Squalamine and Aminosterol Mimics Inhibit the Peptidoglycan Glycosyltransferase Activity of PBP1b

 ,
,
Abstract
:1. Introduction
2. Results
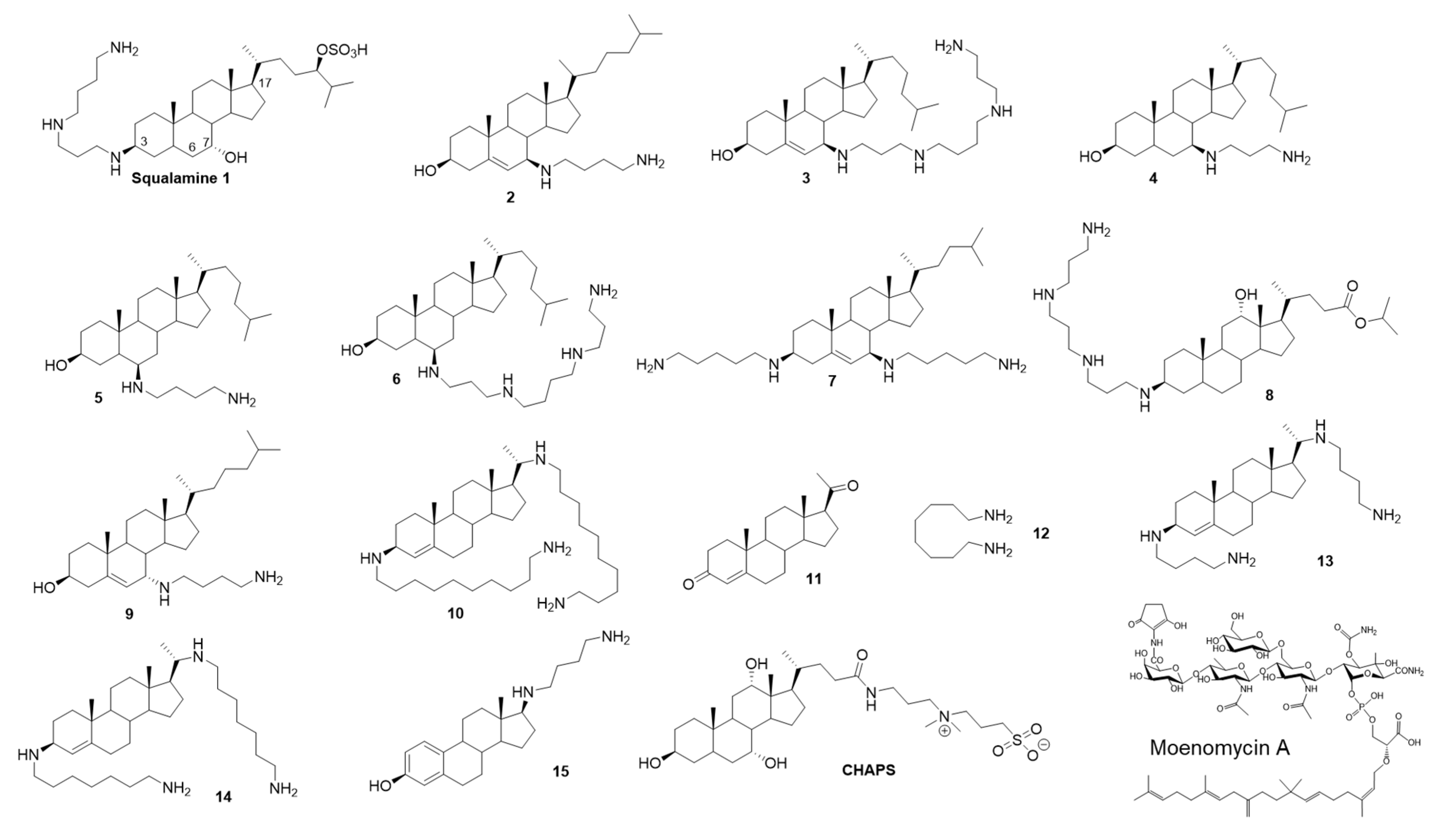
2.1. Squalamine Inhibits the GTase Activity of PBP1b
2.2. Inhibition of the GTase Activity of PBP1b by Aminosterol Mimics of Squalamine
2.3. Mode of Action of Aminosterol Compounds
3. Discussion
4. Material and Methods
4.1. Reagents and Protein
4.2. Compounds Synthesis
4.2.1. General Procedure for the Titanium–Mediated Reductive Amination Reaction
4.2.2. 6β-(1,4-diaminobutane)-cholestan-3β-ol 5
4.2.3. N1,N1’-((3S,7R,10R,13R,17R)-10,13-dimethyl-17-((R)-5-methylhexan-2-yl)-tetradecahydro-1H-cyclopenta[a]phenanthrene-3,7-diyl)bis(pentane-1,5-diamine) 7
4.2.4. (13S,17S)-17-((4-aminobutyl)amino)-13-methyl-7,8,9,11,12,13,14,15,16,17-decahydro-6H-cyclopenta[a]phenanthren-3-ol 15
4.3. GTase Activity Assays
4.4. Fluorescent Anisotropy (FA) Binding Assay
4.5. Modeling
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vollmer, W.; Blanot, D.; de Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [Green Version]
- Sauvage, E.; Kerff, F.; Terrak, M.; Ayala, J.A.; Charlier, P. The penicillin-binding proteins: Structure and role in peptidoglycan biosynthesis. FEMS Microbiol. Rev. 2008, 32, 234–258. [Google Scholar] [CrossRef] [Green Version]
- Meeske, A.J.; Riley, E.P.; Robins, W.P.; Uehara, T.; Mekalanos, J.J.; Kahne, D.; Walker, S.; Kruse, A.C.; Bernhardt, T.G.; Rudner, D.Z. SEDS proteins are a widespread family of bacterial cell wall polymerases. Nature 2016, 537, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, A.; Welsh, M.A.; Marmont, L.S.; Lee, W.; Sjodt, M.; Kruse, A.C.; Kahne, D.; Bernhardt, T.G.; Walker, S. FtsW is a peptidoglycan polymerase that is functional only in complex with its cognate penicillin-binding protein. Nat. Microbiol. 2019, 4, 587–594. [Google Scholar] [CrossRef]
- Cho, H.; Wivagg, C.N.; Kapoor, M.; Barry, Z.; Rohs, P.D.A.; Suh, H.; Marto, J.A.; Garner, E.C.; Bernhardt, T.G. Bacterial cell wall biogenesis is mediated by SEDS and PBP polymerase families functioning semi-autonomously. Nat. Microbiol. 2016, 1, 16172. [Google Scholar] [CrossRef] [PubMed]
- Sung, M.T.; Lai, Y.T.; Huang, C.Y.; Chou, L.Y.; Shih, H.W.; Cheng, W.C.; Wong, C.H.; Ma, C. Crystal structure of the membrane-bound bifunctional transglycosylase PBP1b from Escherichia coli. Proc. Natl. Acad. Sci. USA 2009, 106, 8824–8829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welzel, P. Syntheses around the transglycosylation step in peptidoglycan biosynthesis. Chem. Rev. 2005, 105, 4610–4660. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Influence of different medium components on the in vitro activity of the growth-promoting antibiotic flavomycin against enterococci. J. Antimicrob. Chemother. 2000, 46, 713–716. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.S.; Wehrli, S.; Roder, H.; Rogers, M.; Forrest, J.N.; Mccrimmon, D.; Zasloff, M. Squalamine: An aminosterol antibiotic from the shark. Proc. Natl. Acad. Sci. USA 1993, 90, 1354–1358. [Google Scholar] [CrossRef] [Green Version]
- Douafer, H.; Andrieu, V.; Phanstiel, O.; Brunel, J.M. Antibiotic adjuvants: Make antibiotics great again! J. Med. Chem. 2019, 62, 8665–8681. [Google Scholar] [CrossRef]
- Salmi, C.; Loncle, C.; Vidal, N.; Laget, M.; Letourneux, Y.; Brunel, J.M. Antimicrobial activities of 3-amino- and polyaminosterol analogues of squalamine and trodusquemine. J. Enzym. Inhib. Med. Chem. 2008, 23, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Bernard, E.M.; Sadownik, A.; Regen, S.L.; Armstrong, D. Antimicrobial activities of squalamine mimics. Antimicrob. Agents Chemother. 1997, 41, 1433–1438. [Google Scholar] [CrossRef] [Green Version]
- Djouhri-Bouktab, L.; Vidal, N.; Rolain, J.M.; Brunel, J.M. Synthesis of new 3,20-bispolyaminosteroid squalamine analogues and evaluation of their antimicrobial activities. J. Med. Chem. 2011, 54, 7417–7421. [Google Scholar] [CrossRef]
- Choucair, B.; Dherbomez, M.; Roussakis, C.; El Kihel, L. Synthesis of 7α- and 7β-spermidinylcholesterol, squalamine analogues. Bioorgan. Med. Chem. Lett. 2004, 14, 4213–4216. [Google Scholar] [CrossRef] [PubMed]
- Brycki, B.; Koenig, H.; Pospieszny, T. Quaternary alkylammonium conjugates of steroids: Synthesis, molecular structure, and biological studies. Molecules 2015, 20, 20887–20900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, P.B.; Li, C.; Taotafa, U.; Ding, B.; Guan, Q. Antibacterial properties of cationic steroid antibiotics. FEMS Microbiol. Lett. 2002, 217, 1–7. [Google Scholar] [CrossRef]
- Di Pasquale, E.; Salmi-Smail, C.; Brunel, J.-M.; Sanchez, P.; Fantini, J.; Maresca, M. Biophysical studies of the interaction of squalamine and other cationic amphiphilic molecules with bacterial and eukaryotic membranes: Importance of the distribution coefficient in membrane selectivity. Chem. Phys. Lipids 2010, 163, 131–140. [Google Scholar] [CrossRef]
- Yuan, Y.; Barrett, D.; Zhang, Y.; Kahne, D.; Sliz, P.; Walker, S. Crystal structure of a peptidoglycan glycosyltransferase suggests a model for processive glycan chain synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 5348–5353. [Google Scholar] [CrossRef] [Green Version]
- Boes, A.; Olatunji, S.; Mohammadi, T.; Breukink, E.; Terrak, M. Fluorescence anisotropy assays for high throughput screening of compounds binding to lipid II, PBP1b, FtsW and MurJ. Sci. Rep. 2020, 10, 6280. [Google Scholar] [CrossRef]
- Derouaux, A.; Sauvage, E.; Terrak, M. Peptidoglycan glycosyltransferase substrate mimics as templates for the design of new antibacterial drugs. Front. Immunol. 2013, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Sauvage, E.; Terrak, M. Glycosyltransferases and Transpeptidases/Penicillin-Binding Proteins: Valuable Targets for New Antibacterials. Antibiotics 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Ostash, B.; Walker, S. Moenomycin family antibiotics: Chemical synthesis, biosynthesis, and biological activity. Nat. Prod. Rep. 2010, 27, 1594–1617. [Google Scholar] [CrossRef] [PubMed]
- Bury, D.; Dahmane, I.; Derouaux, A.; Dumbre, S.; Herdewijn, P.; Matagne, A.; Breukink, E.; Mueller-Seitz, E.; Petz, M.; Terrak, M. Positive cooperativity between acceptor and donor sites of the peptidoglycan glycosyltransferase. Biochem. Pharmacol. 2015, 93, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A. Flavophospholipol use in animals: Positive implications for antimicrobial resistance based on its microbiologic properties. Diagn. Microbiol. Infect. Dis. 2006, 56, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, S.; Kusch, D.; Sinell, H.J. Staphylococcus aureus in poultry—Biochemical characteristics, antibiotic resistance and phage pattern (author’s transl). Zentralbl. Bakteriol. B 1979, 168, 546–561. [Google Scholar]
- King, D.T.; Wasney, G.A.; Nosella, M.; Fong, A.; Strynadka, N.C.J.; Peter Guengerich, F. Structural insights into inhibition of Escherichia coli penicillin-binding protein 1B. J. Biol. Chem. 2017, 292, 979–993. [Google Scholar] [CrossRef] [Green Version]
- Dumbre, S.; Derouaux, A.; Lescrinier, E.; Piette, A.; Joris, B.; Terrak, M.; Herdewijn, P. Synthesis of modified peptidoglycan precursor analogues for the inhibition of glycosyltransferase. J. Am. Chem. Soc. 2012, 134, 9343–9351. [Google Scholar] [CrossRef] [Green Version]
- Breukink, E.; van Heusden, H.E.; Vollmerhaus, P.J.; Swiezewska, E.; Brunner, L.; Walker, S.; Heck, A.J.; de Kruijff, B. Lipid II is an intrinsic component of the pore induced by nisin in bacterial membranes. J. Biol. Chem. 2003, 278, 19898–19903. [Google Scholar] [CrossRef] [Green Version]
- van Dam, V.; Sijbrandi, R.; Kol, M.; Swiezewska, E.; de Kruijff, B.; Breukink, E. Transmembrane transport of peptidoglycan precursors across model and bacterial membranes. Mol. Microbiol. 2007, 64, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Bouhss, A.; Crouvoisier, M.; Blanot, D.; Mengin-Lecreulx, D. Purification and characterization of the bacterial MraY translocase catalyzing the first membrane step of peptidoglycan biosynthesis. J. Biol. Chem. 2004, 279, 29974–29980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrak, M.; Ghosh, T.K.; Van Heijenoort, J.; Van Beeumen, J.; Lampilas, M.; Aszodi, J.; Ayala, J.A.; Ghuysen, J.-M.; Nguyen-Distèche, M. The catalytic, glycosyl transferase and acyl transferase modules of the cell wall peptidoglycan-polymerizing penicillin-binding protein 1b of Escherichia coli. Mol. Microbiol. 1999, 34, 350–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Rao, M.N.; Jones, S.R.; Shao, B.; Feibush, P.; Mcguigan, M.; Tzodikov, N.; Feibush, B.; Sharkansky, I.; Snyder, B.; et al. Synthesis of Squalamine Utilizing a Readily Accessible Spermidine Equivalent. J. Org. Chem. 1998, 63, 8599–8603. [Google Scholar] [CrossRef]
- Brunel, J.M.; Letourneux, Y. Recent advances in the synthesis of spermine and spermidine analogs of the shark aminosterol squalamine. Eur. J. Med. Chem. 2003, 2003, 3897–3907. [Google Scholar] [CrossRef]
- Loncle, C.; Salmi, C.; Vidal, N.; Letourneux, Y.; Brunel, J. Antimicrobial activities of 7-aminosterol squalamine analogues. Lett. Drug Des. Discov. 2008, 5, 388–393. [Google Scholar] [CrossRef]
- Loncle, C.; Salmi, C.; Letourneux, Y.; Brunel, J.M. Synthesis of new 7-aminosterol squalamine analogues with high antimicrobial activities through a stereoselective titanium reductive amination reaction. Tetrahedron 2007, 63, 12968–12974. [Google Scholar] [CrossRef]
- Blanchet, M.; Borselli, D.; Rodallec, A.; Peiretti, F.; Vidal, N.; Bolla, J.M.; Digiorgio, C.; Morrison, K.R.; Wuest, W.M.; Brunel, J.M. Claramines: A new class of broad-spectrum antimicrobial agents with bimodal activity. ChemMedChem 2018, 13, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Offant, J.; Terrak, M.; Derouaux, A.; Breukink, E.; Nguyen-Distèche, M.; Zapun, A.; Vernet, T. Optimization of conditions for the glycosyltransferase activity of penicillin-binding protein 1a from Thermotoga maritima. FEBS J. 2010, 277, 4290–4298. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, B.; Markwalder, J.A.; Seitz, S.P.; Wang, Y.; Stein, R.L. A kinetic characterization of the glycosyltransferase activity of Eschericia coli PBP1b and development of a continuous fluorescence assay. Biochemistry 2002, 41, 12552–12561. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Krieger, E.; Vriend, G. YASARA View—Molecular graphics for all devices—From smartphones to workstations|Bioinformatics|Oxford Academic. Bioinformatics 2014, 30, 2981–2982. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | RA % (0.8 mM) | RA % (0.4 mM) | IC50 (µM) |
|---|---|---|---|
| Squalamine (1) | 1.1 ± 0.6 | 6 ± 2 | 291 ± 26 |
| 2 | 0.3 ± 0.1 | 1.2 ± 0.3 | 114 ± 6 |
| 3 | 8.5 ± 3.1 | 16 ± 2 | 220 ± 10 |
| 4 | 4 ± 1 | 39 ± 2 | 303 ± 15 |
| 5 | 3 ± 1 | 16 ± 1.4 | 221 ± 26 |
| 6 | 0.8 ± 0.4 | 3.9 ± 0.1 | 111 ± 6 |
| 7 | 1.5 ± 0.8 | 12.5 ± 4.4 | 229 ± 17 |
| 8 | 97 ± 0.2 | nd | nd |
| 9 | 98 ± 1 | nd | nd |
| 10 | 99 ± 0.5 | nd | nd |
| 11 | 96 ± 2 | nd | nd |
| 12 | 98 ± 1 | nd | nd |
| 13 | 100 | nd | nd |
| 14 | 100 | nd | nd |
| 15 | 105 ± 3 | nd | nd |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boes, A.; Brunel, J.M.; Derouaux, A.; Kerff, F.; Bouhss, A.; Touze, T.; Breukink, E.; Terrak, M. Squalamine and Aminosterol Mimics Inhibit the Peptidoglycan Glycosyltransferase Activity of PBP1b. Antibiotics 2020, 9, 373. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070373
Boes A, Brunel JM, Derouaux A, Kerff F, Bouhss A, Touze T, Breukink E, Terrak M. Squalamine and Aminosterol Mimics Inhibit the Peptidoglycan Glycosyltransferase Activity of PBP1b. Antibiotics. 2020; 9(7):373. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070373
Chicago/Turabian StyleBoes, Adrien, Jean Michel Brunel, Adeline Derouaux, Frédéric Kerff, Ahmed Bouhss, Thierry Touze, Eefjan Breukink, and Mohammed Terrak. 2020. "Squalamine and Aminosterol Mimics Inhibit the Peptidoglycan Glycosyltransferase Activity of PBP1b" Antibiotics 9, no. 7: 373. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070373