Surveillance of Antibiotic Resistance of Maltose-Negative Staphylococcus aureus in South African Dairy Herds

 ,
,
Abstract
:1. Introduction
2. Results
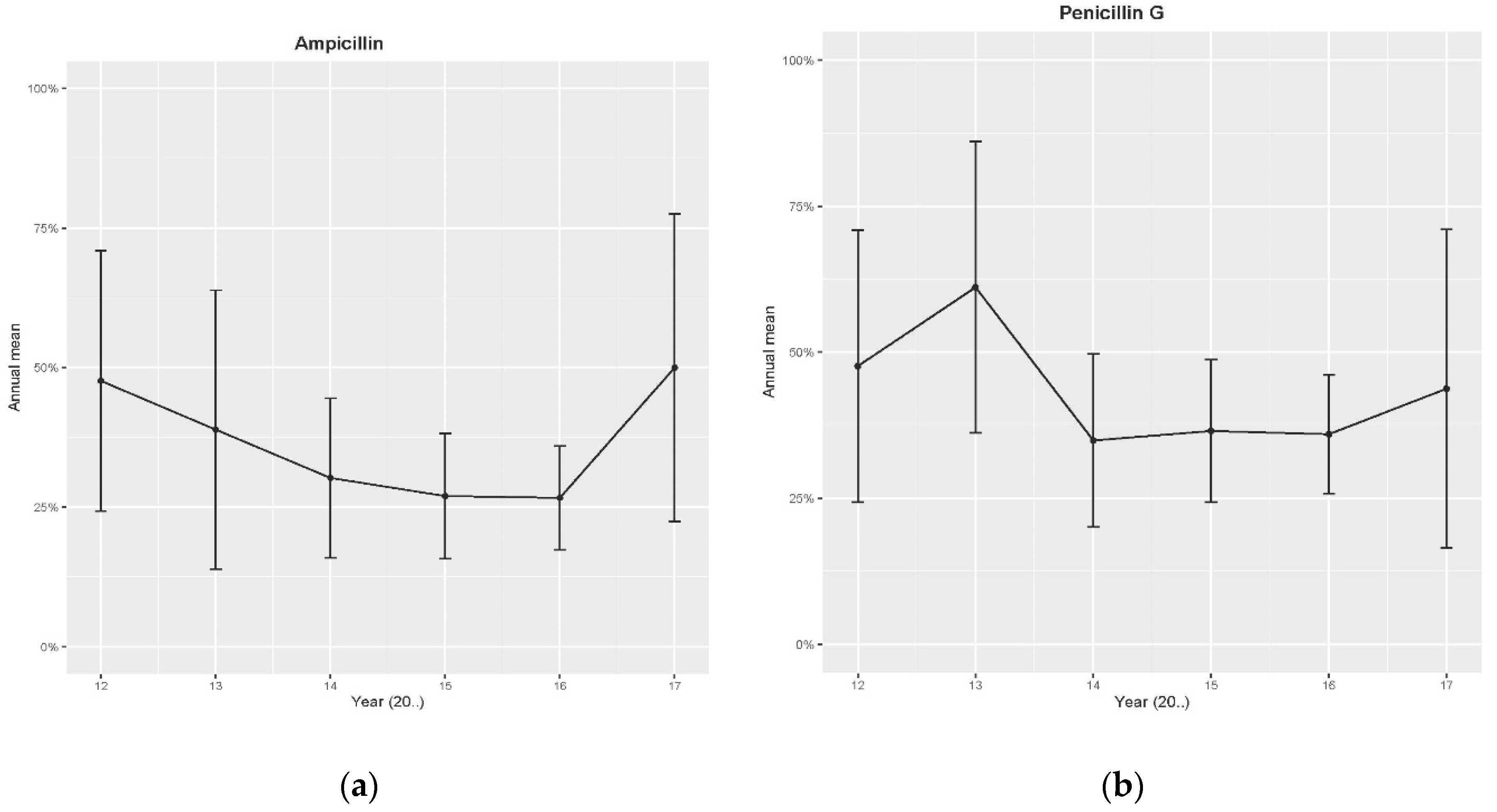
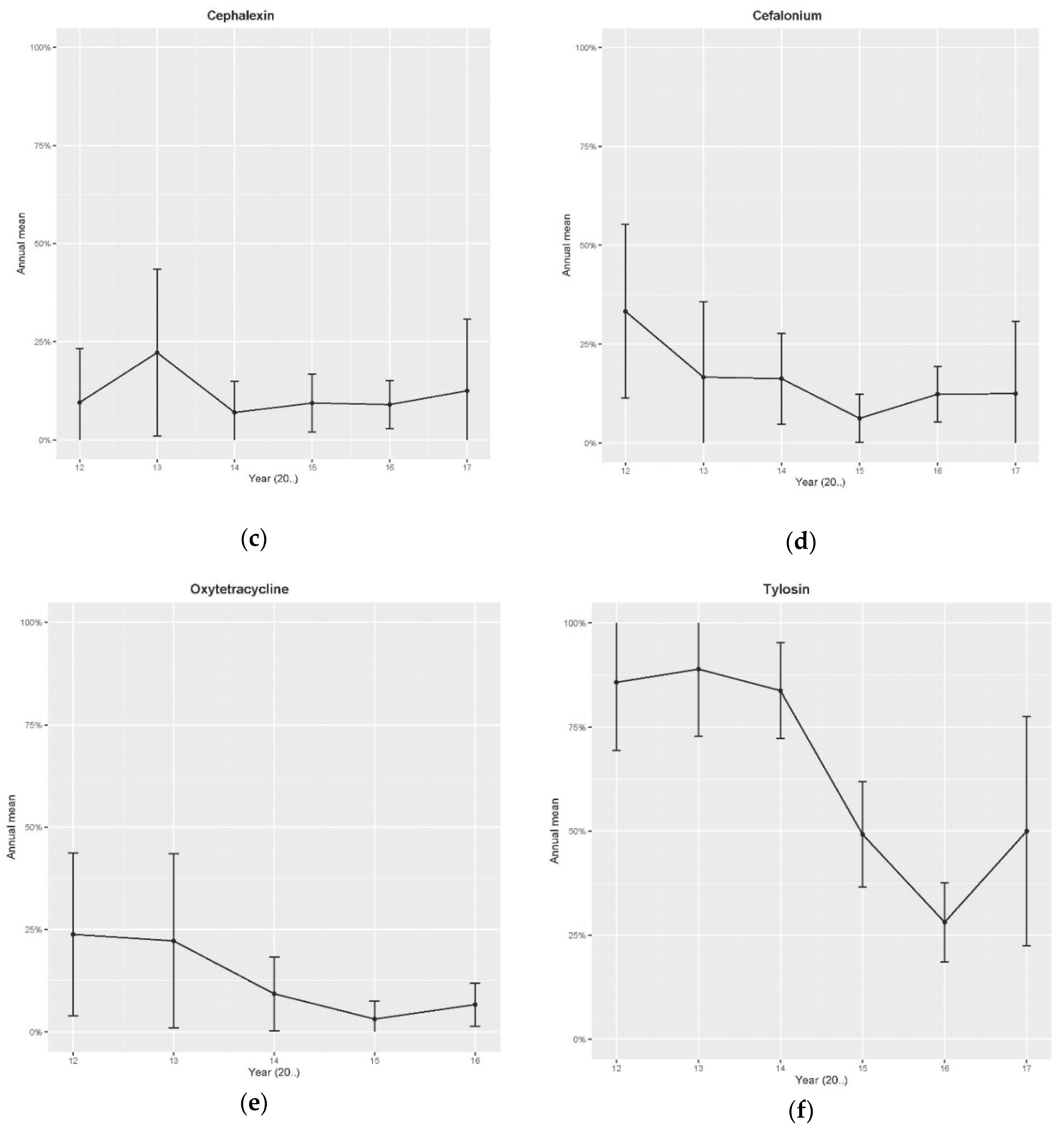
2.1. Retrospective Data—Disk Diffusion Method
2.2. Minimum Inhibitory Concentration (MIC) Analysis
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tenhagen, B.-A.; Hansen, I.; Reinecke, A.; Heuwieser, W. Prevalence of pathogens in milk samples of dairy cows with clinical mastitis and in heifers at first parturition. J. Dairy Res. 2009, 76, 179–187. [Google Scholar] [CrossRef]
- Sasaki, T.; Kikuchi, K.; Tanaka, Y.; Takahashi, N.; Kamata, S.; Hiramatsu, K. Reclassification of Phenotypically Identified Staphylococcus intermedius Strains. J. Clin. Microbiol. 2007, 45, 2770–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piessens, V.; Van Coillie, E.; Verbist, B.; Supré, K.; Braem, G.; Van Nuffel, A.; De Vuyst, L.; Heyndrickx, M.; De Vliegher, S. Distribution of coagulase-negative Staphylococcus species from milk and environment of dairy cows differs between herds. J. Dairy Sci. 2011, 94, 2933–2944. [Google Scholar] [CrossRef] [PubMed]
- Kwon, N.H.; Park, K.-T.; Jung, W.K.; Youn, H.Y.; Lee, Y.; Kim, S.H.; Bae, W.; Lim, J.Y.; Kim, J.Y.; Kim, J.M.; et al. Characteristics of methicillin resistant Staphylococcus aureus isolated from chicken meat and hospitalized dogs in Korea and their epidemiological relatedness. Vet. Microbiol. 2006, 117, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Investigators ABCs AM: Invasive methicillin-resistant Staphylococcus aureus infections in the United States. J. Am. Med. Assoc. 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petzer, I.-M.; Karzis, J.; Watermeyer, J.; Van Der Schans, T.; Van Reenen, R. Trends in udder health and emerging mastitogenic pathogens in South African dairy herds. J. S. Afr. Vet. Assoc. 2009, 80, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Zadoks, R.N.; Fitzpatrick, J. Changing trends in mastitis. Ir. Vet. J. 2009, 62, S59–S70. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, S.C.; Owens, W.E.; Boddie, R.L. Symposium: Mastitis in dairy heifers: Initial studies on prevalence and control. J. Dairy Sci. 1995, 78, 1607–1618. [Google Scholar] [CrossRef]
- Sol, J.; Sampimon, O.; Snoep, J.; Schukken, Y. Factors Associated with Bacteriological Cure During Lactation After Therapy for Subclinical Mastitis Caused by Staphylococcus aureus. J. Dairy Sci. 1997, 80, 2803–2808. [Google Scholar] [CrossRef]
- Karzis, J.; Petzer, I.-M.; Donkin, E.; Naidoo, V.; Labuschagne, C. Short communication: Characterization of an atypical maltose-negative Staphylococcus aureus through the use of phenotypic and molecular techniques. J. Dairy Sci. 2020, 103, 7407–7410. [Google Scholar] [CrossRef]
- Karzis, J. Antibiotic Resistance of Coagulase Positive Staphylococci Isolated from Milk of South African Dairy Herds. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, April 2020. [Google Scholar]
- Johler, S.; Moser, M.; Engl, C.; Tasara, T.; Corti, S.; Chen, J.; Stephan, R. A Coagulase- and -Glucosidase-Negative Variant of Staphylococcus aureus: A Challenge for Routine Microbiological Diagnostics. J. Clin. Microbiol. 2012, 50, 1827–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, Approved Standard, 3rd ed. CLSI Document VET01-A4. Available online: https://www.clsi.org (accessed on 1 November 2019).
- Clinical and Laboratory Standards Institute. Performance Standard for Antimicrobial Susceptibility Testing. Twenty-Fifth Information Supplement M100-SWayne. Available online: https://www.clsi.org (accessed on 1 November 2019).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.E.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toutain, P.; Bousquet-Mélou, A.; Damborg, P.; Ferran, A.; Mevius, D.; Pelligand, L.; Veldman, K.T.; Lees, P. En Route towards European Clinical Breakpoints for Veterinary Antimicrobial Susceptibility Testing: A Position Paper Explaining the VetCAST Approach. Front. Microbiol. 2017, 8, 2344. [Google Scholar] [CrossRef] [Green Version]
- Niemi, R.; Vilar, M.; Dohoo, I.; Hovinen, M.; Simojoki, H.; Rajala-Schultz, P.J. Antibiotic dry cow therapy, somatic cell count, and milk production: Retrospective analysis of the associations in dairy herd recording data using multilevel growth models. Prev. Vet. Med. 2020, 180, 105028. [Google Scholar] [CrossRef] [PubMed]
- Karzis, J.; Petzer, I.-M.; Donkin, E.; Naidoo, V.; Etter, E.M. Climatic and regional antibiotic resistance patterns of Staphylococcus aureus in South African dairy herds. Onderstepoort J. Vet. Res. 2019, 86, e1–e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B. The Farming Handbook; University of KwaZulu Natal Press: KwaZulu Natal, South Africa, 2006; pp. 100–330. ISBN 13:978-1-86914-090-8. [Google Scholar]
- Zadoks, R.N.; Middleton, J.R.; McDougall, S.; Katholm, J.; Schukken, Y. Molecular Epidemiology of Mastitis Pathogens of Dairy Cattle and Comparative Relevance to Humans. J. Mammary Gland. Biol. Neoplasia 2011, 16, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Petzer, I.M.; Karzis, J. Virulence, antibiotic resistance and diagnostics of CNS isolated from milk of SA dairy cows. In Proceedings of the RuVasa Congress, Umhlanga Rocks, South Africa, 20–22 May 2019. [Google Scholar]
- Karzis, J.; Petzer, I.M.; Donkin, E.F.; Naidoo, V. Proactive udder health management in South Africa and antimicrobial resistance monitoring of Staphylococcus aureus in dairy herds from 2001 to 2010. JSAVA 2018, 89, 1–8. [Google Scholar]
- Sydnor, E.R.M.; Perl, T.M. Hospital Epidemiology and Infection Control in Acute-Care Settings. Clin. Microbiol. Rev. 2011, 24, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Kallen, A.J.; Mu, Y.; Bulens, S.; Reingold, A.; Petit, S.; Gershman, K.; Ray, S.M.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; et al. Health Care–Associated Invasive MRSA Infections, 2005–2008. JAMA 2010, 304, 641. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. 2017, 70, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; Available online: http://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 7 June 2017).
- Salmon, S.; Watts, J.; Aarestrup, F.M.; Pankey, J.; Yancey, R. Minimum Inhibitory Concentrations for Selected Antimicrobial Agents Against Organisms Isolated from the Mammary Glands of Dairy Heifers in New Zealand and Denmark. J. Dairy Sci. 1998, 81, 570–578. [Google Scholar] [CrossRef]
- Schmidt, L.H. The MIC50/MIC90: Assessments of in vitro activities of antimicrobial agents that facilitate comparative agent-agent and agent-species susceptibility comparisons. Antimicrob. Newslett. 1987, 4, 1–8. [Google Scholar] [CrossRef]
- Hájek, V. Staphylococcus intermedius, a New Species Isolated from Animals. Int. J. Syst. Bacteriol. 1976, 26, 401–408. [Google Scholar] [CrossRef]
- Haran, K.P.; Godden, S.M.; Boxrud, D.; Jawahir, S.; Bender, J.B.; Sreevatsan, S. Prevalence and Characterization of Staphylococcus aureus, Including Methicillin-Resistant Staphylococcus aureus, Isolated from Bulk Tank Milk from Minnesota Dairy Farms. J. Clin. Microbiol. 2011, 50, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Aubry-Damon, H.; Grenet, K.; Sall-Ndiaye, P.; Che, D.; Cordeiro, E.; Bougnoux, M.-E.; Rigaud, E.; Le Strat, Y.; Lemanissier, V.; Armand-Lefevre, L.; et al. Antimicrobial Resistance in Commensal Flora of Pig Farmers. Emerg. Infect. Dis. 2004, 10, 873–879. [Google Scholar] [CrossRef]
- Voss, A.; Loeffen, F.; Bakker, J.; Klaassen, C.; Wulf, M. Methicillin resistant Staphylococcus aureus in pig farming. Emerg. Infect. Dis. 2005, 11, 1965–1966. [Google Scholar] [CrossRef]
- Schmidt, T.; Kock, M.; Ehlers, M.M. Diversity and antimicrobial susceptibility profiling of staphylococci isolated from bovine mastitis cases and close human contacts. J. Dairy Sci. 2015, 98, 6256–6269. [Google Scholar] [CrossRef]
- Food and Agriculture Organisation (FAO). Afr. Res. Bull. Econ. Financ. Tech. Ser. 2011, 48, 19067. [CrossRef]
- Wong, A. Unknown Risk on the Farm: Does Agricultural Use of Ionophores Contribute to the Burden of Antimicrobial Resistance? mSphere 2019, 4, e00433-19. [Google Scholar] [CrossRef] [Green Version]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, Present, and Future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.B.; Machado, M.F.R.; Picoli, S.U. An investigation of methicillin-resistant Staphylococcus pseudintermedius (MRSP) in domestic and shelter dogs in Montenegro (RS-Brazil). Rev. Bras. Saúde Prod. Anim. 2017, 18, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Wallmann, J.; Kaspar, H.; Kroker, R. The prevalence of antimicrobial susceptibility of veterinary pathogens isolated from cattle and pigs: National antibiotic resistance monitoring 2002/2003 of the BVL. Berl. Munch. Tierarztl. Wochenschr. 2004, 117, 480–492. [Google Scholar]
- Abdi, R.D.; Gillespie, B.E.; Vaughn, J.; Merrill, C.; Headrick, S.I.; Ensermu, D.B.; D’Souza, D.H.; Agga, G.E.; Almeida, R.A.; Oliver, S.P.; et al. Antimicrobial Resistance of Staphylococcus aureus Isolates from Dairy Cows and Genetic Diversity of Resistant Isolates. Foodborne Pathog. Dis. 2018, 15, 449–458. [Google Scholar] [CrossRef]
- National Department of Health. Surveillance Resistance and Consumption of Antibiotics in South Africa. Available online: https://www.health.gov.za (accessed on 1 July 2019).
- Swinkels, J.M.; Cox, P.; Schukken, Y.; Lam, T. Efficacy of extended cefquinome treatment of clinical Staphylococcus aureus mastitis. J. Dairy Sci. 2013, 96, 4983–4992. [Google Scholar] [CrossRef] [Green Version]
- Barkema, H.W.; Schukken, Y.; Zadoks, R.N. Invited Review: The Role of Cow, Pathogen, and Treatment Regimen in the Therapeutic Success of Bovine Staphylococcus aureus Mastitis. J. Dairy Sci. 2006, 89, 1877–1895. [Google Scholar] [CrossRef] [Green Version]
- Petzer, I.-M.; Karzis, J.; Donkin, E.F.; Webb, E.C. A pathogen-specific approach towards udder health management in dairy herds: Using culture and somatic cell counts from routine herd investigations. Onderstepoort J. Vet. Res. 2016, 83, 12. [Google Scholar] [CrossRef] [PubMed]
- Lactodata. Milk SA. Available online: http://www.milksa.co.za/category/all-reports/industry-information (accessed on 7 June 2017).
- National Mastitis Council. Microbiological Procedures for the Diagnosis of Bovine Udder Infection and Determination of Milk Quality—Fourth Edition. Available online: http://www.nmconline.org (accessed on 1 August 2017).
- Petzer, I.-M.; Karzis, J.; Donkin, E.F.; Webb, E.C.; Etter, E.M. Somatic cell count thresholds in composite and quarter milk samples as indicator of bovine intramammary infection status. Onderstepoort J. Vet. Res. 2017, 84, 1269. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Kirby Bauer method antimicrobial susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Tsai, A.C. Achieving consensus on terminology describing multivariable analyses. Am. J. Public Health 2013, 103, e1. [Google Scholar] [CrossRef]
- Dohoo, I.; Ducrot, C.; Fourichon, C.; Donald, A.; Hurnik, D. An overview of techniques for dealing with large numbers of independent variables in epidemiologic studies. Prev. Vet. Med. 1997, 29, 221–239. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 30–35. ISBN 0-387-95364-7. [Google Scholar]



{kind=link}
{kind=link}
| Antibiotics | Year | Season | Province | SCC Category |
|---|---|---|---|---|
| Tylosin | Cubic effect (p < 0.001 for Year, Year2 and Year3) | NS | NS | NS |
| Penicillin G | p = 0.018 | p = 0.05 [Autumn p = 0.03 & Summer p = 0.02] vs. Spring (Lowest R) | NS | NS |
| Ampicillin | p = 0.008 | N/A | NS | NS |
| Clindamycin | N/A | NS | N/A | N/A |
| Cefuroxime | NS | NS | NS | NS |
| Cephalonium (2011–2017) | p = 0.03 * | NS | NS | p = 0.027 [High SCC p = 0.007] vs. Med SCC (Lowest R) |
| Cefoxitin (2014–2017) | NS | NS | NS | NS |
| Oxytetracycline | p < 0.001 | NS | NS | NS |
| Cephalexin | p = 0.017 | p = 0.06 [p = 0.01 Summer] vs. Autumn (Lowest R) | NS | NS |
| Cloxacillin | NS | NS | NS | p = 0.022 [Low SCC p = 0.08] vs. High SCC (Lowest R) |
| Product | % Resistance Maltose-Positive S. aureus (n) | % Resistance Maltose-Negative S. aureus (n) | MIC 90 | MIC 50 | Fisher’s Exact Test p-Value | ||
|---|---|---|---|---|---|---|---|
| Maltose-Positive S. aureus | Maltose-Negative S. aureus | Maltose-Positive S. aureus | Maltose-Negative S. aureus | ||||
| Amox/K Clav | 3.5 (2/57) | 21.1 (12/57) | 1 | 2 | 0.5 | 0.5 | 0.008 ** |
| Ampicillin | 100 (57/57) | 100 (57/57) | 4 | 4 | 0.5 | 0.5 | |
| Azithromycin | 3.5 (2/57) | 1.8 (1/57) | 1 | 1 | _ | _ | |
| Cefepime | 1.8 (1/57) | 22.8 (13/57) | 4 | 4 | _ | _ | |
| Cefotaxime | 1.8 (1/57) | 22.8 (13/57) | 1 | 2 | _ | _ | p < 0.001 ** |
| Cefoxitin | 1.8 (1/57) | 0 (0/57) | 4 | 4 | _ | _ | |
| Cefuroxime | 1.8 (1/57) | 21.1 (12/57) | 4 | 8 | _ | _ | 0.002 ** |
| Ciprofloxacin | 7 (4/57) | 1.8 (1/57) | 1 | 0.5 | 0.5 | _ | |
| Clindamycin | 5.3 (3/57) | 24.6 (14/57) | 0.3 | _ | _ | 0.25 | 0.007 ** |
| Daptomycin | 0 (0/57) | 17.5 (10/57) | 0.5 | _ | _ | 0.5 | 0.001 ** |
| Ertapenem | 1.8 (1/57) | 15.8 (9/57) | 0.5 | 0.5 | _ | 0.5 | 0.016 * |
| Erythromycin | 3.5 (2/57) | 1.8 (1/57) | 1 | _ | _ | 1 | |
| Fosfomycin | 1.8 (1/57) | 12.3 (7/57) | 32 | _ | _ | 32 | |
| Fusidic Acid | 0 (0/57) | 10.5 (6/57) | 2 | _ | _ | 2 | 0.027 * |
| Gentamicin | 7 (4/57) | 22.8 (13/57) | 1 | 2 | _ | 1 | 0.033 * |
| Imipenem | 3.5 (2/57) | 21.1 (12/57) | 2 | 2 | _ | _ | |
| Levofloxacin | 0 (0/57) | 0 (0/57) | 1 | 1 | _ | _ | |
| Linezolid | 0 (0/57) | 15.8 (9/57) | 2 | _ | 1 | 2 | 0.003 * |
| Meropenem | 1.8 (1/57) | 21.1 (12/57) | 2 | 2 | _ | _ | |
| Moxifloxacin | 0 (0/57) | 0 (0/57) | 0.5 | 0.5 | _ | _ | |
| Nitrofurantoin | 0 (0/57) | 8.8 (5/57) | 64 | 64 | _ | _ | |
| Oxacillin | 1.8 (1/57) | 21.1 (12/57) | 0.3 | _ | 0.5 | 0.25 | 0.002 ** |
| Penicillin | 36.8 (21/57) | 47.4 (27/57) | _ | _ | 0.03 | 0.12 | |
| Rifampin | 0 (0/57) | 5.3 (3/57) | 0.5 | 0.5 | _ | _ | |
| Synercid | 5.3 (3/57) | 19.3 (11/57) | 1 | _ | 1 | 0.043 * | |
| Teicoplanin | 0 (0/57) | 14 (8/57) | 1 | _ | _ | 1 | 0.006 ** |
| Tetracycline | 7 (4/57) | 7 (4/57) | 1 | 1 | 1 | _ | |
| Tobramycin | 3.5 (2/57) | 7 (4/57) | 1 | 1 | _ | _ | |
| Trimeth/Sulpha | 0 (0/57) | 0 (0/57) | 1 | 1 | _ | _ | |
| Vancomycin | 0 (0/57) | 31.6 (18/57) | 2 | _ | 1 | 1 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karzis, J.; Petzer, I.-M.; Donkin, E.F.; Naidoo, V.; Etter, E.M.C. Surveillance of Antibiotic Resistance of Maltose-Negative Staphylococcus aureus in South African Dairy Herds. Antibiotics 2020, 9, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090616
Karzis J, Petzer I-M, Donkin EF, Naidoo V, Etter EMC. Surveillance of Antibiotic Resistance of Maltose-Negative Staphylococcus aureus in South African Dairy Herds. Antibiotics. 2020; 9(9):616. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090616
Chicago/Turabian StyleKarzis, Joanne, Inge-Marié Petzer, Edward F. Donkin, Vinny Naidoo, and Eric M.C. Etter. 2020. "Surveillance of Antibiotic Resistance of Maltose-Negative Staphylococcus aureus in South African Dairy Herds" Antibiotics 9, no. 9: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090616