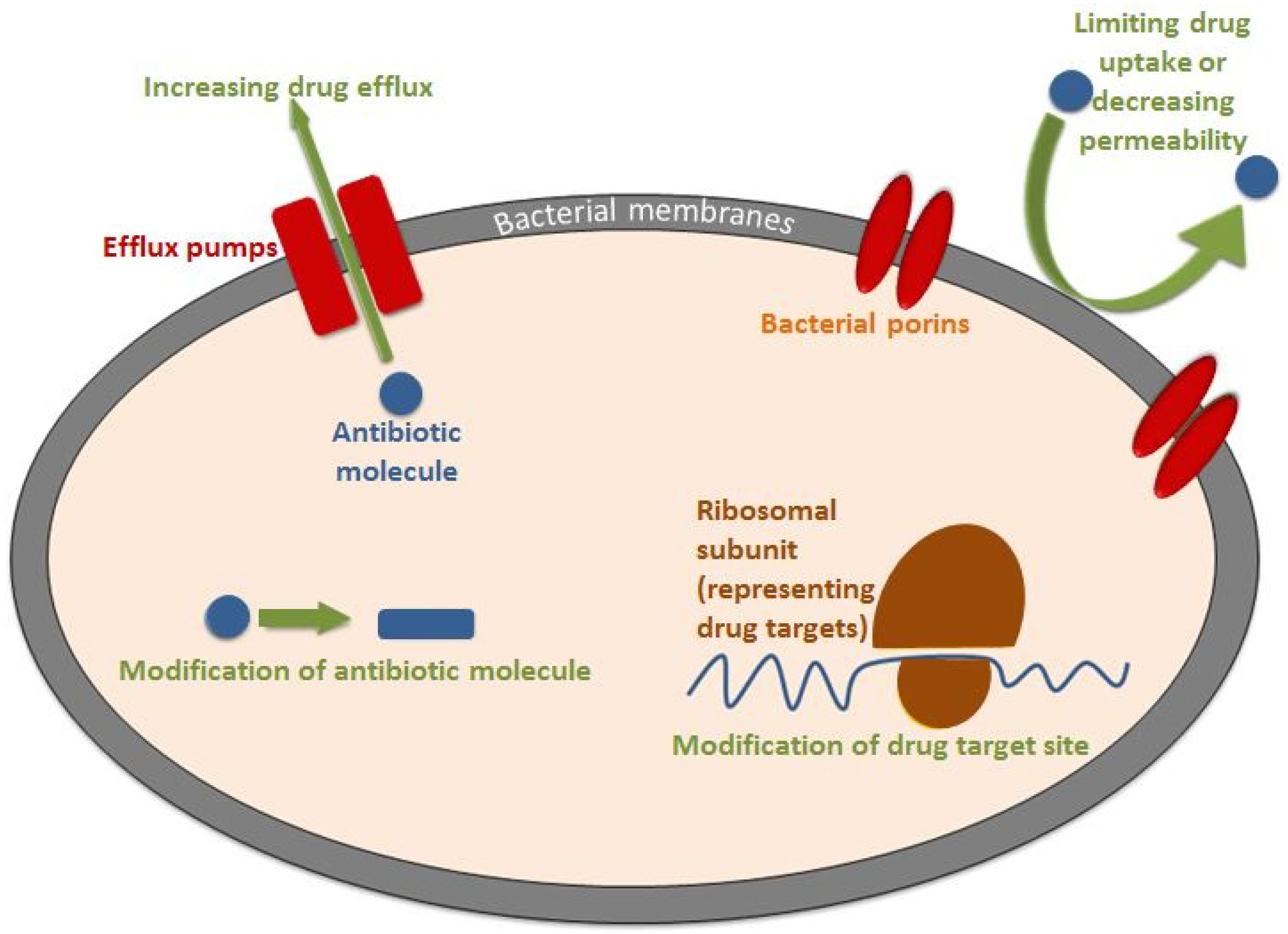
3. Mechanism of Acquiring AMR
3.1. Limiting Drug Uptake and Decreasing Permeability
3.1.1. Lipopolysaccharide (LPS) of Outer Bacterial Membrane
3.1.2. Bacterial Porins
3.1.3. Biofilm Formation
3.2. Enzymatic Destruction of Antibiotic Molecules
3.3. Drug Target Site Modification
3.4. Antibiotic-Specific Efflux Pumps
4. Novel Strategies to Combat AMR
4.1. Small Molecules (SMs)
4.1.1. Mechanisms of Actions of SMs
4.1.2. Limitations of SMs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Name of the SMs | Evaluation | Targets | References |
|---|---|---|---|---|
| M. tuberculosis | Benzimidazole and nitro-triazole | In vitro | Inhibit cell wall biosynthesis | [92] |
| Uropathogenic E. Coli | 120304 and 175472 | In vitro | TonB system | [93] |
| E. coli and P. aeruginosa | Nitrofurans | In vitro | Inhibit bacterial growth through reduction of the nitro group to an amine, followed by damage to bacterial DNA | [94] |
| P. aeruginosa and S. Typhimurium | Class 2,4-disubstituted-4H-[1,3,4]-thiadiazine-5-ones, Fluorothiazinon (FT) | In vitro, in mice | Suppress T3SS | [95,96] |
| Bacillus subtilis | Adamantane derivatives (T6102) | In vitro | Inhibit bacterial protein synthesis and bacterial growth | [97] |
| S. aureus and S. epidermidis | 3-methoxybenzamide derivatives (PC190723) | In vitro (CD-1 mouse hepatocytes), in mice | Disrupt FtsZ | [98] |
| S. aureus | ZY-214-4 (C19H11BrNO4) | In vitro | Suppress biofilm formation | [99] |
| Mycoplasma gallisepticum | SM4 and SM9 | In vitro, in chickens | Alter cell membrane conformation | [76] |
| M. bovis | Methanesulphonic acid, 3-[(2E)-3-(3,4-dihydroxyphenyl) prop-2-enoyloxy](1S,3R,4R,5R)- 1,4,5-trihydroxycyclohexane carboxylic acid, S-carboxymethyl-l-cysteine, l-aspartic acid, dihydrotachysterol, eriodictyol and (+)-a-tocopherol acid succinate) | In vitro | NI* | [100] |
| C. jejuni | Campynexin A | In vitro, in chickens | Inhibit flagellar expression | [101] |
| C. jejuni | Piperazine, aryl amine, piperidine, sulfonamide and pyridazinone molecules | In vitro | NI* | [75] |
| C. jejuni | TH-4 and TH-8 | In vitro, in chickens | Alter cell membrane integrity | [102] |
| S. Typhimurium | JD1 | In vitro, in mice | Inhibit bacterial growth by distorting cytoplasmic membranes through increasing fluidity and disrupting barrier function | [103] |
| S. Typhimurium | Imidazole and methoxybenzylamine | In vitro, Galleria mellonella larvae, in chickens | Alter cell membrane integrity | [104] |
| Avian pathogenic E. coli (APEC) | QSI-5 and GI-7 | In vitro, Galleria mellonella larvae, in chickens | Inhibit quorum-sensing autoinducer-2 and outer membrane proteins | [105,106,107,108] |
| E. faecium | 6-indolyl compounds | In vitro | NI* | [109] |
| Clostridium difficle R20291 | 2-aminoimidazole (2-AI) | In vitro | NI* | [110] |
| Chlamydia | INPs (Innate Pharmaceuticals AB) | Epithelial cells | Supress Type III secretion | [111] |
| Clostridium botulinum | In vitro, In mice | Inhibit neurotoxin serotype A | [112] | |
| L. monocytogenes | Pimozide (antipsychotic drug) | In vitro: murine bone marrow-derived macrophages (BMM) | Decrease the vacuole escape and cell-to-cell spread of L. monocytogenes | [113] |
| L. monocytogenes | SM-3, 5, 7 | In vitro: on catfish fillets | Block the LapB gene, that encodes cell wall surface anchor protein | [114] |
| S. aureus, S. epidermidis, S. pyogenes, S. pneumoniae and Bacillus cereus | F19 and F12 | In vitro on human THP-1 monocytes and mouse macrophage cell line - In mice | Host cell lysis | [115] |
4.2. Quorum-Sensing/Antivirulence Inhibitors
4.2.1. Mechanisms of Action of QSIs
4.2.2. Limitations of QSIs
| Compound Name | Source | Target Pathogen | Mechanism of Inhibition | References |
|---|---|---|---|---|
| C1-C10 | Synthetic | APEC O78 | Inhibit quorum sensing via inhibiting AI-2 production, genes associated with biofilm formation, such as the hha gene, and genes associated with bacterial cell morphology, motility, and division. | [107] |
| Savirin | Synthetic | S. aureus | Inhibits the signaling cascade of bacteria and biofilm formation by targeting AgrA to disrupt agr operon-mediated QS. | [141] |
| N-phenyl-4-(3-phenylthioureido) benzene sulfonamide | Synthetic | E. coli [EHEC] | Inhibits biofilm formation and virulence factors by modifying the AI-3 receptor (QseC). | [142] |
| Anti-autoinducer monoclonal antibody AP4-24H11 | Synthetic | S. aureus [RN4850] | Inhibits the QS signaling molecule autoinducing peptide (AIP)-4 by targeting AgrA, resulting in QS inhibition and biofilm formation. | [143] |
| Limonene nanoemulsion | Synthetic | E. coli [EHEC] | Reduces AI-2 synthesis; inhibits the production of E. coli flagellum by inhibiting QseB and the promoter region of flhDC binding that encodes bacterial motility | [144] |
| N-phenyl-4-phenylaminothioxomen hyl amino-benzenesulfonamide | Synthetic | E. coli [EHEC] S. Typhimurium | Inhibits the histidine kinase QseC and results in a decrease in the expression of virulence factors. | [145] |
| Thiophene inhibitor (TF101) | Synthetic | E. coli (EPEC) | Inhibit the expression of the lsrB gene which encodes the AI-2 receptor, and interferes with fimH, which encodes virulence factors and inhibits biofilm formation. | [146] |
| Grape seed extract | Grape seed extract | E. coli (STEC), E. coli (VTEC), E. coli (EAEC) | Reduces the synthesis of AI and its activity by blocking AI synthase activity. Inhibits the production of E. coli flagellum by inhibiting QseB and the promoter region of flhDC binding that encodes bacterial motility and inhibit Shiga toxin production. | [128] |
| Thymol-carvacrol-chemotype (I and II) oils | Lippia origanoides Thymus vulga0ris oil | E. coli [O157:H7] E. coli [O33] | Inhibits the synthesis of AI-3 and prevents the formation of biofilm. | [147] |
| furocoumarin | Grapefruit juice | S. Typhimurium E. coli [O157:H7] | Inhibits the activity of AI-2, interferes with the activity of AI-1 molecules (AHLs), and inhibits biofilm formation. | [148] |
| Broccoli extracts | Basil, oregano, thyme, rosemary, ginger, and turmeric | E. coli [EHEC] | Inhibits the activity of AI-2 synthase and inhibits synthesis of AI-2. Affects E. coli mobility and inhibits production of virulence factors. | [149] |
| Acetic acid, citric acid, and lactic acid | Vinegar, Lemon, fermented soy products, yogurt | S. Typhimurium E. coli [O157:H7] | Inhibit the producing of the signaling molecules AI-2 by inhibiting AI-2 synthase. They also inhibit the activity of biofilm formation. | [150] |
| Star anise (Illicium verum Hook. f.) | Chinese fruit evergreen tree Illicium verum | S. Typhimurium | Interferes with promoter region flhDC operon which regulates the mobility. Interferes with the signal receptors lux, rhl, and las systems and inhibits biofilm formation. | [151] |
| Chitosan | Shells of crustaceans | E. coli [UPEC] | Reduces E. coli mobility by inhibiting QseB binding to the promoter region of flhDC. Inhibits AI-2 production and biofilm formation. | [152] |
| (Z)-4-Bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one | Synthetic | E. coli [RP437] | Reduces the activity of AI-2 by reducing the activity of AI-2 synthase. | [153] |
| Punicalagin | Pomegranate rind | S. Typhimurium [SL1344] | Decreases the expression of the genes fliA, fliY, fljB, flhC, and fimD encoding the swimming and swarming motility of Salmonella and represses the expression of sdiA and srgE QS-related genes. | [154] |
| 2,3-methyl-N-(2′-phenylethyl)-butyramide | Halobacillus salinus | E. coli [JB525] | Inhibits biofilm formation and decreases the expression of virulence factors by competing with signaling molecules (AHL) for receptor binding. | [155,156] |
| N-(2′-phenylethyl)-isobutyramide | Halobacillus salinus | E. coli [JB525] | Competes with signaling molecules (AHL) for receptor binding and inhibits bacterial QS resulting in biofilm formation. | [155,156] |
| Cyclo(L-Pro-L-Val) | Haloterrigena hispanica | E. coli [JB525] | Inhibits biofilm formation by interacting with signaling molecules (AHL). | [157] |
| Diketopiperazines (DKPs): Cyclo(L-Pro-L-Phe), Cyclo(L-Pro-L-Leu), Cyclo(L-Pro-L-isoLeu), Cyclo(L-Pro-D-Phe) | Marinobacter sp. | E. coli [pSB401] | Inhibits bacterial biofilm formation by inhibiting the production of AHL signaling molecules. | [158] |
| Kojic acid | Altenaria sp., from marine green algae Ulva pertusa | E. coli [pSB401] | Interferes with N-hexanoyl-L-homoserine lactone (C6-HSL) and with LuxR reporters. | [159] |
| O-prenylated flavonoid buchapine and 3-(3-methyl-2-butenyl)-4-[(3-methyl-2-butenyl) oxy]-2(1H)-quinolinone | Melicope lunu-ankenda (leaves extract) | E. coli [pSB401] | Inhibits biofilm formation and decreases violacein production, motility, and bioluminescence production by downregulating the expression of lecA and lux genes. | [160,161] |
| Sesquiterpenes, monoterpenes, hydrocarbon, and phenolic compounds. Eugenyl acetate, eugenol, and β-caryophyllene | Syzygium aromaticum (bud) | E. coli [pSB1075] | Targets lecA and lux genes resulting in the inhibition of QS-regulated phenotypes and violacein factor production, which are considered secondary metabolites responsible for growth and propagation and are a useful indicator of QS systems in bacteria. | [162] |
| Fructose-furoic acid | Aloe africana (plant extract) | E. coli [UPEC] | Represses the expression of biofilm phenotypic characters by competing with quorum regulator (SdiA) native ligand C8HSL. | [163] |
| Cembranoids | Pseudoplexaura flagellosa and Eunicea knighti | E. coli [pSB403] S. aureus | Inhibits biofilm formation by interacting with LuxR receptors. | [164] |
| Brominated alkaloids compounds | Flustra foliacea | E. coli [pSB403] | Inhibits biofilm formation by targeting CepR and LuxR and interferes with N-acyl-homoserine lactone. | [165] |
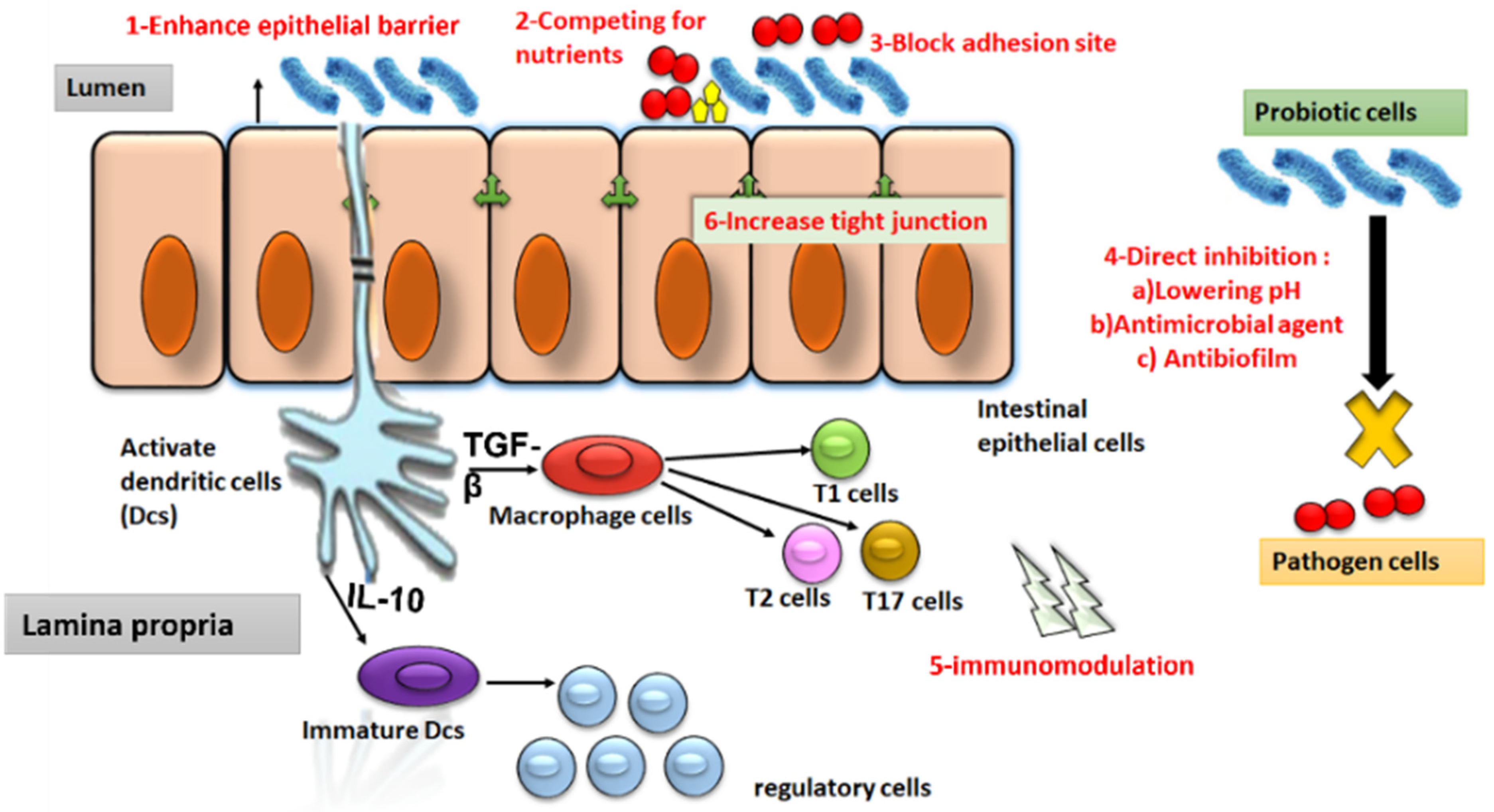
4.3. Probiotics
- Viable and active probiotics
- 2.
- Viable and inactive probiotics
- 3.
- Dead/nonviable probiotics (postbiotics/parabiotics)
- 4.
- Next-generation and genetically modified probiotics
4.3.1. Mechanisms of Action of Probiotics
4.3.2. Limitations of Probiotics
| Probiotic | Target Pathogen | Additional Benefits | Monitoring | References |
|---|---|---|---|---|
| Nissle E. coli 1917 (EcN) | C. jejuni | In vitro: enhance tight junction functions and modulate the innate immune response on HT-29 cells. In chickens: reduce C. Jejuni colonization in the cecum up to 2.5 logs; enhance the immune response and intestinal morphology of the treated chickens without showing adverse effect on the gut microbe. | In vitro (HT-29 cell line) In chickens | [172,173,174,214] |
| L. plantarum | L. monocytogenes, S. Enteritidis, E. coli O157:H7 and Staphylococcus | Attach to epithelial cells, stimulate the production of IL-10 in the colon, and enhance the induction of dopamine and serotonin. | In vitro & in mice | [188,215] |
| L. paracasei & L. rhamnosus | E. coli V517, S. Enteritidis OMS-Ca, S. aureus 76 and L. monocytogenes ATCC 15313 | Boost mineral bioavailability in food products, reduce serum parathyroid hormone via synthesis of short-chain fatty acids, enhance mineral solubilization and absorption, production of phytase, and hydrolyze glycoside linkages of estrogenic food products. | In vitro | [191,216,217] |
| L. helveticus | L. monocytogenes ATCC 19115, S. Typhimurium ATCC 14028, S. aureus ATCC 25923, and E. coli O157:H7 ATCC 43889 | Stop GIT infections, improve protection against pathogens, enhance the immune system of the host, and makeup the composition GIT microbiota. | In vitro | [218,219] |
| L. reuteri | E. coli ATCC25922, S. typhi NCDC113, L. monocytogenes ATCC53135, and E. faecalis NCDC115. | Reduce pro-inflammatory cytokines production, promote regulatory T cells, strengthen the intestinal barrier, and decrease microbial translocation from the gut lumen to the tissues. | In vivo | [220,221] |
| L. acidophilus | S. aureus, P. aeruginosa, L. monocytogenes, V. parahaemolyticus, V. cholerae, H. pylori, Klebsiella, Salmonella, Shigella, Bacillus, Clostridium, Mucor, Aspergillus, Fusarium, Trichoderma and Candida spp. | Production of lactacins B and F, acidophilin, acidocin, acidophilucin, and acidophilicin. | In vitro | [222] |
| L. rhamnosus GG and B. lactis Bb12 | APEC | Reduce the number of colonized APEC in chicken cecum with modulation of the gut microbiota. | In vitro In chickens | [223] |
| S. lactis and L. delbrueckii subsp. Bulgaricus | E. coli ATCC25922 and S. aureus ATCC25923 | Inhibit proliferation via production of acid metabolites. | In vitro & in vivo | [224] |
| B. animalis AHC7 | S. Typhimurium | Mediate weakness of activation of NF-κB that includes recognition of the pathogen by dendritic cells and production of T cells. | In humans | [225] |
| B. adolescentis and B. pseudocatenulatum, and B. longum | Vancomycin-resistant S. aureus and Enterococcus, Propionibacterium acnes, S. aureus, and S. Epidermidis | Reduce pathogen growth and cell adhesion. | In vitro | [226] |
| B. bifidum and B.m infantis | S. enterica serotype Enteritidis | Reduce pathogen growth via production of acids, hydrogen proxide, and bacteriocins. | In vitro | [227] |
| B. lactis | S. Typhimurium | Stimulate transient pro-inflammatory host responses in the epithelial cells of the intestine. | In vivo (rats) | [228] |
| Propionibacterium freudenreichii | Multidrug-resistant S. Heidelberg | Anti-inflammatory effect. | In vitro (HT-29 cell line) | [229,230] |
| Pediococcus acidilactici Kp1 | L. monocytgenes, S. enterica, Shigella sonnei, Klebsiella oxytoca, Enterobacter cloaca and S. pyogenes. | Hender the adherence of pathogens to the intestinal mucosa by forming a barrier via auto-aggregation; production of bacteriocin-like inhibitory substances. | In vitro | [231] |
| Leuconostoc mesenteroides | L. innocua, L. ivanovii, or S. aureus | Production of bacteriocin, which inhibits the growth of pathogens, and lowering the medium pH. | In vivo (mice) | [189,232] |
| E. faecium NCIMB 11181 | C. perfringens | Ameliorate necrotic enteritis and reduce intestinal barrier injury. | In chickens) | [233] |
| S. salivarius K12 | S. mutans and S. hominis | Antibiofilm of Schaalia odontolytica P10 and Enterobacter cloacae. | In vitro | [234] |
| S. thermophilus SMQ-301 | S. aureus, E. coli, and Gardnerella vaginalis | Potential candidate for novel biotherapeutic interventions against inflammation caused in septic mice. | In vitro, in vivo | [235,236] |
| B. coagulans subtilis, B. laterosporus | E. coli, P. aeruginosa, K. pneumoniae, B. subtilis, S. aureu, and Candida albicans | Stimulate human immune cells and change the induction of anti-inflammatory cytokines and chemokines. | In vitro (cell lines) | [237] |
| Saccharomyces boulardii | S. aureus, E. coli, Klebsiella oxytoca, Yersinia enterocolitica, C. perfringens, C. difficile, Salmonella sp., Shigella sp., Candida albicans and Entamoeba hystolitica | Affect the epithelial reconstitution; anti-secretory, anti-inflammatory, and immunomodulating effects. | In vivo (Lymphocyte-transferred SCID mice) | [238,239,240] |
| C. butyricum (CBM 588) | E. coli [EHEC] O157:H7 | Inhibit growth by limiting the adhesion of pathogen to epithelial cells and the production of butyric acid. | In vivo (mice) | [241,242] |
| L. salivarius, L. johnsonii, L. reuteri, L. crispatus, and L. gasseri | C. jejuni 81-176 | Inhibit the quorum-sensing signals of C. jejuni. Reduce the expression of C. jejuni virulence-related genes, including genes responsible for motility (flaA, flaB, and flhA), invasion (ciaB), and AI-2 production (luxS). Enhance the phagocytic activity of macrophages. Increase the expression of cytokines and co-stimulatory molecules in macrophages. | In vitro | [243] |
| Microbial consortia (Aviguard and CEL) | C. jejuni 81-176 | Enhance the intestinal mucosa via the modulation of gut microbiome composition by increasing the relative abundance of Bacteroidaceae and Rikenellaceae | In vivo (chicken) | [244] |
| L. johnsonii, Ligilactobacillus salivarius, Limosilactobacillus reuteri, and L. crispatus | C. perferingens | Induce significant alterations in cytokine gene expression in the intestine. Modify the gut microbiome composition. Improve intestinal morphology. | In vivo (chicken) | [245] |
4.4. Prebiotics
4.4.1. Mechanisms of Action of Prebiotics
4.4.2. Limitations of Prebiotics
4.5. Antimicrobial Peptides (AMPs)
4.5.1. Mechanisms of Action of AMPs
4.5.2. Limitation of AMPs
4.6. Bacteriophages
4.6.1. Mechanisms of Action of Bacteriophages
4.6.2. Limitations of Phage Therapy
| Phage | Target Bacteria | PFU | Application | Reference |
|---|---|---|---|---|
| Cocktail of 12 natural virulent bacteriophages | P. aeruginosa | 106 | In vivo in human | [324] |
| coliphage PhiX174 | S. aureus | NI | Patients with S. aureus bacteremia | [325] |
| Phage cocktail DS-6A, GR-21/T, My-327 | Mycobacterium abscessus | 109 | A cystic fibrosis patient | [326] |
| cocktail 1 (P. aeruginosa 24, P. aeruginosa 25, and P. aeruginosa 7) | P. aeruginosa | 6.2 × 1010 | Mice with chronic bacterial lung infections | [327] |
| IME-AB2 | A. baumannii | 62 PFU/cell | Reduce lung inflammation in mice | [328] |
| Pyo phage phage cocktail from the Eliava Institute | S. aureus, E. coli, Streptococcus, P. aeruginosa, or Proteus mirabilis | 107–109 | Patients with urinary tract infections | [329] |
| T4-like coliphage cocktail | E. coli | 3.6 × 108 | Diarrhea infected children | [330] |
| WPP-201 phage coctail | P. aeruginosa, S. aureus, and E. coli | 8 × 107 | Leg ulcer patients | [331] |
| P. aeruginosa phages 14/1 (Myoviridae) and PNM (Podoviridae) and S. aureus phage ISP (Myoviridae), | P. aeruginosa and S. aureus | 109 | Colonized burn wounds | [332] |
| PP01 phage, | E. coli O157: H7 | 105 | In vitro | [333] |
| PlySs2 and PlySs9 | S. uberis | NI | In vitro (bovine mastitis) | [334] |
| PlySs2 | S. equi, S. agalactiae, S. dysgalactiae, S. pyogenes, S. sanguinis, S. pneumoniae and group E streptococci | NI | In vitro and in vivo (mice) | [335] |
| Φ7-izsam and Φ16-izsam | C. jejuni | 107 | In chickens | [336] |
| Phage cocktail e11/2, e4/1c, pp01 | E. coli O157:H7 | ND | Meat surface | [337] |
| Phage Cj6 | C. jejuni | 5 × 108 | Raw and cooked beef | [338] |
| Phage Φ2 | C. jejuni | 107 | Chicken skin | [339] |
| Salmonella phage (P7) | Salmonella | 5 × 108 | Raw and cooked beef | [338] |
| phage SJ2 | S. Enteritidis | 104 | Cheddar cheese made from raw and pasteurized milk | [340] |
| phage A511 | L. monocytogenes | 5.2 × 10⁷ | Red smear cheese | [341] |
| Cocktail of the two lytic phages | S. aureus | 106 | Fresh and hard cheese type | [342] |
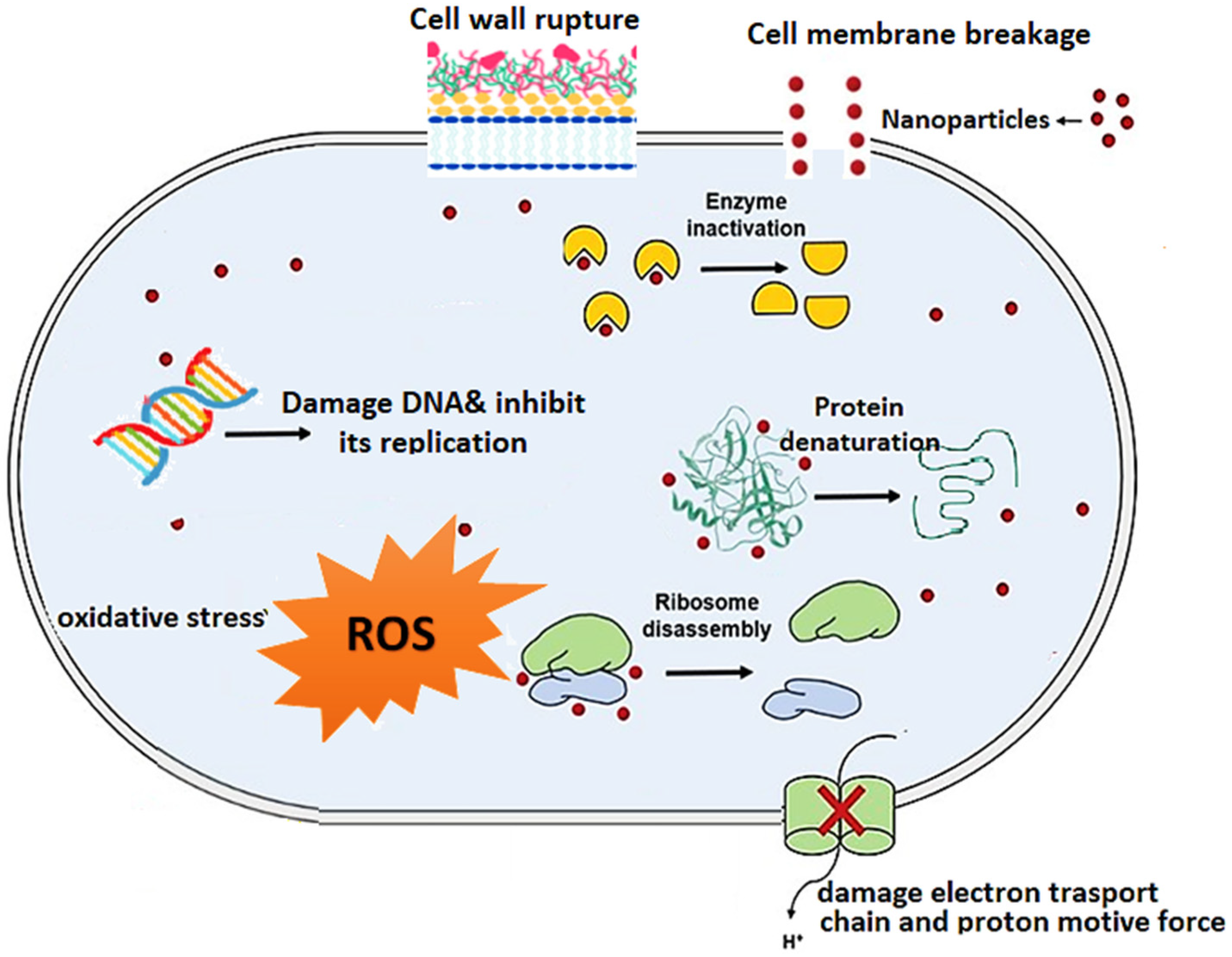
4.7. Nanoparticles (NPs)
4.7.1. Mechanisms of Action of NPs
4.7.2. Limitations of NPs
| NPs | Particle Size | Target Bacteria | Mechanism of Action | Reference |
|---|---|---|---|---|
| Silver (Ag) | 1–100 nm | S. epidermidis, MRSA, vancomycin-resistant Enterococcus (VRE), extended-spectrum beta lactamase (ESBL)-producing organisms, MDR E. coli, P. aeruginosa, K. pneumoniae, carbapenem and polymyxin B-resistant A. baumannii, and carbapenem resistant P. aeruginosa, E. coli | Generate reactive oxygen species (ROS), stopping cytochrome chains, membrane damage, dissipation of proton gradients, destabilization of RNA and DNA | [343,351,352,363] |
| Gold (Au) | 1–100 nm | MRSA | Damage membranes and respiratory chains, inhibit ATPase activity, decrease the binding between tRNA and ribosomes and formation of pores in the cell wall | [344,351,352] |
| Copper (Cu) | 2–350 nm | MDR E. coli, A. baumannii | Dissipation of cell membranes, generation of ROS, lipid peroxidation, protein oxidation, and DNA degradation | [343,364] |
| Silica (Si) | 20–400 nm | MRSA | Generation of ROS and lysis of cell walls | [351,352] |
| Aluminum (Al) | 10–100 nm | E. coli | Generation of ROS and lysis of cell walls | [344] |
| Iron oxide NP | 1–100 nm | MDR E. coli, MRSA, K. pneumoniae, | ROS-generated oxidative stress: superoxide radicals (O−2), hydroxyl radicals (OH−), hydrogen peroxide (H2O2) | [351] |
| Titanium dioxide (TiO2) | 30–45 nm | E. coli, P. aeruginosa, S. aureus, E. Faecium | ROS generation and adsorption to the cell surface | [344] |
| Zinc oxide (ZnO) | 10–100 nm | Enterobacter aerogenes, E. coli, K. oxytoca, K. pneumoniae, MRSA, K. Pneumoniae, ESBL-producing E. coli | Generation of ROS, disruption of membranes, adsorption to cell surface, and damage to lipids and proteins | [365] |
| Magnesium oxide (MgO) | 15–100 nm | S. aureus, E. coli | ROS generation, lipid peroxidation | [343] |
4.8. Organic Acids (OAs)
4.8.1. Mechanisms of Action of OAs
4.8.2. Limitations of OAs
| Organic Acid (pKa1) | Chemical Structure | Main Microbial Producers | Active against | References |
|---|---|---|---|---|
| Acetic acid (4.76) | C2H4O2 | C. formicoaceticum, Acetobacter, Gluconobacter, | L. monocytogenes, S. Typhimurium and E. coli | [367] |
| Adipic acid (4.41) | C6H10O4 | E. coli | Alternaria solani, Botrytis cinerea, Phytophthora capsici, and P. citrophthora | [386] |
| Butyric acid (4.82) | C4H8O2 | C. butyricum, Butyrivibrio sp., Eubacterium sp., Fusobacterium, Megasphera sp., Sarcina sp. | S. Enteritidis, C. perfringens, E. faecalis, and S. pneumoniae | [387,388] |
| Caprylic acid (4.89) | C8H16O2 | Mixculture from brewery wastewater | Vibrio parahaemolyticus & Dermatophilus congolensis | [389] |
| Citric acid (3.13) | C6H8O7 | Aspergillus ficum, Acremonium, Bacillus, Bostrytis, Candida, Aschochyta, Eupenicillium, Debaromyces, Hansenula, Trichoderma, Mucor, Pichia, Saccharomyces, Talaromyces, Penicillium, Torulopsis, Yarrowia, and Zygosaccharomyces | Yersinia enterocolitica Shigella dysenteriae E. coli O157:H7 | [390,391] |
| Fumaric acid (3.02) | C4H4O4 | Rhizopus arrhizus | Talaromyces flavus | [392] |
| Lactic acid (3.86) | C3H6O3 | Rhizopus oryzae, Aspergillus, Bacillus, Carnobacterium, Enterococcus, Escherichia, Lactobacillus, Lactococcus, Rhizopus, Saccharomyces | B. coagulans, L. monocytogenes | [393] |
| Malic acid (3.40) | C4H6O5 | Ustilago trichophora, E. coli, Saccharomyces, Aspergillus sp. and Zygosaccharomyces Aureobasidium pullulans | L. monocytogenes, E. coli O157:H7, S. Enteritidis and S. gaminara, | [394] |
| Phenyllactic acid (4.31) | C9H10O3 | B. coagulans, Lactobacillus, Enterococcus, Leuconostoc, and Weissella, Leuconostoc, L. plantarum 1081, L. acidophilus 1063, L. paracasei 1501 | L. monocytogenes Aspergillus spp. Penicillium spp. | [10,142] |
| Propionic acid (4.87) | C3H6O2 | Propionibacterium acidipropionici | L. plantarum, Sarcina lutea, S. ellipsoideus, Proteus vulgaris, S. aureus, and Torula spp. E. coli K12 and Salmonella | [154] |
| Succinic acid (4.21) | C4H6O4 | Yarrowia lipolytica, Anaerobiospirillum succiniciproducens, Mannheimia succiniciproducens, and Actinobacillus succinogenes | S. Typhimurium, E. coli, B. subtilis, and S. suis | [395,396] |
| Tartaric acid (2.98) | C4H6O6 | Gluconobacter suboxydans | L. monocytogenes, E. coli O157:H7 and S. gaminara | [397] |
| Valeric acid (4.82) | C5H10O2 | Megasphaera elsdenii | C. jejuni | [398] |
4.9. Essential Oils (EOs)
4.9.1. Mechanisms of Action of EOs
4.9.2. Limitations of EO Applications
4.10. Fecal Microbial Transplant (FMT)
4.10.1. Mechanisms of Action of FMT
4.10.2. Limitations of FMT
4.11. Vaccines
4.11.1. Mechanisms of Action of Vaccines
4.11.2. Limitations of Vaccines
4.12. Antibodies
4.12.1. Mechanisms of Action of mAbs
4.12.2. Limitations of Abs
4.13. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 31 October 2022).
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; El-Adawy, H.; Abdelwhab, E.M. A Comprehensive Review of Common Bacterial, Parasitic and Viral Zoonoses at the Human-Animal Interface in Egypt. Pathogens 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailu, W.; Helmy, Y.A.; Carney-Knisely, G.; Kauffman, M.; Fraga, D.; Rajashekara, G. Prevalence and Antimicrobial Resistance Profiles of Foodborne Pathogens Isolated from Dairy Cattle and Poultry Manure Amended Farms in Northeastern Ohio, the United States. Antibiotics 2021, 10, 1450. [Google Scholar] [CrossRef]
- Deblais, L.; Kathayat, D.; Helmy, Y.A.; Closs, G.; Rajashekara, G. Translating ‘big data’: Better understanding of host-pathogen interactions to control bacterial foodborne pathogens in poultry. Anim. Health Res. Rev. 2020, 21, 15–35. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Van Boeckel, T.P.; Glennon, E.E.; Chen, D.; Gilbert, M.; Robinson, T.P.; Grenfell, B.T.; Levin, S.A.; Bonhoeffer, S.; Laxminarayan, R. Reducing antimicrobial use in food animals. Science 2017, 357, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [Green Version]
- FDA. Summary Report on Antimicrobials Sold or Distributed for Use in Food-Producing Animals; Department of Health and Human Services: Washington, DC, USA, 2011.
- Kassem, I.I.; Kehinde, O.O.; Helmy, Y.A.; Kumar, A.; Chandrashekhar, K.; Pina-Mimbela, R.; Rajashekara, G. Campylobacter in poultry: The conundrums of highly adaptable and ubiquitous foodborne pathogens. In Foodborne Diseases: Case Studies of Outbreaks in the Agri-Food Industries; Soon, J.M., Manning, L., Wallace, C.A., Eds.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Kassem, I.; Helmy, Y.A.; Kashoma, I.P.; Rajashekara, G. The emergence of antibiotic resistance on poultry farms. In Achieving Sustainable Production of Poultry Meat: Safety, Quality and Sustainability; Ricke, S., Ed.; Burleigh Dodds Science Publishing: Sawston, UK, 2016; Volume 1, ISBN 978-1-78676-064-7. [Google Scholar]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Algammal, A.M.; Enany, M.E.; El-Tarabili, R.M.; Ghobashy, M.O.; Helmy, Y.A. Prevalence, antimicrobial resistance profiles, virulence and enterotoxins-determinant genes of MRSA isolated from subclinical bovine mastitis in Egypt. Pathogens 2020, 9, 362. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. 2016. Available online: http://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 31 October 2022).
- Scharff, R.L. Food Attribution and Economic Cost Estimates for Meat- and Poultry-Related Illnesses. J. Food Prot. 2020, 83, 959–967. [Google Scholar] [CrossRef] [PubMed]
- CDC. Burden of Foodborne Illness: Findings; Centers for Disease Control and Prevention, United States Department of Health and Human Services: Atlanta, GA, USA, 2018. Available online: https://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 31 October 2022).
- WHO. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 31 October 2022).
- NIAID. NIAID Emerging Infectious Diseases/Pathogens. Available online: https://www.niaid.nih.gov/research/emerging-infectious-diseases-pathogens (accessed on 31 October 2022).
- Truman, A. Antibiotics: Past, present and future Matthew I Hutchings, Andrew W Truman 2 and Barrie Wilkinson 2. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar]
- Sköld, O. Sulfonamide resistance: Mechanisms and trends. Drug Resist. Updates 2000, 3, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Pancu, D.F.; Scurtu, A.; Macasoi, I.G.; Marti, D.; Mioc, M.; Soica, C.; Coricovac, D.; Horhat, D.; Poenaru, M.; Dehelean, C. Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening. Antibiotics 2021, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Hussar, D.A. New Drugs 2019, part 4. Nursing2020 2019, 49, 34–43. [Google Scholar] [CrossRef]
- Andrei, S.; Droc, G.; Stefan, G. FDA approved antibacterial drugs: 2018–2019. Discoveries 2019, 7, e102. [Google Scholar] [CrossRef]
- Voulgaris, G.L.; Voulgari, M.L.; Falagas, M.E. Developments on antibiotics for multidrug resistant bacterial Gram-negative infections. Expert Rev. Anti-Infect. Ther. 2019, 17, 387–401. [Google Scholar] [CrossRef]
- Saxena, D.; Kaul, G.; Dasgupta, A.; Chopra, S. Levonadifloxacin arginine salt to treat acute bacterial skin and skin structure infection due to S. aureus including MRSA. Drugs Today 2020, 56, 583–598. [Google Scholar] [CrossRef]
- Stancil, S.L.; Mirzayev, F.; Abdel-Rahman, S.M. Profiling Pretomanid as a Therapeutic Option for TB Infection: Evidence to Date. Drug Des. Dev. Ther. 2021, 15, 2815. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J. Clin. Med. 2020, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Hedman, H.D.; Krawczyk, E.; Helmy, Y.A.; Zhang, L.; Varga, C. Host Diversity and Potential Transmission Pathways of SARS-CoV-2 at the Human-Animal Interface. Pathogens 2021, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Ilić, T.; Pantelić, I.; Savić, S. The implications of regulatory framework for topical semisolid drug products: From critical quality and performance attributes towards establishing bioequivalence. Pharmaceutics 2021, 13, 710. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report of the Meeting to Review the Paediatric Antituberculosis Drug Optimization Priority List; WHO: Geneva, Switzerland, 2021.
- Peterson, E.; Kaur, P. Antibiotic resistance mechanisms in bacteria: Relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.J.; McEwen, S.A. One health—Its importance in helping to better control antimicrobial resistance. Trop. Med. Infect. Dis. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Alduhaidhawi, A.H.M.; AlHuchaimi, S.N.; Al-Mayah, T.A.; Al-Ouqaili, M.T.; Alkafaas, S.S.; Muthupandian, S.; Saki, M. Prevalence of CRISPR-Cas Systems and Their Possible Association with Antibiotic Resistance in Enterococcus faecalis and Enterococcus faecium Collected from Hospital Wastewater. Infect. Drug Resist. 2022, 15, 1143. [Google Scholar] [CrossRef]
- Goldman, E. Antibiotic abuse in animal agriculture: Exacerbating drug resistance in human pathogens. Hum. Ecol. Risk Assess. 2004, 10, 121–134. [Google Scholar] [CrossRef]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and disease emergence: Dynamics at the wildlife–livestock–human interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Terefe, Y.; Deblais, L.; Ghanem, M.; Helmy, Y.A.; Mummed, B.; Chen, D.; Singh, N.; Ahyong, V.; Kalantar, K.; Yimer, G.; et al. Co-occurrence of Campylobacter species in children from eastern Ethiopia, and their association with environmental enteric dysfunction, diarrhea, and host microbiome. Front. Public Health 2020, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Munk, P.; Njage, P.; Van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurek, L.; Ghosh, A. Insects represent a link between food animal farms and the urban environment for antibiotic resistance traits. Appl. Environ. Microbiol. 2014, 80, 3562–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the essentiality of lipopolysaccharide to Gram-negative bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Motta, S.S.; Cluzel, P.; Aldana, M. Adaptive resistance in bacteria requires epigenetic inheritance, genetic noise, and cost of efflux pumps. PLoS ONE 2015, 10, e0118464. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Schneider, C.L. Bacteriophage-mediated horizontal gene transfer: Transduction. In Bacteriophages; Biology, Technology, Therapy; Springer: Cham, Switzerland, 2021; pp. 151–192. [Google Scholar]
- Sørensen, S.J.; Bailey, M.; Hansen, L.H.; Kroer, N.; Wuertz, S. Studying plasmid horizontal transfer in situ: A critical review. Nat. Rev. Microbiol. 2005, 3, 700–710. [Google Scholar] [CrossRef]
- Sun, D. Pull in and push out: Mechanisms of horizontal gene transfer in bacteria. Front. Microbiol. 2018, 9, 2154. [Google Scholar] [CrossRef]
- Rosenfeld, Y.; Shai, Y. Lipopolysaccharide (Endotoxin)-host defense antibacterial peptides interactions: Role in bacterial resistance and prevention of sepsis. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1758, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- Aguilella, V.M.; Queralt-Martín, M.; Alcaraz, A. Bacterial porins. In Electrophysiology of Unconventional Channels and Pores; Springer: Berlin/Heidelberg, Germany, 2015; pp. 101–121. [Google Scholar]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. Apmis 2013, 121, 1–58. [Google Scholar] [CrossRef]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Jiang, Y.; Geng, M.; Bai, L. Targeting Biofilms Therapy: Current Research Strategies and Development Hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef]
- Singh, T.; Singh, P.K.; Das, S.; Wani, S.; Jawed, A.; Dar, S.A. Transcriptome analysis of beta-lactamase genes in diarrheagenic Escherichia coli. Sci. Rep. 2019, 9, 3626. [Google Scholar] [CrossRef] [Green Version]
- Golkar, T.; Zieliński, M.; Berghuis, A.M. Look and outlook on enzyme-mediated macrolide resistance. Front. Microbiol. 2018, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of resistance to aminoglycoside antibiotics: Overview and perspectives. Medchemcomm 2016, 7, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.A. Mechanisms of Bacterial Resistance. In Overcoming Antimicrobial Resistance of the Skin; Springer: Berlin/Heidelberg, Germany, 2021; pp. 3–25. [Google Scholar]
- Kumar, P. Pharmacology of specific drug groups. In Pharmacology and Therapeutics for Dentistry, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 457–487. [Google Scholar]
- Xu, M.; Zhou, Y.N.; Goldstein, B.P.; Jin, D.J. Cross-resistance of Escherichia coli RNA polymerases conferring rifampin resistance to different antibiotics. J. Bacteriol. 2005, 187, 2783–2792. [Google Scholar] [CrossRef] [Green Version]
- Handzlik, J.; Matys, A.; Kieć-Kononowicz, K. Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2013, 2, 28–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuberger, A.; Du, D.; Luisi, B.F. Structure and mechanism of bacterial tripartite efflux pumps. Res. Microbiol. 2018, 169, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Anes, J.; McCusker, M.P.; Fanning, S.; Martins, M. The ins and outs of RND efflux pumps in Escherichia coli. Front. Microbiol. 2015, 6, 587. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, M.; Hamidi-Farahani, R.; Asgari, A.; Esmailkhani, A.; Soleiman-Meigooni, S. Expression of RND efflux pumps mediated antibiotic resistance in Pseudomonas aeruginosa clinical strains. Microb. Pathog. 2021, 153, 104789. [Google Scholar]
- Zhang, L.; Li, X.-Z.; Poole, K. SmeDEF multidrug efflux pump contributes to intrinsic multidrug resistance in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2001, 45, 3497–3503. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129–145. [Google Scholar] [CrossRef]
- Alcalde-Rico, M.; Hernando-Amado, S.; Blanco, P.; Martínez, J.L. Multidrug efflux pumps at the crossroad between antibiotic resistance and bacterial virulence. Front. Microbiol. 2016, 7, 1483. [Google Scholar] [CrossRef] [Green Version]
- Leeson, P.D.; Springthorpe, B. The influence of drug-like concepts on decision-making in medicinal chemistry. Nat. Rev. Drug Discov. 2007, 6, 881–890. [Google Scholar] [CrossRef]
- Hong-Geller, E.; Micheva-Viteva, S. Small molecule screens to identify inhibitors of infectious disease. In Drug Discovery; El Shelmy, H.A., Ed.; InTech: London, UK, 2013; pp. 157–175. [Google Scholar]
- Selin, C.; Stietz, M.S.; Blanchard, J.E.; Gehrke, S.S.; Bernard, S.; Hall, D.G.; Brown, E.D.; Cardona, S.T. A Pipeline for Screening Small Molecules with Growth Inhibitory Activity against Burkholderia cenocepacia. PLoS ONE 2015, 10, e0128587. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Drozd, M.; Pina-Mimbela, R.; Xu, X.; Helmy, Y.A.; Antwi, J.; Fuchs, J.R.; Nislow, C.; Templeton, J.; Blackall, P.J.; et al. Novel Anti-Campylobacter Compounds Identified Using High Throughput Screening of a Pre-selected Enriched Small Molecules Library. Front. Microbiol. 2016, 7, 405. [Google Scholar] [CrossRef] [Green Version]
- Helmy, Y.A.; Kathayat, D.; Ghanem, M.; Jung, K.; Closs, G., Jr.; Deblais, L.; Srivastava, V.; El-Gazzar, M.; Rajashekara, G. Identification and characterization of novel small molecule inhibitors to control Mycoplasma gallisepticum infection in chickens. Vet. Microbiol. 2020, 247, 108799. [Google Scholar] [CrossRef]
- Mingeot-Leclercq, M.-P.; Decout, J.-L. Bacterial lipid membranes as promising targets to fight antimicrobial resistance, molecular foundations and illustration through the renewal of aminoglycoside antibiotics and emergence of amphiphilic aminoglycosides. MedChemComm 2016, 7, 586–611. [Google Scholar] [CrossRef]
- Garg, S.K.; Singh, O.; Juneja, D.; Tyagi, N.; Khurana, A.S.; Qamra, A.; Motlekar, S.; Barkate, H. Resurgence of Polymyxin B for MDR/XDR Gram-Negative Infections: An Overview of Current Evidence. Crit. Care Res. Pract. 2017, 2017, 3635609. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, A.T.; Barker, R.; Rehal, R.; Vandera, K.A.; Harvey, R.D.; Coates, A.R. Mechanism of Action of a Membrane-Active Quinoline-Based Antimicrobial on Natural and Model Bacterial Membranes. Biochemistry 2017, 56, 1163–1174. [Google Scholar] [CrossRef]
- Hart, E.M.; Mitchell, A.M.; Konovalova, A.; Grabowicz, M.; Sheng, J.; Han, X.; Rodriguez-Rivera, F.P.; Schwaid, A.G.; Malinverni, J.C.; Balibar, C.J. A small-molecule inhibitor of BamA impervious to efflux and the outer membrane permeability barrier. Proc. Natl. Acad. Sci. USA 2019, 116, 21748–21757. [Google Scholar] [CrossRef] [Green Version]
- Vrisman, C.M.; Deblais, L.; Helmy, Y.A.; Johnson, R.; Rajashekara, G.; Miller, S.A. Discovery and Characterization of Low-Molecular Weight Inhibitors of Erwinia tracheiphila. Phytopathology 2020, 110, 989–998. [Google Scholar] [CrossRef]
- Srivastava, V.; Deblais, L.; Kathayat, D.; Rotondo, F.; Helmy, Y.A.; Miller, S.A.; Rajashekara, G. Novel Small Molecule Growth Inhibitors of Xanthomonas spp. Causing Bacterial Spot of Tomato. Phytopathology 2021, 111, 940–953. [Google Scholar] [CrossRef]
- Lu, Y.; Deblais, L.; Rajashekara, G.; Miller, S.A.; Helmy, Y.A.; Zhang, H.; Wu, P.; Qiu, Y.; Xu, X. High-throughput screening reveals small molecule modulators inhibitory to Acidovorax citrulli. Plant Pathol. 2020, 69, 818–826. [Google Scholar] [CrossRef]
- Deblais, L.; Vrisman, C.; Kathayat, D.; Helmy, Y.A.; Miller, S.A.; Rajashekara, G. Imidazole and Methoxybenzylamine Growth Inhibitors Reduce Salmonella Persistence in Tomato Plant Tissues. J. Food Prot. 2019, 82, 997–1006. [Google Scholar] [CrossRef]
- Kathayat, D.; Antony, L.; Deblais, L.; Helmy, Y.A.; Scaria, J.; Rajashekara, G. Small Molecule Adjuvants Potentiate Colistin Activity and Attenuate Resistance Development in Escherichia coli by Affecting pmrAB System. Infect. Drug Resist. 2020, 13, 2205–2222. [Google Scholar] [CrossRef]
- Li, Q.; Kang, C. Mechanisms of Action for Small Molecules Revealed by Structural Biology in Drug Discovery. Int. J. Mol. Sci. 2020, 21, 5262. [Google Scholar] [CrossRef]
- Hatcher, H.M. 4—Principles of systemic therapy. In Specialist Training in Oncology; Ajithkumar, T.V., Hatcher, H.M., Eds.; Mosby: Maryland Heights, MO, USA, 2011; pp. 30–44. [Google Scholar]
- Carro, L. Recent Progress in the Development of Small-Molecule FtsZ Inhibitors as Chemical Tools for the Development of Novel Antibiotics. Antibiotics 2019, 8, 217. [Google Scholar] [CrossRef] [Green Version]
- Linciano, P.; Cavalloro, V.; Martino, E.; Kirchmair, J.; Listro, R.; Rossi, D.; Collina, S. Tackling Antimicrobial Resistance with Small Molecules Targeting LsrK: Challenges and Opportunities. J. Med. Chem. 2020, 63, 15243–15257. [Google Scholar] [CrossRef]
- La Manna, S.; Di Natale, C.; Florio, D.; Marasco, D. Peptides as Therapeutic Agents for Inflammatory-Related Diseases. Int. J. Mol. Sci. 2018, 19, 2714. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, E.V.; Gurevich, V.V. Therapeutic potential of small molecules and engineered proteins. Handb. Exp. Pharmacol. 2014, 219, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.A.; Grant, S.S.; Kawate, T.; Iwase, N.; Shimizu, M.; Wivagg, C.; Silvis, M.; Kazyanskaya, E.; Aquadro, J.; Golas, A.; et al. Identification of novel inhibitors of M. tuberculosis growth using whole cell based high-throughput screening. ACS Chem. Biol. 2012, 7, 1377–1384. [Google Scholar] [CrossRef] [Green Version]
- Yep, A.; McQuade, T.; Kirchhoff, P.; Larsen, M.; Mobley, H.L. Inhibitors of TonB function identified by a high-throughput screen for inhibitors of iron acquisition in uropathogenic Escherichia coli CFT073. MBio 2014, 5, e01089-13. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, R.; Sonawane, N.D.; Arumainayagam, D.; Verkman, A.S. Small molecules with antimicrobial activity against E. coli and P. aeruginosa identified by high-throughput screening. Br. J. Pharmacol. 2006, 149, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Sheremet, A.B.; Zigangirova, N.A.; Zayakin, E.S.; Luyksaar, S.I.; Kapotina, L.N.; Nesterenko, L.N.; Kobets, N.V.; Gintsburg, A.L. Small molecule inhibitor of type three secretion system belonging to a class 2, 4-disubstituted-4H-[1,3,4]-thiadiazine-5-ones improves survival and decreases bacterial loads in an airway Pseudomonas aeruginosa infection in mice. Biomed. Res. Int. 2018, 2018, 5810767. [Google Scholar] [CrossRef] [Green Version]
- Nesterenko, L.N.; Zigangirova, N.A.; Zayakin, E.S.; Luyksaar, S.I.; Kobets, N.V.; Balunets, D.V.; Shabalina, L.A.; Bolshakova, T.N.; Dobrynina, O.Y.; Gintsburg, A.L. A small-molecule compound belonging to a class of 2, 4-disubstituted 1, 3, 4-thiadiazine-5-ones suppresses Salmonella infection in vivo. J. Antibiot. 2016, 69, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Thakral, D.; Tae, H.S. Discovery of a Structurally Unique Small Molecule that Inhibits Protein Synthesis. Yale J. Biol. Med. 2017, 90, 35–43. [Google Scholar] [PubMed]
- Stokes, N.R.; Baker, N.; Bennett, J.M.; Berry, J.; Collins, I.; Czaplewski, L.G.; Logan, A.; Macdonald, R.; MacLeod, L.; Peasley, H. An improved small-molecule inhibitor of FtsZ with superior in vitro potency, drug-like properties, and in vivo efficacy. Antimicrob. Agents Chemother. 2013, 57, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Rao, L.; Zhan, L.; Zhou, Y.; Guo, Y.; Wu, X.; Song, Z.; Yu, F. Antibiofilm Activity of Small-Molecule ZY-214-4 Against Staphylococcus aureus. Front. Microbiol. 2021, 12, 618922. [Google Scholar] [CrossRef] [PubMed]
- Soehnlen, M.K.; Tran, M.A.; Lysczek, H.R.; Wolfgang, D.R.; Jayarao, B.M. Identification of novel small molecule antimicrobials targeting Mycoplasma bovis. J. Antimicrob. Chemother. 2011, 66, 574–577. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.G.; Yuhas, C.; McQuade, T.J.; Larsen, M.J.; DiRita, V.J. Narrow-Spectrum Inhibitors of Campylobacter jejuni Flagellar Expression and Growth. Antimicrob. Agents Chemother. 2015, 59, 3880–3886. [Google Scholar] [CrossRef] [Green Version]
- Deblais, L.; Helmy, Y.A.; Kumar, A.; Antwi, J.; Kathayat, D.; Acuna, U.M.; Huang, H.-C.; de Blanco, E.C.; Fuchs, J.R.; Rajashekara, G. Novel narrow spectrum benzyl thiophene sulfonamide derivatives to control Campylobacter. J. Antibiot. 2019, 72, 555–565. [Google Scholar] [CrossRef]
- Dombach, J.L.; Quintana, J.L.; Nagy, T.A.; Wan, C.; Crooks, A.L.; Yu, H.; Su, C.-C.; Yu, E.W.; Shen, J.; Detweiler, C.S. A small molecule that mitigates bacterial infection disrupts Gram-negative cell membranes and is inhibited by cholesterol and neutral lipids. PLoS Pathog. 2020, 16, e1009119. [Google Scholar] [CrossRef]
- Deblais, L.; Helmy, Y.A.; Kathayat, D.; Huang, H.-c.; Miller, S.A.; Rajashekara, G. Novel Imidazole and Methoxybenzylamine Growth Inhibitors Affecting Salmonella Cell Envelope Integrity and its Persistence in Chickens. Sci. Rep. 2018, 8, 13381. [Google Scholar] [CrossRef] [Green Version]
- Kathayat, D.; Helmy, Y.A.; Deblais, L.; Rajashekara, G. Novel small molecules affecting cell membrane as potential therapeutics for avian pathogenic Escherichia coli. Sci. Rep. 2018, 8, 15329. [Google Scholar] [CrossRef] [Green Version]
- Kathayat, D.; Helmy, Y.A.; Deblais, L.; Srivastava, V.; Closs, G., Jr.; Khupse, R.; Rajashekara, G. Novel Small Molecule Growth Inhibitor Affecting Bacterial Outer Membrane Reduces Extraintestinal Pathogenic Escherichia coli (ExPEC) Infection in Avian Model. Microbiol. Spectr. 2021, 9, e0000621. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Deblais, L.; Kassem, I.I.; Kathayat, D.; Rajashekara, G. Novel small molecule modulators of quorum sensing in avian pathogenic Escherichia coli (APEC). Virulence 2018, 9, 1640–1657. [Google Scholar] [CrossRef] [Green Version]
- Helmy, Y.A.; Kathayat, D.; Deblais, L.; Srivastava, V.; Closs, G., Jr.; Tokarski, R.J., 2nd; Ayinde, O.; Fuchs, J.R.; Rajashekara, G. Evaluation of Novel Quorum Sensing Inhibitors Targeting Auto-Inducer 2 (AI-2) for the Control of Avian Pathogenic Escherichia coli Infections in Chickens. Microbiol. Spectr. 2022, 10, e0028622. [Google Scholar] [CrossRef]
- Ashraf, K.; Yasrebi, K.; Hertlein, T.; Ohlsen, K.; Lalk, M.; Hilgeroth, A. Novel Effective Small-Molecule Antibacterials against Enterococcus Strains. Molecules 2017, 22, 2193. [Google Scholar] [CrossRef] [Green Version]
- Thanissery, R.; Zeng, D.; Doyle, R.G.; Theriot, C.M. A Small Molecule-Screening Pipeline to Evaluate the Therapeutic Potential of 2-Aminoimidazole Molecules Against Clostridium difficile. Front. Microbiol. 2018, 9, 1206. [Google Scholar] [CrossRef]
- Muschiol, S.; Normark, S.; Henriques-Normark, B.; Subtil, A. Small molecule inhibitors of the Yersinia type III secretion system impair the development of Chlamydia after entry into host cells. BMC Microbiol. 2009, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.-P.; Davis, J.; Wang, S.; Park, J.G.; Nambiar, M.P.; Schmidt, J.J.; Millard, C.B. Small molecules showing significant protection of mice against botulinum neurotoxin serotype A. PLoS ONE 2010, 5, e10129. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, L.A.; Higgins, D.E. A small-molecule screen identifies the antipsychotic drug pimozide as an inhibitor of Listeria monocytogenes infection. Antimicrob. Agents Chemother. 2009, 53, 756–764. [Google Scholar] [CrossRef] [Green Version]
- Akgul, A.; Al-Janabi, N.; Das, B.; Lawrence, M.; Karsi, A. Small molecules targeting LapB protein prevent Listeria attachment to catfish muscle. PLoS ONE 2017, 12, e0189809. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, M.; Kuo, D.; Jankowsky, E.; Long, L.; Hager, C.; Bandi, K.; Ma, D.; Manoharan, D.; Shoham, Y.; Harte, W.; et al. Small-molecule AgrA inhibitors F12 and F19 act as antivirulence agents against Gram-positive pathogens. Sci. Rep. 2018, 8, 14578. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Li, C.; He, R.; Zhang, Y.; Wang, B.; Zhang, Z.-H.; Ho, C.-T. Research advances on biogenic amines in traditional fermented foods: Emphasis on formation mechanism, detection and control methods. Food Chem. 2023, 405, 134911. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Preda, V.G.; Săndulescu, O. Communication is the key: Biofilms, quorum sensing, formation and prevention. Discoveries 2019, 7, e100. [Google Scholar] [CrossRef]
- Kose-Mutlu, B.; Ergon-Can, T.; Koyuncu, I.; Lee, C.-H. Quorum quenching for effective control of biofouling in membrane bioreactor: A comprehensive review of approaches, applications, and challenges. Environ. Eng. Res. 2019, 24, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Kaplan, H.B.; Greenberg, E.P. Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system. J. Bacteriol. 1985, 163, 1210–1214. [Google Scholar] [CrossRef] [Green Version]
- Seed, P.C.; Passador, L.; Iglewski, B.H. Activation of the Pseudomonas aeruginosa lasI gene by LasR and the Pseudomonas autoinducer PAI: An autoinduction regulatory hierarchy. J. Bacteriol. 1995, 177, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, C.; Greenberg, E.P. Listening in on bacteria: Acyl-homoserine lactone signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 685–695. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.C.; Coyle, B.J.; Williams, P. Virulence Regulation and Quorum Sensing in Staphylococcal Infections: Competitive AgrC Antagonists as Quorum Sensing Inhibitors. J. Med. Chem. 2004, 47, 4633–4641. [Google Scholar] [CrossRef]
- Singh, V.K.; Kavita, K.; Prabhakaran, R.; Jha, B. Cis-9-octadecenoic acid from the rhizospheric bacterium Stenotrophomonas maltophilia BJ01 shows quorum quenching and anti-biofilm activities. Biofouling 2013, 29, 855–867. [Google Scholar] [CrossRef]
- Asfour, H.Z. Anti-Quorum Sensing Natural Compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef]
- Sheng, L.; Olsen, S.A.; Hu, J.; Yue, W.; Means, W.J.; Zhu, M.J. Inhibitory effects of grape seed extract on growth, quorum sensing, and virulence factors of CDC “top-six” non-O157 Shiga toxin producing E. coli. Int. J. Food Microbiol. 2016, 229, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Ravichandiran, V.; Shanmugam, K.; Solomon, A.P. Screening of SdiA inhibitors from Melia dubia seeds extracts towards the hold back of uropathogenic E.coli quorum sensing-regulated factors. Med. Chem. 2013, 9, 819–827. [Google Scholar] [CrossRef]
- Escobar-Muciño, E.; Arenas-Hernández, M.M.P.; Luna-Guevara, M.L. Mechanisms of Inhibition of Quorum Sensing as an Alternative for the Control of E. coli and Salmonella. Microorganisms 2022, 10, 884. [Google Scholar] [CrossRef]
- Utari, P.D.; Vogel, J.; Quax, W.J. Deciphering Physiological Functions of AHL Quorum Quenching Acylases. Front. Microbiol. 2017, 8, 1123. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [Green Version]
- Ha, J.H.; Eo, Y.; Grishaev, A.; Guo, M.; Smith, J.A.; Sintim, H.O.; Kim, E.H.; Cheong, H.K.; Bentley, W.E.; Ryu, K.S. Crystal structures of the LsrR proteins complexed with phospho-AI-2 and two signal-interrupting analogues reveal distinct mechanisms for ligand recognition. J. Am. Chem. Soc. 2013, 135, 15526–15535. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, T.B.; Givskov, M. Quorum-sensing inhibitors as anti-pathogenic drugs. Int. J. Med. Microbiol. 2006, 296, 149–161. [Google Scholar] [CrossRef]
- De Lamo Marin, S.; Xu, Y.; Meijler, M.M.; Janda, K.D. Antibody catalyzed hydrolysis of a quorum sensing signal found in Gram-negative bacteria. Bioorg. Med. Chem. Lett. 2007, 17, 1549–1552. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.A.; Song, Z.; Kristoffersen, P.; et al. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 2003, 22, 3803–3815. [Google Scholar] [CrossRef]
- Gohil, N.; Ramírez-García, R.; Panchasara, H.; Patel, S.; Bhattacharjee, G.; Singh, V. Book Review: Quorum Sensing vs. Quorum Quenching: A Battle With No End in Sight. Front. Cell Infect. Microbiol. 2018, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Wood, T.K.; Lee, J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol. 2015, 23, 707–718. [Google Scholar] [CrossRef] [PubMed]
- DeLisa, M.P.; Wu, C.F.; Wang, L.; Valdes, J.J.; Bentley, W.E. DNA microarray-based identification of genes controlled by autoinducer 2-stimulated quorum sensing in Escherichia coli. J. Bacteriol. 2001, 183, 5239–5247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Curtis, M.M.; Russell, R.; Moreira, C.G.; Adebesin, A.M.; Wang, C.; Williams, N.S.; Taussig, R.; Stewart, D.; Zimmern, P.; Lu, B. QseC inhibitors as an antivirulence approach for Gram-negative pathogens. MBio 2014, 5, e02165-14. [Google Scholar] [CrossRef] [Green Version]
- Peterson, M.M.; Mack, J.L.; Hall, P.R.; Alsup, A.A.; Alexander, S.M.; Sully, E.K.; Sawires, Y.S.; Cheung, A.L.; Otto, M.; Gresham, H.D. Apolipoprotein B is an innate barrier against invasive Staphylococcus aureus infection. Cell Host Microbe 2008, 4, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, D.; Huang, X.; Yang, H.; Qiu, Z.; Zou, L.; Liang, Q.; Shi, Y.; Wu, Y.; Wu, S.; et al. Study on Antibacterial and Quorum-Sensing Inhibition Activities of Cinnamomum camphora Leaf Essential Oil. Molecules 2019, 24, 3792. [Google Scholar] [CrossRef] [Green Version]
- Rasko, D.A.; Moreira, C.G.; Li, D.R.; Reading, N.C.; Ritchie, J.M.; Waldor, M.K.; Williams, N.; Taussig, R.; Wei, S.; Roth, M.; et al. Targeting QseC signaling and virulence for antibiotic development. Science 2008, 321, 1078–1080. [Google Scholar] [CrossRef] [Green Version]
- Witsø, I.L.; Valen Rukke, H.; Benneche, T.; Aamdal Scheie, A. Thiophenone Attenuates Enteropathogenic Escherichia coli O103:H2 Virulence by Interfering with AI-2 Signaling. PLoS ONE 2016, 11, e0157334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cáceres, M.; Hidalgo, W.; Stashenko, E.; Torres, R.; Ortiz, C. Essential Oils of Aromatic Plants with Antibacterial, Anti-Biofilm and Anti-Quorum Sensing Activities against Pathogenic Bacteria. Antibiotics 2020, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girennavar, B.; Cepeda, M.L.; Soni, K.A.; Vikram, A.; Jesudhasan, P.; Jayaprakasha, G.K.; Pillai, S.D.; Patil, B.S. Grapefruit juice and its furocoumarins inhibits autoinducer signaling and biofilm formation in bacteria. Int. J. Food Microbiol. 2008, 125, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Larrosa, M.; Castro-Ibáñez, I.; Allende, A. Plant food extracts and phytochemicals: Their role as Quorum Sensing Inhibitors. Trends Food Sci. Technol. 2015, 43, 189–204. [Google Scholar] [CrossRef]
- Almasoud, A.; Hettiarachchy, N.; Rayaprolu, S.; Babu, D.; Kwon, Y.M.; Mauromoustakos, A. Inhibitory effects of lactic and malic organic acids on autoinducer type 2 (AI-2) quorum sensing of Escherichia coli O157:H7 and Salmonella Typhimurium. LWT-Food Sci. Technol. 2016, 66, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.R.; Lou, Z.; Zhang, J.; Yu, F.; Timilsena, Y.P.; Zhang, C.; Zhang, Y.; Bakry, A.M. Star Anise (Illicium verum Hook. f.) as Quorum Sensing and Biofilm Formation Inhibitor on Foodborne Bacteria: Study in Milk. J. Food Prot. 2017, 80, 645–653. [Google Scholar] [CrossRef]
- Rubini, D.; Banu, S.F.; Subramani, P.; Hari, B.N.V.; Gowrishankar, S.; Pandian, S.K.; Wilson, A.; Nithyanand, P. Extracted chitosan disrupts quorum sensing mediated virulence factors in Urinary tract infection causing pathogens. Pathog. Dis. 2019, 77, ftz009. [Google Scholar] [CrossRef]
- Pan, J.; Xie, X.; Tian, W.; Bahar, A.A.; Lin, N.; Song, F.; An, J.; Ren, D. (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one sensitizes Escherichia coli persister cells to antibiotics. Appl. Microbiol. Biotechnol. 2013, 97, 9145–9154. [Google Scholar] [CrossRef]
- Li, G.; Yan, C.; Xu, Y.; Feng, Y.; Wu, Q.; Lv, X.; Yang, B.; Wang, X.; Xia, X. Punicalagin inhibits Salmonella virulence factors and has anti-quorum-sensing potential. Appl. Environ. Microbiol. 2014, 80, 6204–6211. [Google Scholar] [CrossRef] [Green Version]
- Maskey, R.P.; Asolkar, R.N.; Kapaun, E.; Wagner-Döbler, I.; Laatsch, H. Phytotoxic arylethylamides from limnic bacteria using a screening with microalgae. J. Antibiot. 2002, 55, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, M.E.; Liu, J.; Wallace, J.; Akhlaghi, F.; Rowley, D.C. Secondary metabolites produced by the marine bacterium Halobacillus salinus that inhibit quorum sensing-controlled phenotypes in gram-negative bacteria. Appl. Environ. Microbiol. 2009, 75, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tommonaro, G.; Abbamondi, G.R.; Iodice, C.; Tait, K.; De Rosa, S. Diketopiperazines produced by the halophilic archaeon, Haloterrigena hispanica, activate AHL bioreporters. Microb. Ecol. 2012, 63, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013, 40, 759–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Jeong, J.H.; Lee, K.T.; Rho, J.R.; Choi, H.D.; Kang, J.S.; Son, B.W. γ-Pyrone derivatives, kojic acid methyl ethers from a marine-derived fungusaltenaria sp. Arch. Pharmacal Res. 2003, 26, 532–534. [Google Scholar] [CrossRef]
- Tan, L.Y.; Yin, W.-F.; Chan, K.-G. Silencing quorum sensing through extracts of Melicope lunu-ankenda. Sensors 2012, 12, 4339–4351. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, M.; D’morris, S.; Paul, V.; Warrier, S.; Vasudevan, A.K.; Vanuopadath, M.; Nair, S.S.; Paul-Prasanth, B.; Mohan, C.G.; Biswas, R. Mechanistic understanding of Phenyllactic acid mediated inhibition of quorum sensing and biofilm development in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 8223–8236. [Google Scholar] [CrossRef]
- Krishnan, T.; Yin, W.-F.; Chan, K.-G. Inhibition of quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa PAO1 by Ayurveda spice clove (Syzygium aromaticum) bud extract. Sensors 2012, 12, 4016–4030. [Google Scholar] [CrossRef] [Green Version]
- Vinothkannan, R.; Tamizh, M.M.; Raj, C.D.; Princy, S.A. Fructose furoic acid ester: An effective quorum sensing inhibitor against uropathogenic Escherichia coli. Bioorg. Chem. 2018, 79, 310–318. [Google Scholar] [CrossRef]
- Tello, E.; Castellanos, L.; Arévalo-Ferro, C.; Duque, C. Disruption in quorum-sensing systems and bacterial biofilm inhibition by cembranoid diterpenes isolated from the octocoral Eunicea knighti. J. Nat. Prod. 2012, 75, 1637–1642. [Google Scholar] [CrossRef]
- Sun, J.; Wu, J.; An, B.; de Voogd, N.J.; Cheng, W.; Lin, W. Bromopyrrole alkaloids with the inhibitory effects against the biofilm formation of Gram negative bacteria. Mar. Drugs 2018, 16, 9. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.R.; de Cássia Orlandi Sardi, J.; de Souza Pitangui, N.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Júnior, A.I.M.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Tharmaraj, N.; Shah, N.P. Antimicrobial effects of probiotics against selected pathogenic and spoilage bacteria in cheese-based dips. Int. Food Res. J. 2009, 16, 261–276. [Google Scholar]
- Parente, E.; Brienza, C.; Moles, M.; Ricciardi, A. A comparison of methods for the measurement of bacteriocin activity. J. Microbiol. Methods 1995, 22, 95–108. [Google Scholar] [CrossRef]
- Adimpong, D.B.; Nielsen, D.S.; Sørensen, K.I.; Derkx, P.M.; Jespersen, L. Genotypic characterization and safety assessment of lactic acid bacteria from indigenous African fermented food products. BMC Microbiol. 2012, 12, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Closs, G., Jr.; Jung, K.; Kathayat, D.; Vlasova, A.; Rajashekara, G. Effect of Probiotic E. coli Nissle 1917 Supplementation on the Growth Performance, Immune Responses, Intestinal Morphology, and Gut Microbes of Campylobacter jejuni Infected Chickens. Infect. Immun. 2022, 90, e0033722. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Kassem, I.I.; Rajashekara, G. Immuno-modulatory effect of probiotic E. coli Nissle 1917 in polarized human colonic cells against Campylobacter jejuni infection. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Kassem, I.I.; Kumar, A.; Rajashekara, G. In vitro evaluation of the impact of the probiotic E. coli Nissle 1917 on Campylobacter jejuni’s invasion and intracellular survival in human colonic cells. Front. Microbiol. 2017, 8, 1588. [Google Scholar] [CrossRef] [Green Version]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. Int. Sch. Res. Not. 2013, 2013, 481651. [Google Scholar] [CrossRef] [Green Version]
- Zendeboodi, F.; Khorshidian, N.; Mortazavian, A.M.; da Cruz, A.G. Probiotic: Conceptualization from a new approach. Curr. Opin. Food Sci. 2020, 32, 103–123. [Google Scholar] [CrossRef]
- Saarela, M.H. Safety aspects of next generation probiotics. Curr. Opin. Food Sci. 2019, 30, 8–13. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Kumar, Y.; Singh, L. Health benefits of fermented and functional foods. J. Plant Dev. Sci. 2009, 1, 151–155. [Google Scholar]
- Holzapfel, W.H.; Schillinger, U. Introduction to pre-and probiotics. Food Res. Int. 2002, 35, 109–116. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Haberer, P.; Snel, J.; Schillinger, U.; in’t Veld, J.H.H. Overview of gut flora and probiotics. Int. J. Food Microbiol. 1998, 41, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, S.J.; Gueimonde, M.; Ouwehand, A.C.; Reinikainen, J.P.; Salminen, S.J. Comparison of four methods to enumerate probiotic bifidobacteria in a fermented food product. Food Microbiol. 2006, 23, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Blinkova, L.; Martirosyan, D.M.; Pakhomov, Y.; Dmitrieva, O.; Vaughan, R.; Altshuler, M. Nonculturable forms of bacteria in lyophilized probiotic preparations. Funct. Foods Health Dis. 2014, 4, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Elshaghabee, F.M.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: Status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Franz, C.M.; Omar, N.B.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [Green Version]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Kutsukake, E.; Fukui, T.; Sato, I.; Shirai, T.; Kurihara, T.; Okada, N.; Danbara, H.; Toba, M.; Kohda, N. Oral administration of heat-killed Lactobacillus plantarum strain b240 protected mice against Salmonella enterica Serovar Typhimurium. Biosci. Biotechnol. Biochem. 2010, 74, 1338–1342. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Kuda, T.; An, C.; Kanno, T.; Takahashi, H.; Kimura, B. Inhibitory effects of Leuconostoc mesenteroides 1RM3 isolated from narezushi, a fermented fish with rice, on Listeria monocytogenes infection to Caco-2 cells and A/J mice. Anaerobe 2012, 18, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Jiang, J.; Zhu, W.; Chu, Y. Heat-killed yogurt-containing lactic acid bacteria prevent cytokine-induced barrier disruption in human intestinal Caco-2 cells. Ann. Microbiol. 2016, 66, 171–178. [Google Scholar] [CrossRef]
- Orlando, A.; Refolo, M.; Messa, C.; Amati, L.; Lavermicocca, P.; Guerra, V.; Russo, F. Antiproliferative and proapoptotic effects of viable or heat-killed Lactobacillus paracasei IMPC2. 1 and Lactobacillus rhamnosus GG in HGC-27 gastric and DLD-1 colon cell lines. Nutr. Cancer 2012, 64, 1103–1111. [Google Scholar] [CrossRef]
- Peng, G.C.; Hsu, C.H. The efficacy and safety of heat-killed Lactobacillus paracasei for treatment of perennial allergic rhinitis induced by house-dust mite. Pediatr. Allergy Immunol. 2005, 16, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Shimada, T.; Chen, R.; Lu, M.; Zhang, Q.; Lu, W.; Yin, M.; Enomoto, T.; Cheng, L. Effects of lysed Enterococcus faecalis FK-23 on experimental allergic rhinitis in a murine model. J. Biomed. Res. 2012, 26, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Tareb, R.; Bernardeau, M.; Gueguen, M.; Vernoux, J.-P. In vitro characterization of aggregation and adhesion properties of viable and heat-killed forms of two probiotic Lactobacillus strains and interaction with foodborne zoonotic bacteria, especially Campylobacter jejuni. J. Med. Microbiol. 2013, 62, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Segawa, S.; Wakita, Y.; Hirata, H.; Watari, J. Oral administration of heat-killed Lactobacillus brevis SBC8803 ameliorates alcoholic liver disease in ethanol-containing diet-fed C57BL/6N mice. Int. J. Food Microbiol. 2008, 128, 371–377. [Google Scholar] [CrossRef]
- Hiippala, K. Epithelial Interactions of Gram-Negative Commensals in Human Gastrointestinal Tract. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2020. [Google Scholar]
- Foligné, B.; Daniel, C.; Pot, B. Probiotics from research to market: The possibilities, risks and challenges. Curr. Opin. Microbiol. 2013, 16, 284–292. [Google Scholar] [CrossRef]
- Howard, F.; Bradley, J.; Flynn, D.; Noone, P.; Szawatkowski, M. Outbreak of necrotising enterocolitis caused by Clostridium butyricum. Lancet 1977, 310, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Cassir, N.; Benamar, S.; La Scola, B. Clostridium butyricum: From beneficial to a new emerging pathogen. Clin. Microbiol. Infect. 2016, 22, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional characterization of novel Faecalibacterium prausnitzii strains isolated from healthy volunteers: A step forward in the use of F. prausnitzii as a next-generation probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Qiu, X.; Zhang, H.; Yang, X.; Hong, N.; Yang, Y.; Chen, H.; Yu, C. Faecalibacterium prausnitzii inhibits interleukin-17 to ameliorate colorectal colitis in rats. PLoS ONE 2014, 9, e109146. [Google Scholar] [CrossRef] [Green Version]
- Foditsch, C.; Pereira, R.V.V.; Ganda, E.K.; Gomez, M.S.; Marques, E.C.; Santin, T.; Bicalho, R.C. Oral administration of Faecalibacterium prausnitzii decreased the incidence of severe diarrhea and related mortality rate and increased weight gain in preweaned dairy heifers. PLoS ONE 2015, 10, e0145485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, R.R.; Preedy, V.R. Bioactive Foods in Promoting Health: Probiotics and Prebiotics; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Kumar, M.; Nagpal, R.; Verma, V.; Kumar, A.; Kaur, N.; Hemalatha, R.; Gautam, S.K.; Singh, B. Probiotic metabolites as epigenetic targets in the prevention of colon cancer. Nutr. Rev. 2013, 71, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Chenoll, E.; Moreno, I.; Sánchez, M.; Garcia-Grau, I.; Silva, Á.; González-Monfort, M.; Genovés, S.; Vilella, F.; Seco-Durban, C.; Simón, C. Selection of new probiotics for endometrial health. Front. Cell. Infect. Microbiol. 2019, 9, 114. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Skonieczna-Żydecka, K.; Kaczmarczyk, M.; Łoniewski, I.; Lara, L.F.; Koulaouzidis, A.; Misera, A.; Maciejewska, D.; Marlicz, W. A systematic review, meta-analysis, and meta-regression evaluating the efficacy and mechanisms of action of probiotics and synbiotics in the prevention of surgical site infections and surgery-related complications. J. Clin. Med. 2018, 7, 556. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Jiang, X.-Y.; Zhou, L.-S.; Song, J.-H.; Zhang, X. Effects of Probiotics on Intestinal Mucosa Barrier in Patients With Colorectal Cancer after Operation: Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e3342. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Helmy, Y.A.; Fritts, Z.; Vlasova, A.; Saif, L.J.; Rajashekara, G. Anti-rotavirus Properties and Mechanisms of Selected Gram-Positive and Gram-Negative Probiotics on Polarized Human Colonic (HT-29) Cells. Probiotics Antimicrob. Proteins 2022, 15, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 3320–3333. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Apetroaie-Constantin, C.; Mikkola, R.; Andersson, M.A.; Teplova, V.; Suominen, I.; Johansson, T.; Salkinoja-Salonen, M. Bacillus subtilis and B. mojavensis strains connected to food poisoning produce the heat stable toxin amylosin. J. Appl. Microbiol. 2009, 106, 1976–1985. [Google Scholar] [CrossRef]
- Mawad, A.; Helmy, Y.A.; Shalkami, A.-G.; Kathayat, D.; Rajashekara, G.E. coli Nissle microencapsulation in alginate-chitosan nanoparticles and its effect on Campylobacter jejuni in vitro. Appl. Microbiol. Biotechnol. 2018, 102, 10675–10690. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Di Lena, M.; Quero, G.M.; Santovito, E.; Verran, J.; De Angelis, M.; Fusco, V. A selective medium for isolation and accurate enumeration of Lactobacillus casei-group members in probiotic milks and dairy products. Int. Dairy J. 2015, 47, 27–36. [Google Scholar] [CrossRef]
- de Matos, F.E.; Santos, T.T.; Burns, P.G.; Reinheimer, J.A.; Vinderola, C.G.; Trindade, C.S.F. Evaluation of lactobacillus paracasei LP11 and lactobacillus rhamnosus 64 potential as candidates for use as probiotics in functional foods. J. Microbiol. Biotechnol. Food Sci. 2020, 9, 1126–1133. [Google Scholar]
- Bian, X.; Evivie, S.E.; Muhammad, Z.; Luo, G.-W.; Liang, H.-Z.; Wang, N.-N.; Huo, G.-C. In vitro assessment of the antimicrobial potentials of Lactobacillus helveticus strains isolated from traditional cheese in Sinkiang China against food-borne pathogens. Food Funct. 2016, 7, 789–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson-Henry, K.C.; Hagen, K.E.; Gordonpour, M.; Tompkins, T.A.; Sherman, P.M. Surface-layer protein extracts from Lactobacillus helveticus inhibit enterohaemorrhagic Escherichia coli O157: H7 adhesion to epithelial cells. Cell. Microbiol. 2007, 9, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic activity of Lactobacillus reuteri strains on the adhesion characteristics of selected pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.P.; Thibodeaux, C.H.; Peña, J.A.; Ferry, G.D.; Versalovic, J. Probiotic Lactobacillus reuteri suppress proinflammatory cytokines via c-Jun. Inflamm. Bowel Dis. 2008, 14, 1068–1083. [Google Scholar] [CrossRef]
- Dinev, T.; Beev, G.; Denev, S.; Dermendzhieva, D.; Tzanova, M.; Valkova, E. Antimicrobial activity of Lactobacillus acidophilus against pathogenic and food spoilage microorganisms: A review. Agric. Sci. Technol. 2017, 9, 3–9. [Google Scholar] [CrossRef]
- Kathayat, D.; Closs, G., Jr.; Helmy, Y.A.; Deblais, L.; Srivastava, V.; Rajashekara, G. In Vitro and In Vivo Evaluation of Lacticaseibacillus rhamnosus GG and Bifidobacterium lactis Bb12 Against Avian Pathogenic Escherichia coli and Identification of Novel Probiotic-Derived Bioactive Peptides. Probiotics Antimicrob. Proteins 2022, 14, 1012–1028. [Google Scholar] [CrossRef]
- Evivie, S.E.; Abdelazez, A.; Li, B.; Lu, S.; Liu, F.; Huo, G. Lactobacillus delbrueckii subsp. bulgaricus KLDS 1.0207 exerts antimicrobial and cytotoxic effects in vitro and improves blood biochemical parameters in vivo against notable foodborne pathogens. Front. Microbiol. 2020, 11, 583070. [Google Scholar] [CrossRef]
- O’Mahony, D.; Murphy, S.; Boileau, T.; Park, J.; O’Brien, F.; Groeger, D.; Konieczna, P.; Ziegler, M.; Scully, P.; Shanahan, F.; et al. Bifidobacterium animalis AHC7 protects against pathogen-induced NF-κB activation in vivo. BMC Immunol. 2010, 11, 63. [Google Scholar]
- Lim, H.J.; Shin, H.S. Antimicrobial and immunomodulatory effects of bifidobacterium strains: A review. J. Microbiol. Biotechnol. 2020, 30, 1793–1800. [Google Scholar] [CrossRef]
- Rahimifard, N.; Naseri, M. Evaluation and comparison of three antimicrobial activity methods using Bifidobacteriabifidum and Bifidobacteria infantis as probiotic bacteria against Salmonella enterica serotype Enteritidis. J. Bacteriol. Mycol. 2016, 2, 24. [Google Scholar]
- Ruiz, P.A.; Hoffmann, M.; Szcesny, S.; Blaut, M.; Haller, D. Innate mechanisms for Bifidobacterium lactis to activate transient pro-inflammatory host responses in intestinal epithelial cells after the colonization of germ-free rats. Immunology 2005, 115, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.V.; Kollanoor Johny, A. Characterizing the antimicrobial function of a dairy-originated probiotic, Propionibacterium freudenreichii, against multidrug-resistant Salmonella enterica serovar Heidelberg in turkey poults. Front. Microbiol. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Prete, R.; Garcia-Gonzalez, N.; Battista, N.; Corsetti, A. Interaction of food-associated Lactobacillus plantarum with human derived intestinal epithelial cells. In Proceedings of the 12th International Scientific Conference on Probiotics, Prebiotics, Gut Microbiota and Health-IPC2018, Budapest, Hungary, 18–21 June 2018; pp. 47–48. [Google Scholar]
- Abbasiliasi, S.; Tan, J.S.; Bashokouh, F.; Ibrahim, T.A.T.; Mustafa, S.; Vakhshiteh, F.; Sivasamboo, S.; Ariff, A.B. In vitro assessment of Pediococcus acidilactici Kp10 for its potential use in the food industry. BMC Microbiol. 2017, 17, 121. [Google Scholar] [CrossRef] [Green Version]
- Benmechernene, Z.; Chentouf, H.F.; Yahia, B.; Fatima, G.; Quintela-Baluja, M.; Calo-Mata, P.; Barros-Velázquez, J. Technological aptitude and applications of Leuconostoc mesenteroides bioactive strains isolated from Algerian raw camel milk. Biomed. Res. Int. 2013, 2013, 418132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef] [Green Version]
- Stašková, A.; Sondorová, M.; Nemcová, R.; Kačírová, J.; Maďar, M. Antimicrobial and Antibiofilm Activity of the Probiotic Strain Streptococcus salivarius K12 against Oral Potential Pathogens. Antibiotics 2021, 10, 793. [Google Scholar] [CrossRef]
- Evivie, S.E.; Ogwu, M.C.; Abdelazez, A.; Bian, X.; Liu, F.; Li, B.; Huo, G. Suppressive effects of Streptococcus thermophilus KLDS 3.1003 on some foodborne pathogens revealed through in vitro, in vivo and genomic insights. Food Funct. 2020, 11, 6573–6587. [Google Scholar] [CrossRef]
- Masumuzzaman, M.; Evivie, S.E.; Ogwu, M.C.; Li, B.; Du, J.; Li, W.; Huo, G.; Liu, F.; Wang, S. Genomic and in vitro properties of the dairy Streptococcus thermophilus SMQ-301 strain against selected pathogens. Food Funct. 2021, 12, 7017–7028. [Google Scholar] [CrossRef]
- Jensen, G.S.; Cash, H.A.; Farmer, S.; Keller, D. Inactivated probiotic Bacillus coagulans GBI-30 induces complex immune activating, anti-inflammatory, and regenerative markers in vitro. J. Inflamm. Res. 2017, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Dalmasso, G.; Cottrez, F.; Imbert, V.; Lagadec, P.; Peyron, J.-F.; Rampal, P.; Czerucka, D.; Groux, H. Saccharomyces boulardii inhibits inflammatory bowel disease by trapping T cells in mesenteric lymph nodes. Gastroenterology 2006, 131, 1812–1825. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Brunser, O.; Szajewska, H. Saccharomyces boulardii in childhood. Eur. J. Pediatr. 2009, 168, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Czerucka, D.; Rampal, P. Diversity of Saccharomyces boulardii CNCM I-745 mechanisms of action against intestinal infections. World J. Gastroenterol. 2019, 25, 2188. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Taguchi, H.; Yamaguchi, H.; Osaki, T.; Komatsu, A.; Kamiya, S. The effect of probiotic treatment with Clostridium butyricum on enterohemorrhagic Escherichia coli O157: H7 infection in mice. FEMS Immunol. Med. Microbiol. 2004, 41, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.; Inoue, I.; Tanaka, M.; Matsuda, N.; Nakano, T.; Awata, T.; Katayama, S.; Alpers, D.H.; Komoda, T. Clostridium butyricum MIYAIRI 588 improves high-fat diet-induced non-alcoholic fatty liver disease in rats. Dig. Dis. Sci. 2013, 58, 3534–3544. [Google Scholar] [CrossRef] [PubMed]
- Taha-Abdelaziz, K.; Astill, J.; Kulkarni, R.R.; Read, L.R.; Najarian, A.; Farber, J.M.; Sharif, S. In vitro assessment of immunomodulatory and anti-Campylobacter activities of probiotic lactobacilli. Sci. Rep. 2019, 9, 17903. [Google Scholar] [CrossRef] [Green Version]
- Ty, M.; Taha-Abdelaziz, K.; Demey, V.; Castex, M.; Sharif, S.; Parkinson, J. Performance of distinct microbial based solutions in a Campylobacter infection challenge model in poultry. Anim. Microbiome 2022, 4, 2. [Google Scholar] [CrossRef]
- Shojadoost, B.; Alizadeh, M.; Boodhoo, N.; Astill, J.; Karimi, S.H.; Shoja Doost, J.; Taha-Abdelaziz, K.; Kulkarni, R.; Sharif, S. Effects of Treatment with Lactobacilli on Necrotic Enteritis in Broiler Chickens. Probiotics Antimicrob. Proteins 2022, 14, 1110–1129. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Manning, T.S.; Gibson, G.R. Prebiotics. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 287–298. [Google Scholar] [CrossRef]
- Mohanty, D.; Misra, S.; Mohapatra, S.; Sahu, P.S. Prebiotics and synbiotics: Recent concepts in nutrition. Food Biosci. 2018, 26, 152–160. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarangi, N.R.; Babu, L.K.; Kumar, A.; Pradhan, C.R.; Pati, P.K.; Mishra, J.P. Effect of dietary supplementation of prebiotic, probiotic, and synbiotic on growth performance and carcass characteristics of broiler chickens. Vet. World 2016, 9, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solis-Cruz, B.; Hernandez-Patlan, D.; Hargis, B.M.; Tellez, G. Use of prebiotics as an alternative to antibiotic growth promoters in the poultry industry. In Prebiotics and Probiotics-Potential Benefits in Nutrition and Health; IntechOpen: London, UK, 2019. [Google Scholar]
- Rajendran, S.R.C.K.; Okolie, C.L.; Udenigwe, C.C.; Mason, B. Structural features underlying prebiotic activity of conventional and potential prebiotic oligosaccharides in food and health. J. Food Biochem. 2017, 41, e12389. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Connolly, M.L.; Lovegrove, J.A.; Tuohy, K.M. Konjac glucomannan hydrolysate beneficially modulates bacterial composition and activity within the faecal microbiota. J. Funct. Foods 2010, 2, 219–224. [Google Scholar] [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Wilson, B.; Whelan, K. Prebiotic inulin-type fructans and galacto-oligosaccharides: Definition, specificity, function, and application in gastrointestinal disorders. J. Gastroenterol. Hepatol. 2017, 32, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Micciche, A.C.; Foley, S.L.; Pavlidis, H.O.; McIntyre, D.R.; Ricke, S.C. A Review of Prebiotics Against Salmonella in Poultry: Current and Future Potential for Microbiome Research Applications. Front. Vet. Sci. 2018, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.A.; Jang, M.J.; Kim, S.Y.; Yang, Y.; Pavlidis, H.O.; Ricke, S.C. Potential for Prebiotics as Feed Additives to Limit Foodborne Campylobacter Establishment in the Poultry Gastrointestinal Tract. Front. Microbiol. 2019, 10, 91. [Google Scholar] [CrossRef] [Green Version]
- Elshaghabee, F.M.F.; Rokana, N. Mitigation of antibiotic resistance using probiotics, prebiotics and synbiotics. A review. Environ. Chem. Lett. 2022, 20, 1295–1308. [Google Scholar] [CrossRef]
- Kondepudi, K.K.; Ambalam, P.; Nilsson, I.; Wadström, T.; Ljungh, A. Prebiotic-non-digestible oligosaccharides preference of probiotic bifidobacteria and antimicrobial activity against Clostridium difficile. Anaerobe 2012, 18, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Koruri, S.S.; Chowdhury, R.; Bhattacharya, P. Potentiation of functional and antimicrobial activities through synergistic growth of probiotic Pediococcus acidilactici with natural prebiotics (garlic, basil). In Microbes in the Spotlight: Recent Progress in the Understanding of Beneficial and Harmful Microorganisms; Brown Walker Press: Irvine, CA, USA, 2016; pp. 219–224. [Google Scholar]
- Bomba, A.; Nemcová, R.; Gancarcíková, S.; Herich, R.; Guba, P.; Mudronová, D. Improvement of the probiotic effect of micro-organisms by their combination with maltodextrins, fructo-oligosaccharides and polyunsaturated fatty acids. Br. J. Nutr. 2002, 88, S95–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.; Burmeister, S.; Brazier, J. Effect of the prebiotic oligofructose on relapse of Clostridium difficile-associated diarrhea: A randomized, controlled study. Clin. Gastroenterol. Hepatol. 2005, 3, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Teng, P.-Y.; Kim, W.K. Review: Roles of Prebiotics in Intestinal Ecosystem of Broilers. Front. Vet. Sci. 2018, 5, 245. [Google Scholar] [CrossRef] [Green Version]
- Ricke, S.C. Impact of Prebiotics on Poultry Production and Food Safety. Yale J. Biol. Med. 2018, 91, 151–159. [Google Scholar]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Taha, A.E.; Soliman, S.M.; Ahmed, A.E.; El-kott, A.F.; et al. Alternatives to antibiotics for organic poultry production: Types, modes of action and impacts on bird’s health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef]
- Svensson, U.; Håkansson, J. Safety of food and beverages: Safety of probiotics and prebiotics. Encycl. Food Saf. 2014, 3, 441–446. [Google Scholar]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.-Y. Antimicrobial peptides: New hope in the war against multidrug resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Di Somma, A.; Moretta, A.; Canè, C.; Cirillo, A.; Duilio, A. Antimicrobial and Antibiofilm Peptides. Biomolecules 2020, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Dennison, S.R.; Harris, F.; Mura, M.; Phoenix, D.A. An Atlas of Anionic Antimicrobial Peptides from Amphibians. Curr. Protein Pept. Sci. 2018, 19, 823–838. [Google Scholar] [CrossRef]
- Almarwani, B.; Phambu, N.; Hamada, Y.Z.; Sunda-Meya, A. Interactions of an Anionic Antimicrobial Peptide with Zinc(II): Application to Bacterial Mimetic Membranes. Langmuir 2020, 36, 14554–14562. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef]
- Gennaro, R.; Zanetti, M. Structural features and biological activities of the cathelicidin-derived antimicrobial peptides. Biopolymers 2000, 55, 31–49. [Google Scholar] [CrossRef]
- Lewies, A.; Wentzel, J.F.; Jacobs, G.; Du Plessis, L.H. The Potential Use of Natural and Structural Analogues of Antimicrobial Peptides in the Fight against Neglected Tropical Diseases. Molecules 2015, 20, 15392–15433. [Google Scholar] [CrossRef]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Starling, S. Innate immunity: A new way out for lysozyme. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 567. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Thomas, U.; Pellegrini, A. A helix-loop-helix peptide at the upper lip of the active site cleft of lysozyme confers potent antimicrobial activity with membrane permeabilization action. J. Biol. Chem. 2001, 276, 43767–43774. [Google Scholar] [CrossRef] [Green Version]
- Genco, C.A.; Maloy, W.L.; Kari, U.P.; Motley, M. Antimicrobial activity of magainin analogues against anaerobic oral pathogens. Int. J. Antimicrob. Agents 2003, 21, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, L.; Weiser, J.N.; Axelsen, P.H. Antibacterial and antimembrane activities of cecropin A in Escherichia coli. Antimicrob. Agents Chemother. 2000, 44, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, A.; Ueno, S.; Zhang, H.; Lee, J.M.; Kato, Y. Cecropin P1 and novel nematode cecropins: A bacteria-inducible antimicrobial peptide family in the nematode Ascaris suum. Biochem. J. 2005, 390, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Franco, I.; Pérez, M.D.; Castillo, E.; Calvo, M.; Sánchez, L. Effect of high pressure on the structure and antibacterial activity of bovine lactoferrin treated in different media. J. Dairy Res. 2013, 80, 283–290. [Google Scholar] [CrossRef]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; de Almeida, R.; Martins, C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A2; of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Cienc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, W.; Du, S.; Xi, X.; Ma, C.; Wang, L.; Zhou, M.; Chen, T. Bioevaluation and Targeted Modification of Temporin-FL From the Skin Secretion of Dark-Spotted Frog (Pelophylax nigromaculatus). Front. Mol. Biosci. 2021, 8, 707013. [Google Scholar] [CrossRef]
- Brancaccio, D.; Pizzo, E.; Cafaro, V.; Notomista, E.; De Lise, F.; Bosso, A.; Gaglione, R.; Merlino, F.; Novellino, E.; Ungaro, F.; et al. Antimicrobial peptide Temporin-L complexed with anionic cyclodextrins results in a potent and safe agent against sessile bacteria. Int. J. Pharm. 2020, 584, 119437. [Google Scholar] [CrossRef]
- Park, C.B.; Yi, K.S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.H.; Cho, Y.; Lehrer, R.I. Effects of pH and salinity on the antimicrobial properties of clavanins. Infect. Immun. 1997, 65, 2898–2903. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, D.A.; Hurst, M.A.; Fujii, C.A.; Kung, A.H.; Ho, J.F.; Cheng, F.C.; Loury, D.J.; Fiddes, J.C. Protegrin-1: A broad-spectrum, rapidly microbicidal peptide with in vivo activity. Antimicrob. Agents Chemother. 1997, 41, 1738–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.G.; Xie, H.W.; Jin, G.; Wang, W.G.; Zhang, Y.; Guo, Y. Preliminary study on high-level expression of tandem-arranged tachyplesin-encoding gene in Bacillus subtilis Wb800 and its antibacterial activity. Mar. Biotechnol. 2009, 11, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Michels, K.; Nemeth, E.; Ganz, T.; Mehrad, B. Hepcidin and Host Defense against Infectious Diseases. PLoS Pathog. 2015, 11, e1004998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaskovich, M.A.T.; Hansford, K.A.; Gong, Y.; Butler, M.S.; Muldoon, C.; Huang, J.X.; Ramu, S.; Silva, A.B.; Cheng, M.; Kavanagh, A.M.; et al. Protein-inspired antibiotics active against vancomycin- and daptomycin-resistant bacteria. Nat. Commun. 2018, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [Green Version]
- Kathayat, D.; Closs, G.; Helmy Yosra, A.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Peptides Affecting the Outer Membrane Lipid Asymmetry System (MlaA-OmpC/F) Reduce Avian Pathogenic Escherichia coli (APEC) Colonization in Chickens. Appl. Environ. Microbiol. 2021, 87, e00567-21. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sun, Y.; Kang, Y.; Shang, D. Antimicrobial peptide temporin-1CEa isolated from frog skin secretions inhibits the proinflammatory response in lipopolysaccharide-stimulated RAW264.7 murine macrophages through the MyD88-dependent signaling pathway. Mol. Immunol. 2021, 132, 227–235. [Google Scholar] [CrossRef]
- Lee, T.H.; Hall, K.N.; Aguilar, M.I. Antimicrobial Peptide Structure and Mechanism of Action: A Focus on the Role of Membrane Structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef]
- Mandal, S.M.; Khan, J.; Mahata, D.; Saha, S.; Sengupta, J.; Silva, O.N.; Das, S.; Mandal, M.; Franco, O.L. A self-assembled clavanin A-coated amniotic membrane scaffold for the prevention of biofilm formation by ocular surface fungal pathogens. Biofouling 2017, 33, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Li, S.; Cheng, Y.; Nie, L.; Wang, G.; Lv, C.; Wei, W.; Cheng, C.; Hou, F.; et al. Novel short antimicrobial peptide isolated from Xenopus laevis skin. J. Pept. Sci. 2017, 23, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.H.; Meneguetti, B.T.; Costa, B.O.; Buccini, D.F.; Oshiro, K.G.N.; Preza, S.L.E.; Carvalho, C.M.E.; Migliolo, L.; Franco, O.L. Non-Lytic Antibacterial Peptides That Translocate Through Bacterial Membranes to Act on Intracellular Targets. Int. J. Mol. Sci. 2019, 20, 4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardirossian, M.; Barrière, Q.; Timchenko, T.; Müller, C.; Pacor, S.; Mergaert, P.; Scocchi, M.; Wilson, D.N. Fragments of the Nonlytic Proline-Rich Antimicrobial Peptide Bac5 Kill Escherichia coli Cells by Inhibiting Protein Synthesis. Antimicrob. Agents Chemother. 2018, 62, e00534-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Fu, J.; Zhao, Y.; Shi, H.; Hu, H.; Wang, H. Escherichia coli PagP Enzyme-Based De Novo Design and In Vitro Activity of Antibacterial Peptide LL-37. Med. Sci. Monit. 2017, 23, 2558–2564. [Google Scholar] [CrossRef] [Green Version]
- Braffman, N.R.; Piscotta, F.J.; Hauver, J.; Campbell, E.A.; Link, A.J.; Darst, S.A. Structural mechanism of transcription inhibition by lasso peptides microcin J25 and capistruin. Proc. Natl. Acad. Sci. USA 2019, 116, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Kurpe, S.R.; Grishin, S.Y.; Surin, A.K.; Panfilov, A.V.; Slizen, M.V.; Chowdhury, S.D.; Galzitskaya, O.V. Antimicrobial and Amyloidogenic Activity of Peptides. Can Antimicrobial Peptides Be Used against SARS-CoV-2? Int. J. Mol. Sci. 2020, 21, 9552. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist. Updates 2016, 26, 43–57. [Google Scholar] [CrossRef]
- Haney, E.F.; Hancock, R.E. Peptide design for antimicrobial and immunomodulatory applications. Biopolymers 2013, 100, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Engelkirk, P.G.; Duben-Engelkirk, J.; Fader, R.C. Burton’s Microbiology for the Health Sciences; Jones & Bartlett Publishers: Burlington, MA, USA, 2020. [Google Scholar]
- Sabour, P.M.; Griffiths, M.W. Bacteriophages in the Control of Food-and Waterborne Pathogens; American Society for Microbiology Press: Washington, DC, USA, 2010. [Google Scholar]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as alternatives to antibiotics in clinical care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Golkar, Z.; Bagasra, O.; Pace, D.G. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries. 2014, 8, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, J.; Sutherland, I.W. The use of phages for the removal of infectious biofilms. Curr. Pharm. Biotechnol. 2008, 9, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Ligonenko, O.V.; Borysenko, M.; Digtyar, I.; Ivashchenko, D.; Zubakha, A.; Chorna, I.; Shumeyko, I.; Storozhenko, O.; Gorb, L.; Ligonenko, O.O. Application of bacteriophages in complex of treatment of a shot-gun wounds of soft tissues in the patients, suffering multiple allergy for antibiotics. Klin. Khirurhiia 2015, 10, 65–66. [Google Scholar]
- El-Shibiny, A.; El-Sahhar, S. Bacteriophages: The possible solution to treat infections caused by pathogenic bacteria. Can. J. Microbiol. 2017, 63, 865–879. [Google Scholar] [CrossRef] [Green Version]
- Kashoma, I.P.; Srivastava, V.; Rajashekara, G. Advances in Vaccines for Controlling Campylobacter in Poultry. In Food Safety in Poultry Meat Production; Venkitanarayanan, K., Thakur, S., Ricke, S.C., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 191–210. [Google Scholar]
- Abubakar, S.; Suleiman, B.H.; Abbagana, B.A.; Mustafa, I.A.; Musa, I.A. Novel uses of bacteriophages in the treatment of human infections and antibiotic resistance. Am. J. Biosci. 2016, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Dufour, N.; Delattre, R.; Ricard, J.-D.; Debarbieux, L. The lysis of pathogenic Escherichia coli by bacteriophages releases less endotoxin than by β-lactams. Clin. Infect. Dis. 2017, 64, 1582–1588. [Google Scholar] [CrossRef]
- Majewska, J.; Beta, W.; Lecion, D.; Hodyra-Stefaniak, K.; Kłopot, A.; Kaźmierczak, Z.; Miernikiewicz, P.; Piotrowicz, A.; Ciekot, J.; Owczarek, B. Oral application of T4 phage induces weak antibody production in the gut and in the blood. Viruses 2015, 7, 4783–4799. [Google Scholar] [CrossRef]
- Love, M.J.; Bhandari, D.; Dobson, R.C.; Billington, C. Potential for bacteriophage endolysins to supplement or replace antibiotics in food production and clinical care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Kucharewicz-Krukowska, A.; Slopek, S. Immunogenic effect of bacteriophage in patients subjected to phage therapy. Arch. Immunol. Ther. Exp. 1987, 35, 553–561. [Google Scholar]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.-A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): A randomised, controlled, double-blind phase 1/2 trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Speck, P.; Smithyman, A. Safety and efficacy of phage therapy via the intravenous route. FEMS Microbiol. Lett. 2016, 363, fnv242. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Pabary, R.; Singh, C.; Morales, S.; Bush, A.; Alshafi, K.; Bilton, D.; Alton, E.W.; Smithyman, A.; Davies, J.C. Antipseudomonal bacteriophage reduces infective burden and inflammatory response in murine lung. Antimicrob. Agents Chemother. 2016, 60, 744–751. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mi, Z.; Niu, W.; An, X.; Yuan, X.; Liu, H.; Li, P.; Liu, Y.; Feng, Y.; Huang, Y. Intranasal treatment with bacteriophage rescues mice from Acinetobacter baumannii-mediated pneumonia. Future Microbiol. 2016, 11, 631–641. [Google Scholar] [CrossRef]
- Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Leitner, L.; Mehnert, U.; Chkhotua, A.; Kessler, T.M.; Sybesma, W. Adapted bacteriophages for treating urinary tract infections. Front. Microbiol. 2018, 9, 1832. [Google Scholar] [CrossRef] [Green Version]
- Sarker, S.A.; Brüssow, H. From bench to bed and back again: Phage therapy of childhood Escherichia coli diarrhea. Ann. N. Y. Acad. Sci. 2016, 1372, 42–52. [Google Scholar] [CrossRef]
- Rhoads, D.; Wolcott, R.; Kuskowski, M.; Wolcott, B.; Ward, L.; Sulakvelidze, A. Bacteriophage therapy of venous leg ulcers in humans: Results of a phase I safety trial. J. Wound Care 2009, 18, 237–243. [Google Scholar] [CrossRef]
- Rose, T.; Verbeken, G.; De Vos, D.; Merabishvili, M.; Vaneechoutte, M.; Lavigne, R.; Jennes, S.; Zizi, M.; Pirnay, J.-P. Experimental phage therapy of burn wound infection: Difficult first steps. Int. J. Burn. Trauma 2014, 4, 66. [Google Scholar]
- Yoichi, M.; Abe, M.; Miyanaga, K.; Unno, H.; Tanji, Y. Alteration of tail fiber protein gp38 enables T2 phage to infect Escherichia coli O157: H7. J. Biotechnol. 2005, 115, 101–107. [Google Scholar] [CrossRef]
- Vander Elst, N.; Linden, S.B.; Lavigne, R.; Meyer, E.; Briers, Y.; Nelson, D.C. Characterization of the Bacteriophage-Derived Endolysins PlySs2 and PlySs9 with In Vitro Lytic Activity against Bovine Mastitis Streptococcus uberis. Antibiotics 2020, 9, 621. [Google Scholar] [CrossRef] [PubMed]
- Gilmer, D.B.; Schmitz, J.E.; Euler, C.W.; Fischetti, V.A. Novel bacteriophage lysin with broad lytic activity protects against mixed infection by Streptococcus pyogenes and methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 2743–2750. [Google Scholar] [CrossRef] [Green Version]
- D’Angelantonio, D.; Scattolini, S.; Boni, A.; Neri, D.; Di Serafino, G.; Connerton, P.; Connerton, I.; Pomilio, F.; Di Giannatale, E.; Migliorati, G.; et al. Bacteriophage Therapy to Reduce Colonization of Campylobacter jejuni in Broiler Chickens before Slaughter. Viruses 2021, 13, 1428. [Google Scholar] [CrossRef] [PubMed]
- O’Flynn, G.; Ross, R.P.; Fitzgerald, G.F.; Coffey, A. Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2004, 70, 3417–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigwood, T.; Hudson, J.A.; Billington, C. Influence of host and bacteriophage concentrations on the inactivation of food-borne pathogenic bacteria by two phages. FEMS Microbiol. Lett. 2009, 291, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Wagenaar, J.A.; Van Bergen, M.A.; Mueller, M.A.; Wassenaar, T.M.; Carlton, R.M. Phage therapy reduces Campylobacter jejuni colonization in broilers. Vet. Microbiol. 2005, 109, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Modi, R.; Hirvi, Y.; Hill, A.; Griffiths, M.W. Effect of phage on survival of Salmonella enteritidis during manufacture and storage of cheddar cheese made from raw and pasteurized milk. J. Food Prot. 2001, 64, 927–933. [Google Scholar] [CrossRef]
- Bigot, B.; Lee, W.J.; McIntyre, L.; Wilson, T.; Hudson, J.A.; Billington, C.; Heinemann, J.A. Control of Listeria monocytogenes growth in a ready-to-eat poultry product using a bacteriophage. Food Microbiol. 2011, 28, 1448–1452. [Google Scholar] [CrossRef]
- Bueno, E.; García, P.; Martínez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef]
- Lee, N.-Y.; Ko, W.-C.; Hsueh, P.-R. Nanoparticles in the treatment of infections caused by multidrug-resistant organisms. Front. Pharmacol. 2019, 10, 1153. [Google Scholar] [CrossRef] [Green Version]
- Hemeg, H.A. Nanomaterials for alternative antibacterial therapy. Int. J. Nanomed. 2017, 12, 8211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014.
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanobiotechnol. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, Q.; Zhao, J.; Urmila, K. Enhancing the antimicrobial activity of natural extraction using the synthetic ultrasmall metal nanoparticles. Sci. Rep. 2015, 5, 11033. [Google Scholar] [CrossRef] [Green Version]
- Armentano, I.; Arciola, C.R.; Fortunati, E.; Ferrari, D.; Mattioli, S.; Amoroso, C.F.; Rizzo, J.; Kenny, J.M.; Imbriani, M.; Visai, L. The interaction of bacteria with engineered nanostructured polymeric materials: A review. Sci. World J. 2014, 2014, 410423. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Thamphiwatana, S.; Angsantikul, P.; Zhang, L. Nanoparticle approaches against bacterial infections. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2014, 6, 532–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.T.; Musarrat, J.; Al-Khedhairy, A.A. Countering drug resistance, infectious diseases, and sepsis using metal and metal oxides nanoparticles: Current status. Colloids Surf. B Biointerfaces 2016, 146, 70–83. [Google Scholar] [CrossRef]
- Zaidi, S.; Misba, L.; Khan, A.U. Nano-therapeutics: A revolution in infection control in post antibiotic era. Nanomedicine 2017, 13, 2281–2301. [Google Scholar] [CrossRef]
- Dizaj, S.M.; Lotfipour, F.; Barzegar-Jalali, M.; Zarrintan, M.H.; Adibkia, K. Antimicrobial activity of the metals and metal oxide nanoparticles. Mater. Sci. Eng. C 2014, 44, 278–284. [Google Scholar] [CrossRef]
- Yang, W.; Shen, C.; Ji, Q.; An, H.; Wang, J.; Liu, Q.; Zhang, Z. Food storage material silver nanoparticles interfere with DNA replication fidelity and bind with DNA. Nanotechnology 2009, 20, 085102. [Google Scholar] [CrossRef]
- Beyth, N.; Houri-Haddad, Y.; Domb, A.; Khan, W.; Hazan, R. Alternative antimicrobial approach: Nano-antimicrobial materials. Evid. -Based Complement. Altern. Med. 2015, 2015, 246012. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Bulut, O.; Some, S.; Mandal, A.K.; Yilmaz, M.D. Green synthesis of silver nanoparticles: Biomolecule-nanoparticle organizations targeting antimicrobial activity. RSC Adv 2019, 9, 2673–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, M.; Khan, H.; Khan, A.; Cameotra, S.S.; Saquib, Q.; Musarrat, J. Interaction of A l2 O 3 nanoparticles with E scherichia coli and their cell envelope biomolecules. J. Appl. Microbiol. 2014, 116, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Baptista, P.V.; McCusker, M.P.; Carvalho, A.; Ferreira, D.A.; Mohan, N.M.; Martins, M.; Fernandes, A.R. Nano-strategies to fight multidrug resistant bacteria—“A Battle of the Titans”. Front. Microbiol. 2018, 9, 1441. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Lu, J.; Xu, H.; Patel, A.; Chen, Z.-S.; Chen, G. Silver nanoparticles: Synthesis, properties, and therapeutic applications. Drug Discov. Today 2015, 20, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Jiang, X. Multiple strategies to activate gold nanoparticles as antibiotics. Nanoscale 2013, 5, 8340–8350. [Google Scholar] [CrossRef] [PubMed]
- Finley, P.J.; Norton, R.; Austin, C.; Mitchell, A.; Zank, S.; Durham, P. Unprecedented silver resistance in clinically isolated Enterobacteriaceae: Major implications for burn and wound management. Antimicrob. Agents Chemother. 2015, 59, 4734–4741. [Google Scholar] [CrossRef] [Green Version]
- Barros, C.H.; Fulaz, S.; Stanisic, D.; Tasic, L. Biogenic nanosilver against multidrug-resistant bacteria (MDRB). Antibiotics 2018, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Huang, R.; Sahu, M.; Feng, X.; Biswas, P.; Tang, Y.J. Bacterial responses to Cu-doped TiO2 nanoparticles. Sci. Total Environ. 2010, 408, 1755–1758. [Google Scholar] [CrossRef]
- Cavassin, E.D.; de Figueiredo, L.F.P.; Otoch, J.P.; Seckler, M.M.; de Oliveira, R.A.; Franco, F.F.; Marangoni, V.S.; Zucolotto, V.; Levin, A.S.S.; Costa, S.F. Comparison of methods to detect the in vitro activity of silver nanoparticles (AgNP) against multidrug resistant bacteria. J. Nanobiotechnol. 2015, 13, 64. [Google Scholar] [CrossRef]
- Chatterjee, A.K.; Chakraborty, R.; Basu, T. Mechanism of antibacterial activity of copper nanoparticles. Nanotechnology 2014, 25, 135101. [Google Scholar] [CrossRef]
- Vandebriel, R.J.; De Jong, W.H. A review of mammalian toxicity of ZnO nanoparticles. Nanotechnol. Sci. Appl. 2012, 5, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic Acids and Potential for Modifying the Avian Gastrointestinal Tract and Reducing Pathogens and Disease. Front. Vet. Sci. 2018, 5, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coban, H.B. Organic acids as antimicrobial food agents: Applications and microbial productions. Bioprocess Biosyst. Eng. 2020, 43, 569–591. [Google Scholar] [CrossRef]
- Davidson, P.M.; Sofos, J.N.; Branen, A.L. Antimicrobials in Food; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Polen, T.; Spelberg, M.; Bott, M. Toward biotechnological production of adipic acid and precursors from biorenewables. J. Biotechnol. 2013, 167, 75–84. [Google Scholar] [CrossRef]
- Blach, P.; Böstrom, Z.; Franceschi-Messant, S.; Lattes, A.; Perez, E.; Rico-Lattes, I. Recyclable process for sustainable adipic acid production in microemulsions. Tetrahedron 2010, 66, 7124–7128. [Google Scholar] [CrossRef]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Citric acid: Emerging applications of key biotechnology industrial product. Chem. Cent. J. 2017, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: Overview and limits. J. Biotechnol. 2011, 156, 286–301. [Google Scholar] [CrossRef]
- Marques, C.; Sotiles, A.R.; Farias, F.O.; Oliveira, G.; Mitterer-Daltoé, M.L.; Masson, M.L. Full physicochemical characterization of malic acid: Emphasis in the potential as food ingredient and application in pectin gels. Arab. J. Chem. 2020, 13, 9118–9129. [Google Scholar] [CrossRef]
- Rodríguez-Pazo, N.; Salgado, J.M.; Cortés-Diéguez, S.; Domínguez, J.M. Biotechnological production of phenyllactic acid and biosurfactants from trimming vine shoot hydrolyzates by microbial coculture fermentation. Appl. Biochem. Biotechnol. 2013, 169, 2175–2188. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Roohi, R. Kinetic models for production of propionic acid by Propionibacter freudenrechii subsp. shermanii and Propionibacterium freudenreichii subsp. freudenreichii in date syrup during sonication treatments. Biocatal. Agric. Biotechnol. 2019, 17, 613–619. [Google Scholar] [CrossRef]
- Galli, G.M.; Aniecevski, E.; Petrolli, T.G.; da Rosa, G.; Boiago, M.M.; Simões, C.A.D.P.; Wagner, R.; Copetti, P.M.; Morsch, V.M.; Araujo, D.N.; et al. Growth performance and meat quality of broilers fed with microencapsulated organic acids. Anim. Feed. Sci. Technol. 2021, 271, 114706. [Google Scholar] [CrossRef]
- Kuenz, A.; Hoffmann, L.; Goy, K.; Bromann, S.; Prüße, U. High-level production of succinic acid from crude glycerol by a wild type organism. Catalysts 2020, 10, 470. [Google Scholar] [CrossRef]
- Brul, S.; Coote, P. Preservative agents in foods. Mode of action and microbial resistance mechanisms. Int. J. Food Microbiol. 1999, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Theron, M.M.; Lues, J.F.R. Organic Acids and Meat Preservation: A Review. Food Rev. Int. 2007, 23, 141–158. [Google Scholar] [CrossRef]
- Stratford, M.; Anslow, P.A. Evidence that sorbic acid does not inhibit yeast as a classic ‘weak acid preservative’. Lett. Appl. Microbiol. 1998, 27, 203–206. [Google Scholar] [CrossRef]
- Kundukad, B.; Udayakumar, G.; Grela, E.; Kaur, D.; Rice, S.A.; Kjelleberg, S.; Doyle, P.S. Weak acids as an alternative anti-microbial therapy. Biofilm 2020, 2, 100019. [Google Scholar] [CrossRef]
- Ben Braïek, O.; Smaoui, S. Chemistry, Safety, and Challenges of the Use of Organic Acids and Their Derivative Salts in Meat Preservation. J. Food Qual. 2021, 2021, 6653190. [Google Scholar] [CrossRef]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Hauser, C.; Thielmann, J.; Muranyi, P. Chapter 46—Organic Acids: Usage and Potential in Antimicrobial Packaging. In Antimicrobial Food Packaging; Barros-Velázquez, J., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 563–580. [Google Scholar]
- Hajati, H. Application of organic acids in poultry nutrition. Int. J. Avian Wildl. Biol. 2018, 3, 324–329. [Google Scholar] [CrossRef] [Green Version]
- Flors, V.; Miralles, M.C.; Varas, E.; Company, P.; González-Bosch, C.; García-Agustín, P. Effect of analogues of plant growth regulators on in vitro growth of eukaryotic plant pathogens. Plant Pathol. 2004, 53, 58–64. [Google Scholar] [CrossRef]
- Fernández-Rubio, C.; Ordóñez, C.; Abad-González, J.; Garcia-Gallego, A.; Honrubia, M.P.; Mallo, J.J.; Balaña-Fouce, R. Butyric acid-based feed additives help protect broiler chickens from Salmonella Enteritidis infection. Poult. Sci. 2009, 88, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambabu, K.; Bharath, G.; Banat, F.; Show, P.L.; Cocoletzi, H.H. Mango leaf extract incorporated chitosan antioxidant film for active food packaging. Int. J. Biol. Macromol. 2019, 126, 1234–1243. [Google Scholar] [CrossRef]
- Show, P.L.; Oladele, K.O.; Siew, Q.Y.; Aziz Zakry, F.A.; Lan, J.C.-W.; Ling, T.C. Overview of citric acid production from Aspergillus niger. Front. Life Sci. 2015, 8, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Laury, A.M.; Alvarado, M.V.; Nace, G.; Alvarado, C.Z.; Brooks, J.C.; Echeverry, A.; Brashears, M.M. Validation of a lactic acid- and citric acid-based antimicrobial product for the reduction of Escherichia coli O157: H7 and Salmonella on beef tips and whole chicken carcasses. J. Food Prot. 2009, 72, 2208–2211. [Google Scholar] [CrossRef]
- Beuchat, L.R. Influence of organic acids on heat resistance characteristics of Talaromyces flavus ascospores. Int. J. Food Microbiol. 1988, 6, 97–105. [Google Scholar] [CrossRef]
- Zhang, S.; Farber, J.M. The effects of various disinfectants againstListeria monocytogeneson fresh-cut vegetables. Food Microbiol. 1996, 13, 311–321. [Google Scholar] [CrossRef]
- Zou, X.; Zhou, Y.; Yang, S.T. Production of polymalic acid and malic acid by Aureobasidium pullulans fermentation and acid hydrolysis. Biotechnol. Bioeng. 2013, 110, 2105–2113. [Google Scholar] [CrossRef]
- Beauprez, J.J.; De Mey, M.; Soetaert, W.K. Microbial succinic acid production: Natural versus metabolic engineered producers. Process Biochem. 2010, 45, 1103–1114. [Google Scholar] [CrossRef]
- Li, C.; Yang, X.; Gao, S.; Chuh, A.H.; Lin, C.S.K. Hydrolysis of fruit and vegetable waste for efficient succinic acid production with engineered Yarrowia lipolytica. J. Clean. Prod. 2018, 179, 151–159. [Google Scholar] [CrossRef]
- Over, K.F.; Hettiarachchy, N.; Johnson, M.G.; Davis, B. Effect of organic acids and plant extracts on Escherichia coli O157:H7, Listeria monocytogenes, and Salmonella Typhimurium in broth culture model and chicken meat systems. J. Food Sci. 2009, 74, M515–M521. [Google Scholar] [CrossRef] [PubMed]
- Kovanda, L.; Zhang, W.; Wei, X.; Luo, J.; Wu, X.; Atwill, E.R.; Vaessen, S.; Li, X.; Liu, Y. In vitro antimicrobial activities of organic acids and their derivatives on several species of gram-negative and gram-positive bacteria. Molecules 2019, 24, 3770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed. Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Stefanakis, M.K.; Touloupakis, E.; Anastasopoulos, E.; Ghanotakis, D.; Katerinopoulos, H.E.; Makridis, P. Antibacterial activity of essential oils from plants of the genus Origanum. Food Control 2013, 34, 539–546. [Google Scholar] [CrossRef]
- Djilani, A.; Dicko, A.J.N. The therapeutic benefits of essential oils. Nutr. Well-Being Health 2012, 7, 155–179. [Google Scholar]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils-Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, H.A.E.; El-Ghorab, A.H.; Shibamoto, T. Bioactivity of essential oils and their volatile aroma components: Review. J. Essent. Oil Res. 2012, 24, 203–212. [Google Scholar] [CrossRef]
- Nikolić, M.; Jovanović, K.K.; Marković, T.; Marković, D.; Gligorijević, N.; Radulović, S.; Soković, M. Chemical composition, antimicrobial, and cytotoxic properties of five Lamiaceae essential oils. Ind. Crops Prod. 2014, 61, 225–232. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef]
- Verma, S.K.; Goswami, P.; Verma, R.S.; Padalia, R.C.; Chauhan, A.; Singh, V.R.; Darokar, M.P. Chemical composition and antimicrobial activity of bergamot-mint (Mentha citrata Ehrh.) essential oils isolated from the herbage and aqueous distillate using different methods. Ind. Crops Prod. 2016, 91, 152–160. [Google Scholar] [CrossRef]
- Radaelli, M.; da Silva, B.P.; Weidlich, L.; Hoehne, L.; Flach, A.; da Costa, L.A.; Ethur, E.M. Antimicrobial activities of six essential oils commonly used as condiments in Brazil against Clostridium perfringens. Braz. J. Microbiol. 2016, 47, 424–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajer, T.; Šilha, D.; Ventura, K.; Bajerová, P. Composition and antimicrobial activity of the essential oil, distilled aromatic water and herbal infusion from Epilobium parviflorum Schreb. Ind. Crops Prod. 2017, 100, 95–105. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control 2016, 59, 282–289. [Google Scholar] [CrossRef]
- El Kolli, M.; Laouer, H.; El Kolli, H.; Akkal, S.; Sahli, F. Chemical analysis, antimicrobial and anti-oxidative properties of Daucus gracilis essential oil and its mechanism of action. Asian Pac. J. Trop. Biomed. 2016, 6, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Churklam, W.; Chaturongakul, S.; Ngamwongsatit, B.; Aunpad, R. The mechanisms of action of carvacrol and its synergism with nisin against Listeria monocytogenes on sliced bologna sausage. Food Control 2020, 108, 106864. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- da Silva, B.D.; Bernardes, P.C.; Pinheiro, P.F.; Fantuzzi, E.; Roberto, C.D. Chemical composition, extraction sources and action mechanisms of essential oils: Natural preservative and limitations of use in meat products. Meat Sci. 2021, 176, 108463. [Google Scholar] [CrossRef]
- Ait-Ouazzou, A.; Cherrat, L.; Espina, L.; Lorán, S.; Rota, C.; Pagán, R. The antimicrobial activity of hydrophobic essential oil constituents acting alone or in combined processes of food preservation. Innov. Food Sci. Emerg. Technol. 2011, 12, 320–329. [Google Scholar] [CrossRef]
- Gupta, S.; Allen-Vercoe, E.; Petrof, E.O. Fecal microbiota transplantation: In perspective. Ther. Adv. Gastroenterol. 2016, 9, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.; Kumar, R.; Sharma, S.; Mahanta, M.; Vayuuru, S.K.; Nayak, B.; Kumar, S. Fecal microbiota transplantation in Hepatitis B e antigen-positive chronic Hepatitis B patients: A pilot study. Dig. Dis. Sci. 2021, 66, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Ueckermann, V.; Hoosien, E.; De Villiers, N.; Geldenhuys, J. Fecal microbial transplantation for the treatment of persistent multidrug-resistant Klebsiella pneumoniae infection in a critically ill patient. Case Rep. Infect. Dis. 2020, 2020, 8462659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hourigan, S.K.; Ahn, M.; Gibson, K.M.; Pérez-Losada, M.; Felix, G.; Weidner, M.; Leibowitz, I.; Niederhuber, J.E.; Sears, C.L.; Crandall, K.A. Fecal transplant in children with Clostridioides difficile gives sustained reduction in antimicrobial resistance and potential pathogen burden. Open Forum Infect. Dis. 2019, 6, ofz379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xing, Y.; Ji, Y.; Xi, H.; Liu, X.; Yang, L.; Lei, L.; Han, W.; Gu, J. The Combination of Phages and Faecal Microbiota Transplantation Can Effectively Treat Mouse Colitis Caused by Salmonella enterica Serovar Typhimurium. Front. Microbiol. 2022, 13, 944495. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Gong, J.; Zhu, W.; Guo, D.; Gu, L.; Li, N.; Li, J. Fecal microbiota transplantation restores dysbiosis in patients with methicillin resistant Staphylococcus aureus enterocolitis. BMC Infect. Dis. 2015, 15, 265. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; de Groot, P.F.; Geerlings, S.E.; Hodiamont, C.J.; Belzer, C.; Berge, I.; de Vos, W.M.; Bemelman, F.J.; Nieuwdorp, M. Fecal microbiota transplantation against intestinal colonization by extended spectrum beta-lactamase producing Enterobacteriaceae: A proof of principle study. BMC Res Notes 2018, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Mańkowska-Wierzbicka, D.; Stelmach-Mardas, M.; Gabryel, M.; Tomczak, H.; Skrzypczak-Zielińska, M.; Zakerska-Banaszak, O.; Sowińska, A.; Mahadea, D.; Baturo, A.; Wolko, Ł. The effectiveness of multi-session FMT treatment in active ulcerative colitis patients: A pilot study. Biomedicines 2020, 8, 268. [Google Scholar] [CrossRef]
- Paramsothy, S.; Paramsothy, R.; Rubin, D.T.; Kamm, M.A.; Kaakoush, N.O.; Mitchell, H.M.; Castaño-Rodríguez, N. Faecal microbiota transplantation for inflammatory bowel disease: A systematic review and meta-analysis. J. Crohn’s Colitis 2017, 11, 1180–1199. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Madsen, K.L. Impact of fecal microbiota transplantation on obesity and metabolic syndrome—A systematic review. Nutrients 2019, 11, 2291. [Google Scholar] [CrossRef] [Green Version]
- Kragsnaes, M.S.; Kjeldsen, J.; Horn, H.C.; Munk, H.L.; Pedersen, F.M.; Holt, H.M.; Pedersen, J.K.; Holm, D.K.; Glerup, H.; Andersen, V. Efficacy and safety of faecal microbiota transplantation in patients with psoriatic arthritis: Protocol for a 6-month, double-blind, randomised, placebo-controlled trial. BMJ Open 2018, 8, e019231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Angulo, M.T.; Lao, S.; Weiss, S.T.; Liu, Y.-Y. An ecological framework to understand the efficacy of fecal microbiota transplantation. Nat. Commun. 2020, 11, 3329. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Ren, K.; Ning, R.; Li, C.; Zhang, H.; Li, D.; Xu, L.; Sun, F.; Dai, M. Fecal microbiota transplantation provides new insight into wildlife conservation. Glob. Ecol. Conserv. 2020, 24, e01234. [Google Scholar] [CrossRef]
- Khoruts, A.; Sadowsky, M.J. Understanding the mechanisms of faecal microbiota transplantation. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, L.J.; Aroniadis, O.C.; Mellow, M.; Kanatzar, A.; Kelly, C.; Park, T.; Stollman, N.; Rohlke, F.; Surawicz, C. Long-Term Follow-Up of Colonoscopic Fecal Microbiota Transplant for RecurrentClostridium difficileInfection. Off. J. Am. Coll. Gastroenterol. ACG 2012, 107, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Leshem, A.; Horesh, N.; Elinav, E. Fecal microbial transplantation and its potential application in cardiometabolic syndrome. Front. Immunol. 2019, 10, 1341. [Google Scholar] [CrossRef] [Green Version]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojärvi, J.; Voigt, A.Y. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef]
- Li, N.; Tian, H.; Ma, C.; Ding, C.; Ge, X.; Gu, L.; Zhang, X.; Yang, B.; Hua, Y.; Zhu, Y. Efficacy analysis of fecal microbiota transplantation in the treatment of 406 cases with gastrointestinal disorders. Zhonghua Wei Chang. Wai Ke Za Zhi Chin. J. Gastrointest. Surg. 2017, 20, 40–46. [Google Scholar]
- Gweon, T.-G.; Na, S.-Y. Next generation fecal microbiota transplantation. Clin. Endosc. 2021, 54, 152. [Google Scholar] [CrossRef]
- Allegretti, J.R.; Elliott, R.J.; Ladha, A.; Njenga, M.; Warren, K.; O’Brien, K.; Budree, S.; Osman, M.; Fischer, M.; Kelly, C.R. Stool processing speed and storage duration do not impact the clinical effectiveness of fecal microbiota transplantation. Gut Microbes 2020, 11, 1806–1808. [Google Scholar] [CrossRef]
- Greenwood, B. The contribution of vaccination to global health: Past, present and future. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369, 20130433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elaish, M.; Ngunjiri, J.M.; Ali, A.; Xia, M.; Ibrahim, M.; Jang, H.; Hiremath, J.; Dhakal, S.; Helmy, Y.A.; Jiang, X.; et al. Supplementation of inactivated influenza vaccine with norovirus P particle-M2e chimeric vaccine enhances protection against heterologous virus challenge in chickens. PLoS ONE 2017, 12, e0171174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawzy, M.; Helmy, Y.A. The One Health Approach is Necessary for the Control of Rift Valley Fever Infections in Egypt: A Comprehensive Review. Viruses 2019, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Hoelzer, K.; Bielke, L.; Blake, D.P.; Cox, E.; Cutting, S.M.; Devriendt, B.; Erlacher-Vindel, E.; Goossens, E.; Karaca, K.; Lemiere, S.; et al. Vaccines as alternatives to antibiotics for food producing animals. Part 2: New approaches and potential solutions. Vet. Res. 2018, 49, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klugman, K.P.; Black, S. Impact of existing vaccines in reducing antibiotic resistance: Primary and secondary effects. Proc. Natl. Acad. Sci. USA 2018, 115, 12896–12901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The role of vaccines in combatting antimicrobial resistance. Nat. Rev. Microbiol. 2021, 19, 287–302. [Google Scholar] [CrossRef]
- Kennedy, D.A.; Read, A.F. Why does drug resistance readily evolve but vaccine resistance does not? Proc. Biol. Sci. 2017, 284, 20162562. [Google Scholar] [CrossRef] [Green Version]
- Sihvonen, R.; Siira, L.; Toropainen, M.; Kuusela, P.; Patari-Sampo, A. Streptococcus pneumoniae antimicrobial resistance decreased in the Helsinki Metropolitan Area after routine 10-valent pneumococcal conjugate vaccination of infants in Finland. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2109–2116. [Google Scholar] [CrossRef]
- Klugman, K.P.; Madhi, S.A.; Huebner, R.E.; Kohberger, R.; Mbelle, N.; Pierce, N.; Grp, V.T. A trial of a 9-valent pneumococcal conjugate vaccine in children with and those without HIV infection. N. Engl. J. Med. 2003, 349, 1341–1348. [Google Scholar] [CrossRef]
- WHO. Pakistan First Country to Introduce New Typhoid Vaccine into Routine Immunization Programme; WHO: Geneva, Switzerland, 2019.
- Kaufhold, S.; Yaesoubi, R.; Pitzer, V.E. Predicting the Impact of Typhoid Conjugate Vaccines on Antimicrobial Resistance. Clin. Infect. Dis. 2019, 68, S96–S104. [Google Scholar] [CrossRef] [Green Version]
- House, J.K.; Ontiveros, M.M.; Blackmer, N.M.; Dueger, E.L.; Fitchhorn, J.B.; McArthur, G.R.; Smith, B.P. Evaluation of an autogenous Salmonella bacterin and a modified live Salmonella serotype Choleraesuis vaccine on a commercial dairy farm. Am. J. Vet. Res. 2001, 62, 1897–1902. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.R.; Friedman, C.R.; Crim, S.M.; Judd, M.; Barrett, K.A.; Tolar, B.; Folster, J.P.; Griffin, P.M.; Brown, A.C. Epidemiology of Salmonella enterica Serotype Dublin Infections among Humans, United States, 1968–2013. Emerg. Infect. Dis. 2017, 23, 1493–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, D.V.T.; Venkitanarayanan, K.; Kollanoo, J.A. Antibiotic-Resistant Salmonella in the Food Supply and the Potential Role of Antibiotic Alternatives for Control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, K.J.; Rodriguez-Rivera, L.D.; Capel, M.B.; Rankin, S.C.; Nydam, D.V. Short communication: Oral and intranasal administration of a modified-live Salmonella Dublin vaccine in dairy calves: Clinical efficacy and serologic response. J. Dairy Sci. 2019, 102, 3474–3479. [Google Scholar] [CrossRef] [Green Version]
- Hosomi, K.; Hinenoya, A.; Suzuki, H.; Nagatake, T.; Nishino, T.; Tojima, Y.; Hirata, S.-I.; Matsunaga, A.; Kondoh, M.; Yamasaki, S.; et al. Development of a bivalent food poisoning vaccine: Augmented antigenicity of the C-terminus of Clostridium perfringens enterotoxin by fusion with the B subunit of Escherichia coli Shiga toxin 2. Int. Immunol. 2019, 31, 91–100. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Alkie, T.N.; Hodgins, D.C.; Yitbarek, A.; Shojadoost, B.; Sharif, S. Gene expression profiling of chicken cecal tonsils and ileum following oral exposure to soluble and PLGA-encapsulated CpG ODN, and lysate of Campylobacter jejuni. Vet. Microbiol. 2017, 212, 67–74. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Yitbarek, A.; Alkie, T.N.; Hodgins, D.C.; Read, L.R.; Weese, J.S.; Sharif, S. PLGA-encapsulated CpG ODN and Campylobacter jejuni lysate modulate cecal microbiota composition in broiler chickens experimentally challenged with C. jejuni. Sci. Rep. 2018, 8, 12076. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, M.; Shojadoost, B.; Boodhoo, N.; Astill, J.; Taha-Abdelaziz, K.; Hodgins, D.C.; Kulkarni, R.R.; Sharif, S. Necrotic enteritis in chickens: A review of pathogenesis, immune responses and prevention, focusing on probiotics and vaccination. Anim. Health Res. Rev. 2021, 22, 147–162. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Hodgins, D.C.; Alkie, T.N.; Quinteiro-Filho, W.; Yitbarek, A.; Astill, J.; Sharif, S. Oral administration of PLGA-encapsulated CpG ODN and Campylobacter jejuni lysate reduces cecal colonization by Campylobacter jejuni in chickens. Vaccine 2018, 36, 388–394. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Singh, M.; Sharif, S.; Sharma, S.; Kulkarni, R.R.; Alizadeh, M.; Yitbarek, A.; Helmy, Y.A. Intervention Strategies to Control Campylobacter at Different Stages of the Food Chain. Microorganisms 2023, 11, 113. [Google Scholar] [CrossRef]
- Nothaft, H.; Perez-Muñoz, M.E.; Gouveia, G.J.; Duar, R.M.; Wanford, J.J.; Lango-Scholey, L.; Panagos, C.G.; Srithayakumar, V.; Plastow, G.S.; Coros, C.; et al. Coadministration of the Campylobacter jejuni N-Glycan-Based Vaccine with Probiotics Improves Vaccine Performance in Broiler Chickens. Appl. Environ. Microbiol. 2017, 83, e01523-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchy, P.; Ascioglu, S.; Buisson, Y.; Datta, S.; Nissen, M.; Tambyah, P.A.; Vong, S. Impact of vaccines on antimicrobial resistance. Int. J. Infect. Dis. 2020, 90, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.P.; Oviedo-Orta, E.; Prachi, P.; Rappuoli, R.; Bagnoli, F. Vaccines and antibiotic resistance. Curr. Opin. Microbiol. 2012, 15, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Poolman, J.T. Expanding the role of bacterial vaccines into life-course vaccination strategies and prevention of antimicrobial-resistant infections. NPJ Vaccines 2020, 5, 84. [Google Scholar] [CrossRef] [PubMed]
- Tekle, Y.I.; Nielsen, K.M.; Liu, J.; Pettigrew, M.M.; Meyers, L.A.; Galvani, A.P.; Townsend, J.P. Controlling antimicrobial resistance through targeted, vaccine-induced replacement of strains. PLoS ONE 2012, 7, e50688. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.K.; Yadav, N.; Khurana, S.M.P. Vaccines. In Animal Biotechnology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 491–508. [Google Scholar]
- Mak, T.; Saunders, M.; Jett, B. Vaccination. In Primer to the Immune Response; Elsevier: Amsterdam, The Netherlands, 2014; pp. 333–375. [Google Scholar]
- Parham, P.; Janeway, C. The Immune System, 4th ed.; Garland Science, Taylor & Francis Group: New York, NY, USA, 2015; p. 1. [Google Scholar]
- Lidder, P.; Sonnino, A. Biotechnologies for the Management of Genetic Resources for Food and Agriculture. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 2012; Volume 78, pp. 1–167. [Google Scholar]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef]
- Rappuoli, R. Glycoconjugate vaccines: Principles and mechanisms. Sci. Transl. Med. 2018, 10, eaat4615. [Google Scholar] [CrossRef]
- Ura, T.; Okuda, K.; Shimada, M. Developments in Viral Vector-Based Vaccines. Vaccines 2014, 2, 624–641. [Google Scholar] [CrossRef] [Green Version]
- Gheibi Hayat, S.M.; Darroudi, M. Nanovaccine: A novel approach in immunization. J. Cell. Physiol. 2019, 234, 12530–12536. [Google Scholar] [CrossRef]
- Lyon, C.E.; Sadigh, K.S.; Carmolli, M.P.; Harro, C.; Sheldon, E.; Lindow, J.C.; Larsson, C.J.; Martinez, T.; Feller, A.; Ventrone, C.H.; et al. In a randomized, double-blinded, placebo-controlled trial, the single oral dose typhoid vaccine, M01ZH09, is safe and immunogenic at doses up to 1.7×1010 colony-forming units. Vaccine 2010, 28, 3602–3608. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Bronzo, V.; Locatelli, C.; Pollera, C.; Rota, N.; Casula, A.; Testa, F.; Scaccabarozzi, L.; March, R.; Zalduendo, D.; et al. Efficacy of vaccination on Staphylococcus aureus and coagulase-negative staphylococci intramammary infection dynamics in 2 dairy herds. J. Dairy Sci. 2014, 97, 5250–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, J.R.; Ma, J.; Rinehart, C.L.; Taylor, V.N.; Luby, C.D.; Steevens, B.J. Efficacy of different Lysigin™ formulations in the prevention of Staphylococcus aureus intramammary infection in dairy heifers. J. Dairy Res. 2006, 73, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.W.; Smith, F.; Zuidhof, S.; Foster, D.M. Short communication: Characterization of the serologic response induced by vaccination of late-gestation cows with a Salmonella Dublin vaccine. J. Dairy Sci. 2015, 98, 2529–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, C.F.; Withanage, G.S.K.; de Haas, V.; Etore, F.; Francis, M.J. Safety and efficacy of a maternal vaccine for the passive protection of broiler chicks against necrotic enteritis. Avian Pathol. 2010, 39, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Huberman, Y.D.; Velilla, A.V.; Terzolo, H.R. Evaluation of different live Salmonella enteritidis vaccine schedules administered during layer hen rearing to reduce excretion, organ colonization, and egg contamination. Poult. Sci. 2019, 98, 2422–2431. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Wiedermann, U.; Garner-Spitzer, E.; Wagner, A. Primary vaccine failure to routine vaccines: Why and what to do? Hum. Vaccines Immunother. 2016, 12, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Moyle, P.M.; Toth, I. Modern Subunit Vaccines: Development, Components, and Research Opportunities. ChemMedChem 2013, 8, 360–376. [Google Scholar] [CrossRef]
- Deng, F. Advances and challenges in enveloped virus-like particle (VLP)-based vaccines. J. Immunol. Sci. 2018, 2, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Rossi, O.; Citiulo, F.; Mancini, F. Outer membrane vesicles: Moving within the intricate labyrinth of assays that can predict risks of reactogenicity in humans. Hum. Vaccines Immunother. 2021, 17, 601–613. [Google Scholar] [CrossRef]
- Mettu, R.; Chen, C.-Y.; Wu, C.-Y. Synthetic carbohydrate-based vaccines: Challenges and opportunities. J. Biomed. Sci. 2020, 27, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Chaudhary, S.; Agarwal, A.; Verma, A.S. Antibodies. In Animal Biotechnology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 265–287. [Google Scholar]
- Liddell, E. Antibodies. In The Immunoassay Handbook; Elsevier: Amsterdam, The Netherlands, 2013; pp. 245–265. [Google Scholar]
- van Erp, E.A.; Luytjes, W.; Ferwerda, G.; van Kasteren, P.B. Fc-Mediated Antibody Effector Functions During Respiratory Syncytial Virus Infection and Disease. Front. Immunol. 2019, 10, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiGiandomenico, A.; Sellman, B.R. Antibacterial monoclonal antibodies: The next generation? Curr. Opin. Microbiol. 2015, 27, 78–85. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J. Where are we with monoclonal antibodies for multidrug-resistant infections? Drug Discov. Today 2019, 24, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, M.; Castanho, M.A.R.B.; Neves, V. The Use of Antibody-Antibiotic Conjugates to Fight Bacterial Infections. Front. Microbiol. 2022, 13, 835677. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T. Therapeutic monoclonal antibodies. Keio J. Med. 2011, 60, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Reichert, J.M.; Dewitz, M.C. Ooutlook—Anti-infective monoclonal antibodies: Perils and promise of development. Nat. Rev. Drug Discov. 2006, 5, 191–195. [Google Scholar] [CrossRef]
- Chan, C.E.Z.; Chan, A.H.Y.; Hanson, B.J.; Ooi, E.E. The use of antibodies in the treatment of infectious diseases. Singap. Med. J. 2009, 50, 663–672. [Google Scholar]
- Kohler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity (Reprinted from Nature, vol 256, 1975). J. Immunol. 2005, 174, 2453–2455. [Google Scholar]
- Pelfrene, E.; Mura, M.; Cavaleiro Sanches, A.; Cavaleri, M. Monoclonal antibodies as anti-infective products: A promising future? Clin. Microbiol. Infect. 2019, 25, 60–64. [Google Scholar] [CrossRef]
- Zurawski, D.V.; McLendon, M.K. Monoclonal Antibodies as an Antibacterial Approach Against Bacterial Pathogens. Antibiotics 2020, 9, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, E.; Nagy, G.; Power, C.A.; Badarau, A.; Szijarto, V. Anti-bacterial Monoclonal Antibodies. Recomb. Antibodies Infect. Dis. 2017, 1053, 119–153. [Google Scholar]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond binding: Antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.; Rothenberg, M.E.; Deng, R.; Lewin-Koh, N.; She, G.; Kamath, A.V.; Carrasco-Triguero, M.; Saad, O.; Castro, A.; Teufel, L.; et al. A Phase 1, Randomized, Single-Ascending-Dose Study To Investigate the Safety, Tolerability, and Pharmacokinetics of DSTA4637S, an Anti- Staphylococcus aureus Thiomab Antibody-Antibiotic Conjugate, in Healthy Volunteers. Antimicrob. Agents Chemother. 2019, 63, e02588-18. [Google Scholar] [CrossRef] [Green Version]
- Guptill, J.T.; Raja, S.M.; Juel, V.C.; Walter, E.B.; Cohen-Wolkowiez, M.; Hill, H.; Sendra, E.; Hauser, B.; Jackson, P.; Swamy, G.K. Safety, Tolerability, and Pharmacokinetics of NTM-1632, a Novel Mixture of Three Monoclonal Antibodies against Botulinum Toxin B. Antimicrob. Agents Chemother. 2021, 65, e02329-20. [Google Scholar] [CrossRef] [PubMed]
- Greig, S.L. Obiltoxaximab: First Global Approval. Drugs 2016, 76, 823–830. [Google Scholar] [CrossRef]
- Mazumdar, S. Raxibacumab. mAbs 2009, 1, 531–538. [Google Scholar] [CrossRef]
- Weisman, L.E.; Thackray, H.M.; Garcia-Prats, J.A.; Nesin, M.; Schneider, J.H.; Fretz, J.; Kokai-Kun, J.F.; Mond, J.J.; Kramer, W.G.; Fischer, G.W. Phase 1/2 Double-Blind, Placebo-Controlled, Dose Escalation, Safety, and Pharmacokinetic Study of Pagibaximab (BSYX-A110), an Antistaphylococcal Monoclonal Antibody for the Prevention of Staphylococcal Bloodstream Infections, in Very-Low-Birth-Weight Neonates. Antimicrob. Agents Chemother. 2009, 53, 2879–2886. [Google Scholar] [CrossRef] [Green Version]
- Soon, T.L. Recombinant Antibodies for Infectious Diseases; Springer Berlin Heidelberg: New York, NY, USA, 2018. [Google Scholar]
- Merakou, C.; Schaefers, M.M.; Priebe, G.P. Progress Toward the Elusive Pseudomonas aeruginosa Vaccine. Surg. Infect. 2018, 19, 757–768. [Google Scholar] [CrossRef]
- Sousa, S.A.; Seixas, A.M.M.; Marques, J.M.M.; Leitão, J.H. Immunization and Immunotherapy Approaches against Pseudomonas aeruginosa and Burkholderia cepacia Complex Infections. Vaccines 2021, 9, 670. [Google Scholar] [CrossRef]
- Chames, P.; Van Regenmortel, M.; Weiss, E.; Baty, D. Therapeutic antibodies: Successes, limitations and hopes for the future: Therapeutic antibodies: An update. Br. J. Pharmacol. 2009, 157, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Spadiut, O.; Capone, S.; Krainer, F.; Glieder, A.; Herwig, C. Microbials for the production of monoclonal antibodies and antibody fragments. Trends Biotechnol. 2014, 32, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaranayake, H.; Wirth, T.; Schenkwein, D.; Räty, J.K.; Ylä-Herttuala, S. Challenges in monoclonal antibody-based therapies. Ann. Med. 2009, 41, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.H. The history of monoclonal antibody development—Progress, remaining challenges and future innovations. Ann. Med. Surg. 2014, 3, 113–116. [Google Scholar] [CrossRef]
- Iannello, A.; Ahmad, A. Role of antibody-dependent cell-mediated cytotoxicity in the efficacy of therapeutic anti-cancer monoclonal antibodies. Cancer Metastasis Rev. 2005, 24, 487–499. [Google Scholar] [CrossRef] [PubMed]






| Vaccine Name | Target Pathogen | Indicated for Use in | Notable Observations | Reference |
|---|---|---|---|---|
| M01ZH09 vaccine Live attenuated S. Typhi, strain S. Typhi ZH9 | S. Typhi | Humans | Vaccine extremely well tolerated. Adverse events did not differ between cohorts or from subjects receiving placebo; M01ZH09 was highly immunogenic in all dose ranges. Serologic responses measured by S. Typhi LPS-specific IgA and IgG ELISA were seen in most volunteers at all dose levels and time points post-vaccination. | [471] |
| Whole cell S. aureus vaccine (StartVac) | S. aureus | Cattle | The 45% observed reduction in the basic reproduction ratio of S. aureus is encouraging, but highlights that vaccination is only an additional tool in the control of S. aureus infections on dairy farms. Efficacy was dependent upon the age group of the animals, where first-lactation animals showed a higher value. Compared with animals in third and higher lactation. | [472] |
| Whole cell S. aureus vaccine (Lysigin) | S. aureus | Cattle | Lower mean duration of clinical mastitis. No evidence that the vaccine reliably prevented S. aureus, but Lysigin showed a benefit in reducing the clinical severity and duration of clinical disease after challenging. | [473] |
| Modified live S. dublin vaccine (EnterVene-d) | S. enterica serotype Dublin | Cattle | The vaccine induced the immune response via stimulation of cell-mediated, humoral, and mucosal immunity. Calves that received colostrum from vaccinated cows had significantly higher S. Dublin titers compared to calves born to unvaccinated cows. | [474] |
| Alpha toxin (CPA) toxoid vaccine (NetVax) | C. perfringens | Poultry | Overall, the vaccine appeared to be safe, with no observed systemic reactions or adverse effects on performance or reproduction. Vaccination of broiler breeder hens induces the production of antibodies in the circulation of the hen, which remain at significant levels throughout the laying cycle. Antibodies are transferred from the hen to egg yolk, resulting in antibodies in the circulation of 7-day-old chicks. | [475] |
| N-glycan-based vaccine * | C. jejuni | Poultry | Reduce the cecal Campylobacter by 6 log10 | [458] |
| AviPro Megan Vac 1 | S. Typhimurium, S. Enteritidis and S. Heidelberg | Poultry | MV1 was effective at reducing cecal S. Enteritidis counts. The live attenuated vaccine had the added advantage of not persisting in the chicks. | [476] |
| PLGA-encapsulated CpG ODN and C. jejuni lysate | C. jejuni | Poultry | Reduced C. jejuni colonization by up to 2.4 log10, modulated intestinal immune responses, modulated the gut microbiome composition, enhanced the production of C. jejuni-specific antibodies | [453,454,456] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helmy, Y.A.; Taha-Abdelaziz, K.; Hawwas, H.A.E.-H.; Ghosh, S.; AlKafaas, S.S.; Moawad, M.M.M.; Saied, E.M.; Kassem, I.I.; Mawad, A.M.M. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics 2023, 12, 274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020274
Helmy YA, Taha-Abdelaziz K, Hawwas HAE-H, Ghosh S, AlKafaas SS, Moawad MMM, Saied EM, Kassem II, Mawad AMM. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics. 2023; 12(2):274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020274
Chicago/Turabian StyleHelmy, Yosra A., Khaled Taha-Abdelaziz, Hanan Abd El-Halim Hawwas, Soumya Ghosh, Samar Sami AlKafaas, Mohamed M. M. Moawad, Essa M. Saied, Issmat I. Kassem, and Asmaa M. M. Mawad. 2023. "Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens" Antibiotics 12, no. 2: 274. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020274