
Shifting Perspectives of Translational Research in Bio-Bactericides: Reviewing the Bacillus amyloliquefaciens Paradigm

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. An Overview of Bacterial Pesticides
2.1. The Global Market
2.2. The Discovery of Bacterial Biopesticides
2.3. The Traditional Approach to Develop Bacterial Pesticides
3. Shifting Perspectives in BCA Mode of Action and Application
3.1. “Let Them Eat Cake”: Antibiotic Production Is Conditional
3.2. The Role of Siderophores in Biocontrol
3.3. Reciprocal Perception Enables Host-Microbe Interactions
3.4. BCAs Trigger Multifaceted Defense Responses
3.5. The Environmental Impact of Bio-Microbicides
3.6. The Multidisciplinary Approach to Study BCAs
4. Future Challenges
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R. Bacterial diseases of crops: Elucidation of the factors that lead to differences between field and experimental infections. Adv. Agron. 2015, 134, 227–246. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J. Fungal trunk pathogens in the grapevine propagation process: Potential inoculum sources, detection, identification, and management strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicard, A.; Zeilinger, A.R.; Vanhove, M.; Schartel, T.E.; Beal, D.J.; Daugherty, M.P.; Almeida, R.P.P. Xylella fastidiosa: Insights into an emerging plant pathogen. Annu. Rev. Phytopathol. 2018, 56, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, J.L. The Scientific, Economic, and Social Impacts of the New Zealand Outbreak of Bacterial Canker of Kiwifruit (Pseudomonas syringae pv. actinidiae). Annu. Rev. Phytopathol. 2017, 55, 377–399. [Google Scholar] [CrossRef]
- Kramer, J.; Simnitt, S.; Calvin, L. Fruit and Tree Nuts Outlook: September 2020; United States Department of Agriculture: Washington, DC, USA, 2020; pp. 1–45.
- Food, E.; Authority, S. Review of the existing maximum residue levels for copper compounds according to Article 12 of Regulation (EC) No 396/2005. EFSA J. 2018, 16, e05212. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; Court Marques, D.; et al. Peer review of the pesticide risk assessment of the active substance copper compounds copper(I), copper(II) variants namely copper hydroxide, copper oxychloride, tribasic copper sulfate, copper(I) oxide, Bordeaux mixture. EFSA J. 2018, 16, 1–25. [Google Scholar] [CrossRef]
- World Health Organization. Copper in Drinking-Water; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Pal, K.K.; McSpadden Gardener, B. Biological Control of Plant Pathogens. Plant Health Instr. 2006, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Cawoy, H.; Bettiol, W.; Fickers, P.; Ongena, M. Bacillusbased biological control of plant diseases. In Pesticides in the Modern Worldpesticides Use and Management; Stoytcheva, M., Ed.; InTech: Rijeka, Croatia, 2011; pp. 273–302. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Veerman, C.; Pinto Correia, T.; Bastioli, C.; Biro, B.; Bouma, J.; Cienciala, E.; Emmett, B.; Frison, E.; Grand, A.; Hristov Filchew, L.; et al. Caring for Soil is Caring for Life—Ensure 75% of Soils are Healthy by 2030 for food, People, Nature and Climate; Report of the Mission Board of Soil Health and Food; European Commission: Brussels, Belgium, 2020; p. 82. [Google Scholar]
- Van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Fravel, D.R. Commercialization and Implementation of Biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Velivelli, S.L.S.; De Vos, P.; Kromann, P.; Declerck, S.; Prestwich, B.D. Biological control agents: From field to market, problems, and challenges. Trends Biotechnol. 2014, 32, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Marrone, P.G. Pesticidal natural products—Status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef]
- Lambert, B.; Peferoen, M. Promise of Insecticidal Bacillus thuringiensis Facts and mysteries about a successful biopesticide. Bioscience 1992, 42, 112–122. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Stack, J.P. Using Pseudomonas spp. for integrated biological control. In Proceedings of the The Nature and Application of Biocontrol Microbes III: Pseudomonas spp. Phytopathology 2007, 97, 244–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regulation (EC) No. 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Off. J. Eur. Union 2009, 309, 1–50.
- Frederiks, C.; Wesseler, J.H.H. A comparison of the EU and US regulatory frameworks for the active substance registration of microbial biological control agents. Pest Manag. Sci. 2019, 75, 87–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Leahy, J.; Mendelsohn, M.; Kough, J.; Jones, R.; Berckes, N. Biopesticide oversight and registration at the U.S. Environmental Protection Agency. ACS Symp. Ser. 2014, 1172, 3–18. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological control of plant diseases: The European situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Environmental Protection Agency. Bacillus subtilis MBI 600 (129082) Fact Sheet; Environmental Protection Agency: Washington, DC, USA, 1999; pp. 2–3.
- Lilly, J. US EPA, Pesticide Product Label, Serifel, 02/11/2019; Environmental Protection Agency: Washington, DC, USA, 2019; Volume 130, ISBN 1800424930.
- Montesinos, E. Development, registration and commercialization of microbial pesticides for plant protection. Int. Microbiol. 2003, 6, 245–252. [Google Scholar] [CrossRef]
- Gardener, B.B.M.; Fravel, D.R. Biological Control of Plant Pathogens: Research, Commercialization, and Application in the USA. Plant Health Prog. 2002, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Borriss, R. Use of Plant-Associated Bacillus Strains as Biofertilizers and Biocontrol Agents in Agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. ISBN 978-3-642-20331-2. [Google Scholar]
- Hartley, C. Damping-Off in Forest Nurseries; U.S. Department of Agriculture: Washington, DC, USA, 1921; Volume 934, pp. 1–99.
- Millard, W.A.; Taylor, C.B. Antagonism of Micro-Organisms As the Controlling Factor in the: Inhibition of Scab By Green-Manuring. Ann. Appl. Biol. 1927, 14, 202–216. [Google Scholar] [CrossRef]
- Weindling, R.; Fawcett, H.S. Experiments in the control of Rhizoctonia damping-off of citrus seedlings. Hilgardia 1936, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Escobar, M.A.; Dandekar, A.M. Agrobacterium tumefaciens as an agent of disease. Trends Plant Sci. 2003, 8, 380–386. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Control of Fire Blight by Pseudomonas fluorescens A506 and Pantoea vagans C9-1 Applied as Single Strains and Mixed Inocula. Biol. Control 2010, 1, 44–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, R.J.; Rovira, A.D. The role of bacteria in the biological control of Gaeumannomyces graminis by suppressive soils. Soil Biol. Biochem. 1976, 8, 269–273. [Google Scholar] [CrossRef]
- Kandel, S.; Joubert, P.; Doty, S. Bacterial Endophyte Colonization and Distribution within Plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Doornbos, R.F.; Van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Erlacher, A.; Cardinale, M.; Grosch, R.; Grube, M.; Berg, G. The impact of the pathogen Rhizoctonia solani and its beneficial counterpart Bacillus amyloliquefaciens on the indigenous lettuce microbiome. Front. Microbiol. 2014, 5, 175. [Google Scholar] [CrossRef] [Green Version]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niazi, A.; Manzoor, S.; Asari, S.; Bejai, S.; Meijer, J.; Bongcam-Rudloff, E. Genome Analysis of Bacillus amyloliquefaciens Subsp. plantarum UCMB5113: A Rhizobacterium That Improves Plant Growth and Stress Management. PLoS ONE 2014, 9, e104651. [Google Scholar] [CrossRef]
- De Jong, I.G.; Beilharz, K.; Kuipers, O.P.; Veening, J.-W. Live Cell Imaging of Bacillus subtilis and Streptococcus pneumoniae using Automated Time-lapse Microscopy. J. Vis. Exp. 2011, e3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelopoulou, D.J.; Naska, E.J.; Paplomatas, E.J.; Tjamos, S.E. Biological control agents (BCAs) of verticillium wilt: Influence of application rates and delivery method on plant protection, triggering of host defence mechanisms and rhizosphere populations of BCAs. Plant Pathol. 2014, 63, 1062–1069. [Google Scholar] [CrossRef]
- Bonaterra, A.; Cabrefiga, J.; Camps, J.; Montesinos, E. Increasing survival and efficacy of a bacterial biocontrol agent of fire blight of rosaceous plants by means of osmoadaptation. FEMS Microbiol. Ecol. 2007, 61, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Lindow, S.E.; Suslow, T.V. Temporal dynamics of the biocontrol agent Pseudomonas fluorescens strain A506 in flowers in inoculated pear trees. Phytopathology 2003, 93, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auld, B.A.; Morin, L. Constraints in the Development of Bioherbicides. Weed Technol. 1995, 9, 638–652. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Saha, D.; Purkayastha, G.D.; Ghosh, A.; Isha, M.; Saha, A. Isolation and Characterization of Two New Bacillus Subtilis Strains From the Rhizosphere of Eggplant As Potential Biocontrol Agents. J. Plant Pathol. 2012, 94, 109–118. [Google Scholar]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturz, A. Bacterial root zone communities, beneficial allelopathies and plant disease control. In Allelochemical Biological Control of Plant Pathogens and Diseases; Springer: Dordrecht, The Netherlands, 2006; pp. 123–142. ISBN 9781402044458. [Google Scholar]
- De La Fuente, L.; Quagliotto, L.; Bajsa, N.; Fabiano, E.; Altier, N.; Arias, A. Inoculation with Pseudomonas fiuorescens biocontrol strains does not affect the symbiosis between rhizobia and forage legumes. Soil Biol. Biochem. 2002, 34, 545–548. [Google Scholar] [CrossRef]
- Velusamy, P.; Ebenezar Immanuel, J.; Gnanamanickam, S.S.; Thomashow, L. Biological control of rice bacterial blight by plant-associated bacteria producing 2,4-diacetylphloroglucinol. Can. J. Microbiol. 2006, 52, 56–65. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Delves-Broughton, J. Nisin and its application as a food preservative. J. Soc. Dairy Technol. 1990, 43, 73–76. [Google Scholar] [CrossRef]
- Parisot, J.; Carey, S.; Breukink, E.; Chan, W.C.; Narbad, A.; Bonev, B. Molecular mechanism of target recognition by subtilin, a class I lanthionine antibiotic. Antimicrob. Agents Chemother. 2008, 52, 612–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammami, I.; Rhouma, A.; Jaouadi, B.; Rebai, A.; Nesme, X. Optimization and biochemical characterization of a bacteriocin from a newly isolated Bacillus subtilis strain 14B for biocontrol of Agrobacterium spp. strains. Lett. Appl. Microbiol. 2009, 48, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; Mögel, G.; Kunz, S.; Borriss, R. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wu, H.; Chen, L.; Yu, X.; Borriss, R.; Gao, X. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens. Sci. Rep. 2015, 5, 12975. [Google Scholar] [CrossRef]
- Zeriouh, H.; Romero, D.; Garcia-Gutierrez, L.; Cazorla, F.M.; de Vicente, A.; Perez-Garcia, A. The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol. Plant. Microbe. Interact. 2011, 24, 1540–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Wang, J.P.; Zhu, Y.J.; Liu, B.; Yang, W.J.; Ruan, C.Q. Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349. J. Appl. Microbiol. 2019, 126, 1519–1529. [Google Scholar] [CrossRef]
- Haas, D.; Keel, C. Regulation of Antibiotic Production in Root-Colonizing Pseudomonas spp. and Relevance for Biological Control of Plant Disease. Annu. Rev. Phytopathol. 2003, 41, 117–153. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Chavez, A.; Forero, A.; Garcia-Huante, Y.; Romero, A.; Sanchez, M.; Rocha, D.; Sanchez, B.; Avalos, M.; Guzman-Trampe, S.; et al. Carbon source regulation of antibiotic production. J. Antibiot. 2010, 63, 442–459. [Google Scholar] [CrossRef] [Green Version]
- Jacques, P. Biosurfactants: From Genes to Applications; Soberon-Chavez, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 57–91. ISBN 9783642144899. [Google Scholar]
- Duffy, B.K.; Défago, G. Environmental factors modulating antibiotic and siderophore biosynthesis by Pseudomonas fluorescens biocontrol strains. Appl. Environ. Microbiol. 1999, 65, 2429–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, J.L.; Silo-Suh, L.; Lee, J.C.; Haiyin, H.E.; Clardy, J.; Handelsman, J.O. Production of kanosamine by Bacillus cereus UW85. Appl. Environ. Microbiol. 1996, 62, 3061–3065. [Google Scholar] [CrossRef] [Green Version]
- Lugtenberg, B.; Rozen, D.E.; Kamilova, F. Wars between microbes on roots and fruits. F1000Research 2017, 6, 343. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Silby, M.W.; Raaijmakers, J.M.; Levy, S.B.; Boer, W. De Transcriptional and antagonistic responses of Pseudomonas fluorescens Pf0-1 to phylogenetically different bacterial competitors. ISME J. 2011, 5, 973–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traxler, M.F.; Watrous, J.D.; Alexandrov, T.; Dorrestein, P.C.; Kolter, R. Interspecies Interactions Stimulate Diversification of the Streptomyces coelicolor Secreted Metabolome. MBio 2013, 4, e00459-13. [Google Scholar] [CrossRef] [Green Version]
- Tyc, O.; van den Berg, M.; Gerards, S.; van Veen, J.A.; Raaijmakers, J.M.; de Boer, W.; Garbeva, P. Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 2014, 5, 567. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Diversity and Natural Functions of Antibiotics Produced by Beneficial and Plant Pathogenic Bacteria. Annu. Rev. Phytopathol. 2012, 50, 403–424. [Google Scholar] [CrossRef]
- Koch, E.; Becker, J.; Berg, G.; Hauschild, R.; Jehle, J.; Kohl, J.; Smalla, K. Biocontrol of plant diseases is not an unsafe technology! J. Plant Dis. Prot. 2018, 125, 121–125. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus amyloliquefaciens MBI600. mSphere 2021, 6, e00376-21. [Google Scholar] [CrossRef] [PubMed]
- Quadri, L.E.N. MicroReview Assembly of aryl-capped siderophores by modular peptide synthetases and polyketide synthases. Mol. Microbiol. 2000, 37, 1–12. [Google Scholar] [CrossRef]
- Su, C.; Zhang, M.; Lin, L.; Yu, G.; Zhong, H.; Chong, Y. Reduction of iron oxides and microbial community composition in iron-rich soils with different organic carbon as electron donors. Int. Biodeterior. Biodegradation 2020, 148, 104881. [Google Scholar] [CrossRef]
- Winkelmann, G. Ecology of siderophores with special reference to the fungi. BioMetals 2007, 20, 379–392. [Google Scholar] [CrossRef]
- Sullivan, T.S.; Ramkissoon, S.; Garrison, V.H.; Ramsubhag, A.; Thies, J.E. Siderophore production of African dust microorganisms over Trinidad and Tobago. Aerobiologia 2012, 28, 391–401. [Google Scholar] [CrossRef]
- Masalha, J.; Kosegarten, H.; Elmaci, Ö. The central role of microbial activity for iron acquisition in maize and sunflower. Biol Fertil Soils 2000, 433–439. [Google Scholar] [CrossRef]
- Greenshields, D.L.; Liu, G.; Feng, J.; Selvaraj, G.; Wei, Y. The siderophore biosynthetic gene SID1, but not the ferroxidase gene FET3, is required for full Fusarium graminearum virulence. Mol. Plant Pathol. 2007, 8, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Heymann, P.; Ernst, J.F.; Winkelmann, G. A gene of the major facilitator superfamily encodes a transporter for enterobactin (Enb1p) in Saccharomyces cerevisiae. BioMetals 2000, 13, 65–72. [Google Scholar] [CrossRef]
- Arias, A.A.; Lambert, S.; Martinet, L.; Adam, D.; Tenconi, E.; Hayette, M.P.; Ongena, M.; Rigali, S. Growth of desferrioxamine-deficient Streptomyces mutants through xenosiderophore piracy of airborne fungal contaminations. FEMS Microbiol. Ecol. 2015, 91, fiv080. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, Y.; Wang, H.; Wang, J.; Cheol, M.; Byun, Y.; He, H.; Liang, Y.; Yang, V.C. Curb challenges of the “Trojan Horse” approach: Smart strategies in achieving effective yet safe cell-penetrating peptide-based drug delivery. Adv. Drug Deliv. Rev. 2013, 65, 1299–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.; Sun, L.; Qi, G.; Chen, S.; Zhao, X. Microbial community composition is related to soil biological and chemical properties and bacterial wilt outbreak. Sci. Rep. 2017, 7, 343. [Google Scholar] [CrossRef] [Green Version]
- Badri, D.V.; Weir, T.L.; van der Lelie, D.; Vivanco, J.M. Rhizosphere chemical dialogues: Plant–microbe interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Zhang, N.; Du, W.; Zhang, H.; Liu, Y.; Fu, R.; Shao, J.; Zhang, G.; Shen, Q.; Zhang, R. Identification of chemotaxis compounds in root exudates and their sensing chemoreceptors in plant-growth-promoting rhizobacteria bacillus amyloliquefaciens SQR9. Mol. Plant-Microbe Interact. 2018, 31, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- el Zahar Haichar, F.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C. Plant responses to plant growth-promoting rhizobacteria. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Springer: Dordrecht, The Netherlands, 2007; pp. 243–254. [Google Scholar]
- Fan, B.; Borriss, R.; Bleiss, W.; Wu, X. Gram-positive rhizobacterium Bacillus amyloliquefaciens FZB42 colonizes three types of plants in different patterns. J. Microbiol. 2012, 50, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietel, K.; Beator, B.; Budiharjo, A.; Fan, B.; Borriss, R. Bacterial Traits Involved in colonization of Arabidopsis thaliana roots by Bacillus amyloliquefaciens FZB42. Plant Pathol. J. 2013, 29, 59. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Feng, H.; Chen, L.; Zhang, H.; Dong, X.; Xiong, Q.; Zhang, R. Root-Secreted Spermine Binds to Bacillus amyloliquefaciens SQR9 Histidine Kinase KinD and Modulates Biofilm Formation. Mol. Plant-Microbe Interact. 2020, 33, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Calvio, C.; Celandroni, F.; Ghelardi, E.; Amati, G.; Salvetti, S.; Ceciliani, F.; Galizzi, A.; Senesi, S. Swarming differentiation and swimming motility in Bacillus subtilis are controlled by swrA, a newly identified dicistronic operon. J. Bacteriol. 2005, 187, 5356–5366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [Green Version]
- Thapa, S.; Prasanna, R.; Ranjan, K.; Velmourougane, K.; Ramakrishnan, B. Nutrients and host attributes modulate the abundance and functional traits of phyllosphere microbiome in rice. Microbiol. Res. 2017, 204, 55–64. [Google Scholar] [CrossRef]
- Skandalis, N.; Sarris, P.F.; Panopoulos, N.J. Targeting injectisomes of virulence: Benefits of thirty years of research on bacterial secretion systems, to crop protection. Hell. Plant Prot. J. 2012, 5, 31–47. Available online: https://www.bpi.gr/files/journal/2012/july/Hellenic%20Plant%20Protection%20Journal%20-%20Skandalis%20(July%202012).indd.pdf (accessed on 4 July 2021).
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Beckers, G.J.M.; Conrath, U. Priming for stress resistance: From the lab to the field. Curr. Opin. Plant Biol. 2007, 10, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Spoel, S.H. NPR1 Modulates Cross-Talk between Salicylate- and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol. Plant Cell Online 2003, 15, 760–770. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alqu, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic Lipopeptides of Bacillus amyloliquefaciens subsp. plantarum Colonizing the Lettuce Rhizosphere Enhance Plant Defense Responses Toward the Bottom Rot Pathogen Rhizoctonia solani. Mol. Plant-Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulou, A.; Theologidis, I.; Liebmann, B.; Kalantidis, K.; Vassilakos, N.; Skandalis, N. Bacillus amyloliquefaciens MBI600 differentially induces tomato defense signaling pathways depending on plant part and dose of application. Sci. Rep. 2019, 9, 19120. [Google Scholar] [CrossRef] [Green Version]
- Beris, D.; Theologidis, I.; Skandalis Nicholasand Vassilakos, N. Bacillus amyloliquefaciens strain MBI600 induces salicylic aciddependent resistance in tomato plants against Tomato spotted wiltvirus and Potato virus Y. Sci. Rep. 2018, 8, 10320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.-M.; Murphy, J.F.; Mysore, K.S.; Kloepper, J.W. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J. 2004, 39, 381–392. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.; Zhang, S. Induced Systemic Resistance and Promotion of Plant Growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fousia, S.; Paplomatas, E.J.; Tjamos, S.E. Bacillus subtilis QST 713 Confers Protection to Tomato Plants Against Pseudomonas syringae pv tomato and Induces Plant Defence-related Genes. J. Phytopathol. 2016, 164, 264–270. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Liu, Y.; Xu, Y.; Zhang, G.; Shen, Q.; Zhang, R. Exploring elicitors of the beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 to induce plant systemic resistance and their interactions with plant signaling pathways. Mol. Plant-Microbe Interact. 2018, 31, 560–567. [Google Scholar] [CrossRef] [Green Version]
- Beris, D.; Vassilakos, N. Plant beneficial microbes: Do they have a role as antiviral agents in agriculture? Mol. Asp. Plant Benef. Microbes Agric. 2020, 19–33. [Google Scholar] [CrossRef]
- Brown, L.; Kessler, A.; Cabezas-Sanchez, P.; Luque-Garcia, J.L.; Casadevall, A. Extracellular vesicles produced by the Gram-positive bacterium Bacillus subtilis are disrupted by the lipopeptide surfactin. Mol. Microbiol. 2014, 93, 183–198. [Google Scholar] [CrossRef] [Green Version]
- Pangesti, N.; Reichelt, M.; van de Mortel, J.E.; Kapsomenou, E.; Gershenzon, J.; van Loon, J.J.A.; Dicke, M.; Pineda, A. Jasmonic Acid and Ethylene Signaling Pathways Regulate Glucosinolate Levels in Plants During Rhizobacteria-Induced Systemic Resistance Against a Leaf-Chewing Herbivore. J. Chem. Ecol. 2016, 42, 1212–1225. [Google Scholar] [CrossRef] [Green Version]
- Zebelo, S.; Song, Y.; Kloepper, J.W.; Fadamiro, H. Rhizobacteria activates (+)-δ-cadinene synthase genes and induces systemic resistance in cotton against beet armyworm (Spodoptera exigua). Plant Cell Environ. 2016, 39, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Acero, J.L.; Benítez, F.J.; Real, F.J.; González, M. Chlorination of organophosphorus pesticides in natural waters. J. Hazard. Mater. 2008, 153, 320–328. [Google Scholar] [CrossRef]
- Kole, R.K.; Banerjee, H.; Bhattacharyya, A. Monitoring of market fish samples for endosulfan and hexachlorocyclohexane residues in and around Calcutta. Bull. Environ. Contam. Toxicol. 2001, 67, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Flores, M. Effects of glyphosate on nitrogen fixation of free-living heterotrophic bacteria. Lett. Appl. Microbiol. 1995, 20, 349–352. [Google Scholar] [CrossRef]
- Arias, R.N.; Fabra de Peretti, A. Effects of 2,4-dichlorophenoxyacetic acid on Rhizobium sp. growth and characterization of its transport. Toxicol. Lett. 1993, 68, 267–273. [Google Scholar] [CrossRef]
- Fabra, A.; Duffard, R.; Evangelista De Duffard, A. Toxicity of 2,4-dichlorophenoxyacetic acid to Rhizobium sp. in pure culture. Bull. Environ. Contam. Toxicol. 1997, 59, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Folmar, L.C.; Sanders, H.O.; Julin, A.M. Toxicity of the herbicide glyphosate and several of its formulations to fish and aquatic invertebrates. Arch. Environ. Contam. Toxicol. 1979, 8, 269–278. [Google Scholar] [CrossRef]
- Gadeva, P.; Dimitrov, B. Genotoxic effects of the pesticides Rubigan, Omite and Rovral in root-meristem cells of Crepis capillaris L. Mutat. Res.-Genet. Toxicol. Environ. Mutagen. 2008, 652, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Igbedioh, S.O. Effects of agricultural pesticides on humans, animals, and higher plants in developing countries. Arch. Environ. Health 1991, 46, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, A.; Longnecker, M.P.; Birnbaum, L.S.; Cogliano, J.; Kostyniak, P.; Moore, J.; Schantz, S.; Winneke, G. Characterization of potential endocrine-related health effects at low-dose levels of exposure to PCBs. Environ. Health Perspect. 1999, 107, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Crisp, T.M.; Clegg, E.D.; Cooper, R.L.; Wood, W.P.; Andersen, D.G.; Baetcke, K.P.; Hoffmann, J.L.; Morrow, M.S.; Rodier, D.J.; Schaeffer, J.E.; et al. Environmental endocrine disruption: An effects assessment and analysis. Environ. Health Perspect. 1998, 106, 11–56. [Google Scholar] [PubMed]
- Berlec, A. Novel techniques and findings in the study of plant microbiota: Search for plant probiotics. Plant Sci. 2012, 193–194, 96–102. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Novinscak, A.; Filion, M. Inoculation with the Plant-Growth-Promoting Rhizobacterium Pseudomonas fluorescens LBUM677 Impacts the Rhizosphere Microbiome of Three Oilseed Crops. Front. Microbiol. 2020, 11, 2534. [Google Scholar] [CrossRef] [PubMed]
- Grosch, R.; Dealtry, S.; Schreiter, S.; Berg, G.; Mendonça-Hagler, L.; Smalla, K. Biocontrol of Rhizoctonia solani: Complex interaction of biocontrol strains, pathogen and indigenous microbial community in the rhizosphere of lettuce shown by molecular methods. Plant Soil 2012, 361, 343–357. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on Lettuce Growth and Health under Pathogen Pressure and Its Impact on the Rhizosphere Bacterial Community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [Green Version]
- Scherwinski, K.; Wolf, A.; Berg, G. Assessing the risk of biological control agents on the indigenous microbial communities: Serratia plymuthica HRO-C48 and Streptomyces sp. HRO-71 as model bacteria. BioControl 2007, 52, 87–112. [Google Scholar] [CrossRef]
- Sang, M.K.; Kim, K.D. Plant growth-promoting rhizobacteria suppressive to Phytophthora blight affect microbial activities and communities in the rhizosphere of pepper (Capsicum annuum L.) in the field. Appl. Soil Ecol. 2012, 62, 88–97. [Google Scholar] [CrossRef]
- Chen, Y.; Yan, F.; Chai, Y.; Liu, H.; Kolter, R.; Losick, R.; Guo, J.H. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Wang, N.; Xia, F.; Li, Q.; Wang, W. Impact of biocontrol agents Pseudomonas fluorescens 2P24 and CPF10 on the bacterial community in the cucumber rhizosphere. Eur. J. Soil Biol. 2013, 59, 36–42. [Google Scholar] [CrossRef]
- Kröber, M.; Wibberg, D.; Grosch, R.; Eikmeyer, F.; Verwaaijen, B.; Chowdhury, S.P.; Hartmann, A.; Pühler, A.; Schlüter, A. Effect of the strain Bacillus amyloliquefaciens FZB42 on the microbial community in the rhizosphere of lettuce under field conditions analyzed by whole metagenome sequencing. Front. Microbiol. 2014, 5, 252. [Google Scholar] [CrossRef]
- Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pindo, M.; Pertot, I. Resilience of the natural phyllosphere microbiota of the grapevine to chemical and biological pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiter, S.; Ding, G.-C.; Grosch, R.; Kropf, S.; Antweiler, K.; Smalla, K. Soil type-dependent effects of a potential biocontrol inoculant on indigenous bacterial communities in the rhizosphere of field-grown lettuce. FEMS Microbiol. Ecol. 2014, 90, 718–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylla, J.; Alsanius, B.W.; Krüger, E.; Reineke, A.; Strohmeier, S.; Wohanka, W. Leaf microbiota of strawberries as affected by biological control agents. Phytopathology 2013, 103, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Hu, X.; Xu, X. Dispersal of Bacillus subtilis and its effect on strawberry phyllosphere microbiota under open field and protection conditions. Sci. Rep. 2016, 6, 22611. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Tao, J.; Liu, T.; Liu, Y.; Xiao, N.; Li, T.; Gu, Y.; Yin, H.; Meng, D. Responses of phyllosphere microbiota and plant health to application of two different biocontrol agents. AMB Express 2019, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Favilla, M.; Macchia, L.; Gallo, A.; Altomare, C. Toxicity assessment of metabolites of fungal biocontrol agents using two different (Artemia salina and Daphnia magna) invertebrate bioassays. Food Chem. Toxicol. 2006, 44, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Grandlic, C.J.; Green, W.A.; Kenovuo, J.S.; McCann, R. Compositions and Methods for Controlling Head Blight Disease. U.S. Patent MX368976B, 23 October 2019. [Google Scholar]
- Chen, F.; Wang, M.; Zheng, Y.; Li, S.; Wang, H.; Han, D.; Guo, S. The effect of biocontrol bacteria on rhizosphere bacterial communities analyzed by plating and pcr-dgge. Curr. Microbiol. 2013, 67, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Wang, Y.; Li, J.; Shen, Q.; Zhang, R. Enhanced root colonization and biocontrol activity of Bacillus amyloliquefaciens SQR9 by abrB gene disruption. Appl. Microbiol. Biotechnol. 2013, 97, 8823–8830. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.S.; Agostini, F.; Leifert, C.; Killham, K.; Mullins, C.E. Influence of Soil Temperature and Matric Potential on Sugar Beet Seedling Colonization and Suppression of Pythium Damping-Off by the Antagonistic Bacteria Pseudomonas fluorescens and Bacillus subtilis. Phytopathology 2004, 94, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Samaras, A.; Nikolaidis, M.; Antequera-Gómez, M.L.; Cámara-Almirón, J.; Romero, D.; Moschakis, T.; Amoutzias, G.D.; Karaoglanidis, G.S. Whole Genome Sequencing and Root Colonization Studies Reveal Novel Insights in the Biocontrol Potential and Growth Promotion by Bacillus subtilis MBI 600 on Cucumber. Front. Microbiol. 2021, 11, 600393. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.R.; Marco, M.L. Phyllosphere Microbiota Composition and Microbial Community Transplantation on Lettuce Plants Grown Indoors Thomas. MBio 2014, 5, e01564-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Pusey, P.L.; Stockwell, V.O.; Mazzola, M. Epiphytic Bacteria and Yeasts on Apple Blossoms and Their Potential as Antagonists of Erwinia amylovora. Phytopathology 2009, 99, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Escalante, A.E.; Barbolla, L.J.; Ramírez-Barahona, S.; Eguiarte, L.E. The study of biodiversity in the era of massive sequencing. Rev. Mex. Biodivers. 2014, 85, 1249–1264. [Google Scholar] [CrossRef] [Green Version]
- Land, M.; Hauser, L.; Jun, S.R.; Nookaew, I.; Leuze, M.R.; Ahn, T.H.; Karpinets, T.; Lund, O.; Kora, G.; Wassenaar, T.; et al. Insights from 20 years of bacterial genome sequencing. Funct. Integr. Genomics 2015, 15, 141–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Yang, D.; Wang, D.; Miao, Y.; Shao, J.; Zhou, X.; Xu, Z.; Li, Q.; Feng, H.; Li, S.; et al. Whole transcriptomic analysis of the plant-beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 during enhanced biofilm formation regulated by maize root exudates. BMC Genom. 2015, 16, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, M.; Chopra, S.; Mortelmans, K.; Kodukula, K.; Talcott, C.; Galande, A.K. In Silico Pathway Analysis Predicts Metabolites that are Potential Antimicrobial Targets. J. Comput. Sci. Syst. Biol. 2011, 04, 20–26. [Google Scholar] [CrossRef]
- Padilla-González, G.F.; Frey, M.; Gómez-Zeledón, J.; Da Costa, F.B.; Spring, O. Metabolomic and gene expression approaches reveal the developmental and environmental regulation of the secondary metabolism of yacón (Smallanthus sonchifolius, Asteraceae). Sci. Rep. 2019, 9, 13178. [Google Scholar] [CrossRef]
- Covington, B.C.; McLean, J.A.; Bachmann, B.O. Comparative mass spectrometry-based metabolomics strategies for the investigation of microbial secondary metabolites. Nat. Prod. Rep. 2017, 34, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Wielkopolan, B.; Obrępalska-Stęplowska, A. Three-way interaction among plants, bacteria, and coleopteran insects. Planta 2016, 244, 313–332. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, P.T.; Whiteman, N.K. Insect herbivory reshapes a native leaf microbiome. Nat. Ecol. Evol. 2020, 4, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Nair, S. Dynamics of Insect–Microbiome Interaction Influence Host and Microbial Symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef]
- Sugio, A.; Dubreuil, G.; Giron, D.; Simon, J.C. Plant-insect interactions under bacterial influence: Ecological implications and underlying mechanisms. J. Exp. Bot. 2015, 66, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Prasad, V.; Chauhan, P.S.; Lata, C. Bacillus amyloliquefaciens Confers Tolerance to Various Abiotic Stresses and Modulates Plant Response to Phytohormones through Osmoprotection and Gene Expression Regulation in Rice. Front. Plant Sci. 2017, 8, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Khurana, S.M.P. Plant Growth-Promoting Rhizobacteria for Alleviating Abiotic Stresses in Medicinal Plants; Springer: Cham, Switzerland, 2015; pp. 167–200. [Google Scholar]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Zhang, N.; Shen, Q.; Zhang, R. Beneficial Rhizobacterium bacillus amyloliquefaciens SQR9 induces plant salt tolerance through spermidine production. Mol. Plant-Microbe Interact. 2017, 30, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Prudent, M.; Salon, C.; Souleimanov, A.; Emery, R.J.N.; Smith, D.L. Soybean is less impacted by water stress using Bradyrhizobium japonicum and thuricin-17 from Bacillus thuringiensis. Agron. Sustain. Dev. 2015, 35, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Köberl, M.; Müller, H.; Ramadan, E.M.; Berg, G. Desert farming benefits from microbial potential in arid soils and promotes diversity and plant health. PLoS ONE 2011, 6, e24452. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Mirseyed Hosseini, H.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant Growth-Promoting Rhizobacteria Enhance Salinity Stress Tolerance in Okra through ROS-Scavenging Enzymes. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.M.; Khan, A.L.; Waqas, M.; You, Y.H.; Hamayun, M.; Joo, G.J.; Shahzad, R.; Choi, K.S.; Lee, I.J. Gibberellin-producing Serratia nematodiphila PEJ1011 ameliorates low temperature stress in Capsicum annuum L. Eur. J. Soil Biol. 2015, 68, 85–93. [Google Scholar] [CrossRef]
- Fernandez, O.; Theocharis, A.; Bordiec, S.; Feil, R.; Jacquens, L.; Clément, C.; Fontaine, F.; Barka, E.A. Burkholderia phytofirmans PsJN acclimates grapevine to cold by modulating carbohydrate metabolism. Mol. Plant-Microbe Interact. 2012, 25, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Slininger, P.J.; Schisler, D.A.; Ericsson, L.D.; Brandt, T.L.; Frazier, M.J.; Woodell, L.K.; Olsen, N.L.; Kleinkopf, G.E. Biological control of post-harvest late blight of potatoes. Biocontrol Sci. Technol. 2007, 17, 647–663. [Google Scholar] [CrossRef]
- Raupach, G.S.; Kloepper, J.W. Mixtures of plant growth-promoting rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology 1998, 88, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, D.P.; Lohrke, S.M.; Meyer, S.L.F.; Buyer, J.S.; Bowers, J.H.; Baker, C.J.; Li, W.; De Souza, J.T.; Lewis, J.A.; Chung, S. Biocontrol agents applied individually and in combination for suppression of soilborne diseases of cucumber. Crop Prot. 2005, 24, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Muthukumar, A.; Eswaran, A.; Sangeetha, G. Induction of systemic resistance by mixtures of fungal and endophytic bacterial isolates against Pythium aphanidermatum. Acta Physiol. Plant. 2011, 33, 1933–1944. [Google Scholar] [CrossRef]
- Gyenis, L.; Anderson, N.A.; Ostry, M.E. Biological control of Septoria leaf spot disease of hybrid poplar in the field. Plant Dis. 2003, 87, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Jeffries, P.; Pautasso, M.; Jeger, M.J. Combined use of biocontrol agents to manage plant diseases in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno-Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A compatible interaction controlling Fusarium wilt of cape gooseberry. Sci. Rep. 2020, 10, 6857. [Google Scholar] [CrossRef] [Green Version]
- Ayoubi, N.; Zafari, D.; Mirabolfathy, M. Evaluation of β-1,3-glucanase and β-1,4-glucanase enzymes production in some Trichoderma species. Arch. Phytopathol. Plant Prot. 2014, 47, 1929–1941. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.H.; Bernard, S.M.; Andersen, G.L.; Chen, W. Developing microbe-plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2009, 2, 428–440. [Google Scholar] [CrossRef]
- Gera Hol, W.H.; Martijn Bezemer, T.; Biere, A. Getting the ecology into interactions between plants and the plant growth-promoting bacterium Pseudomonas fluorescens. Front. Plant Sci. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 2019, 219, 26–39. [Google Scholar] [CrossRef]
- Strobel, G. The emergence of endophytic microbes and their biological promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Muthukumar, A.; Udhayakumar, R.; Naveenkumar, R. Role of Bacterial Endophytes in Plant Disease Control; Springer: Cham, Switzerland, 2017; pp. 133–161. [Google Scholar] [CrossRef]
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of endophytes for sustainable agricultural intensification. Mol. Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, M.O. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef] [PubMed]
- Hogervorst, P.A.M.; van den Akker, H.C.M.; Glandorf, D.C.M.; Klaassen, P.; van der Vlugt, C.J.B.; Westra, J. Assessment of Human Health and Environmental Risks of New Developments in Modern Biotechnology; RIVM Letter Report 2018-0089; National Institute for Public Health and the Environment: Utrecht, The Netherlands, 2018; 85p. [Google Scholar] [CrossRef]
- Scheepmaker, J.W.A.; Hogervorst, P.A.M.; Glandorf, D.C.M. Future Introductions of Genetically Modified Microbial Biocontrol Agents in the Eu: Are Current eu Legislation and Risk Assessment Fit for Purpose? RIVM Letter Report 2016-0057; National Institute for Public Health and the Environment: Utrecht, The Netherlands, 2016. [Google Scholar]




{kind=link}
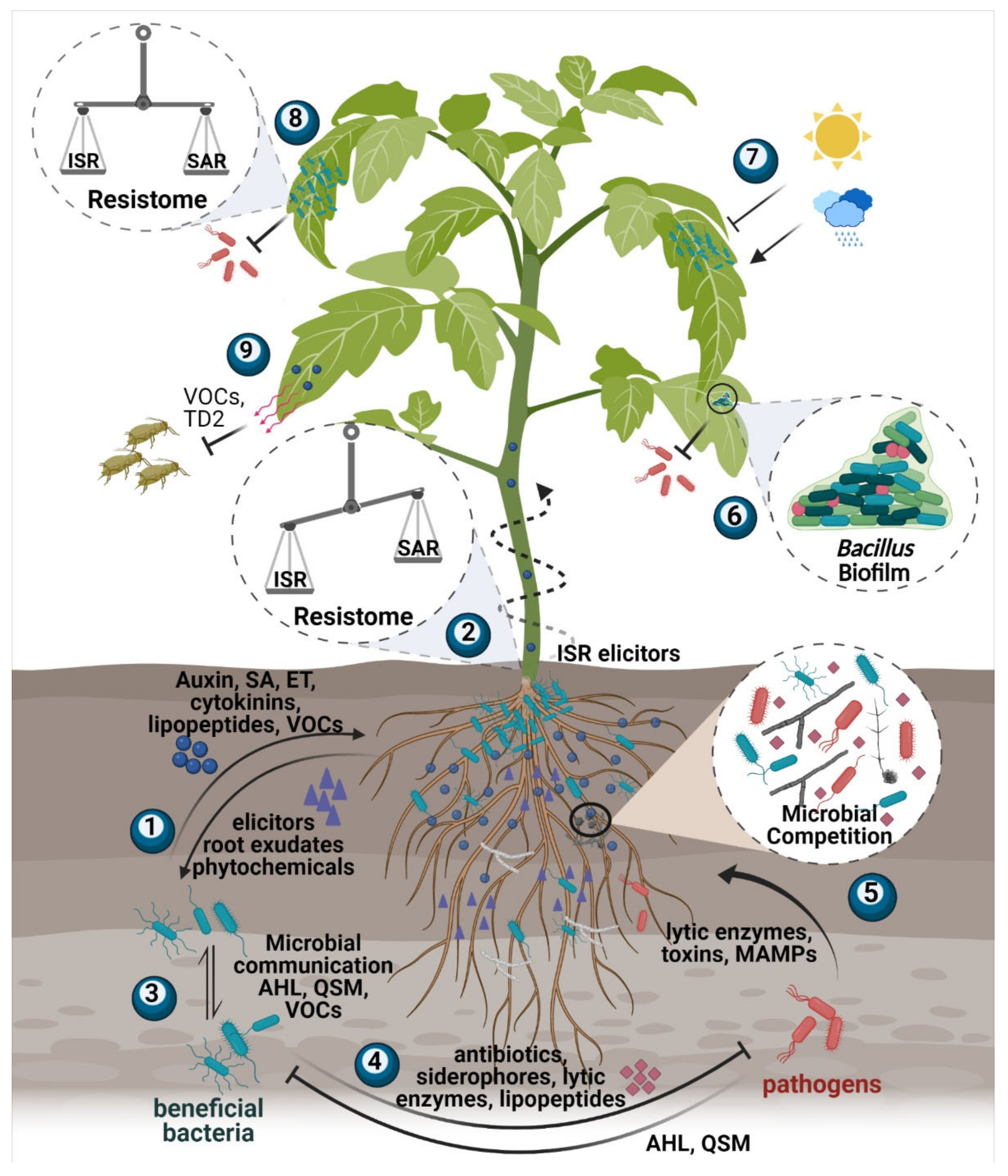{kind=link}
{kind=link}
| Bacterial BCAs | Brand Name | Manufacturer | Formulation | Application | Registration | Target Fungal Diseases | Target Bacterial Diseases | Crops | Authorizations | |
|---|---|---|---|---|---|---|---|---|---|---|
| Use | Region | |||||||||
| Agrobacterium radiobacter K84 | Galltrol-A | AgBioChem | Cell culture | Cuts, roots, seeds and stems application | Bactericide | US | Crown gall (Agrobacterium tumefaciens) | Bact: Non-food bearing stone fruit and nut trees | US:40230-1 | |
| Gallex | Flowable emulsion | Gals and cuts application | Crown gall (Agrobacterium tumefaciens), olive knot (Pseudomonas savastanoi) | Bact: Oleander, olive, ornamentals, stone and nut fruit trees | US:40230-2 | |||||
| Agrobacterium radiobacter (syn. strain K1062 | Nogall | Basf | Cells in peat carrier (wettable powder) | Cuts, roots, seeds and stems application | Bactericide | US | Crown gall (Agrobacterium Tumefaciens) | Bact: Non-food bearing stone fruit and nut trees, ornamentals | US:62388-1 | |
| Bacillus amyloliquefaciens D747 | Double Nickel 55 | Certis USA L.L.C | Bacterial endospores (wettable powder) | Chemigation, cut/root, foliar and ground application | Bactericide, fungicide | EU | Blasts, blights, blotches, damping-off diseases, downy mildews, drops, eutypa, flyspecks, leaf curls, melanoses, molds, mummy berry, phomopsis, powdery mildews, rots, rusts, scabs, scurfs, shanks, shot hole, sigatoka, smuts, spots, vine decline | Blights, cankers, specks and spots (Erwinia spp. Pseudomonas spp., Xanthomonas spp.) | Both: Citrus, pome and stone fruit trees, coffee, herbs and spices, nut trees, tropical fruits, vegetables F:Berries, cereal grains, grapes, oilseed crops, pomegranates, sugar beets and tobacco | US: 70051-108 |
| Amylo-X | Foliar application | US | Blights, downy mildews, molds, powdery mildews, rots, sclerotinia | Fire blight (Erwinia amylovora), kiwi canker (Pseudomonas syringae pv. actinidiae) | Bact: Kiwifruit pome F: Berries grapes, mushrooms, and stone fruit trees, vegetables | EU: Reg. No 1316/2014 (Dossier complete (2011/253/EU) | ||||
| Bacillus amyloliquefaciens strain FZB24 | Taergo 2 | Novozymes | Bacterial endospores (wettable powder) | Cut/root, foliar and ground application, ground incorporation | Bactericide, fungicide | US | Damping off diseases, blights, downy mildews, molds, powdery mildews, sclerotinia, spots | Specks (Pseudomonas spp.), spots (Xanthomonas spp.) | Both: Vegetables F: Ornamentals | US:70127-12 |
| Taergo | Foliar application | EU | Blights, damping off diseases, downy mildews, molds, powdery mildews, rots, spots | Specks (Pseudomonas spp.) | Both: vegetables F: Berries, grapes, | EU: Reg. (EU) 2017/806 | ||||
| Bacillus amyloliquefaciens (subtilis) MBI 600 | Serifel | Basf | Bacterial endospores (wettable power | Foliar and ground application | Bactericide, fungicide | US | Anthracnose, blights, blotches, damping off diseases, diebacks, dots, downy mildews, drops, esca, eutypa, flyspeck, molds, mummy berry, phomopsis, powdery mildews, rots, rusts, scabs, shanks, scorches, scurfs, shot hole, spots | Aerial stem rot (Erwinia carotovora) cankers, specks and spots (Pseudomonas spp., Xanthomonas spp.), fire blight (Erwinia amylovora), walnut blight (Xanthomonas campestris | Both: Cereal grains, grapes, herbs and spices, oilseed crops, soybean, tobacco, sugar beets berries citrus, pome and stone fruit trees, tree nuts, vegetables | US:71840-18 |
| Foliar application | Fungicide | EU | Molds, sclerotinia | F: Berries, herbs and spices, grapes | EU: Reg. (EU) 2016/1429Reg. (EU) 540/2011 | |||||
| Histick N/T (Beans, peanut, soybean) | Bacterial endospores (wettable power | Seed application | Fungicide | US | Damping off diseases | F: Beans, peanut, soybean | US: 71840-2 | |||
| Integral | Aqueous suspension | Ground application | Fungicide | US | Damping off diseases | F: Peanut, soybean | US: 71840-5 | |||
| Subtilex | Bacterial endospores (wettable powder) | Foliar and ground application | Fungicide | US | Damping off diseases, downy mildews, molds, powdery mildews | F: Bedding plants, ornamentals, tropical plants, vegetables | US: 71840-8 | |||
| Bacillus amyloliquefaciens (subtilis) strain QST 713 | Cease | Bioworks | Aqueous suspension | Aerial and ground application, chemigation | Bactericide, fungicide | US | Anthracose, blights, downy mildews, molds, powedery mildews, rots, rusts and sclerotinia | Blight (Pseudomonas syringae), fruit blotch (Acidovorax avenae), rots, specks and spots (Erwinia spp, Pseudomonas spp., Xanthomonas spp.) | Both: Berries, herbs and vegetables | US:264-1155-68539 |
| Rhapsody | Bayer Crop science | Concentrated cell suspension | Chemigation, foliar and ground application | Bactericide, fungicide | US | Anthracnose, brown patch, damping off diseases, gray mold, spots, powdery mildews, red thread, rust, septoria | Soft rot (Erwinia spp.), spots (Erwinia spp., Pseudomonas spp., Xanthomonas spp.) | Both: Landscape plants F: Berries, citrus, pome and stone fruit trees, golf courses lawns, turfs | US:264-1155 | |
| Serenade Aso | Concentrated cell suspension | Foliar and ground application | Bactericide, fungicide | US | Anthracnose, bakanae, blights, molds, damping off diseases, melanose, mummy berry, phomopsis, pink root, post bloom fruit drop, powdery mildews, ramularia, rots, rusts, scabs, sclerotinia, shot hole, sigatoka, smut, spots, web blotch | Fruit blotch (Actinovorax avenae), blights, cankers, rots, specks and spots (Erwinia spp., Pseudomonas spp., Xanthomonas spp.), gumming disease (Xanthomonas spp.), olive knot (Pseudomonas savastanoi), pustule (Xanthomonas spp.) | Both: Tropical fruits, vegetables soybeans, sugarcanes, tree nuts, oilseed crops, olive, kiwifruit, F: Cereal grains, coffee, cotton, herbs and spices, grass seeds, grapes, nongrass animal feeds, peanut, pomegranate, tobacco | US:264-1152 | ||
| Chemigation, foliar and ground application | EU | Anthracnose, blights, blotches, clubroot, molds, damping off diseases, downy mildews, phomopsis, powdery mildews, rots, rusts, scabs, sclerotinia, sigatoga, spots | Blights, cankers, rots, specks and spots (Erwinia spp., Pseudomonas spp., Xanthomonas spp.) | Both: Citrus, pome and stone fruit trees, vegetables tropical fruits, pomegranate, Berries, F: herbs and spices, grapes, mushrooms, oilseed crops, ornamentals, tobacco, sugar beets | EU: Reg. (EU) 2015/1396Reg. (EU) 2020/421Reg. (EU) No 540/2011 (07/6/EC, Reg. (EU) 2019/168,Reg. (EU) 2018/524,Reg. (EU) No 487/2014) | |||||
| Serenade Max | Bacterial endospores (wettable powder) | Aerial and foliar application, chemigation ground incorporation | Bactericide, fungicide | US | Anthracnose, blights, blotches, molds, damping off diseases, downy mildews, eutypa, flyspeck, melanose, mummy berry, phomopsis, post bloom fruit drop, powdery mildews, rots, rusts, scabs, smuts, shot hole, spots | Fruit blotch (Actinovorax avenae), blights, cankers, rots, specks and spots (Erwinia spp., Pseudomonas spp., Xanthomonas spp.), gumming disease (Xanthomonas spp.), olive knot (Pseudomonas savastanoi), pustule (Xanthomonas spp.) | Both: Berries, cereal grains, pome and stone fruit trees, tree nuts, tropical fruits, vegetables F: citrus, grass seeds, grapes, nongrass animal feeds, oilseed crops, peanut | US: 264-1151 | ||
| Foliar application | EU | Blights, molds, rusts, scabs, sclerotinia | Blights, cankers, rots, specks and spots (Erwinia spp., Pseudomonas spp., Xanthomonas spp.), crown gall (Agrobacterium tumefaciens | Both: Berries, herbs and spices, grapes, pome and stone fruit trees, tobacco, tropical fruits, vegetables | Reg. (EU) 2015/1396Reg. (EU) 2020/421Reg. (EU) No 540/2011 (07/6/EC, Reg. (EU) 2019/168,Reg. (EU) 2018/524,Reg. (EU) No 487/2014) | |||||
| Serenade Opti | Bacterial endospores (wettable powder) | Aerial and ground application, chemigation | Bactericide, fungicide | US | Anthracnose, blights, blotches, molds, flyspeck, mummy berry, phomopsis, post bloom fruit drop, powdery mildews, rots, rusts, sclerotinia, shot hole, spots | Aerial Stem Rot (Erwinia carotovora), canker, (Pseudomonas spp.), fire blight (Erwinia amylovora), shot hole (Xanthomonas pruni), spots (Pseudomonas spp., Xanthomonas spp.), | Botth: Berries, pome and stone fruit trees, tree nuts, vegetables citrus, pomegranate, stevia F: Herbs and spices, grapes, kiwifruit, oilseed crops, peanut | US: 264-1160 | ||
| Serenade Soil | Concentrated cell suspension | Ground application | Bactericide, fungicide | US | damping off diseases, pink root, rots, sclerotinia | Rots (Erwinia spp.) | Both: Vegetables F: Berries, citrus fruit trees, peanut | US: 264-1152 | ||
| Jazz | Bacterial endospores (wettable powder) | Chemigation, ground incorporation | Fungicide | US | Molds | F: Mushrooms | US: 264-1151 | |||
| Bacillus mycoides isolate J | Lifeguard | Certis USA L.L.C | Bacterial endospores (wettable powder) | Aerial and ground application, chemigation | Bactericide, fungicide | US | Anthracnose, blights, blotches, flyspeck, molds, downy mildews, mummy berry, powdery mildews, rusts, scabs, spots | Canker, spot and speck (Pseudomonas spp., Xanthomonas spp.), fire blight (Erwinia amylovora) | Both: Citrus and pome fruit trees, vegetables, grapes, peanuts, tobacco | US:70051-119 |
| Bacillus pumilus QST 2808 | Ballad Plus | Bayer Crop Science | Aqueous suspension of bacterial endospores, solids and solubles | Aerial and ground application, chemigation | Bactericide, fungicide | US | Blights, molds, downy mildews, powdery mildews, rots, rusts, smuts, spots, ramularia | Blights and speck (Pseudomonas spp., Xanthomonas spp.), pustule (Xanthomonas spp.) | Both: Cereal grains, oilseed crops, vegetables F: Grass grown for seed production, sugar beets | US:264-1153 |
| Bay 2000 | Concentrated cell suspension | Seed application | Fungicide | Damping off diseases | F: Cereal grains, vegetables | US:264-118 | ||||
| Sonata | Water suspension of bacterial endospores, solids and solubles | Aerial and ground application, chemigation | Fungicide | Blights, downy mildews, powdery mildews, rusts, scabs, spots | F: Berries, cereal grains, citrus, pome and stone fruit trees, herbs and spices, grapes, grass grown for seed production oilseed crops, roses, sugar beets, sweet corn, tree nuts, vegetables | US:264-1153 | ||||
| Concentrated cell suspension | Foliar application | Fungicide | EU | Powdery mildews | F: Herbs and spices, grapes, seed production, tobacco, vegetables | EU: Reg. (EU) No 485/2014 (2011/253/EU) | ||||
| Pseudomonas fluorescens A506 | BlightBan A506 | Nufarm Americas Inc. | Bacterial endospores (wettable powder) | Foliar application | Bactericide, fungicide | US | Molds | Fire blight (Erwinia amylovora), sour rot (Acetobacter spp.) | Both: Berries, pome and stone fruit trees, vegetables | US:228-710 |
| Sreptomyces lydicus WYE 108 | Actinovate AG | Novozymes | Bacterial endospores (wettable powder) | Chemigation, cutting/root, foliar, ground and seed application | Bactericide, fungicide | US | Damping off diseases, molds, rots | Bacterial spot (Xanthomonas perforans), walnut blight (Xanthomonas arboricola pv. juglandis) | Both: Stone fruit, vegetables F: Berries, cereal grains, citrus, pome fruit trees, herbs and spices, grapes, mushroom, oilseed crops, soybean, tree nuts, tropical fruits | US:73314-1 |
| Actinovate Lawn and Garden | Foliar, ground application | Bactericide, fungicide | US | Club root, damping off diseases, downy mildews, leaf curl, molds, patches, powdery mildews, rots, spots | Bacterial spot (Xanthomonas perforans), bacterial blast (Pseudomonas syringae), citrus canker (Xanthomonas axopodis pv. citri) | Both: Lawns, pome and stone fruit trees, ornamentals, vegetables | US:73314-1 | |||
| Foliar, ground application | Fungicide | EU | blights, damping off diseases, downy mildews, powdery mildews, rusts, spots | F: Lawns, ornamentals | Reg. (EU) No 917/2014 (Dossier complete (2011/253/EU)) | |||||
| Actinovate SP | Cut/root, ground and seed application | Fungicide | US | Anthracnose, blights, damping off diseases, downy mildews, molds, patches, powdery mildews, rusts, spots | F: Lawns, ornamentals, vegetables | US:73314-1 | ||||
| Actinovate STP | Dry seed application | Collapses, damping off diseases, rots, smuts | F: Cereal grains, cotton, herbs and spices, oilseed crops, peanut, sugar beets, vegetables | US:73314-4 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimopoulou, A.; Theologidis, I.; Varympopi, A.; Papafotis, D.; Mermigka, G.; Tzima, A.; Panopoulos, N.J.; Skandalis, N. Shifting Perspectives of Translational Research in Bio-Bactericides: Reviewing the Bacillus amyloliquefaciens Paradigm. Biology 2021, 10, 1202. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111202
Dimopoulou A, Theologidis I, Varympopi A, Papafotis D, Mermigka G, Tzima A, Panopoulos NJ, Skandalis N. Shifting Perspectives of Translational Research in Bio-Bactericides: Reviewing the Bacillus amyloliquefaciens Paradigm. Biology. 2021; 10(11):1202. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111202
Chicago/Turabian StyleDimopoulou, Anastasia, Ioannis Theologidis, Adamantia Varympopi, Dimitris Papafotis, Glykeria Mermigka, Aliki Tzima, Nick J. Panopoulos, and Nicholas Skandalis. 2021. "Shifting Perspectives of Translational Research in Bio-Bactericides: Reviewing the Bacillus amyloliquefaciens Paradigm" Biology 10, no. 11: 1202. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10111202