
Antiviral Function of NKEF against VHSV in Rainbow Trout

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Statements
2.2. Cell Culture and Virus
2.3. Time Course of NKEF Expression in VHSV-Exposed RBCs and RTG-2 Cells
2.4. VHSV Challenged of Rainbow Trout
2.5. NKEF siRNA Assay
2.6. Evaluation of VHSV Infectivity in Cells Treated with siNKEF
2.7. RNA Isolation, RT-PCR, and RT-qPCR
2.8. Flow Cytometry
2.9. nkef cDNA Cloning in pmTFP Plasmid
2.10. Transfection Assays of RBCs and EPC Cells with pmTFP1-NKEF
2.11. VHSV Infectivity in pmTFP1-NKEF-Transfected EPC Cells
2.12. Software and Statistics
3. Results
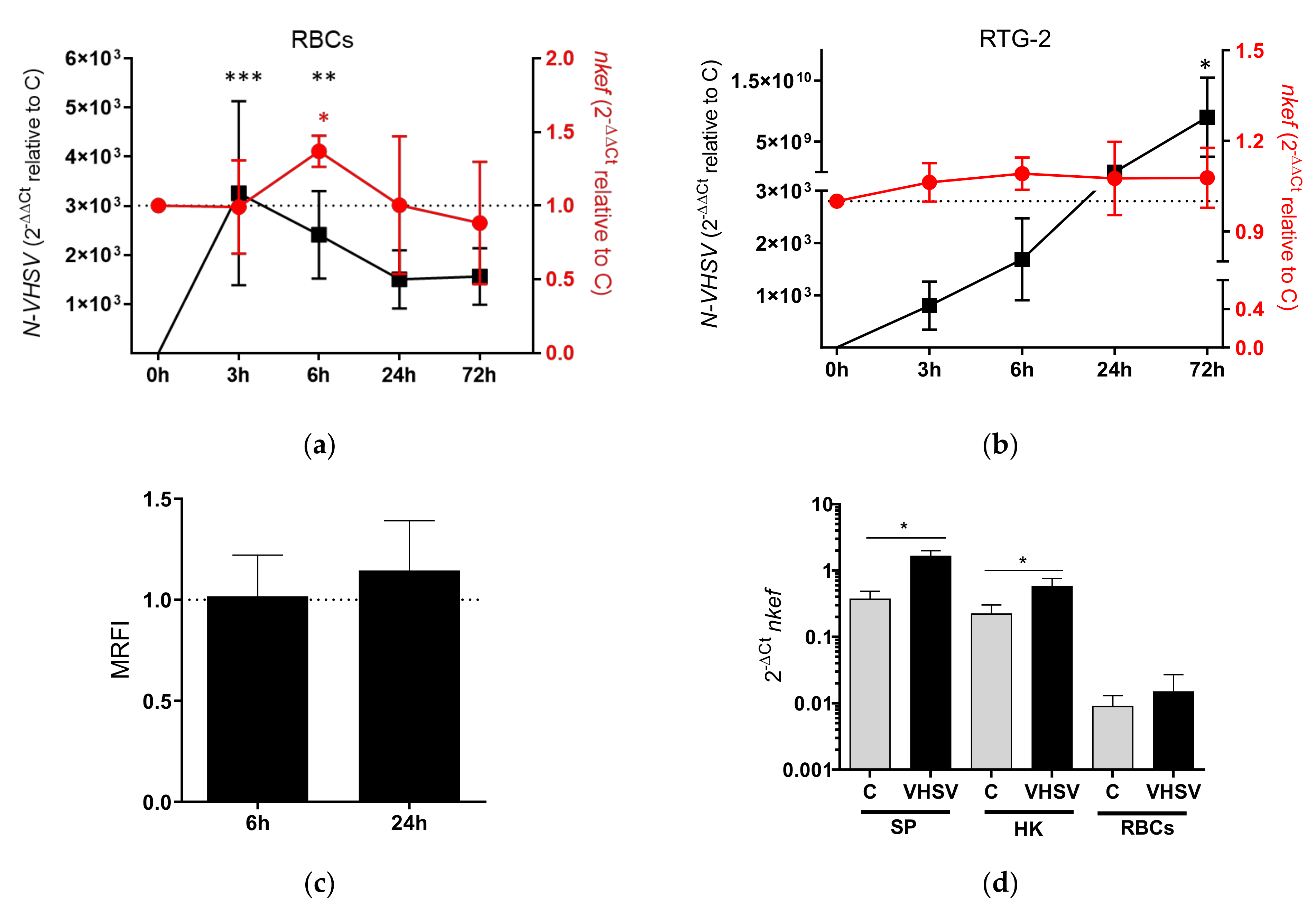
3.1. Time Course of NKEF Expression in RBCs Exposed to VHSV
3.2. nkef Gene Silencing in RBCs and RTG-2 Cell Line
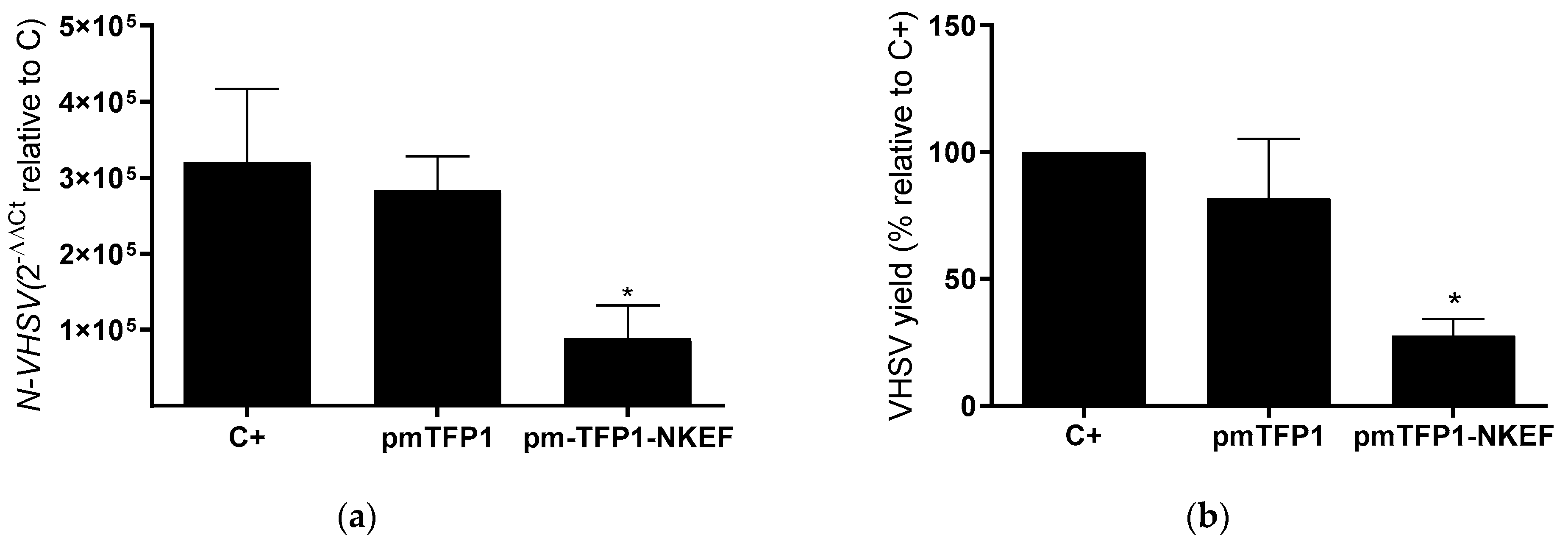
3.3. Overexpression of NKEF in RBCs
3.4. Evaluation of NKEF Overexpression on VHSV Infectivity in EPC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shau, H.; Butterfield, L.H.; Chiu, R.; Kim, A. Cloning and sequence analysis of candidate human natural killer-enhancing factor genes. Immunogenetics 1994, 40, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, I.H.; Lee, K.Y.; Rhee, S.G.; Stadtman, E.R. The isolation and purification of a specific “protector” protein which inhibits enzyme inactivation by a thiol/Fe(III)/O2 mixed-function oxidation system. J. Biol. Chem. 1988, 263, 4704–4711. [Google Scholar] [CrossRef]
- Shau, H.; Gupta, R.K.; Golub, S.H. Identification of a natural killer enhancing factor (NKEF) from human erythroid cells. Cell. Immunol. 1993, 147, 1–11. [Google Scholar] [CrossRef]
- Shau, H.; Kim, A. Identification of natural killer enhancing factor as a major antioxidant in human red blood cells. Biochem. Biophys. Res. Commun. 1994, 199, 83–88. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, B.; Kang, S.W.; Seo, M.S.; Rhee, S.G.; Obeid, L.M. Thioredoxin peroxidase is a novel inhibitor of apoptosis with a mechanism distinct from that of Bcl-2. J. Biol. Chem. 1997, 272, 30615–30618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, T.H.; Park, E.S.; Suh, J.M.; Park, S.J.; Chung, H.K.; Kwon, O.Y.; Kim, Y.K.; Ro, H.K.; Shong, M. Role of peroxiredoxins in regulating intracellular hydrogen peroxide and hydrogen peroxide-induced apoptosis in thyroid cells. J. Biol. Chem. 2000, 275, 18266–18270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Matsui, Y.; Natori, S.; Obinata, M. Cloning of a housekeeping-type gene (MER5) preferentially expressed in murine erythroleukemia cells. Gene 1989, 80, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Prosperi, M.T.; Ferbus, D.; Karczinski, I.; Goubin, G. A human cDNA corresponding to a gene overexpressed during cell proliferation encodes a product sharing homology with amoebic and bacterial proteins. J. Biol. Chem. 1993, 268, 11050–11056. [Google Scholar] [CrossRef]
- Ishii, T.; Warabi, E.; Yanagawa, T. Novel roles of peroxiredoxins in inflammation, cancer and innate immunity. J. Clin. Biochem. Nutr. 2012, 50, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T. Close teamwork between Nrf2 and peroxiredoxins 1 and 6 for the regulation of prostaglandin D2 and E2 production in macrophages in acute inflammation. Free Radic. Biol. Med. 2015, 88, 189–198. [Google Scholar] [CrossRef]
- Geiben-Lynn, R.; Kursar, M.; Brown, N.V.; Addo, M.M.; Shau, H.; Lieberman, J.; Luster, A.D.; Walker, B.D. HIV-1 antiviral activity of recombinant natural killer cell enhancing factors, NKEF-A and NKEF-B, members of the peroxiredoxin family. J. Biol. Chem. 2003, 278, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Mourich, D.V.; Hansen, J.; Leong, J. Natural killer cell enhancement factor-like gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics 1995, 42, 438–439. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Martinez-Morcillo, F.J.; Esteban, M.A.; Chaves-Pozo, E.; Cuesta, A. Fish Peroxiredoxins and Their Role in Immunity. Biology 2015, 4, 860–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, A.; Tafalla, C. Transcription of immune genes upon challenge with viral hemorrhagic septicemia virus (VHSV) in DNA vaccinated rainbow trout (Oncorhynchus mykiss). Vaccine 2009, 27, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Utke, K.; Bergmann, S.; Lorenzen, N.; Kollner, B.; Ototake, M.; Fischer, U. Cell-mediated cytotoxicity in rainbow trout, Oncorhynchus mykiss, infected with viral haemorrhagic septicaemia virus. Fish Shellfish Immunol. 2007, 22, 182–196. [Google Scholar] [CrossRef]
- Huang, R.; Gao, L.Y.; Wang, Y.P.; Hu, W.; Guo, Q.L. Structure, organization and expression of common carp (Cyprinus carpio L.) NKEF-B gene. Fish Shellfish Immunol. 2009, 26, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Ordas, M.C.; Cuesta, A.; Mercado, L.; Bols, N.C.; Tafalla, C. Viral hemorrhagic septicaemia virus (VHSV) up-regulates the cytotoxic activity and the perforin/granzyme pathway in the rainbow trout RTS11 cell line. Fish Shellfish Immunol. 2011, 31, 252–259. [Google Scholar] [CrossRef]
- Nombela, I.; Puente-Marin, S.; Chico, V.; Villena, A.J.; Carracedo, B.; Ciordia, S.; Mena, M.C.; Mercado, L.; Perez, L.; Coll, J.; et al. Identification of diverse defense mechanisms in rainbow trout red blood cells in response to halted replication of VHS virus. F1000Research 2017, 6, 1958. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.A.; Chaves-Pozo, E.; Arizcun, M.; Meseguer, J.; Cuesta, A. Regulation of natural killer enhancing factor (NKEF) genes in teleost fish, gilthead seabream and European sea bass. Mol. Immunol. 2013, 55, 275–282. [Google Scholar] [CrossRef]
- Chico, V.; Salvador-Mira, M.E.; Nombela, I.; Puente-Marin, S.; Ciordia, S.; Mena, M.C.; Perez, L.; Coll, J.; Guzman, F.; Encinar, J.A.; et al. IFIT5 Participates in the Antiviral Mechanisms of Rainbow Trout Red Blood Cells. Front. Immunol. 2019, 10, 613. [Google Scholar] [CrossRef] [Green Version]
- Nombela, I.; Lopez-Lorigados, M.; Salvador-Mira, M.E.; Puente-Marin, S.; Chico, V.; Ciordia, S.; Mena, M.C.; Mercado, L.; Coll, J.; Perez, L.; et al. Integrated Transcriptomic and Proteomic Analysis of Red Blood Cells from Rainbow Trout Challenged with VHSV Point Towards Novel Immunomodulant Targets. Vaccines 2019, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Nombela, I.; Requena-Platek, R.; Morales-Lange, B.; Chico, V.; Puente-Marin, S.; Ciordia, S.; Mena, M.C.; Coll, J.; Perez, L.; Mercado, L.; et al. Rainbow Trout Red Blood Cells Exposed to Viral Hemorrhagic Septicemia Virus Up-Regulate Antigen-Processing Mechanisms and MHC I&II, CD86, and CD83 Antigen-presenting Cell Markers. Cells 2019, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Nombela, I.; Carrion, A.; Puente-Marin, S.; Chico, V.; Mercado, L.; Perez, L.; Coll, J.; Ortega-Villaizan, M.d.M. Infectious pancreatic necrosis virus triggers antiviral immune response in rainbow trout red blood cells, despite not being infective. F1000Research 2017, 6, 1968. [Google Scholar] [CrossRef]
- LeBerre, M.; De Kinkelin, P.; Metzger, A. Identification sérologique des rhabdovirus des salmonidés. Bull. Off. Int. Epizoot. 1977, 87, 391–393. [Google Scholar]
- Basurco, B.; Coll, J.M. Spanish isolates and reference strains of viral haemorrhagic septicaemia virus shown similar protein size patterns. Bull. Eur. Assoc. Fish Pathol. 1989, 9, 92–95. [Google Scholar]
- Bethke, J.; Rojas, V.; Berendsen, J.; Cardenas, C.; Guzman, F.; Gallardo, J.A.; Mercado, L. Development of a new antibody for detecting natural killer enhancing factor (NKEF)-like protein in infected salmonids. J. Fish Dis. 2012, 35, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Chico, V.; Gomez, N.; Estepa, A.; Perez, L. Rapid detection and quantitation of viral hemorrhagic septicemia virus in experimentally challenged rainbow trout by real-time RT-PCR. J. Virol. Methods 2006, 132, 154–159. [Google Scholar] [CrossRef]
- Wang, T.; Johnson, N.; Zou, J.; Bols, N.; Secombes, C.J. Sequencing and expression of the second allele of the interleukin-1beta1 gene in rainbow trout (Oncorhynchus mykiss): Identification of a novel SINE in the third intron. Fish Shellfish Immunol. 2004, 16, 335–358. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Temperature-dependent expression of immune-relevant genes in rainbow trout following Yersinia ruckeri vaccination. Dis. Aquat. Organ. 2007, 77, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Villaizan, M.; Chico, V.; Martinez-Lopez, A.; Falco, A.; Perez, L.; Coll, J.M.; Estepa, A. In vitro analysis of the factors contributing to the antiviral state induced by a plasmid encoding the viral haemorrhagic septicaemia virus glycoprotein G in transfected trout cells. Vaccine 2011, 29, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Purcell, M.K.; Smith, K.D.; Hood, L.; Winton, J.R.; Roach, J.C. Conservation of Toll-Like Receptor Signaling Pathways in Teleost Fish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Garcia, I.; Galiana, A.; Falco, A.; Estepa, A.; Perez, L. Characterization of an infectious pancreatic necrosis (IPN) virus carrier cell culture with resistance to superinfection with heterologous viruses. Vet. Microbiol. 2011, 149, 48–55. [Google Scholar] [CrossRef]
- Xinxian, W.; Peng, J.; Guixiang, T.; Jinjin, W.; Xiaocong, Z.; Junqiang, H.; Xianle, Y.; Hong, L. Effect of common carp (Cyprinus carpio) TLR9 overexpression on the expression of downstream interferon-associated immune factor mRNAs in epithelioma papulosum cyprini cells. Vet. Immunol. Immunopathol. 2016, 170, 47–53. [Google Scholar] [CrossRef]
- Ai, H.W.; Henderson, J.N.; Remington, S.J.; Campbell, R.E. Directed evolution of a monomeric, bright and photostable version of Clavularia cyan fluorescent protein: Structural characterization and applications in fluorescence imaging. Biochem. J. 2006, 400, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Lopez, A.; Fernandez-Alonso, M.; Rocha, A.; Estepa, A.; Coll, J. Transfection of epithelioma papulosum cyprini (EPC) carp cells. Biotechnol. Lett. 2001, 23, 481–487. [Google Scholar] [CrossRef]
- Sanz, F.; Basurco, B.; Babin, M.; Dominguez, J.; Coll, J.M. Monoclonal antibodies against the structural proteins of viral haemorrhagic septicaemia virus isolates. J. Fish Dis. 1993, 16, 53–63. [Google Scholar] [CrossRef]
- Olesen, N.J.; Jørgensen, P.E.V. Comparative susceptibility of three fish cell lines to Egtved virus, the virus of viral haemorrhagic septicaemia (VHS). Dis. Aquat. Org. 1992, 12, 235–237. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018: Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Crane, M.; Hyatt, A. Viruses of fish: An overview of significant pathogens. Viruses 2011, 3, 2025–2046. [Google Scholar] [CrossRef]
- Morera, D.; Mackenzie, S.A. Is there a direct role for erythrocytes in the immune response? Vet. Res. 2011, 42, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nombela, I.; Ortega-Villaizan, M.D.M. Nucleated red blood cells: Immune cell mediators of the antiviral response. PLoS Pathog. 2018, 14, e1006910. [Google Scholar] [CrossRef]
- Madusanka, R.K.; Tharuka, M.D.N.; Madhuranga, W.S.P.; Lee, S.; Lee, J. Transcriptional modifications and the cytoprotective, DNA protective, and wound healing effects of peroxiredoxin-1 from Sebastes schlegelii. Fish Shellfish Immunol. 2020, 107, 73–83. [Google Scholar] [CrossRef]
- Kjærner-Semb, E.; Ayllon, F.; Furmanek, T.; Wennevik, V.; Dahle, G.; Niemelä, E.; Ozerov, M.; Vähä, J.-P.; Glover, K.A.; Rubin, C.J.; et al. Atlantic salmon populations reveal adaptive divergence of immune related genes—A duplicated genome under selection. BMC Genom. 2016, 17, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Gan, X.; Ito, M.; Chen, M.; Aly, H.H.; Matsui, C.; Abe, T.; Watashi, K.; Wakita, T.; Suzuki, T.; et al. Peroxiredoxin 1, a Novel HBx-Interacting Protein, Interacts with Exosome Component 5 and Negatively Regulates Hepatitis B Virus (HBV) Propagation through Degradation of HBV RNA. J. Virol. 2019, 93, e02203-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Li, N.; Roller, R.J. Peroxiredoxin 1 protein interacts with influenza virus ribonucleoproteins and is required for efficient virus replication. Vaccine 2018, 36, 4540–4547. [Google Scholar] [CrossRef]
- Chang, C.J.; Robertsen, C.; Sun, B.; Robertsen, B. Protection of Atlantic salmon against virus infection by intramuscular injection of IFNc expression plasmid. Vaccine 2014, 32, 4695–4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Yan, J.; Chen, H.; Li, J.; Tian, Y.; Tang, L.; Feng, H. Mx1 of black carp functions importantly in the antiviral innate immune response. Fish Shellfish Immunol. 2016, 58, 584–592. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Y.H.; Xiao, Z.Z.; Sun, L. Megalocytivirus-induced proteins of turbot (Scophthalmus maximus): Identification and antiviral potential. J. Proteom. 2013, 91, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Rual, J.-F.; Venkatesan, K.; Hao, T.; Hirozane-Kishikawa, T.; Dricot, A.; Li, N.; Berriz, G.F.; Gibbons, F.D.; Dreze, M.; Ayivi-Guedehoussou, N.; et al. Towards a proteome-scale map of the human protein–protein interaction network. Nature 2005, 437, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Denison, C.; Huibregtse, J.M.; Gygi, S.; Krug, R.M. Human ISG15 conjugation targets both IFN-induced and constitutively expressed proteins functioning in diverse cellular pathways. Proc. Natl. Acad. Sci. USA 2005, 102, 10200–10205. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | siRNA Design Sequence (5′–3′) | Start on Target |
|---|---|---|
| siNKEF-1 sense | CGUAUAGCUUGGAGAUGUUdTdT | 840 |
| siNKEF-1 antisense | AACAUCUCCAAGCUAUACGdTdT | 840 |
| siNKEF-2 sense | CAAACUAUGAAGAUUAUAUdTdT | 818 |
| siNKEF-2 antisense | AUAUAAUCUUCAUAGUUUGdTdT | 818 |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Annealing, Ta (°C) | Reference or Accession Number |
|---|---|---|---|---|
| nkef | TCCAAGCAGCAGTAAGACGA | CATGAGATAAGGGGATGCTGA | 60 | AF250194.1 |
| gapdh | ATGTCAGACCTCTGTGTTGG | TCCTCGATGCCGAAGTTGTCG | 52 | [28] |
| Β-actin | AAGTGTGACGTGGACATCCG | CAGAGCTGAAGTGGTAGTCGG | 60 | NM_001124235.1 |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Probe (5′–3′) | References or Accession Number |
|---|---|---|---|---|
| Om-nkef | CGCTGGACTTCACCTTTGTGT | ACCTCACAACCGATCTTCCTAAAC | - | [18] |
| N-VHSV | GACTCAACGGGACAGGAATGA | GGGCAATGCCCAAGTTGTT | TGGGTTGTTCACCCAGGCCGC | [27] |
| Om-ef1α | ACCCTCCTCTTGGTCGTTTC | TGATGACACCAACAGCAACA | GCTGTGCGTGACATGAGGCA | [29] |
| Om-mx | TGAAGCCCAGGATGAAATGG | TGGCAGGTCGATGAGTGTGA | ACCTCATCAGCCTAGAGATTGGCTCCCC | [30] |
| Om-isg15 | GTTAGGCGTCAATGGGAACAA | GGCCATAGTCGCTCAAAGTTTT | - | XM_036979883.1 |
| Om-irf3 | AACAAGGCATGCAGGGTTCTAAAT | ACGTGTGCAATCAGTACCAGCA | - | [31] |
| Carp-ef1α | CTGGAGGCCAGCTCAAACT | CATTTCCCTCCTTACGCTCAAC | - | AY643400 |
| Carp-mx | GGA GAA GAG GTT AAA TGT GGA TCA G | TGA CCG AAT CAA GAA GTC ATT CC | - | [32] |
| Carp-isg15 | TAATGCCACAGTCGGTGAA | AGGTCCAGTGTTAGTGATGAGC | [33] | |
| Carp-irf3 | GTTTAGAGGGACAATTAACTGGACTA | GAGGGTCCACTCTTTGAAAATG | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chico, V.; Salvador-Mira, M.E.; Nombela, I.; Puente-Marin, S.; Perez, L.; Mercado, L.; Ortega-Villaizan, M.d.M. Antiviral Function of NKEF against VHSV in Rainbow Trout. Biology 2021, 10, 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10101045
Chico V, Salvador-Mira ME, Nombela I, Puente-Marin S, Perez L, Mercado L, Ortega-Villaizan MdM. Antiviral Function of NKEF against VHSV in Rainbow Trout. Biology. 2021; 10(10):1045. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10101045
Chicago/Turabian StyleChico, Veronica, Maria Elizabhet Salvador-Mira, Ivan Nombela, Sara Puente-Marin, Luis Perez, Luis Mercado, and Maria del Mar Ortega-Villaizan. 2021. "Antiviral Function of NKEF against VHSV in Rainbow Trout" Biology 10, no. 10: 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10101045