
Comprehensive Analysis of CPA4 as a Poor Prognostic Biomarker Correlated with Immune Cells Infiltration in Bladder Cancer

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. CPA4 Methylation Level Analysis
2.3. Analysis of Differentially Expressed Genes (DEGs)
2.4. Gene Set Enrichment Analysis (GSEA)
2.5. Comprehensive Analysis of Protein–Protein Interaction
2.6. Analysis of the Tumor Immune Estimation Resource (TIMER2)
2.7. Univariate and Multivariate Logistic Regression Analysis
2.8. Identification of CPA4 Coexpression Genes and Construction of a Prognostic Nomogram
2.9. Immune Infiltration Analysis by ssGSEA
2.10. Analysis of the Gene Expression Profiling Interactive Analysis 2
2.11. Statistical Analysis
3. Results
3.1. Characteristics of BLCA Patients
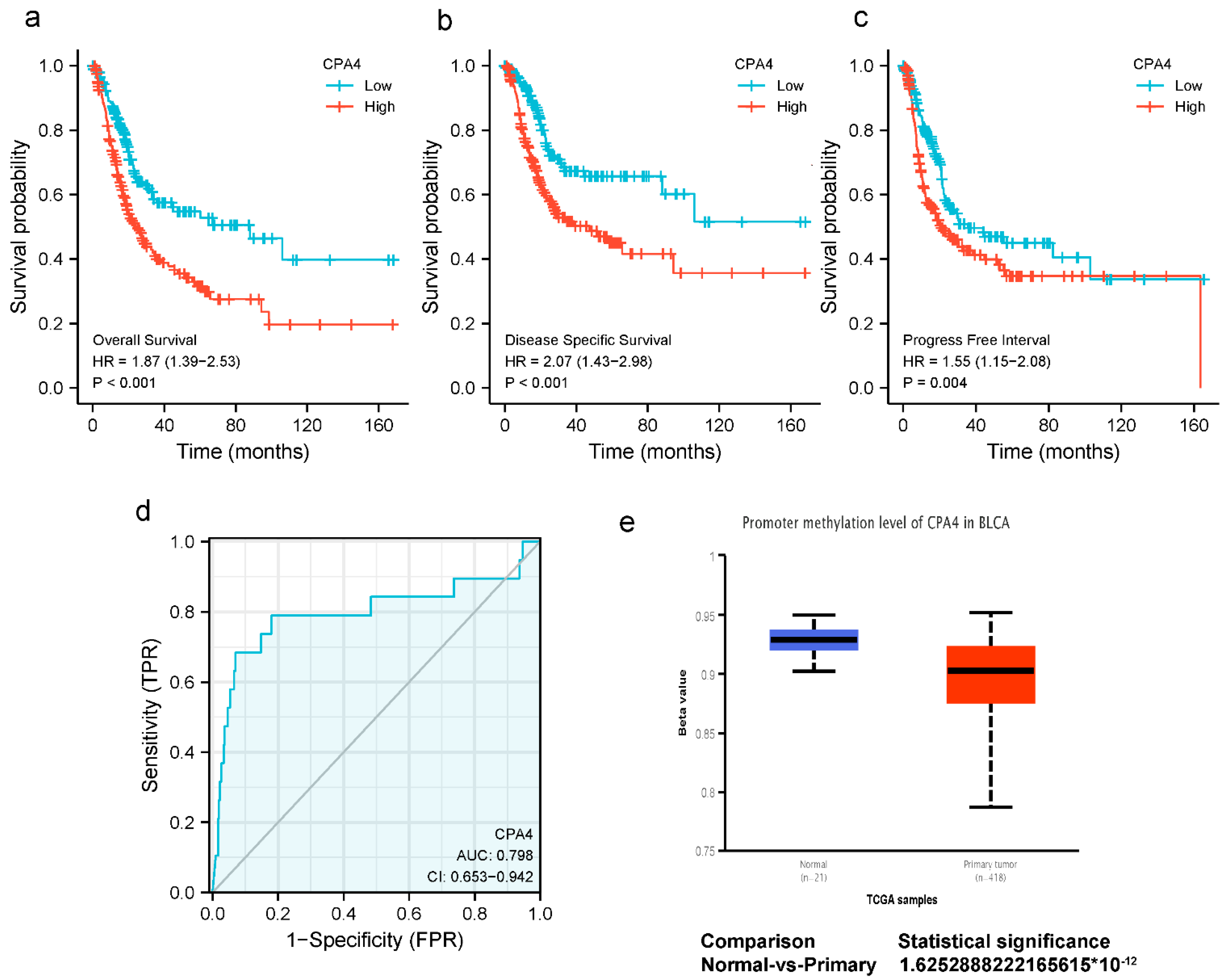
3.2. Tumor Tissues Express Higher CPA4 Than Normal Tissue
3.3. Impact of High CPA4 Expression on the Detection and Prognosis of BLCA Patients
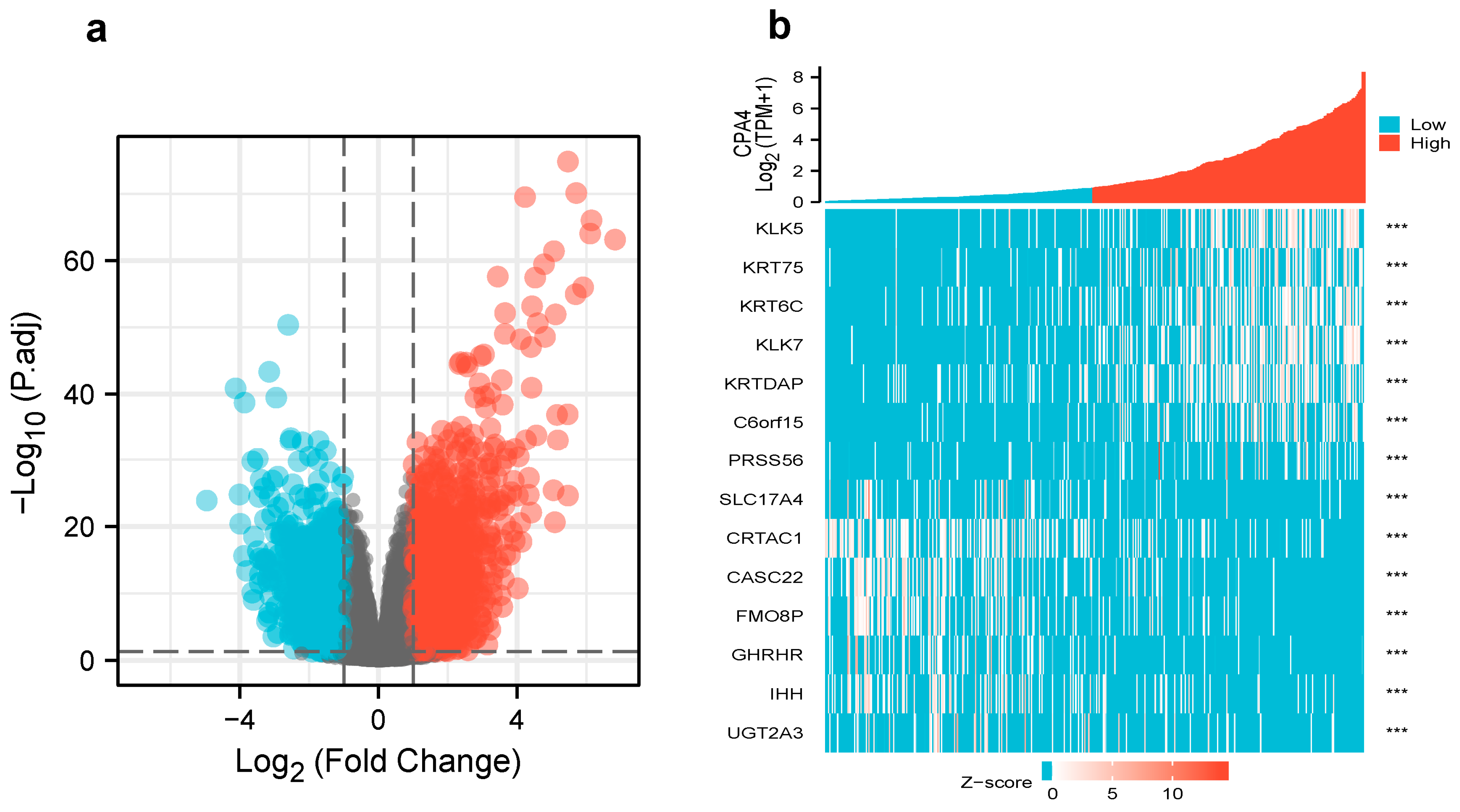
3.4. Differentially Expressed Genes and GO Enrichment Analysis in High- and Low-CPA4 Expression Samples
3.5. Gene Set Enrichment Analysis for CPA4-Related Signaling Pathways
3.6. CPA4 Expression Predicts Poor Prognosis in Different Cancer Stages
3.7. Construction of Nomogram for Predicting OS and Validation by Calibration
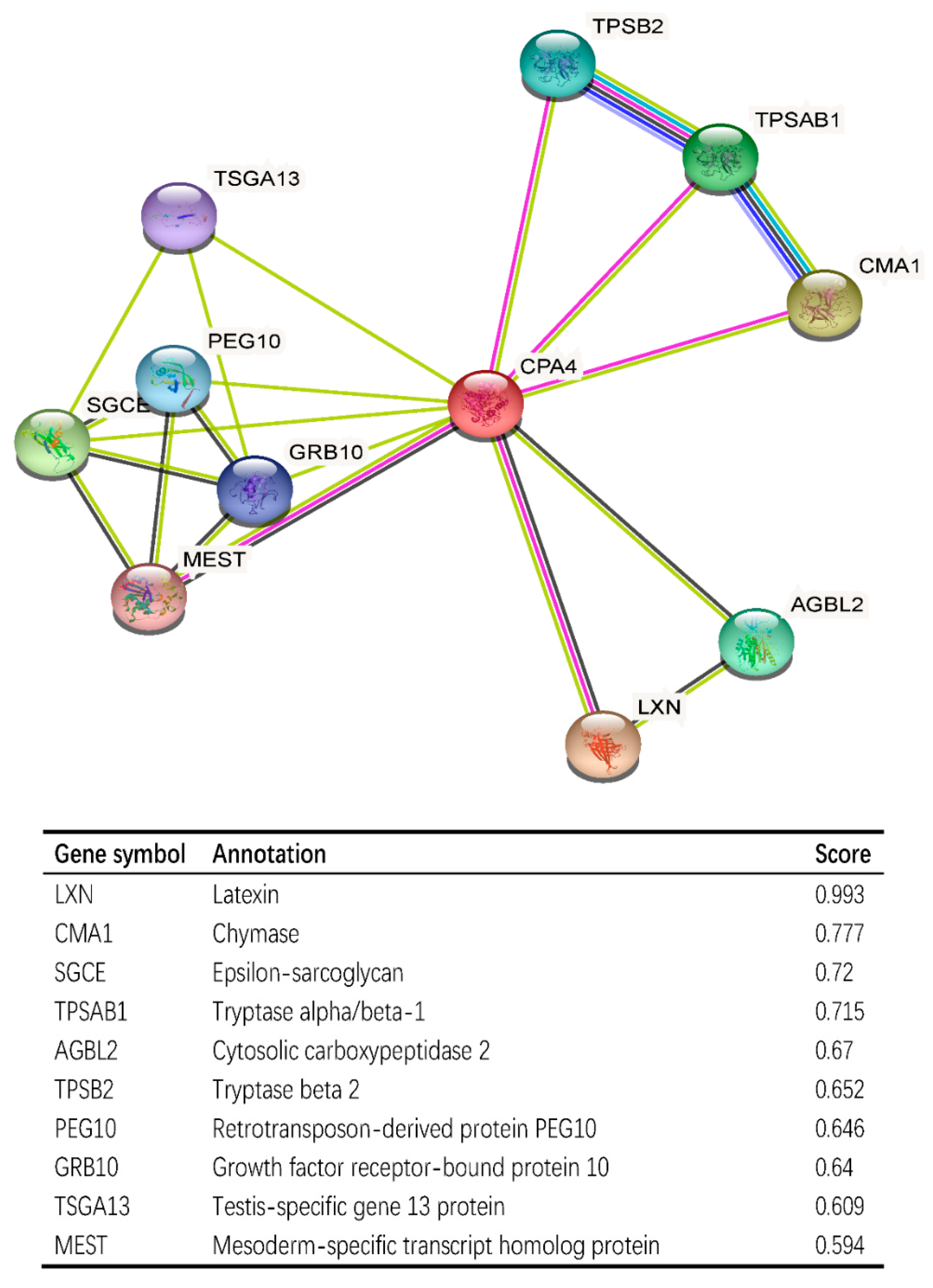
3.8. CPA4-Interaction Protein Networks in BLCA Tissue
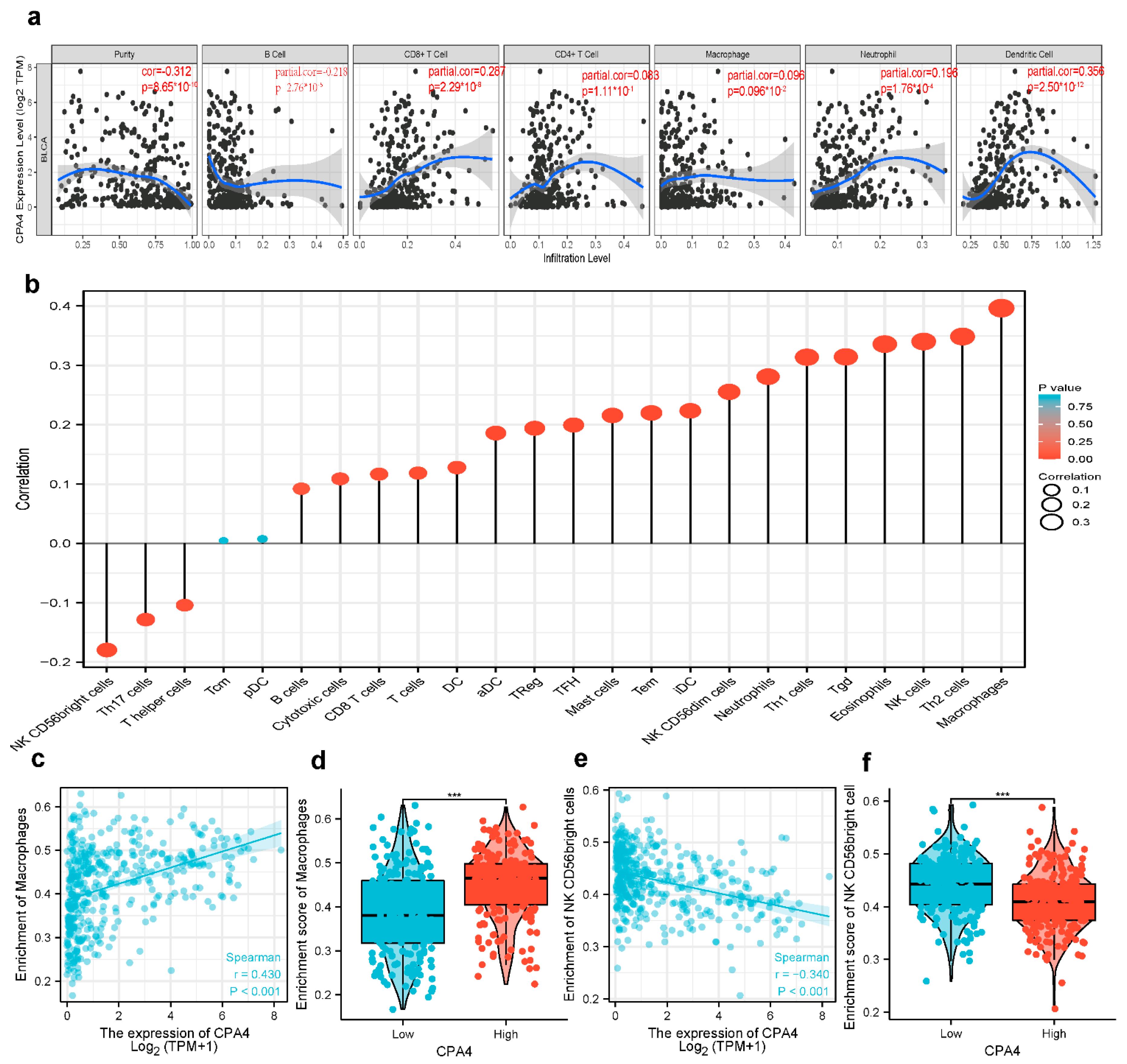
3.9. Correlation Analysis between CPA4 Expression and Infiltrating Immune Cells
3.10. Possible Role of the Expression of CPA4 in Various Infiltrating Immune Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenis, A.T.; Lec, P.M.; Chamie, K.; Mshs, M.D. Bladder Cancer: A Review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- McConkey, D.J.; Choi, W. Molecular Subtypes of Bladder Cancer. Curr. Oncol. Rep. 2018, 20, 77. [Google Scholar] [CrossRef]
- Bhanvadia, S.K. Bladder Cancer Survivorship. Curr. Urol. Rep. 2018, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- Babjuk, M.; Burger, M.; Compérat, E.M.; Gontero, P.; Mostafid, A.H.; Palou, J.; van Rhijn, B.W.G.; Rouprêt, M.; Shariat, S.F.; Sylvester, R.; et al. European Association of Urology Guidelines on Non-muscle-invasive Bladder Cancer (TaT1 and Carcinoma In Situ)—2019 Update. Eur. Urol. 2019, 76, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Berdik, C. Unlocking bladder cancer. Nature 2017, 551, S34–S35. [Google Scholar] [CrossRef]
- James, A.C.; Gore, J.L. The costs of non-muscle invasive bladder cancer. Urol. Clin. N. Am. 2013, 40, 261–269. [Google Scholar] [CrossRef]
- Abdollah, F.; Gandaglia, G.; Thuret, R.; Schmitges, J.; Tian, Z.; Jeldres, C.; Passoni, N.M.; Briganti, A.; Shariat, S.F.; Perrotte, P.; et al. Incidence, survival and mortality rates of stage-specific bladder cancer in United States: A trend analysis. Cancer Epidemiol. 2013, 37, 219–225. [Google Scholar] [CrossRef]
- Grayson, M. Bladder cancer. Nature 2017, 551, S33. [Google Scholar] [CrossRef] [Green Version]
- Tanco, S.; Zhang, X.; Morano, C.; Avilés, F.X.; Lorenzo, J.; Fricker, L.D. Characterization of the substrate specificity of human carboxypeptidase A4 and implications for a role in extracellular peptide processing. J. Biol. Chem. 2010, 285, 18385–18396. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.L.; Cheng, I.; Liu, X.; Cicek, M.S.; Carroll, P.R.; Casey, G.; Witte, J.S. Carboxypeptidase 4 gene variants and early-onset intermediate-to-high risk prostate cancer. BMC Cancer 2009, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Kayashima, T.; Yamasaki, K.; Yamada, T.; Sakai, H.; Miwa, N.; Ohta, T.; Yoshiura, K.; Matsumoto, N.; Nakane, Y.; Kanetake, H.; et al. The novel imprinted carboxypeptidase A4 gene (CPA4) in the 7q32 imprinting domain. Hum. Genet. 2003, 112, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Burnett, J.; Guo, C.; Xie, Y.; Pan, J.; Yang, Z.; Ran, Y.; Sun, D. CPA4 is a promising diagnostic serum biomarker for pancreatic cancer. Am. J. Cancer Res. 2016, 6, 91–96. [Google Scholar] [PubMed]
- Hong, W.; Xue, M.; Jiang, J.; Zhang, Y.; Gao, X. Circular RNA circ-CPA4/ let-7 miRNA/PD-L1 axis regulates cell growth, stemness, drug resistance and immune evasion in non-small cell lung cancer (NSCLC). J. Exp. Clin. Cancer Res. CR 2020, 39, 149. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Zhang, Z.; Cao, R.; Zang, H.; Pei, W.; Sun, T. CPA4 Promotes EMT in Pancreatic Cancer via Stimulating PI3K-AKT-mTOR Signaling. OncoTargets Ther. 2020, 13, 8567–8580. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Reed, C.P.; Zhang, J.S.; Shridhar, V.; Wang, L.; Smith, D.I. Carboxypeptidase A3 (CPA3): A novel gene highly induced by histone deacetylase inhibitors during differentiation of prostate epithelial cancer cells. Cancer Res. 1999, 59, 2981–2988. [Google Scholar]
- Sun, L.; Guo, C.; Burnett, J.; Pan, J.; Yang, Z.; Ran, Y.; Sun, D. Association between expression of Carboxypeptidase 4 and stem cell markers and their clinical significance in liver cancer development. J. Cancer 2017, 8, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Su, L.; Cai, M.; Yao, B.; Xiao, S.; He, Q.; Xu, L.; Yang, L.; Zhao, C.; Wan, T.; et al. Downregulation of CPA4 inhibits non small-cell lung cancer growth by suppressing the AKT/c-MYC pathway. Mol. Carcinog. 2019, 58, 2026–2039. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, K.; Czerwińska, P.; Wiznerowicz, M. The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge. Contemp. Oncol. (Pozn. Pol.) 2015, 19, A68–A77. [Google Scholar] [CrossRef]
- Blum, A.; Wang, P.; Zenklusen, J.C. SnapShot: TCGA-Analyzed Tumors. Cell 2018, 173, 530. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia (N. Y.) 2017, 19, 649–658. [Google Scholar] [CrossRef]
- He, J.; Chen, D.L.; Samocha-Bonet, D.; Gillinder, K.R.; Barclay, J.L.; Magor, G.W.; Perkins, A.C.; Greenfield, J.R.; Yang, G.; Whitehead, J.P. Fibroblast growth factor-1 (FGF-1) promotes adipogenesis by downregulation of carboxypeptidase A4 (CPA4)—A negative regulator of adipogenesis implicated in the modulation of local and systemic insulin sensitivity. Growth Factors (Chur Switz.) 2016, 34, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Dinu, I.; Potter, J.D.; Mueller, T.; Liu, Q.; Adewale, A.J.; Jhangri, G.S.; Einecke, G.; Famulski, K.S.; Halloran, P.; Yasui, Y. Improving gene set analysis of microarray data by SAM-GS. BMC Bioinform. 2007, 8, 242. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Liu, J.; Wang, J.; Zhong, X.; Dong, D.; Yang, X.; Wang, Y. Development and validation of a prognostic immune-associated gene signature in clear cell renal cell carcinoma. Int. Immunopharmacol. 2020, 81, 106274. [Google Scholar] [CrossRef]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Tang, Z.; Zhang, W.; Ye, Z.; Liu, F. GEPIA2021: Integrating multiple deconvolution-based analysis into GEPIA. Nucleic Acids Res. 2021, 49, W242–W246. [Google Scholar] [CrossRef]
- Wei, S.; Segura, S.; Vendrell, J.; Aviles, F.X.; Lanoue, E.; Day, R.; Feng, Y.; Fricker, L.D. Identification and characterization of three members of the human metallocarboxypeptidase gene family. J. Biol. Chem. 2002, 277, 14954–14964. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, Y.; Yuan, H.; Burnett, J.; Pan, J.; Yang, Z.; Ran, Y.; Myers, I.; Sun, D. CPA4 is a Novel Diagnostic and Prognostic Marker for Human Non-Small-Cell Lung Cancer. J. Cancer 2016, 7, 1197–1204. [Google Scholar] [CrossRef]
- Sun, L.; Guo, C.; Yuan, H.; Burnett, J.; Pan, J.; Yang, Z.; Ran, Y.; Myers, I.; Sun, D. Overexpression of carboxypeptidase A4 (CPA4) is associated with poor prognosis in patients with gastric cancer. Am. J. Transl. Res. 2016, 8, 5071–5075. [Google Scholar]
- Sun, L.; Guo, C.; Burnett, J.; Yang, Z.; Ran, Y.; Sun, D. Serum carboxypeptidaseA4 levels predict liver metastasis in colorectal carcinoma. Oncotarget 2016, 7, 78688–78697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, T.; Katayama, A.; Yokobori, T.; Yamane, A.; Fujii, T.; Obayashi, S.; Kurozumi, S.; Kawabata-Iwakawa, R.; Gombodorj, N.; Nishiyama, M.; et al. Carboxypeptidase A4 accumulation is associated with an aggressive phenotype and poor prognosis in triple-negative breast cancer. Int. J. Oncol. 2019, 54, 833–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Cao, J.; Guo, C.; Burnett, J.; Yang, Z.; Ran, Y.; Sun, D. Associations of carboxypeptidase 4 with ALDH1A1 expression and their prognostic value in esophageal squamous cell carcinoma. Dis. Esophagus Off. J. Int. Soc. Dis. Esophagus 2017, 30, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Abudurexiti, M.; Xie, H.; Jia, Z.; Zhu, Y.; Zhu, Y.; Shi, G.; Zhang, H.; Dai, B.; Wan, F.; Shen, Y.; et al. Development and External Validation of a Novel 12-Gene Signature for Prediction of Overall Survival in Muscle-Invasive Bladder Cancer. Front. Oncol. 2019, 9, 856. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Niu, Y.; Song, P.; Liu, Y.; Burnett, J.; Yang, Z.; Sun, D.; Ran, Y.; Li, Y.; et al. Carboxypeptidase A4 negatively correlates with p53 expression and regulates the stemness of breast cancer cells. Int. J. Med Sci. 2021, 18, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Qin, C.; Zhang, C.; Su, J.; Xiao, Q.; Xiao, Y.; Xiao, K.; Liu, Q. circCPA4 acts as a prognostic factor and regulates the proliferation and metastasis of glioma. J. Cell. Mol. Med. 2019, 23, 6658–6665. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Qu, J.; Cao, H.; Jing, C.; Wang, Z.; Xu, H.; Ma, R. Monoclonal antibody AC10364 inhibits cell proliferation in 5-fluorouracil resistant hepatocellular carcinoma via apoptotic pathways. OncoTargets Ther. 2019, 12, 5053–5067. [Google Scholar] [CrossRef] [Green Version]
- Pallarès, I.; Bonet, R.; García-Castellanos, R.; Ventura, S.; Avilés, F.X.; Vendrell, J.; Gomis-Rüth, F.X. Structure of human carboxypeptidase A4 with its endogenous protein inhibitor, latexin. Proc. Natl. Acad. Sci. USA 2005, 102, 3978–3983. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Ye, S.; Mao, J.; Ru, Y.; Lu, Y.; Wu, X.; Xu, M.; Zhu, T.; Wang, Y.; Chen, Y.; et al. CMA1 is potent prognostic marker and associates with immune infiltration in gastric cancer. Autoimmunity 2020, 53, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. Th1/Th2 cells. Inflamm. Bowel Dis. 1999, 5, 285–294. [Google Scholar] [CrossRef]
- Sasidharan Nair, V.; Toor, S.M.; Ali, B.R.; Elkord, E. Dual inhibition of STAT1 and STAT3 activation downregulates expression of PD-L1 in human breast cancer cells. Expert Opin. Ther. Targets 2018, 22, 547–557. [Google Scholar] [CrossRef]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Walker, L.S.; Sansom, D.M. The emerging role of CTLA4 as a cell-extrinsic regulator of T cell responses. Nat. Rev. Immunol. 2011, 11, 852–863. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Low Expression of CPA4 | High Expression of CPA4 | p |
|---|---|---|---|
| n | 207 | 207 | |
| Age, n (%) | 0.921 | ||
| ≤70 | 116 (28%) | 118 (28.5%) | |
| >70 | 91 (22%) | 89 (21.5%) | |
| Race, n (%) | 0.003 | ||
| Asian | 32 (8.1%) | 12 (3%) | |
| Black or African American | 8 (2%) | 15 (3.8%) | |
| White | 159 (40.1%) | 171 (43.1%) | |
| Gender, n (%) | 0.372 | ||
| Female | 50 (12.1%) | 59 (14.3%) | |
| Male | 157 (37.9%) | 148 (35.7%) | |
| T stage, n (%) | 0.004 | ||
| T1 | 4 (1.1%) | 1 (0.3%) | |
| T2 | 73 (19.2%) | 46 (12.1%) | |
| T3 | 89 (23.4%) | 107 (28.2%) | |
| T4 | 23 (6.1%) | 37 (9.7%) | |
| N stage, n (%) | 0.494 | ||
| N0 | 120 (32.4%) | 119 (32.2%) | |
| N1 | 18 (4.9%) | 28 (7.6%) | |
| N2 | 39 (10.5%) | 38 (10.3%) | |
| N3 | 3 (0.8%) | 5 (1.4%) | |
| M stage, n (%) | 0.810 | ||
| M0 | 109 (51.2%) | 93 (43.7%) | |
| M1 | 5 (2.3%) | 6 (2.8%) | |
| Pathologic stage, n (%) | 0.014 | ||
| Stage I | 4 (1%) | 0 (0%) | |
| Stage II | 76 (18.4%) | 54 (13.1%) | |
| Stage III | 63 (15.3%) | 79 (19.2%) | |
| Stage IV | 63 (15.3%) | 73 (17.7%) | |
| Radiation therapy, n (%) | 0.369 | ||
| No | 181 (46.6%) | 186 (47.9%) | |
| Yes | 13 (3.4%) | 8 (2.1%) | |
| Primary therapy outcome, n (%) | <0.001 | ||
| PD | 18 (5%) | 52 (14.6%) | |
| SD | 14 (3.9%) | 17 (4.8%) | |
| PR | 12 (3.4%) | 10 (2.8%) | |
| CR | 136 (38.1%) | 98 (27.5%) | |
| Histologic grade, n (%) | <0.001 | ||
| High Grade | 186 (45.3%) | 204 (49.6%) | |
| Low Grade | 19 (4.6%) | 2 (0.5%) | |
| Lymphovascular invasion, n (%) | 0.666 | ||
| No | 62 (21.9%) | 68 (24%) | |
| Yes | 78 (27.6%) | 75 (26.5%) | |
| Subtype, n (%) | 0.003 | ||
| Non-Papillary | 124 (30.3%) | 151 (36.9%) | |
| Papillary | 82 (20%) | 52 (12.7%) | |
| OS event, n (%) | <0.001 | ||
| Alive | 139 (33.6%) | 92 (22.2%) | |
| Dead | 68 (16.4%) | 115 (27.8%) | |
| Age, meidan (IQR) | 69 (60, 76) | 68 (61, 76) | 0.990 |
| Cell Type | Gene Marker | None Cor | p | Purity Cor | p | Tumor R | p | Normal R | p |
|---|---|---|---|---|---|---|---|---|---|
| B cell | CD19 | −0.042 | 0.397 | −0.138 | ** | −0.032 | 0.52 | −0.033 | 0.89 |
| CD20(KRT20) | −0.314 | *** | −0.226 | *** | −0.13 | * | −0.18 | 0.47 | |
| CD38 | 0.301 | *** | 0.148 | ** | 0.1 | * | −0.032 | 0.9 | |
| CD8+ T cell | CD8A | 0.267 | *** | 0.12 | * | −0.032 | 0.9 | −0.074 | 0.76 |
| CD8B | 0.15 | ** | 0.018 | 0.727 | 0.0031 | 0.95 | −0.096 | 0.69 | |
| Tfh | BCL6 | −0.247 | *** | −0.214 | *** | −0.14 | ** | −0.28 | 0.25 |
| ICOS | −0.307 | *** | 0.154 | ** | 0.13 | ** | −0.095 | 0.7 | |
| CXCR5 | 0.109 | * | −0.095 | 0.0677 | 0.075 | 0.13 | 0.039 | 0.87 | |
| Th1 | T-bet(TBX21) | 0.227 | *** | 0.046 | 0.375 | 0.2 | *** | 0.041 | 0.87 |
| STAT4 | 0.37 | *** | 0.223 | *** | 0.16 | ** | 0.0068 | 0.98 | |
| IL12RB2 | 0.403 | *** | 0.327 | *** | 0.24 | *** | −0.22 | 0.37 | |
| WSX1(IL27RA) | 0.39 | *** | 0.291 | *** | 0.18 | *** | 0.027 | 0.91 | |
| STAT1 | 0.386 | *** | 0.282 | *** | 0.24 | *** | −0.14 | 0.56 | |
| IFN-γ(IFNG) | 0.278 | *** | 0.161 | ** | 0.13 | ** | −0.085 | 0.73 | |
| TNF-α(TNF) | 0.287 | *** | 0.194 | *** | 0.098 | * | 0.27 | 0.26 | |
| Th2 | GATA3 | −0.484 | *** | −0.402 | *** | −0.26 | *** | −0.26 | 0.28 |
| CCR3 | 0.188 | *** | 0.131 | * | 0.67 | * | −0.14 | 0.58 | |
| STAT6 | −0.228 | *** | −0.209 | *** | −0.13 | ** | −0.32 | 0.18 | |
| STAT5A | 0.004 | 0.936 | −0.158 | ** | −0.015 | 0.76 | −0.53 | * | |
| Th9 | TGFBR2 | 0.087 | 0.079 | −0.014 | 0.792 | 0.038 | 0.45 | −0.45 | 0.056 |
| IRF4 | 0.188 | *** | −0.03 | 0.571 | 0.043 | 0.39 | −0.12 | 0.63 | |
| PU.1(SPI1) | 0.356 | *** | 0.181 | *** | 0.15 | ** | −0.17 | 0.49 | |
| Th17 | STAT3 | 0.325 | *** | 0.232 | *** | 0.15 | ** | −0.11 | 0.64 |
| IL-21R | 0.318 | *** | 0.132 | * | 0.073 | 0.14 | −0.1 | 0.68 | |
| IL-23R | −0.003 | 0.945 | −0.076 | 0.143 | −0.0048 | 0.92 | −0.019 | 0.94 | |
| IL-17A | −0.019 | 0.705 | −0.057 | 0.274 | −0.051 | 0.31 | −0.18 | 0.47 | |
| Th22 | CCR10 | −0.025 | 0.626 | −0.068 | 0.195 | 0.029 | 0.57 | −0.34 | 0.16 |
| AHR | −0.271 | *** | −0.195 | *** | −0.11 | * | −0.29 | 0.23 | |
| Treg | FOXP3 | 0.287 | *** | 0.15 | ** | 0.16 | ** | 0.037 | 0.88 |
| CD25(IL2RA) | 0.369 | *** | 0.22 | *** | 0.037 | 0.88 | −0.066 | 0.79 | |
| CCR8 | 0.218 | *** | 0.083 | 0.113 | 0.083 | 0.094 | −0.0059 | 0.98 | |
| T cell exhaustion | PD-1(PDCD1) | 0.255 | *** | 0.089 | * | 0.089 | 0.073 | −0.099 | 0.69 |
| CTLA4 | 0.311 | *** | 0.16 | ** | 0.23 | *** | −0.11 | 0.64 | |
| LAG3 | 0.362 | *** | 0.227 | *** | 0.22 | *** | −0.19 | 0.45 | |
| TIM-3(HAVCR2) | 0.375 | *** | 0.218 | *** | 0.21 | *** | −0.097 | 0.69 | |
| Macrophage | CD68 | 0.316 | *** | 0.193 | *** | 0.14 | ** | 0.49 | * |
| CD11b(ITGAM) | 0.303 | *** | 0.119 | * | 0.72 | * | −0.29 | 0.23 | |
| M1 | INOS(NOS2) | −0.033 | 0.511 | −0.092 | 0.0774 | −0.0068 | 0.89 | −0.14 | 0.57 |
| IRF5 | −0.123 | * | −0.116 | * | −0.063 | 0.2 | −0.026 | 0.92 | |
| COX2(PTGS2) | 0.209 | *** | 0.164 | ** | 0.057 | 0.25 | −0.24 | 0.32 | |
| M2 | CD16 | 0.408 | *** | 0.273 | *** | 0.17 | *** | −0.2 | 0.42 |
| ARG1 | −0.049 | 0.322 | −0.007 | 0.894 | 0.076 | 0.13 | 0.68 | ** | |
| MRC1 | 0.334 | *** | 0.164 | ** | 0.042 | 0.4 | −0.23 | 0.34 | |
| MS4A4A | 0.353 | *** | 0.199 | *** | 0.12 | ** | −0.23 | 0.34 | |
| TAM | CCL2 | 0.26 | *** | 0.113 | * | 0.022 | 0.66 | −0.12 | 0.62 |
| CD80 | 0.413 | *** | 0.285 | *** | 0.17 | *** | −0.18 | 0.46 | |
| CD86 | 0.396 | *** | 0.244 | *** | 0.17 | *** | −0.074 | 0.76 | |
| CCR5 | 0.29 | *** | 0.101 | 0.0522 | 0.13 | * | −0.08 | 0.75 | |
| Monocyte | CD14 | 0.406 | *** | 0.253 | *** | 0.11 | * | −0.21 | 0.38 |
| CD16(FCGR3B) | 0.316 | *** | 0.22 | *** | 0.15 | ** | −0.073 | 0.77 | |
| CD115(CSF1R) | 0.353 | *** | 0.178 | *** | 0.14 | ** | −0.28 | 0.24 | |
| Neutrophil | CD66b(CEACAM8) | 0.089 | 0.0721 | 0.098 | 0.0609 | −0.031 | 0.53 | −0.084 | 0.73 |
| CD15(FUT4) | 0.141 | ** | 0.047 | 0.369 | 0.0041 | 0.93 | −0.33 | 0.17 | |
| CD11b(ITGAM) | 0.303 | *** | 0.119 | * | 0.018 | 0.72 | −0.29 | 0.23 | |
| Natural killer cell | XCL1 | −0.01 | 0.844 | −0.005 | 0.93 | −0.06 | 0.23 | 0.13 | 0.59 |
| CD7 | 0.304 | *** | 0.131 | * | 0.15 | ** | −0.029 | 0.91 | |
| KIR3DL1 | 0.136 | ** | 0.049 | 0.346 | 0.075 | 0.13 | 0.19 | 0.44 | |
| Dendritic cell | CD1C(BDCA-1) | 0.086 | 0.0823 | −0.054 | 0.305 | −0.023 | 0.65 | −0.02 | 0.93 |
| CD141(THBD) | 0.356 | *** | 0.322 | *** | 0.055 | 0.27 | 0.37 | 0.12 | |
| CD11c(ITGAX) | 0.35 | *** | 0.181 | *** | 0.099 | * | −0.2 | 0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, C.; Zhou, Y.; Xiong, Q.; Xiong, M.; Hou, Y.; Yang, X.; Chen, Z. Comprehensive Analysis of CPA4 as a Poor Prognostic Biomarker Correlated with Immune Cells Infiltration in Bladder Cancer. Biology 2021, 10, 1143. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111143
Wei C, Zhou Y, Xiong Q, Xiong M, Hou Y, Yang X, Chen Z. Comprehensive Analysis of CPA4 as a Poor Prognostic Biomarker Correlated with Immune Cells Infiltration in Bladder Cancer. Biology. 2021; 10(11):1143. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111143
Chicago/Turabian StyleWei, Chengcheng, Yuancheng Zhou, Qi Xiong, Ming Xiong, Yaxin Hou, Xiong Yang, and Zhaohui Chen. 2021. "Comprehensive Analysis of CPA4 as a Poor Prognostic Biomarker Correlated with Immune Cells Infiltration in Bladder Cancer" Biology 10, no. 11: 1143. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111143