Delivery of the 5-HT2A Receptor Agonist, DOI, Enhances Activity of the Sphincter Muscle during the Micturition Reflex in Rats after Spinal Cord Injury



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Spinal Cord Surgery
2.3. Metabolic Cages
2.4. Bladder Cystometry and EUS Electromyography (EMG) Recordings
2.5. Drugs
2.6. Fresh Tissue Harvesting and Protein Isolation
2.7. Western Blot
2.8. RNA Extraction/Quantitative Real-Time PCR
2.9. Statistical Analyses
3. Results
3.1. 5-HT2A Protein and Gene Expression Is Sustained in the Spinal Cord Following SCI
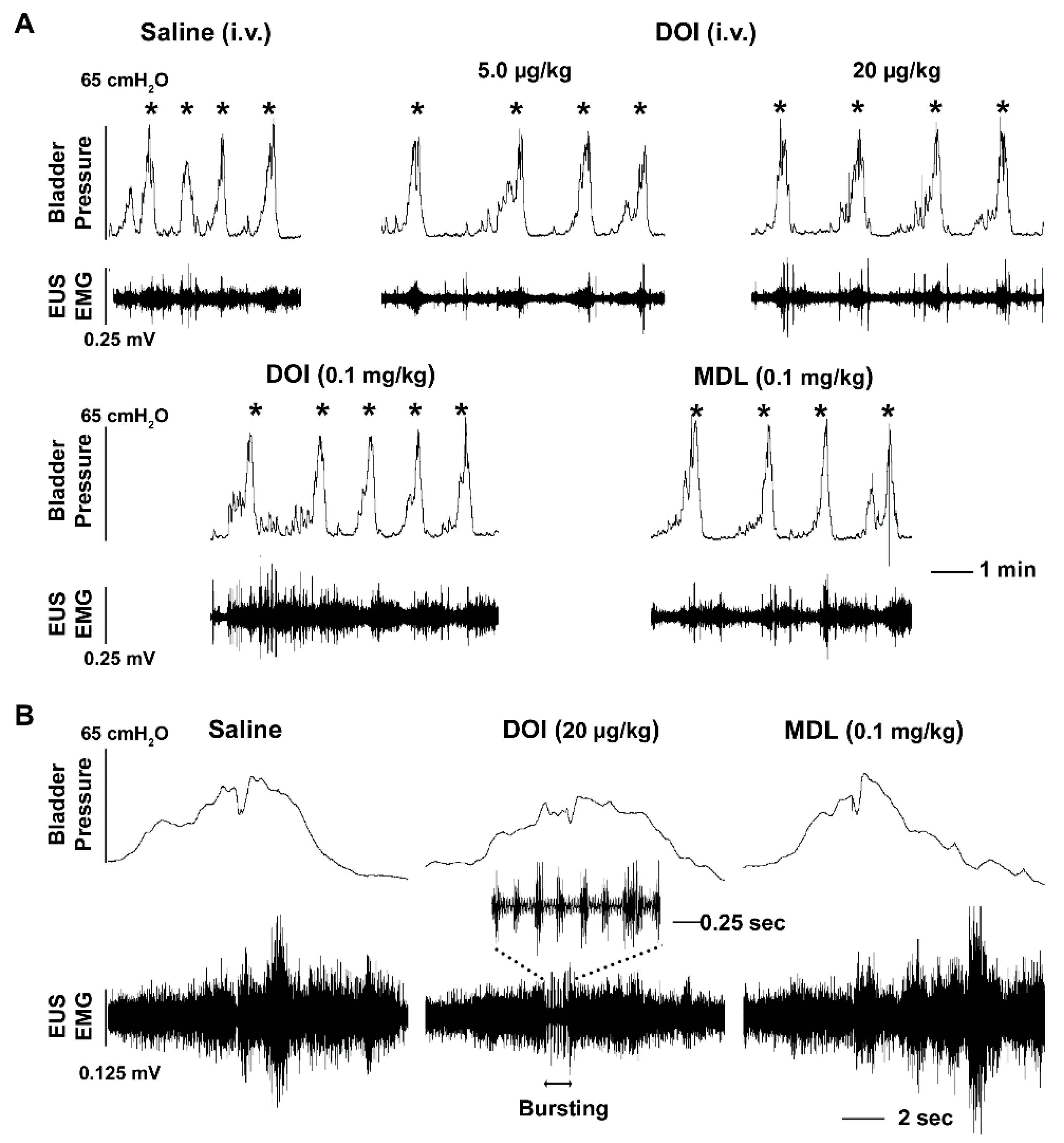
3.2. I.v. Drug Delivery and its Effects on Bladder and EUS Reflexes
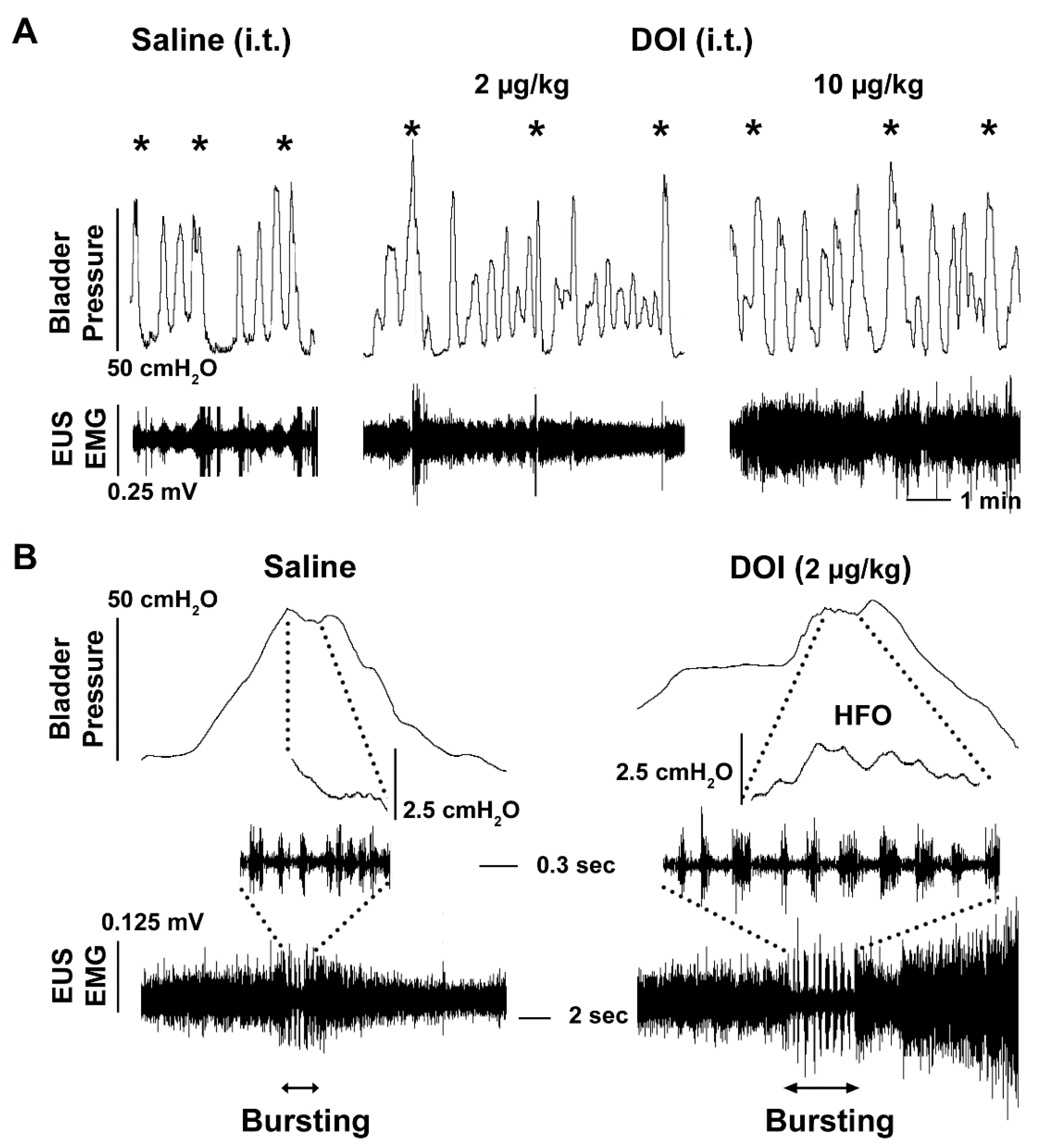
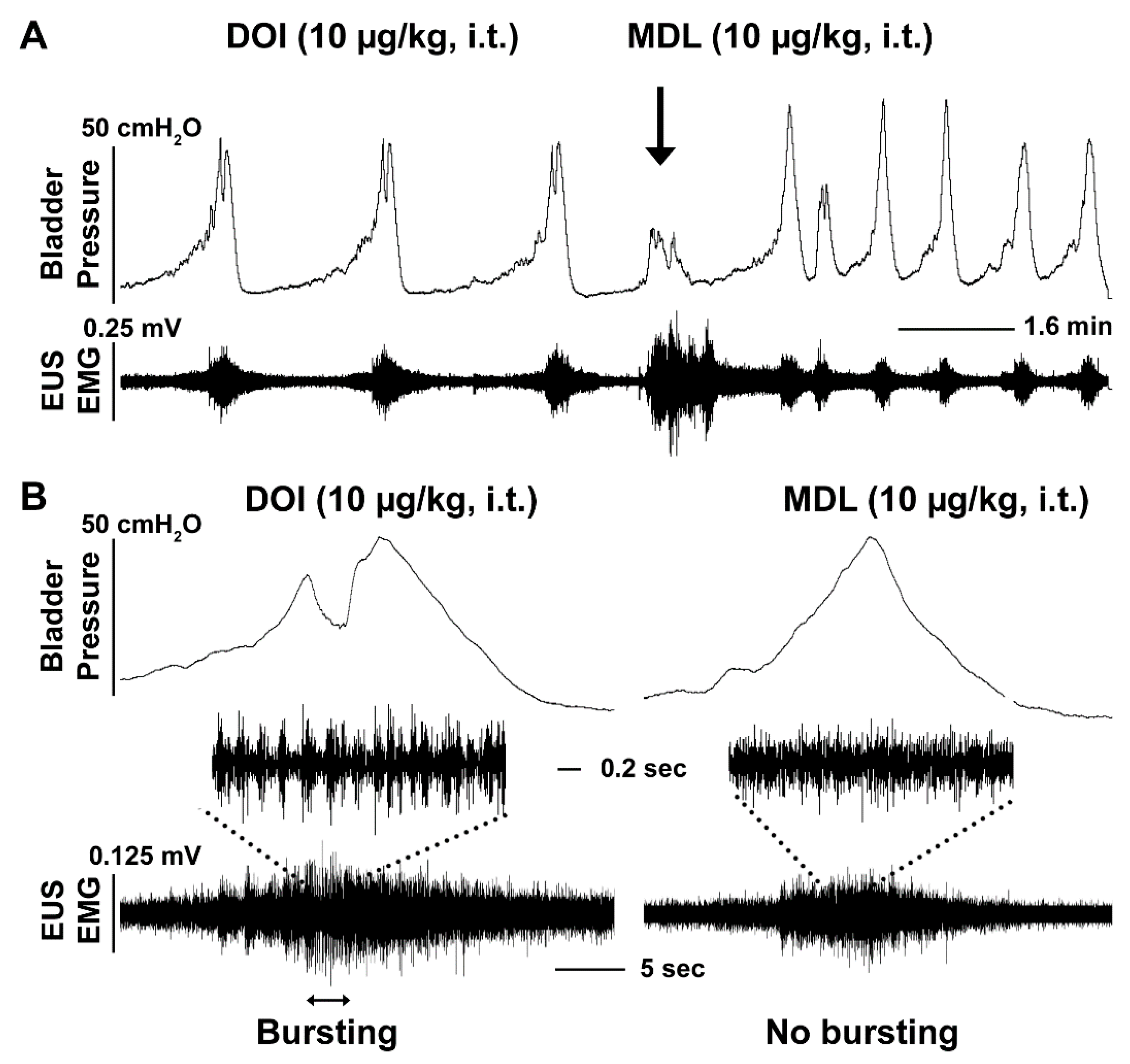
3.3. I.t. Drug Delivery and Its Effects on Bladder and EUS Reflexes
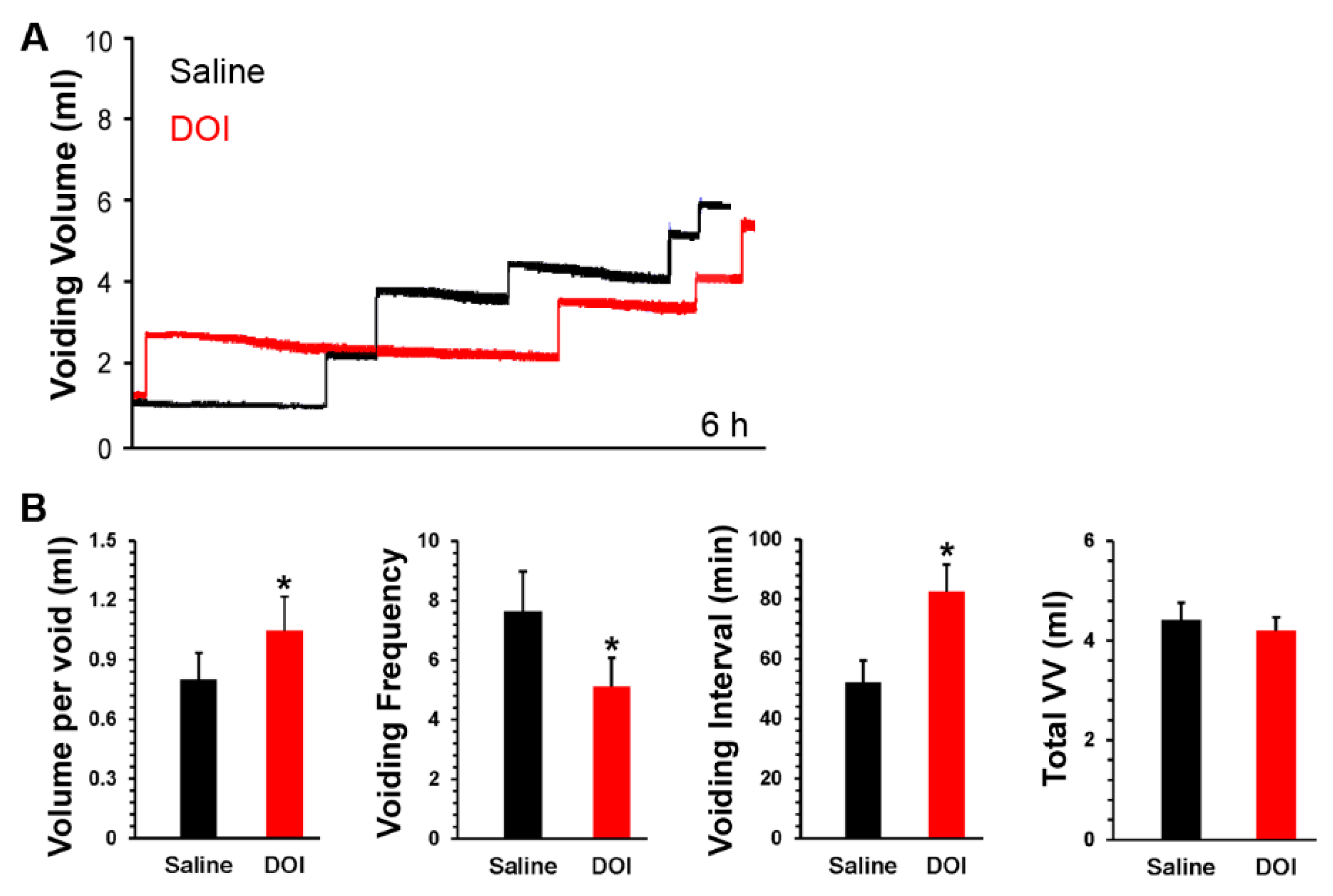
3.4. S.c. Delivery of DOI Improves Spontaneous Micturition Performance Following SCI
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nadelhaft, I.; Vera, P.L. Central nervous system neurons infected by pseudorabies virus injected into the rat urinary bladder following unilateral transection of the pelvic nerve. J. Comp. Neurol. 1995, 359, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, K.; Roppolo, J.R.; Yoshimura, N.; Card, J.P.; de Groat, W.C. The central neural pathways involved in micturition in the neonatal rat as revealed by the injection of pseudorabies virus into the urinary bladder. Neurosci. Lett. 1997, 223, 197–200. [Google Scholar] [CrossRef]
- Vizzard, M.A.; Erickson, V.L.; Card, J.P.; Roppolo, J.R.; de Groat, W.C. Transneuronal labeling of neurons in the adult rat brainstem and spinal cord after injection of pseudorabies virus into the urethra. J. Comp. Neurol. 1995, 355, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Mbaki, Y.; Ramage, A.G. Investigation of the role of 5-HT2 receptor subtypes in the control of the bladder and the urethra in the anaesthetized female rat. Br. J. Pharmacol. 2008, 155, 343–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norouzi-Javidan, A.; Javanbakht, J.; Barati, F.; Fakhraei, N.; Mohammadi, F.; Dehpour, A.R. Effect of 5-HT7 receptor agonist, LP-211, on micturition following spinal cord injury in male rats. Am. J. Transl. Res. 2016, 8, 2525–2533. [Google Scholar] [PubMed]
- De Groat, W.C. Influence of central serotonergic mechanisms on lower urinary tract function. Urology 2002, 59, 30–36. [Google Scholar] [CrossRef]
- Bowker, R.M.; Westlund, K.N.; Coulter, J.D. Origins of serotonergic projections to the spinal cord in rat: An immunocytochemical-retrograde transport study. Brain Res. 1981, 226, 187–199. [Google Scholar] [CrossRef]
- Ahn, J.; Saltos, T.M.; Tom, V.J.; Hou, S. Transsynaptic tracing to dissect supraspinal serotonergic input regulating the bladder reflex in rats. Neurourol. Urodyn. 2018, 37, 2487–2494. [Google Scholar] [CrossRef]
- Thor, K.B.; Nickolaus, S.; Helke, C.J. Autoradiographic localization of 5-hydroxytryptamine1A, 5-hydroxytryptamine1B and 5-hydroxytryptamine1C/2 binding sites in the rat spinal cord. Neuroscience 1993, 55, 235–252. [Google Scholar] [CrossRef]
- de Groat, W.C.; Yoshimura, N. Plasticity in reflex pathways to the lower urinary tract following spinal cord injury. Exp. Neurol. 2012, 235, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taweel, W.A.; Seyam, R. Neurogenic bladder in spinal cord injury patients. Res. Rep. Urol. 2015, 7, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Romo, P.G.B.; Smith, C.P.; Cox, A.; Averbeck, M.A.; Dowling, M.; Beckford, C.; Manohar, P.; Duran, S.; Cameron, A.P. Non-surgical urologic management of neurogenic bladder after spinal cord injury. World J. Urol. 2018, 36, 1555–1568. [Google Scholar] [CrossRef] [PubMed]
- Madersbacher, H.; Murtz, G.; Stohrer, M. Neurogenic detrusor overactivity in adults: A review on efficacy, tolerability and safety of oral antimuscarinics. Spinal Cord 2013, 51, 432–441. [Google Scholar] [CrossRef]
- Al Taweel, W.; Alzyoud, K.M. The effect of spinal cord-injury level on the outcome of neurogenic bladder treatment using OnabotulinumtoxinA. Urol. Ann. 2015, 7, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Schurch, B.; Schmid, D.M.; Stohrer, M. Treatment of neurogenic incontinence with botulinum toxin A. N. Engl. J. Med. 2000, 342, 665. [Google Scholar] [CrossRef]
- Jamil, F. Towards a catheter free status in neurogenic bladder dysfunction: A review of bladder management options in spinal cord injury (SCI). Spinal Cord 2001, 39, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Leu, R.; Stearns, G.L. Complications of Botox and their Management. Curr. Urol. Rep. 2018, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, T. Effect of chronic transection on dopamine, noradrenaline and 5-hydroxytryptamine in the rat spinal cord. Naunyn Schmiedebergs Arch. Pharmacol. 1973, 278, 13–22. [Google Scholar] [CrossRef]
- Newton, B.W.; Hamill, R.W. The morphology and distribution of rat serotoninergic intraspinal neurons: An immunohistochemical study. Brain Res. Bull. 1988, 20, 349–360. [Google Scholar] [CrossRef]
- Wienecke, J.; Ren, L.Q.; Hultborn, H.; Chen, M.; Møller, M.; Zhang, Y.; Zhang, M. Spinal cord injury enables aromatic L-amino acid decarboxylase cells to synthesize monoamines. J. Neurosci. 2014, 34, 11984–12000. [Google Scholar] [CrossRef] [Green Version]
- Sawynok, J.; Reid, A. Spinal supersensitivity to 5-HT1, 5-HT2 and 5-HT3 receptor agonists following 5,7-dihydroxytryptamine. Eur. J. Pharmacol. 1994, 264, 249–257. [Google Scholar] [CrossRef]
- Chen, J.; Gu, B.; Wu, G.; Tu, H.; Si, J.; Xu, Y.; Andersson, K.E. The effect of the 5-HT2A/2C receptor agonist DOI on micturition in rats with chronic spinal cord injury. J. Uro. 2013, 189, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; Condino, A.M.; Franceschetti, G.P.; Tonini, M. Characterization of prejunctional serotonin receptors modulating [3H]acetylcholine release in the human detrusor. J. Pharmacol. Exp. Ther. 2006, 316, 129–135. [Google Scholar] [CrossRef]
- Ochodnicky, P.; Humphreys, S.; Eccles, R.; Poljakovic, M.; Wiklund, P.; Michel, M.C. Expression profiling of G-protein-coupled receptors in human urothelium and related cell lines. BJU Int. 2012, 110, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto-Miyai, K.; Yoshizumi, M.; Kawatani, M. Regulatory Effects of 5-Hydroxytryptamine Receptors on Voiding Function. Adv. Ther. 2015, 32, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, M.; Nezu, F.M.; Yokoyama, O.; de Groat, W.C.; Chancellor, M.B. Changes in micturition after spinal cord injury in conscious rats. Urology 1999, 54, 929–933. [Google Scholar] [CrossRef]
- Kadekawa, K.; Nishijima, S.; Sugaya, K.; Miyazato, M.; Saito, S. Mechanisms by which the serotonergic system inhibits micturition in rats. Life Sci. 2009, 85, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, T.; Shumsky, J.S.; Lepore, A.C.; Murray, M.; Fischer, I. Transplantation of neuronal and glial restricted precursors into contused spinal cord improves bladder and motor functions, decreases thermal hypersensitivity, and modifies intraspinal circuitry. J. Neurosci. 2005, 25, 9624–9636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlstroem, A.; Fuxe, K. Evidence for the Existence of Monoamine-Containing Neurons in the Central Nervous System. I. Demonstration of Monoamines in the Cell Bodies of Brain Stem Neurons. Acta Physiol. Scand. Suppl. 1964, 232, 1–55. [Google Scholar]
- Dahlstroem, A.; Fuxe, K. Evidence for the Existence of Monoamine Neurons in the Central Nervous System. Ii. Experimentally Induced Changes in the Intraneuronal Amine Levels of Bulbospinal Neuron Systems. Acta Physiol. Scand. Suppl. 1965, 247, 1–36. [Google Scholar]
- Xu, C.; Giuliano, F.; Sun, X.Q.; Brisorgueil, M.-J.; Leclerc, P.; Vergé, D.; Conrath, M. Serotonin 5-HT2A and 5-HT5A receptors are expressed by different motoneuron populations in rat Onuf’s nucleus. J. Comp. Neurol. 2007, 502, 620–634. [Google Scholar] [CrossRef]
- Helton, L.; Thor, K.B.; Baez, M. 5-Hydroxytryptamine2a, 5-Hydroxytryptamine2B, 5-Hydroxytryptamine2C receptor mRNA expression in the spinal cord of rat, cat, monkey, and human. Neuroreport 1994, 5, 2617–2620. [Google Scholar] [CrossRef]
- Kong, X.Y.; Wienecke, J.; Hultborn, H.; Zhang, M. Robust upregulation of serotonin 2A receptors after chronic spinal transection of rats: An immunohistochemical study. Brain Res. 2010, 1320, 60–68. [Google Scholar] [CrossRef]
- Chang, H.Y.; Cheng, C.L.; Chen, J.-J.J.; de Groat, W.C. Roles of glutamatergic and serotonergic mechanisms in reflex control of the external urethral sphincter in urethane-anesthetized female rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R224–R234. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.Y.; Cheng, C.L.; Chen, J.-J.J.; de Groat, W.C. Serotonergic drugs and spinal cord transections indicate that different spinal circuits are involved in external urethral sphincter activity in rats. Am. J. Physiol Renal Physiol 2007, 292, F1044–F1053. [Google Scholar] [CrossRef]
- Cheng, C.L.; de Groat, W.C. Role of 5-HT1A receptors in control of lower urinary tract function in anesthetized rats. Am. J. Physiol. Renal. Physiol. 2010, 298, F771–F778. [Google Scholar] [CrossRef] [Green Version]
- Dolber, P.C.; Gu, B.; Zhang, X.; Fraser, M.O.; Thor, K.B.; Reiter, J.P. Activation of the external urethral sphincter central pattern generator by a 5-HT(1A) receptor agonist in rats with chronic spinal cord injury. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1699–R1706. [Google Scholar] [CrossRef]
- Ramage, A.G. The role of central 5-hydroxytryptamine (5-HT, serotonin) receptors in the control of micturition. Br. J. Pharmacol. 2006, 147, S120–S131. [Google Scholar] [CrossRef]
- Cao, N.; Ni, J.; Wang, X.; Tu, H.; Gu, B.; Si, J.; Wu, G.; Andersson, K.E. Chronic spinal cord injury causes upregulation of serotonin (5-HT)2A and 5-HT2C receptors in lumbosacral cord motoneurons. BJU Int. 2018, 121, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Doly, S.; Madeira, A.; Fischer, J.; Brisorgueil, M.J.; Daval, G.; Bernard, R.; Vergé, D.; Conrath, M. The 5-HT2A receptor is widely distributed in the rat spinal cord and mainly localized at the plasma membrane of postsynaptic neurons. J. Comp. Neurol. 2004, 472, 496–511. [Google Scholar] [CrossRef]
- Kruse, M.N.; Belton, A.L.; de Groat, W.C. Changes in bladder and external urethral sphincter function after spinal cord injury in the rat. Am. J. Physiol. 1993, 264, R1157–R1163. [Google Scholar] [CrossRef] [PubMed]
- Kakizaki, H.; Fraser, M.O.; De Groat, W.C. Reflex pathways controlling urethral striated and smooth muscle function in the male rat. Am. J. Physiol. 1997, 272, R1647–R1656. [Google Scholar] [CrossRef] [PubMed]
- Maggi, C.A.; Giuliani, S.; Santicioli, P.; Meli, A. Analysis of factors involved in determining urinary bladder voiding cycle in urethan-anesthetized rats. Am. J. Physiol. 1986, 251, R250–R257. [Google Scholar] [CrossRef]
- Danuser, H.; Thor, K.B. Spinal 5-HT2 receptor-mediated facilitation of pudendal nerve reflexes in the anaesthetized cat. Br. J. Pharmacol. 1996, 118, 150–154. [Google Scholar] [CrossRef] [Green Version]
- Karnup, S.V.; de Groat, W.C. Propriospinal Neurons of L3-L4 Segments Involved in Control of the Rat External Urethral Sphincter. Neuroscience 2020, 425, 12–28. [Google Scholar] [CrossRef]
- Hoyer, D. Molecular pharmacology and biology of 5-HT1C receptors. Trends Pharmacol. Sci. 1988, 9, 89–94. [Google Scholar] [CrossRef]
- Lopez-Gimenez, J.F.; Vilaró, M.T.; Palacios, J.M.; Mengod, G. Multiple conformations of 5-HT2A and 5-HT 2C receptors in rat brain: An autoradiographic study with [125I](+/-)DOI. Exp. Brain Res. 2013, 230, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Gimenez, J.F.; Mengod, G.; Palacios, J.M.; Vilaró, M.T. Selective visualization of rat brain 5-HT2A receptors by autoradiography with [3H]MDL 100,907. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 356, 446–454. [Google Scholar] [CrossRef]
- Wang, X.; Cao, N.; Ni, J.; Si, J.; Gu, B.; Karl-Erik, A. Effect of 5-HT2A receptor antagonist ketanserin on micturition in male rats. Neurosci. Lett. 2018, 687, 196–201. [Google Scholar] [CrossRef]
- Brouillard, C.B.J.; Crook, J.J.; Lovick, T.A. Suppression of Urinary Voiding “on Demand” by High-Frequency Stimulation of the S1 Sacral Nerve Root in Anesthetized Rats. Neuromodulation 2019, 22, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, M.; Roppolo, J.R.; De Groat, W.C. Alteration by urethane of glutamatergic control of micturition. Eur. J. Pharmacol. 1994, 264, 417–425. [Google Scholar] [CrossRef]
- Morikawa, K.; Ichihashi, M.; Kakiuchi, M.; Yamauchi, T.; Kato, H.; Ito, Y.; Gomi, Y. Effects of various drugs on bladder function in conscious rats. Jpn. J. Pharmacol. 1989, 50, 369–376. [Google Scholar] [CrossRef]
- Hou, S.; Rabchevsky, A.G. Autonomic consequences of spinal cord injury. Compr. Physiol. 2014, 4, 1419–1453. [Google Scholar] [PubMed]
- Van Middendorp, J.J.; Goss, B.; Urquhart, S.; Atresh, S.; Williams, R.P.; Schuetz, M. Diagnosis and prognosis of traumatic spinal cord injury. Global. Spine J. 2011, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wyndaele, M.; Wyndaele, J.J. Incidence, prevalence and epidemiology of spinal cord injury: What learns a worldwide literature survey? Spinal Cord 2006, 44, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Abelson, B.; Sun, D.; Que, L.; Nebel, R.A.; Baker, D.; Popiel, P.; Amundsen, C.L.; Chai, T.; Close, C.; DiSanto, M.; et al. Sex differences in lower urinary tract biology and physiology. Biol. Sex. Differ. 2018, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yang, L.; Pu, C.; Tang, Y.; Yun, H.; Han, P. The role of duloxetine in stress urinary incontinence: A systematic review and meta-analysis. Int. Urol. Nephrol. 2013, 45, 679–686. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs | Doses (/kg) | Bladder CMG | EUS EMG | VV | ||||
|---|---|---|---|---|---|---|---|---|
| VA | CD | VI | # NVCs | RMS | MA | |||
| MDL (n = 6) (5HT2A receptor antagonist) | Saline | 1.11 ± 0.10 | 0.96 ± 0.05 | 1.00 ± 0.05 | 1.31 ± 0.31 | 0.95 ± 0.06 | 0.82 ± 0.12 | 0.95 ± 0.09 |
| 20 µg | 1.01 ± 0.19 | 0.93 ± 0.05 | 1.07 ± 0.14 | 1.25 ± 0.70 | 0.87 ± 0.02 | 0.71 ± 0.09 | 1.00 ± 0.15 | |
| 0.1 mg | 0.97 ± 0.06 | 0.98 ± 0.06 | 1.22 ± 0.11 * | 1.56 ± 0.86 | 0.96 ± 0.09 | 0.81 ± 0.10 | 1.15 ± 0.10 | |
| 0.5 mg | 1.04 ± 0.04 | 0.95 ± 0.06 | 1.10 ± 0.07 | 0.88 ± 0.79 | 1.11 ± 0.16 | 1.00 ± 0.37 | 1.09 ± 0.14 | |
| DOI (n = 6) (5HT2A receptor agonist) | Saline | 1.11 ± 0.10 | 0.96 ± 0.05 | 1.00 ± 0.05 | 1.31 ± 0.31 | 0.95 ± 0.06 | 0.82 ± 0.12 | 0.95 ± 0.09 |
| 5.0 µg | 1.08 ± 0.04 | 0.98 ± 0.04 | 1.34 ± 0.11 | 1.00 ± 0.84 | 1.03 ± 0.14 | 0.69 ± 0.97 | 1.23 ± 0.15 | |
| 20 µg | 1.06 ± 0.08 | 1.12 ± 0.04 | 1.73 ± 0.17 * | 2.19 ± 1.59 | 1.13 ± 0.13 | 0.84 ± 0.21 | 1.32 ± 0.12 * | |
| 0.1 mg | 1.00 ± 0.08 | 1.01 ± 0.04 | 1.51 ± 0.30 | 1.31 ± 0.80 | 1.53 ± 0.24 * | 1.64 ± 0.55 | 1.24 ± 0.17 | |
| Drugs | Doses (/kg) | Bladder CMG | EUS EMG | VV | ||||
|---|---|---|---|---|---|---|---|---|
| VA | CD | VI | # NVCs | RMS | MA | |||
| MDL (n = 6) (5HT2A receptor antagonist) | Saline | 1.23 ± 0.11 | 1.01 ± 0.01 | 1.12 ± 0.11 | 0.29 ± 0.15 | 0.97 ± 0.19 | 0.92 ± 0.07 | 0.81 ± 0.03 |
| 2 µg | 1.13 ± 0.10 | 0.97 ± 0.01 | 1.12 ± 0.11 | 0.20 ± 0.13 | 1.09 ± 0.06 | 0.95 ± 0.08 | 0.78 ± 0.03 | |
| 10 µg | 1.22 ± 0.18 | 1.08 ± 0.09 | 1.34 ± 0.20 | 0.29 ± 0.15 | 0.98 ± 0.05 | 0.74 ± 0.11 | 1.07 ± 0.09 | |
| 50 µg | 0.81 ± 0.08 | 0.83 ± 0.05 | 1.19 ± 0.15 | 0.58 ± 0.26 | 1.11 ± 0.05 | 1.46 ± 0.30 | 0.95 ± 0.09 | |
| DOI (n = 6) (5HT2A receptor agonist) | Saline | 0.94 ± 0.03 | 1.09 ± 0.04 | 1.24 ± 0.14 | 1.08 ± 0.60 | 1.01 ± 0.03 | 1.17 ± 0.20 | 0.95 ± 0.10 |
| 0.5 µg | 1.03 ± 0.09 | 1.14 ± 0.05 | 1.42 ± 0.46 | 0.79 ± 0.64 | 1.01 ± 0.03 | 1.05 ± 0.09 | 0.88 ± 0.12 | |
| 2 µg | 1.21 ± 0.13 | 1.13 ± 0.08 | 3.27 ± 1.04 * | 2.70 ± 1.77 | 1.14 ± 0.17 | 1.12 ± 0.28 | 1.14 ± 0.18 | |
| 10 µg | 1.12 ± 0.11 | 1.09 ± 0.05 | 3.98 ± 1.58 * | 3.33 ± 2.41 | 1.46 ± 0.21 | 1.96 ± 0.31 | 1.33 ± 0.18 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeFinis, J.H.; Weinberger, J.; Hou, S. Delivery of the 5-HT2A Receptor Agonist, DOI, Enhances Activity of the Sphincter Muscle during the Micturition Reflex in Rats after Spinal Cord Injury. Biology 2021, 10, 68. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010068
DeFinis JH, Weinberger J, Hou S. Delivery of the 5-HT2A Receptor Agonist, DOI, Enhances Activity of the Sphincter Muscle during the Micturition Reflex in Rats after Spinal Cord Injury. Biology. 2021; 10(1):68. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010068
Chicago/Turabian StyleDeFinis, Jaclyn H., Jeremy Weinberger, and Shaoping Hou. 2021. "Delivery of the 5-HT2A Receptor Agonist, DOI, Enhances Activity of the Sphincter Muscle during the Micturition Reflex in Rats after Spinal Cord Injury" Biology 10, no. 1: 68. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010068