Characterization of Clinical MRSA Isolates from Northern Spain and Assessment of Their Susceptibility to Phage-Derived Antimicrobials

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. DNA Preparation and Genomic Analysis
2.3. Biofilm Formation Assays
2.4. Quantification of Specific Lytic Activity
2.5. Phage Susceptibility
2.6. Data Analysis
3. Results
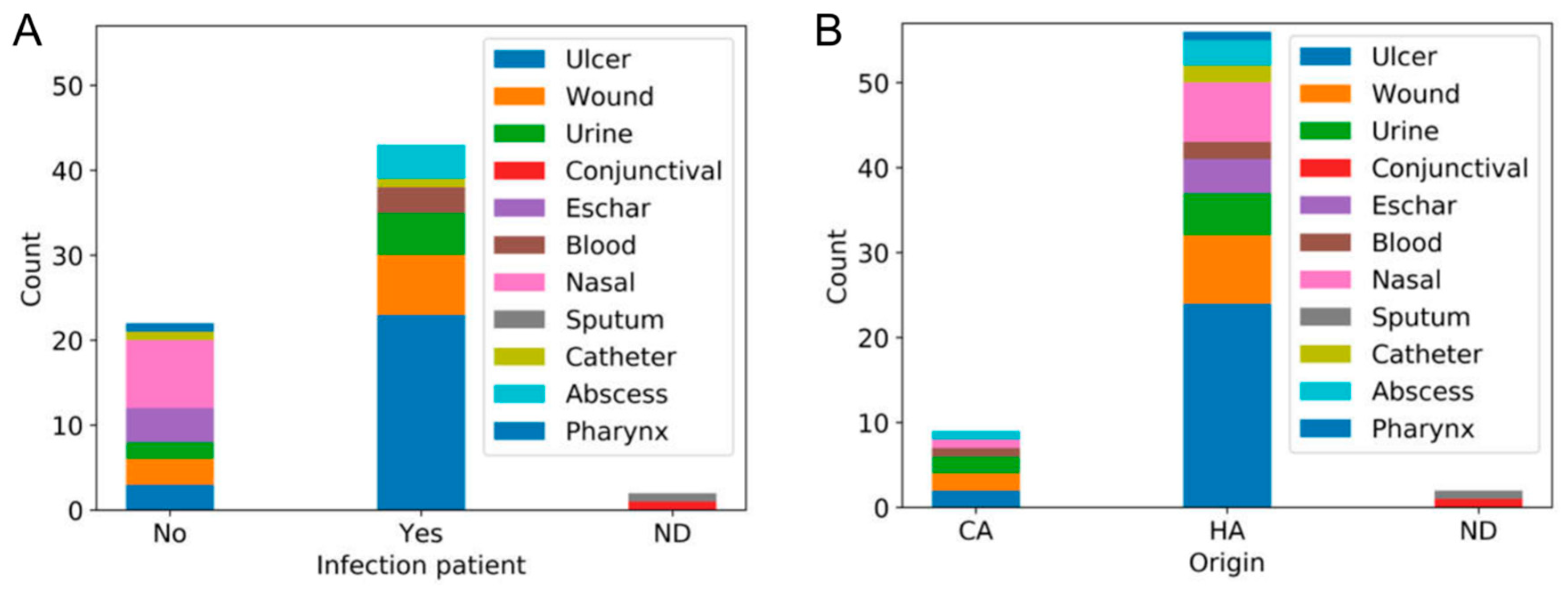
3.1. Isolation of MRSA Strains at San Agustín Hospital
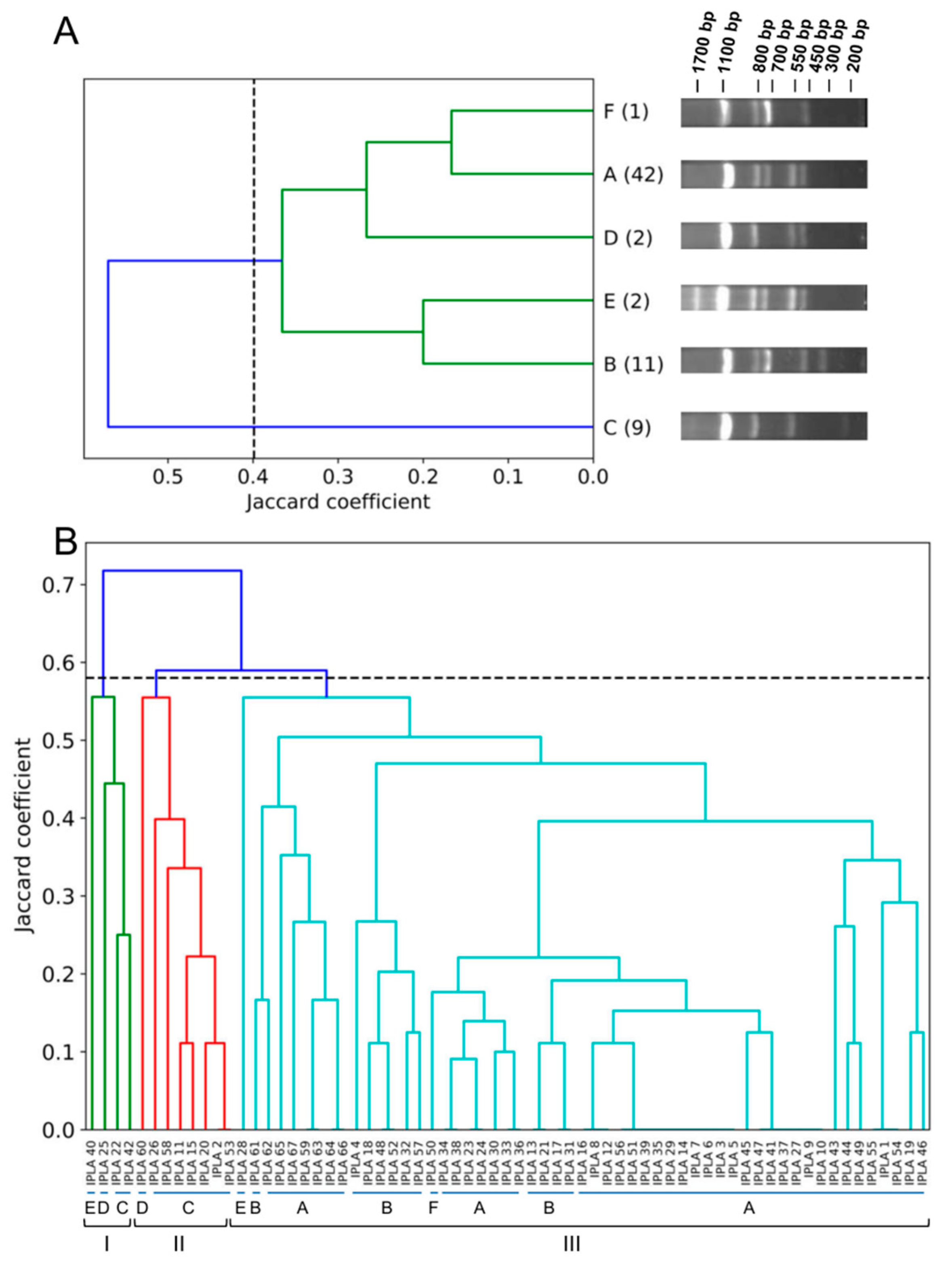
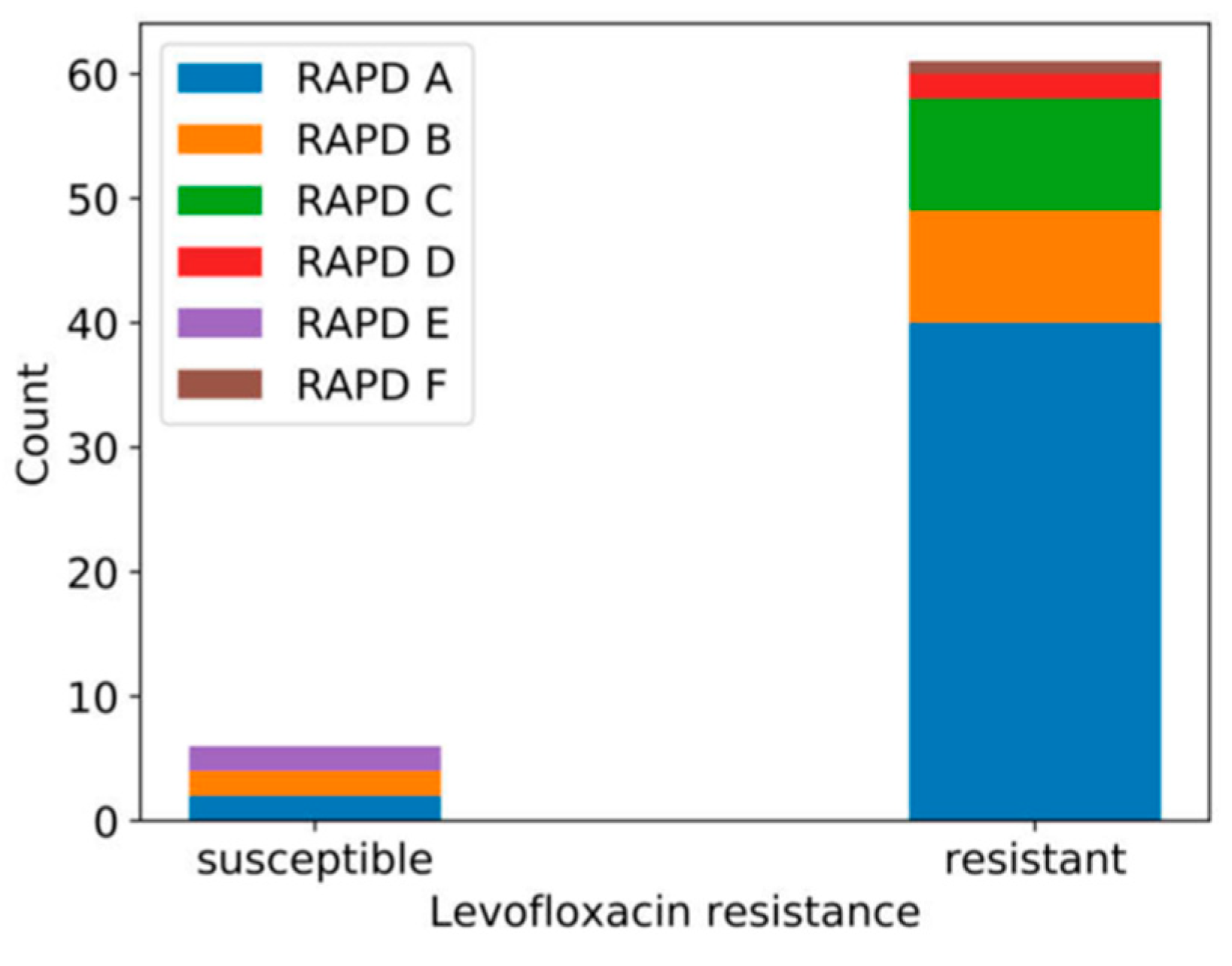
3.2. Genetic Diversity of Human MRSA Isolates
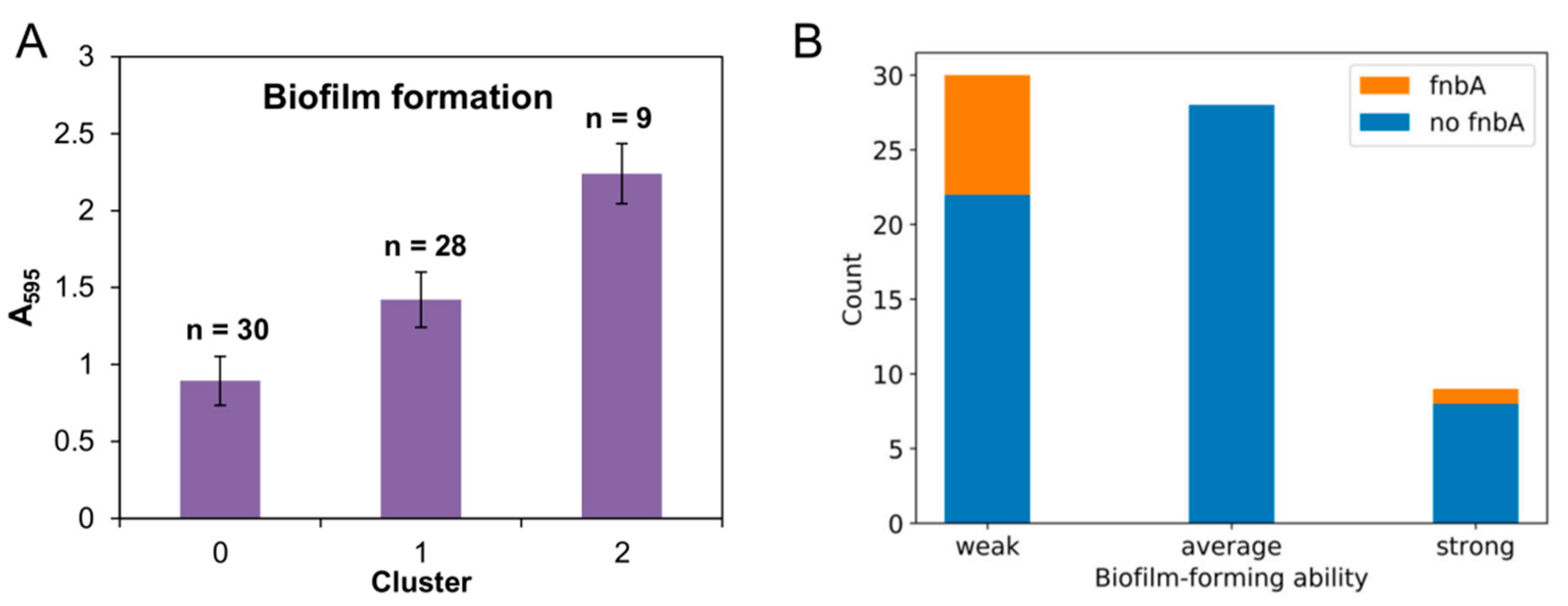
3.3. Biofilm Formation Ability
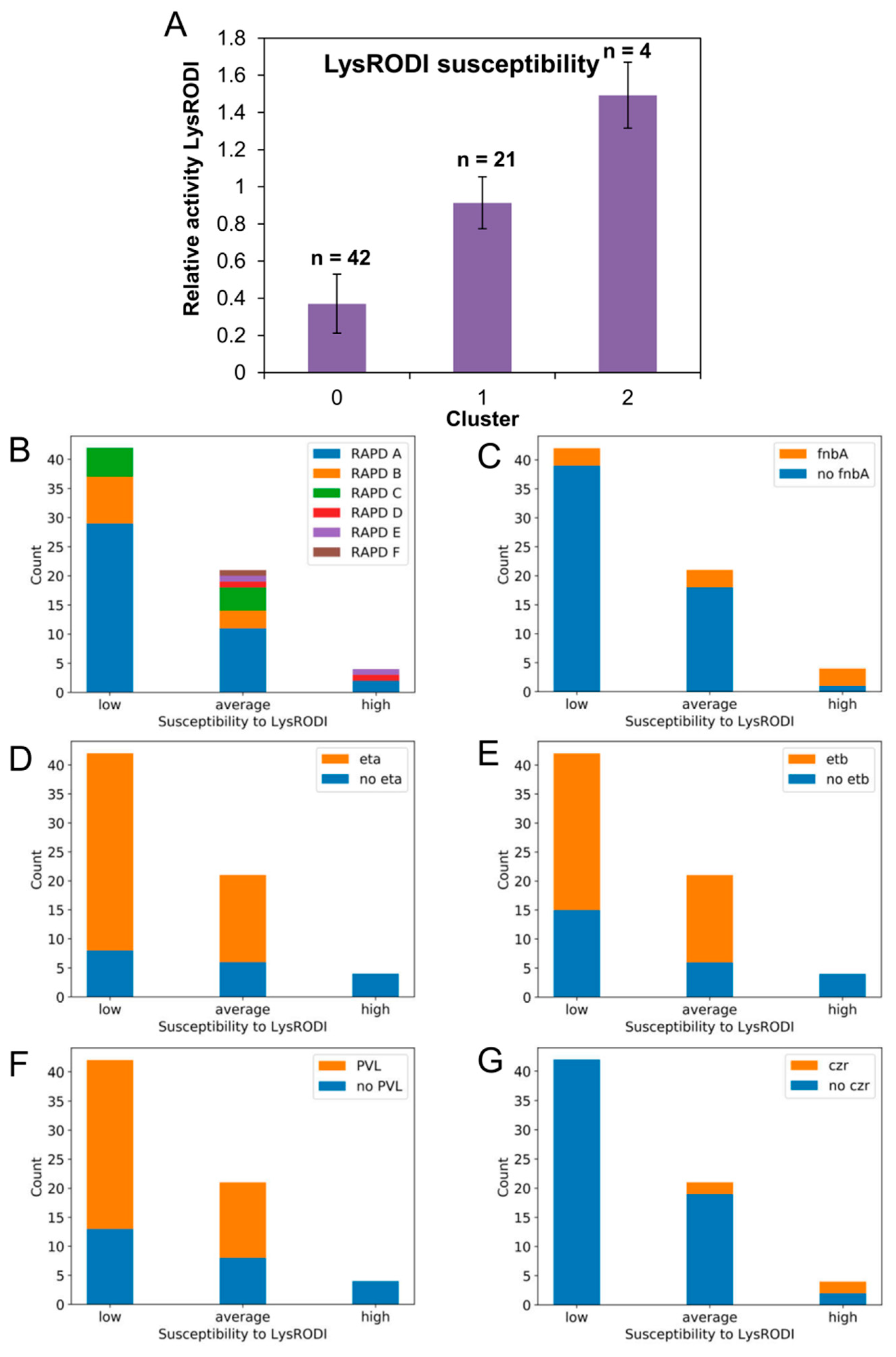
3.4. Susceptibility to Phages phiIPLA-RODI and phiIPLA-C1C
3.5. Susceptibility to Endolysin LysRODI
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Otto, M. Staphylococcal infections: Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 2008, 46, S350–S359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Quiles-Puchalt, N.; Chiang, Y.N.; Bacigalupe, R.; Fillol-Salom, A.; Chee, M.S.J.; Fitzgerald, J.R.; Penades, J.R. Genome hypermobility by lateral transduction. Science 2018, 362, 207–212. [Google Scholar] [CrossRef] [Green Version]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014; p. 257. [Google Scholar]
- Stefani, S.; Chung, D.R.; Lindsay, J.A.; Friedrich, A.W.; Kearns, A.M.; Westh, H.; Mackenzie, F.M. Meticillin-resistant Staphylococcus aureus (MRSA): Global epidemiology and harmonisation of typing methods. Int. J. Antimicrob. Agents 2012, 39, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Wyllie, D.H.; Walker, A.S.; Miller, R.; Moore, C.; Williamson, S.R.; Schlackow, I.; Finney, J.M.; O’Connor, L.; Peto, T.E.; Crook, D.W. Decline of meticillin-resistant Staphylococcus aureus in Oxfordshire hospitals is strain-specific and preceded infection-control intensification. BMJ Open 2011, 1, e000160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations-a review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 211. [Google Scholar] [CrossRef] [Green Version]
- Barbier, F.; Andremont, A.; Wolff, M.; Bouadma, L. Hospital-acquired pneumonia and ventilator-associated pneumonia: Recent advances in epidemiology and management. Curr. Opin. Pulm. Med. 2013, 19, 216–228. [Google Scholar] [CrossRef]
- DeRonde, K.J.; Girotto, J.E.; Nicolau, D.P. Management of Pediatric Acute Hematogenous Osteomyelitis, Part I: Antimicrobial stewardship approach and review of therapies for methicillin-susceptible Staphylococcus aureus, Streptococcus pyogenes, and Kingella kingae. Pharmacotherapy 2018, 38, 947–966. [Google Scholar] [CrossRef]
- Weiser, M.C.; Moucha, C.S. The current state of screening and decolonization for the prevention of Staphylococcus aureus surgical site infection after total hip and knee arthroplasty. J. Bone Jt. Surg. Am. 2015, 97, 1449–1458. [Google Scholar] [CrossRef]
- Alvarez, A.; Fernandez, L.; Gutierrez, D.; Iglesias, B.; Rodriguez, A.; Garcia, P. Methicillin-resistant Staphylococcus aureus in hospitals: Latest trends and treatments based on bacteriophages. J. Clin. Microbiol. 2019, 57, e01006-19. [Google Scholar] [CrossRef]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage therapy in clinical practice: Treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef]
- Schuch, R.; Nelson, D.; Fischetti, V.A. A bacteriolytic agent that detects and kills Bacillus anthracis. Nature 2002, 418, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Letrado, P.; Corsini, B.; Diez-Martinez, R.; Bustamante, N.; Yuste, J.E.; Garcia, P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-based antibacterials as therapeutics. Adv. Exp. Med. Biol. 2019, 1148, 233–253. [Google Scholar] [PubMed]
- Harhala, M.; Nelson, D.C.; Miernikiewicz, P.; Heselpoth, R.D.; Brzezicka, B.; Majewska, J.; Linden, S.B.; Shang, X.; Szymczak, A.; Lecion, D.; et al. Safety studies of pneumococcal endolysins Cpl-1 and Pal. Viruses 2018, 10, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briers, Y.; Lavigne, R. Breaking barriers: Expansion of the use of endolysins as novel antibacterials against Gram-negative bacteria. Future Microbiol. 2015, 10, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Heselpoth, R.D.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Lysocins: Bioengineered antimicrobials that deliver lysins across the outer membrane of Gram-negative bacteria. Antimicrob. Agents Chemother. 2019, 63, e00342-19. [Google Scholar] [CrossRef] [Green Version]
- Jun, S.Y.; Jang, I.J.; Yoon, S.; Jang, K.; Yu, K.S.; Cho, J.Y.; Seong, M.W.; Jung, G.M.; Yoon, S.J.; Kang, S.H. Pharmacokinetics and tolerance of the phage endolysin-based candidate drug SAL200 after a single intravenous administration among healthy volunteers. Antimicrob. Agents Chemother. 2017, 61, e02629-16. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://clinicaltrials.gov/ct2/show/NCT03163446#wrapper (accessed on 1 October 2019).
- García, P.; Martínez, B.; Obeso, J.M.; Lavigne, R.; Lurz, R.; Rodríguez, A. Functional genomic analysis of two Staphylococcus aureus phages isolated from the dairy environment. Appl. Environ. Microbiol. 2009, 75, 7663–7673. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species staphylococcal biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Garrido, V.; Fernández, L.; Portilla, S.; Rodríguez, A.; Grilló, M.J.; García, P. Phage lytic protein LysRODI prevents staphylococcal mastitis in mice. Front. Microbiol. 2020, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obeso, J.M.; Martinez, B.; Rodriguez, A.; Garcia, P. Lytic activity of the recombinant staphylococcal bacteriophage PhiH5 endolysin active against Staphylococcus aureus in milk. Int. J. Food Microbiol. 2008, 128, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Rubio, L.; Martinez, B.; Donovan, D.M.; Garcia, P.; Rodriguez, A. Potential of the virion-associated peptidoglycan hydrolase HydH5 and its derivative fusion proteins in milk biopreservation. PLoS ONE 2013, 8, e54828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, D.; Ruas-Madiedo, P.; Martínez, B.; Rodríguez, A.; García, P. Effective removal of staphylococcal biofilms by the endolysin LysH5. PLoS ONE 2014, 9, e107307. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; González, S.; Campelo, A.B.; Martínez, B.; Rodríguez, A.; García, P. Downregulation of autolysin-encoding genes by phage-derived lytic proteins inhibits biofilm formation in Staphylococcus aureus. Antimicrob. Agents Chemother. 2017, 61, e02724-16. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.L.; Quednau, M.; Molin, G.; Ahrne, S. Randomly amplified polymorphic DNA (RAPD) for rapid typing of Lactobacillus plantarum strains. Lett. Appl. Microbiol. 1995, 21, 155–159. [Google Scholar] [CrossRef]
- Herrera, J.J.; Cabo, M.L.; Gonzalez, A.; Pazos, I.; Pastoriza, L. Adhesion and detachment kinetics of several strains of Staphylococcus aureus subsp. aureus under three different experimental conditions. Food Microbiol. 2007, 24, 585–591. [Google Scholar] [CrossRef]
- Tristan, A.; Ying, L.; Bes, M.; Etienne, J.; Vandenesch, F.; Lina, G. Use of multiplex PCR to identify Staphylococcus aureus adhesins involved in human hematogenous infections. J. Clin. Microbiol. 2003, 41, 4465–4467. [Google Scholar] [CrossRef] [Green Version]
- Verdier, I.; Durand, G.; Bes, M.; Taylor, K.L.; Lina, G.; Vandenesch, F.; Fattom, A.I.; Etienne, J. Identification of the capsular polysaccharides in Staphylococcus aureus clinical isolates by PCR and agglutination tests. J. Clin. Microbiol. 2007, 45, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Delgado, S.; Vázquez-Sánchez, D.; Martínez, B.; Cabo, M.L.; Rodríguez, A.; Herrera, J.J.; García, P. Incidence of Staphylococcus aureus and analysis of associated bacterial communities on food industry surfaces. Appl. Environ. Microbiol. 2012, 78, 8547–8554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; Roth, R.; Peters, G. Rapid and specific detection of toxigenic Staphylococcus aureus: Use of two multiplex PCR enzyme immunoassays for amplification and hybridization of staphylococcal enterotoxin genes, exfoliative toxin genes, and toxic shock syndrome toxin 1 gene. J. Clin. Microbiol. 1998, 36, 2548–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.L.; Ueda, S.; Shinagawa, K. Detection of seg, seh, and sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates harboring seg, seh, or sei genes. J. Clin. Microbiol. 2002, 40, 857–862. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Kar, S.; Choudhury, R.; Sharma, S.; Singh, D.V. Development and evaluation of hexaplex PCR for rapid detection of methicillin, cadmium/zinc and antiseptic-resistant staphylococci, with simultaneous identification of PVL-positive and -negative Staphylococcus aureus and coagulase negative staphylococci. FEMS Microbiol. Lett. 2014, 352, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Planet, P.J.; Narechania, A.; Chen, L.; Mathema, B.; Boundy, S.; Archer, G.; Kreiswirth, B. Architecture of a species: Phylogenomics of Staphylococcus aureus. Trends Microbiol. 2017, 25, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Bowler, P.G. Antibiotic resistance and biofilm tolerance: A combined threat in the treatment of chronic infections. J. Wound Care 2018, 27, 273–277. [Google Scholar] [CrossRef] [PubMed]
- O’Gara, J.P. Ica and beyond: Biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flock, J.I. Extracellular-matrix-binding proteins as targets for the prevention of Staphylococcus aureus infections. Mol. Med. Today 1999, 5, 532–537. [Google Scholar] [CrossRef]
- Chhibber, S.; Shukla, A.; Kaur, S. Transfersomal phage cocktail is an effective treatment against methicillin-resistant Staphylococcus aureus-mediated skin and soft tissue infections. Antimicrob. Agents Chemother. 2017, 61, e02146-16. [Google Scholar] [CrossRef] [Green Version]
- Takemura-Uchiyama, I.; Uchiyama, J.; Osanai, M.; Morimoto, N.; Asagiri, T.; Ujihara, T.; Daibata, M.; Sugiura, T.; Matsuzaki, S. Experimental phage therapy against lethal lung-derived septicemia caused by Staphylococcus aureus in mice. Microbes Infect. 2014, 16, 512–517. [Google Scholar] [CrossRef]
- Chhibber, S.; Kaur, J.; Kaur, S. Liposome entrapment of bacteriophages improves wound healing in a diabetic mouse MRSA Infection. Front. Microbiol. 2018, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- Lobocka, M.; Hejnowicz, M.S.; Dabrowski, K.; Gozdek, A.; Kosakowski, J.; Witkowska, M.; Ulatowska, M.I.; Weber-Dabrowska, B.; Kwiatek, M.; Parasion, S.; et al. Genomics of staphylococcal Twort-like phages—Potential therapeutics of the post-antibiotic era. Adv. Virus Res. 2012, 83, 143–216. [Google Scholar] [PubMed]
- Gutierrez, D.; Fernandez, L.; Rodriguez, A.; Garcia, P. Are phage lytic proteins the secret weapon to kill Staphylococcus aureus? MBio 2018, 9, e01923-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaack, D.; Idelevich, E.A.; Schleimer, N.; Molinaro, S.; Kriegeskorte, A.; Peters, G.; Becker, K. Bactericidal activity of bacteriophage endolysin HY-133 against Staphylococcus aureus in comparison to other antibiotics as determined by minimum bactericidal concentrations and time-kill analysis. Diagn. Microbiol. Infect. Dis. 2019, 93, 362–368. [Google Scholar] [CrossRef]
- Idelevich, E.A.; Schaumburg, F.; Knaack, D.; Scherzinger, A.S.; Mutter, W.; Peters, G.; Peschel, A.; Becker, K. The recombinant bacteriophage endolysin HY-133 exhibits in vitro activity against different african clonal lineages of the Staphylococcus aureus complex, Including Staphylococcus schweitzeri. Antimicrob. Agents Chemother. 2016, 60, 2551–2553. [Google Scholar] [CrossRef] [Green Version]
- Endl, J.; Seidl, H.P.; Fiedler, F.; Schleifer, K.H. Chemical composition and structure of cell wall teichoic acids of staphylococci. Arch. Microbiol. 1983, 135, 215–223. [Google Scholar] [CrossRef]
- Idelevich, E.A.; von Eiff, C.; Friedrich, A.W.; Iannelli, D.; Xia, G.; Peters, G.; Peschel, A.; Wanninger, I.; Becker, K. In vitro activity against Staphylococcus aureus of a novel antimicrobial agent, PRF-119, a recombinant chimeric bacteriophage endolysin. Antimicrob. Agents Chemother. 2011, 55, 4416–4419. [Google Scholar] [CrossRef] [Green Version]
- Bibek, G.C.; Sahukhal, G.S.; Elasri, M.O. Role of the msaABCR operon in cell wall biosynthesis, autolysis, integrity, and antibiotic resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2019, 63, e00680-19. [Google Scholar]
- Nair, R.; Thapaliya, D.; Su, Y.; Smith, T.C. Resistance to zinc and cadmium in Staphylococcus aureus of human and animal origin. Infect. Control Hosp. Epidemiol. 2014, 35, S32–S39. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, I.; Uchiyama, J.; Tsukui, T.; Hisatsune, J.; Ide, K.; Matsuzaki, S.; Sugai, M.; Nishifuji, K. Therapeutic potential of an endolysin derived from kayvirus S25-3 for staphylococcal impetigo. Viruses 2019, 11, 769. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.; Li, X.; Wang, B.; Gong, P.; Xiao, F.; Yang, M.; Zhang, L.; Song, J.; Hu, L.; Cheng, M.; et al. Combination therapy of LysGH15 and apigenin as a new strategy for treating pneumonia caused by Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.S.; Singh, R.; McKinnell, J.A.; Park, S.; Gombosev, A.; Eells, S.J.; Gillen, D.L.; Kim, D.; Rashid, S.; Macias-Gil, R.; et al. Decolonization to reduce postdischarge infection risk among MRSA carriers. N. Engl. J. Med. 2019, 380, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Schleimer, N.; Kaspar, U.; Knaack, D.; von Eiff, C.; Molinaro, S.; Grallert, H.; Idelevich, E.A.; Becker, K. In vitro activity of the bacteriophage endolysin HY-133 against Staphylococcus aureus small-colony variants and their corresponding wild types. Int. J. Mol. Sci. 2019, 20, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, A.; Euler, C.; Collin, M.; Chahales, P.; Gorelick, K.J.; Fischetti, V.A. Synergism between a novel chimeric lysin and oxacillin protects against infection by methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 1603–1612. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Poonacha, N.; Desai, S.; Hiremath, D.; Tuppad, D.; Mohan, T.; Chikkamadaiah, R.; Durgaiah, M.; Kumar, S.; Channabasappa, S.; et al. Restoration of sensitivity of a diverse set of drug-resistant Staphylococcus clinical strains by bactericidal protein P128. J. Med. Microbiol. 2018, 67, 296–307. [Google Scholar] [CrossRef]
- Schuch, R.; Lee, H.M.; Schneider, B.C.; Sauve, K.L.; Law, C.; Khan, B.K.; Rotolo, J.A.; Horiuchi, Y.; Couto, D.E.; Raz, A.; et al. Combination therapy with lysin CF-301 and antibiotic is superior to antibiotic alone for treating methicillin-resistant Staphylococcus aureus-induced murine bacteremia. J. Infect. Dis. 2014, 209, 1469–1478. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sample | Infection | Origin | RAPD Profile | Susceptibility phiIPLA-RODI | Relative Specific Activity LysRODI * | Biofilm Formation * | Virulence/Resistance Genes * |
|---|---|---|---|---|---|---|---|---|
| 1 | Ulcer exudate | + | HA | A | S | 0.869 ± 0.060 | 1.208 ± 0.736 | ebpS, fib, cap5, seg, sei, PVL/LfxR |
| 2 | Ulcer exudate | + | HA | C | S | 0.778 ± 0.019 | 1.069 ± 0.611 | cna, cap5, sec, seg, sei, eta, etb, PVL/LfxR |
| 3 | Ulcer exudate | + | HA | A | S | 0.256 ± 0.020 | 0.627 ± 0.469 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 4 | Ulcer exudate | + | CA | B | S | 0.506 ± 0.173 | 1.084 ± 0.739 | fnbB, fib, cap5, see, seg, sei, eta, etb, PVL |
| 5 | Urine | + | HA | A | S | 0.895 ± 0.053 | 1.458 ± 0.773 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 6 | Ulcer exudate | + | HA | A | S | 0.592 ± 0.058 | 0.995 ± 0.344 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 7 | Ulcer exudate | + | HA | A | S | 0.439 ± 0.047 | 0.872 ± 0.559 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 8 | Ulcer exudate | + | HA | A | S | 0.330 ± 0.040 | 1.334 ± 0.895 | fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 9 | Ulcer exudate | - | HA | A | S | 0.345 ± 0.025 | 1.765 ± 0.618 | ebpS, fib, cap5, seg, sei, eta, etb/CLIR |
| 10 | Ulcer exudate | + | HA | A | S | 0.409 ± 0.017 | 1.506 ± 0.343 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR, CLIR |
| 11 | Conjunctival exudate | ND | ND | C | R | 1.136 ± 0.097 | 0.895 ± 0.552 | cna, ebpS, cap5, seg, sei, eta, etb/LfxR |
| 12 | Ulcer exudate | + | HA | A | S | 0.101 ± 0.030 | 1.196 ± 0.896 | fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 13 | Wound exudate | - | HA | B | S | 0.281 ± 0.127 | 1.353 ± 0.885 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 14 | Ulcer exudate | + | HA | A | S | 0.564 ± 0.043 | 2.236 ± 0.720 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR, CLIR |
| 15 | Ulcer exudate | + | HA | C | S | 0.458 ± 0.044 | 1.861 ± 0.348 | cna, ebpS, cap5, seg, sei, eta, etb, PVL/LfxR* |
| 16 | Ulcer exudate | + | HA | A | R | 0.380 ± 0.030 | 0.954 ± 0.389 | fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 17 | Ulcer exudate | + | HA | B | S | 0.910 ± 0.144 | 1.375 ± 0.694 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 18 | Ulcer exudate | - | HA | B | S | 0.619 ± 0.076 | 0.804 ± 0.496 | ebpS, fnbB, fib, cap5, seg, sei, eta, PVL/LfxR |
| 19 | Urine | - | HA | A | R | 0.681 ± 0.076 | 2.300 ± 0.324 | ebpS, fib, cap5, sed, see, seg, sei/LfxR |
| 20 | Eschar exudate | - | HA | C | S | 0.569 ± 0.121 | 1.630 ± 0.796 | cna, cap5, seg, sei, eta, etb, PVL/LfxR, CLIR |
| 21 | Blood culture | + | CA | B | R | 0.771 ± 0.099 | 1.781 ± 0.441 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 22 | Blood culture | + | HA | C | S | 0.427 ± 0.109 | 0.626 ± 0.474 | cna, fnbA, cap5, seg, sei, PVL/LfxR |
| 23 | Urine | - | CA | A | S | 1.068 ± 0.081 | 0.828 ± 0.291 | ebpS, fib, cap5, sed, see, seg, sei, eta, etb, PVL/LfxR |
| 24 | Nasal exudate | - | HA | A | R | 0.800 ± 0.041 | 2.214 ± 0.519 | ebpS, fib, cap5, sed, see, seg, sei, eta, etb, PVL/LfxR |
| 25 | Nasal exudate | - | HA | D | S | 1.364 ± 0.153 | 0.786 ± 0.421 | cna, fnbA, fnbB, cap5, seg, sei, czr/LfxR |
| 26 | Eschar exudate | - | HA | C | R | 0.575 ± 0.062 | 1.143 ± 0.401 | cap5, sed, see, seg, sei, eta, etb, PVL/LfxR |
| 27 | Nasal exudate | - | CA | A | R | 0.614 ± 0.034 | 0.739 ± 0.347 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 28 | Nasal exudate | - | HA | E | S | 1.755 ± 0.169 | 0.892 ± 0.317 | ebpS, fnbA, fib, cap5, sec, seg, sei |
| 29 | Sputum | ND | ND | A | S | 0.857 ± 0.093 | 2.287 ± 0.322 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR, CLIR |
| 30 | Eschar exudate | - | HA | A | S | 0.566 ± 0.100 | 1.384 ± 0.275 | ebpS, fib, cap5, sed, see, seg, sei, eta, etb/LfxR, CLIR |
| 31 | Urine | + | HA | B | S | 1.076 ± 0.083 | 1.160 ± 0.456 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 32 | Nasal exudate | - | HA | B | S | 0.316 ± 0.054 | 0.979 ± 0.309 | fnbA, fnbB, fib, cap5, seg, sei, PVL/LfxR |
| 33 | Wound exudate | + | HA | A | S | 1.115 ± 0.311 | 0.919 ± 0.146 | ebpS, fib, cap5, sed, seg, sei, eta, etb/LfxR |
| 34 | Ulcer exudate | + | HA | A | R | 0.383 ± 0.045 | 0.693 ± 0.646 | ebpS, fib, cap5, sed, seg, sei, eta, etb, PVL/LfxR |
| 35 | Nasal exudate | - | HA | A | R | 0.456 ± 0.121 | 1.131 ± 0.360 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 36 | Eschar exudate | - | HA | A | R | 0.884 ± 0.048 | 0.555 ± 0.325 | ebpS, fib, cap5, sed, seg, sei, eta, etb/LfxR |
| 37 | Wound exudate | - | HA | A | S | 0.333 ± 0.035 | 1.288 ± 0.615 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 38 | Pericatheter | + | HA | A | S | 0.946 ± 0.008 | 1.326 ± 0.440 | ebpS, fib, cap5, sed, seg, sei, eta, etb, PVL/LfxR |
| 39 | Ulcer exudate | + | HA | A | S | 0.576 ± 0.061 | 0.992 ± 0.408 | ebpS, fib, cap5, seg, sei, eta, etb, PVL |
| 40 | Abscess | + | HA | E | S | 1.100 ± 0.105 | 0.985 ± 0.338 | bbp, cna, ebpS, fnbA, cap8, seg, sei, PVL |
| 41 | Nasal exudate | - | HA | A | S | 0.121 ± 0.019 | 0.648 ± 0.235 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 42 | Ulcer exudate | + | HA | C | S | 0.913 ± 0.221 | 0.893 ± 0.447 | cna, fnbA, cap5, cap8, seg, sei/LfxR |
| 43 | Abscess | + | HA | A | S | 0.363 ± 0.039 | 0.882 ± 0.484 | ebpS, fnbA, fib, cap5, cap8, seg, sei/LfxR |
| 44 | Pharyngeal exudate | - | HA | A | S | 0.964 ± 0.068 | 0.865 ± 0.430 | ebpS, fnbA, fib, cap5, sed, seg, sei, czr/LfxR |
| 45 | Nasal exudate | - | HA | A | S | 0.108 ± 0.015 | 1.571 ± 0.465 | ebpS, fib, cap5, seg, sei, eta/LfxR |
| 46 | Blood | + | HA | A | S | 1.433 ± 0.091 | 1.043 ± 0.840 | ebpS, fib, cap5, sed, seg, sei/LfxR, CLIR |
| 47 | Urine | + | CA | A | R | 0.366 ± 0.047 | 2.214 ± 0.788 | ebpS, fib, cap5, seg, sei, eta, etb/LfxR |
| 48 | Abscess | + | HA | B | S | 0.102 ± 0.023 | 1.350 ± 0.353 | fnbB, fib, cap5, seg, sei, eta, PVL/LfxR |
| 49 | Telescoping catheter | - | HA | A | R | 1.418 ± 0.080 | 2.361 ± 0.863 | ebpS, fnbA, fib, cap5, seg, sei, czr/LfxR, CLIR |
| 50 | Ulcer exudate | + | HA | F | S | 0.654 ± 0.126 | 1.301 ± 0.339 | ebpS, fib, cap5, sed, seg, sei, eta, etb, PVL/LfxR |
| 51 | Ulcer exudate | - | HA | A | R | 0.353 ± 0.052 | 1.683 ± 0.482 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 52 | Abscess | + | CA | B | S | 0.528 ± 0.136 | 1.404 ± 0.562 | fnbB, fib, cap5, seg, sei, eta, PVL/LfxR |
| 53 | Ulcer exudate | + | HA | C | S | 0.213 ± 0.035 | 1.000 ± 0.186 | cna, cap5, sec, seg, sei, eta, etb, PVL/LfxR |
| 54 | Ulcer exudate | + | HA | A | S | 0.353 ± 0.035 | 1.398 ± 0.566 | ebpS, fib, cap5, seg, sei, PVL/LfxR, TMP/SMXR |
| 55 | Ulcer exudate | + | CA | A | R | 1.007 ± 0.083 | 1.204 ± 0.608 | ebpS, fib, cap5, seg, sei, PVL/LfxR |
| 56 | Ulcer exudate | + | CA | A | S | 0.072 ± 0.021 | 2.583 ± 0.572 | ebpS, fib, cap5, seg, sei, eta, etb, PVL/LfxR |
| 57 | Wound exudate | + | CA | B | R | 0.218 ± 0.031 | 0.905 ± 0.416 | fnbB, fib, cap5, seg, sei, PVL |
| 58 | Wound exudate | + | HA | C | S | 0.769 ± 0.055 | 0.879 ± 0.398 | cna, cap5, sec, eta, etb, PVL/LfxR |
| 59 | Ulcer exudate | + | HA | A | S | 0.253 ± 0.031 | 1.561 ± 0.417 | ebpS, fib, cap5, PVL/LfxR |
| 60 | Wound exudate | + | HA | D | S | 1.001 ± 0.089 | 1.463 ± 0.147 | cna, fnbB, cap5, eta, etb, PVL, czr/LfxR, CLIR |
| 61 | Wound exudate | + | HA | B | R | 0.163 ± 0.030 | 1.718 ± 0.622 | ebpS, fib, cap5, eta, etb/LfxR |
| 62 | Ulcer exudate | + | HA | A | R | 0.584 ± 0.141 | 1.223 ± 0.720 | ebpS, fib, cap5, eta, etb/LfxR |
| 63 | Wound exudate | + | HA | A | R | 0.169 ± 0.071 | 1.307 ± 0.655 | ebpS, fib, cap5, PVL/LfxR |
| 64 | Urine | + | HA | A | S | 0.335 ± 0.013 | 1.133 ± 0.625 | ebpS, fib, cap5, eta, PVL/LfxR, CLIR |
| 65 | Wound exudate | - | HA | A | R | 0.325 ± 0.005 | 1.280 ± 0.456 | ebpS, fib, cap5, sed, eta/LfxR, CLIR |
| 66 | Urine | + | HA | A | S | 0.444 ± 0.358 | 2.102 ± 0.704 | ebpS, fib, cap5, eta, PVL/LfxR |
| 67 | Ulcer exudate | + | HA | A | R | 0.397 ± 0.024 | 1.575 ± 0.444 | ebpS, fib, cap5/LfxR |
| Virulence factor | Gene (pb) | Primers | Reference |
|---|---|---|---|
| Bone sialoprotein binding protein | bbp (575) | BBP-1, BBP-2 | [31] |
| Collagen binding protein | cna (423) | CNA-1, CNA-2 | |
| Laminin binding protein | eno (302) | ENO-1, ENO-2 | |
| Elastin binding protein | ebpS (186) | EBP-1, EBP-1 | |
| Fibronectin binding protein | fnbA (643) | FNBA-1, FNBA-2 | |
| fnbB (524) | FNBB-1, FNBB-2 | ||
| Fibrinogen binding protein | fib (404) | FIB-1, FIB-2 | |
| Clumping factor | clfA (292) | CLFA-1, CLFA-2 | |
| clfB (205) | CLFB-1, CLFB-2 | ||
| Capsule | cap5 (361) | Cap5 k1, Cap5 k2 | [32] |
| cap8 (173) | Cap8 k1, Cap8 k2 | ||
| Polysaccharide matrix | icaA (1315) | icaA-R, icaA-R | [33] |
| icaD (381) | icaD-F, icaD-R | ||
| Proteinaceous matrix | bap (971) | BAP-sasp-6m, BAP-sasp-7c | |
| Enterotoxins | sea (127) | SEA3, SEA4 | [34] |
| seb (477) | SEB1, SEB4 | ||
| sec (271) | SEC3, SEC4 | ||
| sed (319) | SED3, SED4 | ||
| see (178) | SEE3, SEE2 | ||
| seg (287) | SEG1, SEG2 | [35] | |
| seh (213) | SEH1, SEH2 | ||
| sei (454) | SEI1, SEI2 | ||
| Toxic shock syndrome toxin | tst (445) | TST3, TST6 | [34] |
| Exfoliative toxins | eta (119) | ETA3, ETA4 | |
| etb (262) | ETB3, ETB4 | ||
| Panton-Valentine leukocidin | PVL (505) | PVL505-F, PVL505-R | [36] |
| Cadmium and zinc resistance | czr (655) | czrC-F, czrC-R | |
| Methicillin resistance | mecA (264) | mecA264-F, mecA264-R | |
| Quaternary ammonium compounds resistance | qacA/B (192) | QacA/B-F, QacA/B-R |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salas, M.; Wernecki, M.; Fernández, L.; Iglesias, B.; Gutiérrez, D.; Álvarez, A.; García, L.; Prieto, E.; García, P.; Rodríguez, A. Characterization of Clinical MRSA Isolates from Northern Spain and Assessment of Their Susceptibility to Phage-Derived Antimicrobials. Antibiotics 2020, 9, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9080447
Salas M, Wernecki M, Fernández L, Iglesias B, Gutiérrez D, Álvarez A, García L, Prieto E, García P, Rodríguez A. Characterization of Clinical MRSA Isolates from Northern Spain and Assessment of Their Susceptibility to Phage-Derived Antimicrobials. Antibiotics. 2020; 9(8):447. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9080447
Chicago/Turabian StyleSalas, Marina, Maciej Wernecki, Lucía Fernández, Beatriz Iglesias, Diana Gutiérrez, Andrea Álvarez, Laura García, Elisabeth Prieto, Pilar García, and Ana Rodríguez. 2020. "Characterization of Clinical MRSA Isolates from Northern Spain and Assessment of Their Susceptibility to Phage-Derived Antimicrobials" Antibiotics 9, no. 8: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9080447