
Exploring Interactions between Primary Hepatocytes and Non-Parenchymal Cells on Physiological and Pathological Liver Stiffness


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
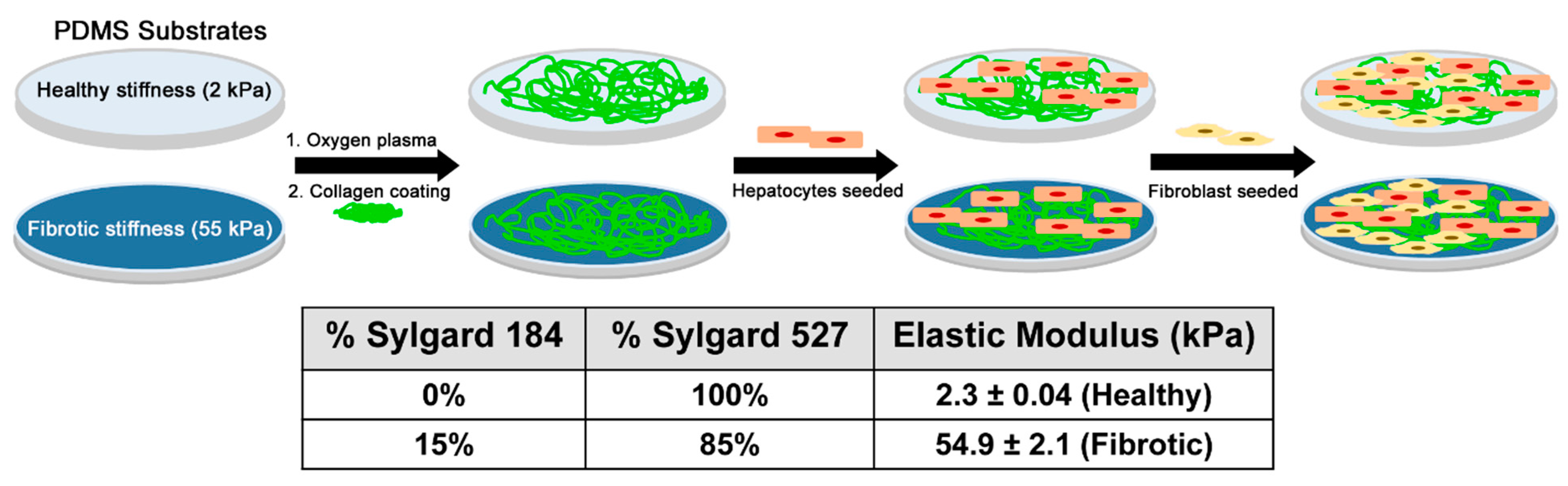
2.1. Preparation of PDMS Substrates
2.2. Collagen Coating of the Culture Substrates
2.3. Isolation and Culture of Primary Hepatocytes
2.4. Primary Hepatocyte Culture Medium
2.5. Culture of 3T3 Fibroblasts
2.6. Coculture on PDMS Surfaces
2.7. Differential Trypsinization for Separation of Fibroblast from Primary Hepatocytes
2.8. Urea Assay
2.9. Albumin Assay
2.10. Cytochrome P450 Activity Assay
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. Measuring Elastic Modulus of the PDMS Substrates
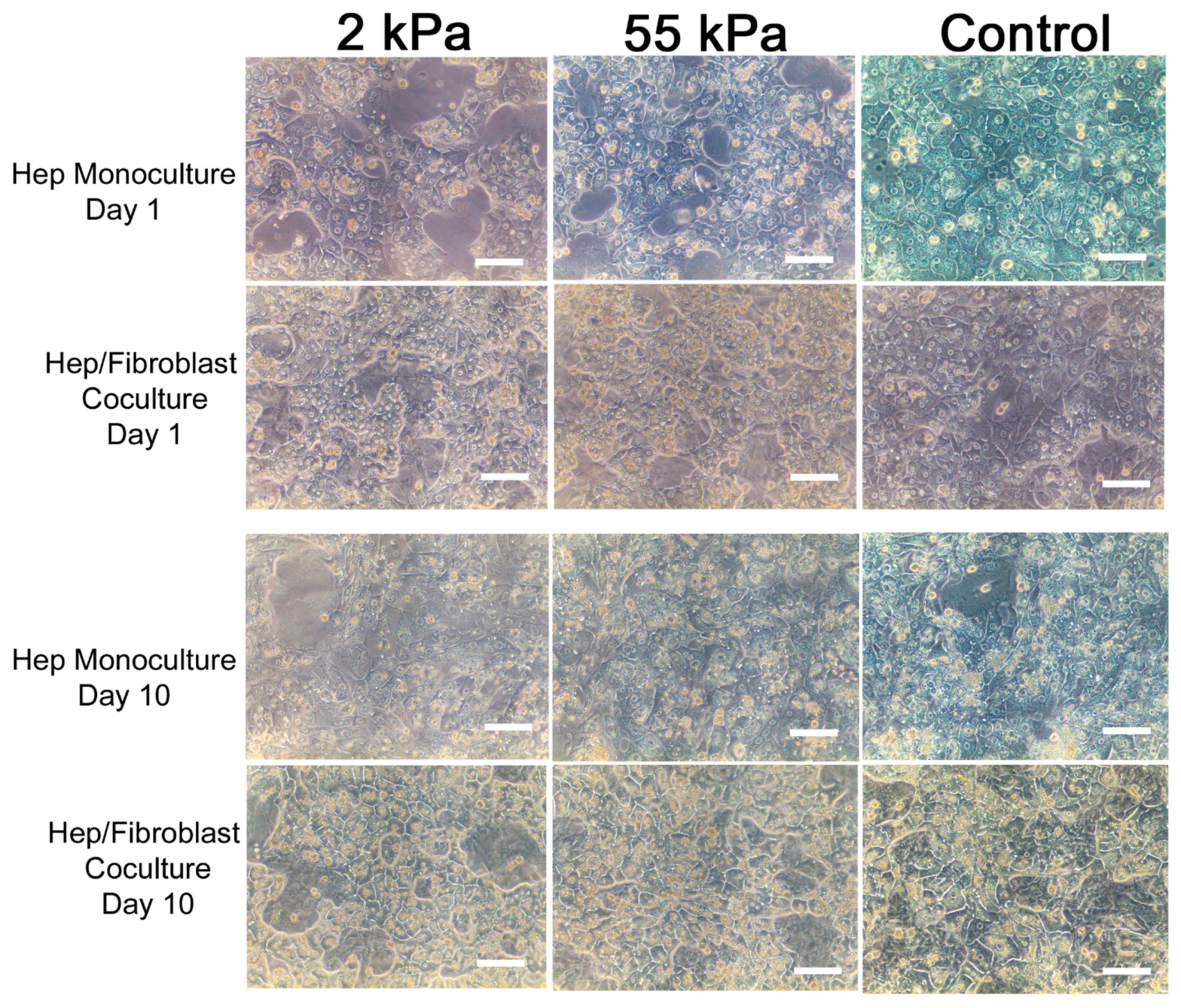
3.2. Primary Hepatocytes/Fibroblast Coculture on PDMS Substrates
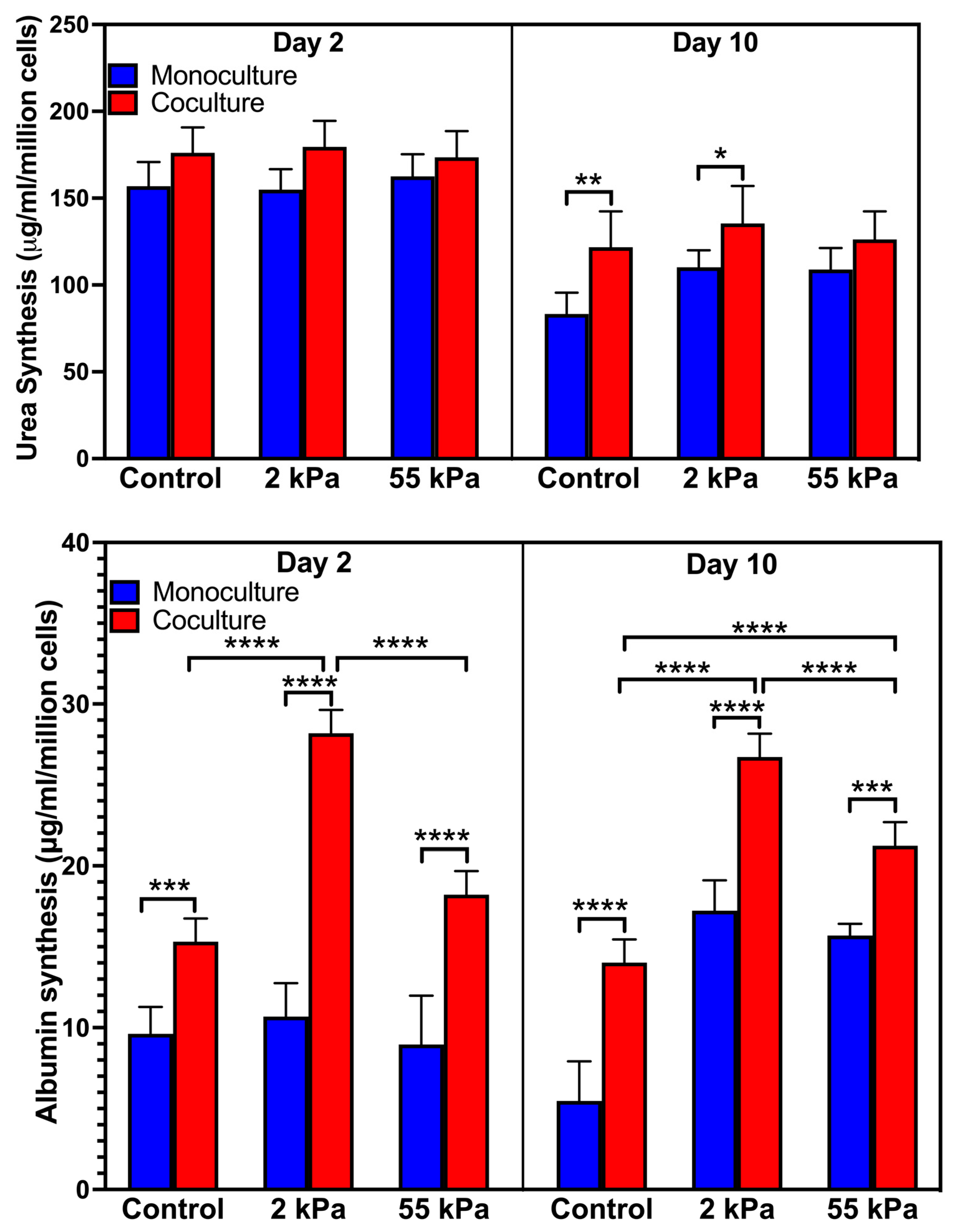
3.3. Effect of Stiffness on Primary Hepatocytes Urea Production in the Coculture
3.4. Effect of Stiffness on Primary Hepatocytes Albumin Synthesis in Coculture
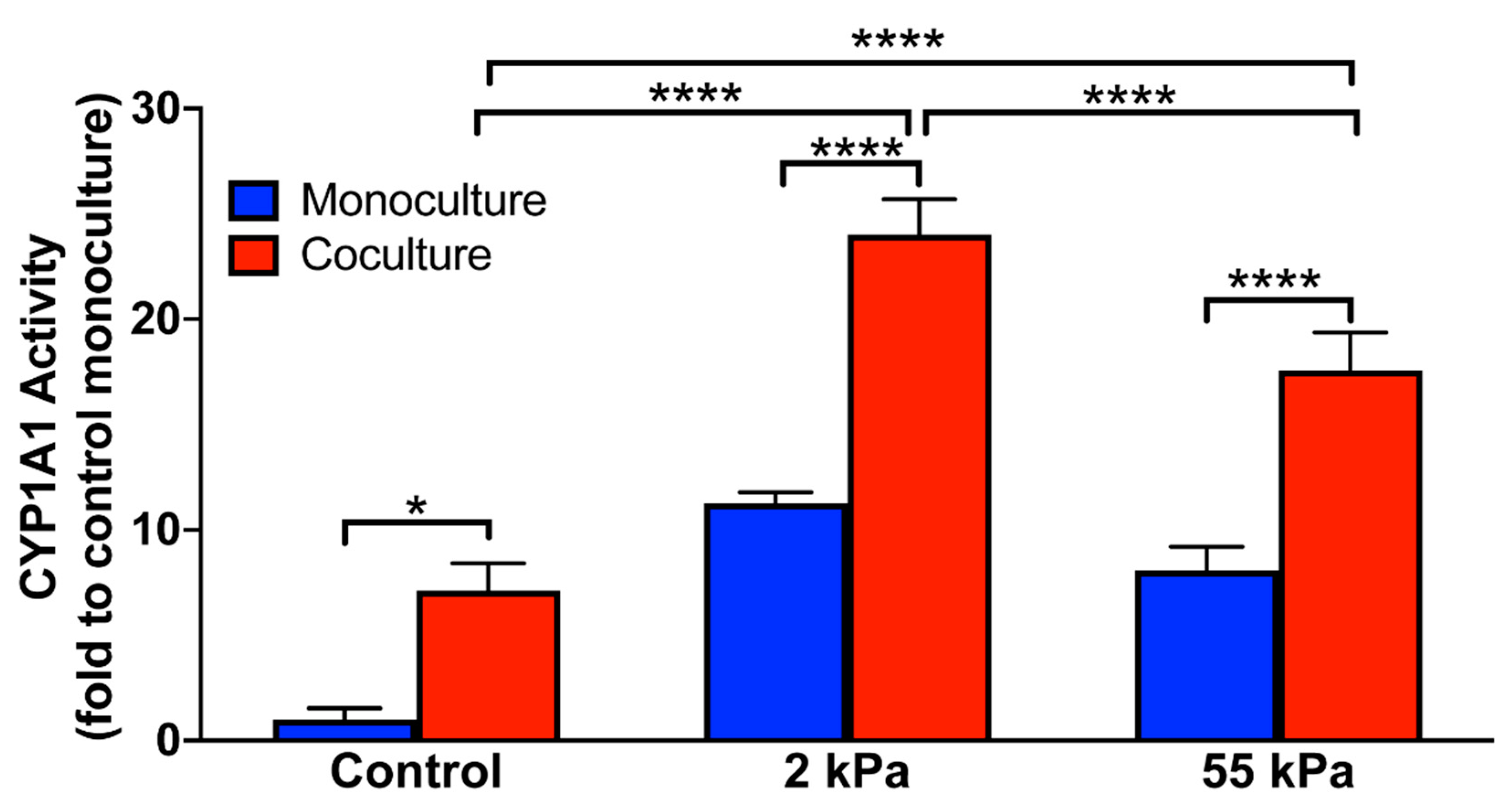
3.5. Effect of Stiffness on Hepatocytes CYP1A1 Activity in Coculture
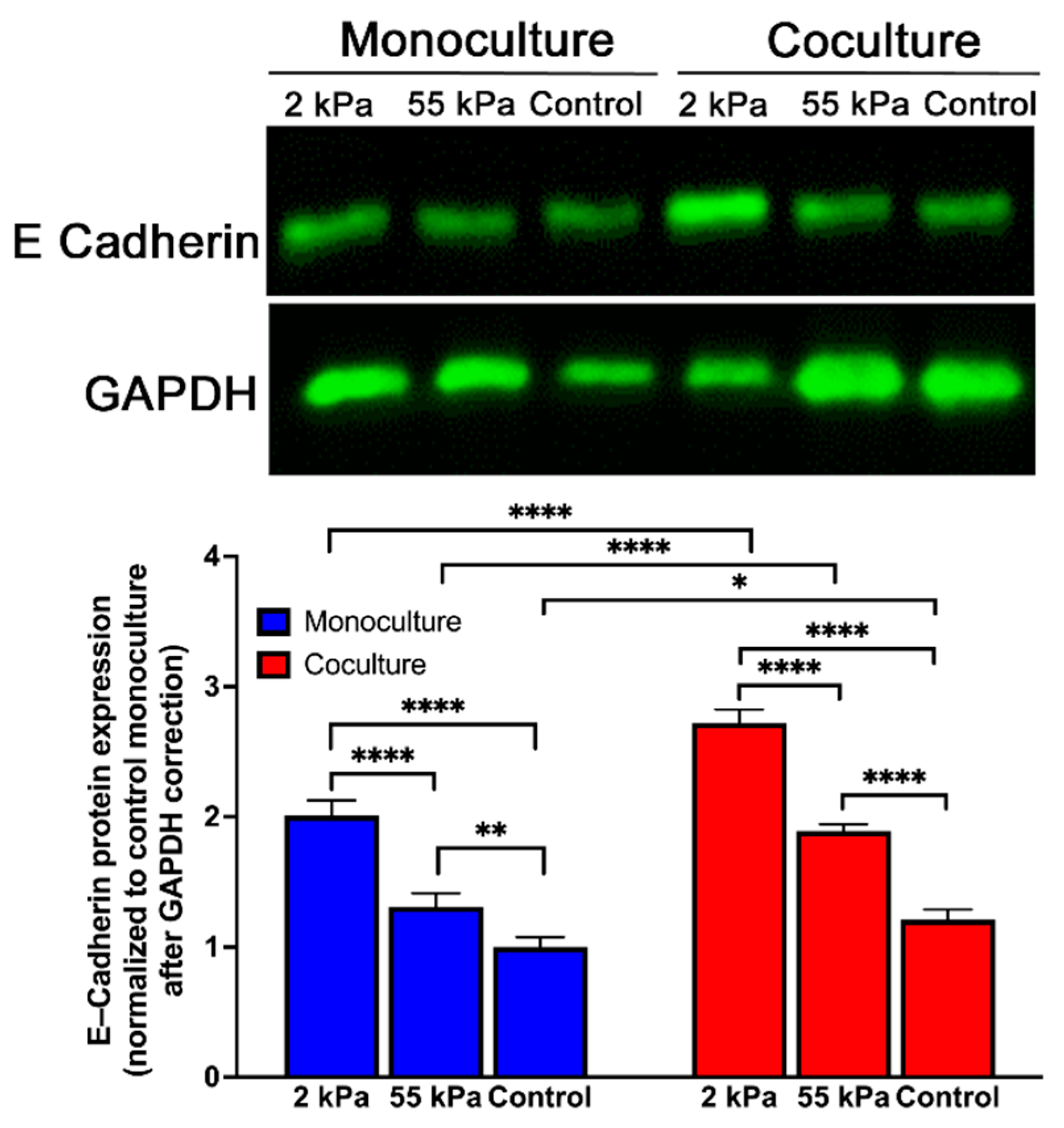
3.6. Effect of Stiffness Primary Hepatocytes E-Cadherin Expression in Coculture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedman, S.L. Hepatic Fibrosis: Emerging Therapies. Dig. Dis. 2015, 33, 504–507. [Google Scholar] [CrossRef]
- Trautwein, C.; Friedman, S.L.; Schuppan, D.; Pinzani, M. Hepatic fibrosis: Concept to treatment. J. Hepatol. 2015, 62, S15–S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.A.; Wallace, M.C.; Friedman, S.L. Pathobiology of liver fibrosis: A translational success story. Gut 2015, 64, 830–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Castera, L. Non-invasive assessment of liver fibrosis in chronic hepatitis C. Hepatol. Int. 2011, 5, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.; Talwalkar, J.A.; Glaser, K.J.; Manduca, A.; Grimm, R.C.; Rossman, P.J.; Fidler, J.L.; Ehman, R.L. Assessment of hepatic fibrosis with magnetic resonance elastography. Clin. Gastroenterol. Hepatol. 2007, 5, 1207–1213 e1202. [Google Scholar] [CrossRef] [Green Version]
- Arena, U.; Vizzutti, F.; Corti, G.; Ambu, S.; Stasi, C.; Bresci, S.; Moscarella, S.; Boddi, V.; Petrarca, A.; Laffi, G.; et al. Acute viral hepatitis increases liver stiffness values measured by transient elastography. Hepatology 2008, 47, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Foucher, J.; Chanteloup, E.; Vergniol, J.; Castera, L.; Le Bail, B.; Adhoute, X.; Bertet, J.; Couzigou, P.; de Ledinghen, V. Diagnosis of cirrhosis by transient elastography (FibroScan): A prospective study. Gut 2006, 55, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.; Moller, P.; Meyer, C.; Kolbe, C.C.; Bogs, C.; Pohlmann, A.; Schierwagen, R.; Praktiknjo, M.; Abdullah, Z.; Lehmann, J.; et al. Increase in liver stiffness after transjugular intrahepatic portosystemic shunt is associated with inflammation and predicts mortality. Hepatology 2018, 67, 1472–1484. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Horn, A.; Duckers, H.; Yagmur, E.; Sanson, E.; Bruensing, J.; Buendgens, L.; Voigt, S.; Trautwein, C.; Tacke, F. Increased liver stiffness denotes hepatic dysfunction and mortality risk in critically ill non-cirrhotic patients at a medical ICU. Crit. Care 2011, 15, R266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, M.; Shimakawa, Y.; Njie, R.; Njai, H.F.; Nayagam, S.; Khalil, M.; Goldin, R.; Ingiliz, P.; Taal, M.; Nyan, O.; et al. Food intake increases liver stiffness measurements and hampers reliable values in patients with chronic hepatitis B and healthy controls: The PROLIFICA experience in The Gambia. Aliment. Pharmacol. Ther. 2014, 39, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millonig, G.; Friedrich, S.; Adolf, S.; Fonouni, H.; Golriz, M.; Mehrabi, A.; Stiefel, P.; Poschl, G.; Buchler, M.W.; Seitz, H.K.; et al. Liver stiffness is directly influenced by central venous pressure. J. Hepatol. 2010, 52, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Rockey, D.C. Hepatic blood flow regulation by stellate cells in normal and injured liver. Semin Liver Dis. 2001, 21, 337–349. [Google Scholar] [CrossRef]
- Arima, S.; Uto, H.; Ibusuki, R.; Kumamoto, R.; Tanoue, S.; Mawatari, S.; Oda, K.; Numata, M.; Fujita, H.; Oketani, M.; et al. Hypertension exacerbates liver injury and hepatic fibrosis induced by a choline-deficient L-amino acid-defined diet in rats. Int. J. Mol. Med. 2014, 33, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.B.; Bhathal, P.S.; O'Brien, P.E. Nonalcoholic fatty liver disease: Predictors of nonalcoholic steatohepatitis and liver fibrosis in the severely obese. Gastroenterology 2001, 121, 91–100. [Google Scholar] [CrossRef]
- Wells, R.G. Location, location, location: Cell-level mechanics in liver fibrosis. Hepatology 2016. [Google Scholar] [CrossRef] [Green Version]
- Wells, R.G. Tissue mechanics and fibrosis. Biochim. Biophys. Acta 2013, 1832, 884–890. [Google Scholar] [CrossRef] [Green Version]
- Tschumperlin, D.J.; Ligresti, G.; Hilscher, M.B.; Shah, V.H. Mechanosensing and fibrosis. J. Clin. Investig. 2018, 128, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Kennedy, L.; Liangpunsakul, S.; Kusumanchi, P.; Yang, Z.; Meng, F.; Glaser, S.; Francis, H.; Alpini, G. Intercellular Communication between Hepatic Cells in Liver Diseases. Int. J. Mol. Sci. 2019, 20, 2180. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.L. Hepatic fibrosis—overview. Toxicology 2008, 254, 120–129. [Google Scholar] [CrossRef]
- Michalopoulos, G.K.; DeFrances, M.C. Liver regeneration. Science 1997, 276, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Hurrell, T.; Kastrinou-Lampou, V.; Fardellas, A.; Hendriks, D.F.G.; Nordling, A.; Johansson, I.; Baze, A.; Parmentier, C.; Richert, L.; Ingelman-Sundberg, M. Human Liver Spheroids as a Model to Study Aetiology and Treatment of Hepatic Fibrosis. Cells 2020, 9, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbero-Becerra, V.J.; Giraudi, P.J.; Chavez-Tapia, N.C.; Uribe, M.; Tiribelli, C.; Rosso, N. The interplay between hepatic stellate cells and hepatocytes in an in vitro model of NASH. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2015, 29, 1753–1758. [Google Scholar] [CrossRef]
- Giraudi, P.J.; Becerra, V.J.; Marin, V.; Chavez-Tapia, N.C.; Tiribelli, C.; Rosso, N. The importance of the interaction between hepatocyte and hepatic stellate cells in fibrogenesis induced by fatty accumulation. Exp. Mol. Pathol. 2015, 98, 85–92. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell (Cambridge, MA, United States) 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, A.M.; Noce, V.; Battistelli, C.; Marchetti, A.; Grassi, G.; Cicchini, C.; Tripodi, M.; Amicone, L. Modulating the Substrate Stiffness to Manipulate Differentiation of Resident Liver Stem Cells and to Improve the Differentiation State of Hepatocytes. Stem Cells Int. 2016, 2016, 5481493. [Google Scholar] [CrossRef] [Green Version]
- Lozoya, O.A.; Wauthier, E.; Turner, R.A.; Barbier, C.; Prestwich, G.D.; Guilak, F.; Superfine, R.; Lubkin, S.R.; Reid, L.M. Regulation of hepatic stem/progenitor phenotype by microenvironment stiffness in hydrogel models of the human liver stem cell niche. Biomaterials 2011, 32, 7389–7402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidambi, S. Chapter 55. Stiffness and Hepatocytes Function In Vitro in “Liver Elastography-Clinical Use and Interpretation”. Liver Elastography Clin. Use Interpret. 2020. [Google Scholar] [CrossRef]
- Ganesan, M.; Dagur, R.S.; Makarov, E.; Poluektova, L.I.; Kidambi, S.; Osna, N.A. Matrix stiffness regulate apoptotic cell death in HIV-HCV co-infected hepatocytes: Importance for liver fibrosis progression. Biochem. Biophys. Res. Commun. 2018, 500, 717–722. [Google Scholar] [CrossRef]
- Natarajan, V.; Berglund, E.J.; Chen, D.X.; Kidambi, S. Substrate stiffness regulates primary hepatocyte functions. Rsc Adv. 2015, 5, 80956–80966. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.S.; Tung, J.C.; Zhou, V.X.; Grenert, J.P.; Malato, Y.; Rezvani, M.; Espanol-Suner, R.; Willenbring, H.; Weaver, V.M.; Chang, T.T. Physiological ranges of matrix rigidity modulate primary mouse hepatocyte function in part through hepatocyte nuclear factor 4 alpha. Hepatology 2016, 64, 261–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semler, E.J.; Lancin, P.A.; Dasgupta, A.; Moghe, P.V. Engineering hepatocellular morphogenesis and function via ligand-presenting hydrogels with graded mechanical compliance. Biotechnol. Bioeng. 2005, 89, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Deegan, D.B.; Zimmerman, C.; Skardal, A.; Atala, A.; Shupe, T.D. Stiffness of hyaluronic acid gels containing liver extracellular matrix supports human hepatocyte function and alters cell morphology. J. Mech. Behav. Biomed. Mater. 2015, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Park, S.A.; Shin, D.S.; Patel, D.; Raghunathan, V.K.; Kim, M.; Murphy, C.J.; Tae, G.; Revzin, A. Characterizing the effects of heparin gel stiffness on function of primary hepatocytes. Tissue Eng. Part A 2013, 19, 2655–2663. [Google Scholar] [CrossRef]
- Chen, A.A.; Khetani, S.R.; Lee, S.; Bhatia, S.N.; Van Vliet, K.J. Modulation of hepatocyte phenotype in vitro via chemomechanical tuning of polyelectrolyte multilayers. Biomaterials 2009, 30, 1113–1120. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Hang, R.; Wang, X.; Lin, N.; Zhang, X.; Tang, B. Matrix Stiffness in Three-Dimensional Systems Effects on the Behavior of C3A Cells. Artif. Organs 2013, 37, 166–174. [Google Scholar] [CrossRef]
- Ben-Ze'ev, A.; Robinson, G.S.; Bucher, N.; Farmer, S.R. Cell-cell and cell-matrix interactions differentially regulate the expression of hepatic and cytoskeletal genes in primary cultures of rat hepatocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 2161–2165. [Google Scholar] [CrossRef] [Green Version]
- Mooney, D.; Hansen, L.; Vacanti, J.; Langer, R.; Farmer, S.; Ingber, D. Switching from differentiation to growth in hepatocytes: Control by extracellular matrix. J. Cell. Physiol. 1992, 151, 497–505. [Google Scholar] [CrossRef]
- Hansen, L.K.; Wilhelm, J.; Fassett, J.T. Regulation of hepatocyte cell cycle progression and differentiation by type I collagen structure. Curr. Top. Dev. Biol. 2006, 72, 205–236. [Google Scholar] [CrossRef]
- Fassett, J.; Tobolt, D.; Hansen, L.K. Type I collagen structure regulates cell morphology and EGF signaling in primary rat hepatocytes through cAMP-dependent protein kinase A. Mol. Biol. Cell 2005, 17, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Khetani, S.R.; Chen, A.A.; Ranscht, B.; Bhatia, S.N. T-cadherin modulates hepatocyte functions in vitro. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 3768–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khetani, S.R.; Szulgit, G.; Del Rio, J.A.; Barlow, C.; Bhatia, S.N. Exploring interactions between rat hepatocytes and nonparenchymal cells using gene expression profiling. Hepatology 2004, 40, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.N.; Yarmush, M.L.; Toner, M. Controlling cell interactions by micropatterning in co-cultures: Hepatocytes and 3T3 fibroblasts. J. Biomed. Mater. Res. 1997, 34, 189–199. [Google Scholar] [CrossRef]
- Marois, Y.; Sigot-Luizard, M.-F.; Guidoin, R. Endothelial cell behavior on vascular prosthetic grafts: Effect of polymer chemistry, surface structure, and surface treatment. ASAIO J. 1999, 45, 272–280. [Google Scholar] [CrossRef]
- Park, J.H.; Park, K.D.; Bae, Y.H. PDMS-based polyurethanes with MPEG grafts: Synthesis, characterization and platelet adhesion study. Biomaterials 1999, 20, 943–953. [Google Scholar] [CrossRef]
- Sherman, M.A.; Kennedy, J.P.; Ely, D.L.; Smith, D. Novel polyisobutylene/polydimethyl siloxane bicomponent networks: III. Tissue compatibility. J. Biomater. Sci. Polym. Ed. 1999, 10, 259–269. [Google Scholar] [CrossRef]
- Bordenave, L.; Bareille, R.; Lefebvre, F.; Caix, J.; Baquey, C. Cytocompatibility study of NHLBI primary reference materials using human endothelial cells. J. Biomater. Sci. Polym. Ed. 1992, 3, 409–416. [Google Scholar] [CrossRef]
- van Kooten, T.G.; Whitesides, J.F.; von Recum, A.F. Influence of silicone (PDMS) surface texture on human skin fibroblast proliferation as determined by cell cycle analysis. J. Biomed. Mater. Res. 1998, 43, 1–14. [Google Scholar] [CrossRef]
- Ertel, S.I.; Ratner, B.D.; Kaul, A.; Schway, M.B.; Horbett, T.A. In vitro study of the intrinsic toxicity of synthetic surfaces to cells. J. Biomed. Mater. Res. 1994, 28, 667–675. [Google Scholar] [CrossRef]
- Dahrouch, M.; Schmidt, A.; Leemans, L.; Linssen, H.; Goetz, H. Synthesis and properties of poly(butylene terephthalate)-poly(ethylene oxide)-poly(dimethylsiloxane) block copolymers. Macromol. Symp. 2003, 199, 147–162. [Google Scholar] [CrossRef]
- Interrante, L.V.; Shen, Q.; Li, J. Poly(dimethylsilylenemethylene-co- dimethylsiloxane): A Regularly Alternating Copolymer of Poly(dimethylsiloxane) and Poly(dimethylsilylenemethylene). Macromolecules 2001, 34, 1545–1547. [Google Scholar] [CrossRef]
- Mueller, S. Noninvasive assessment of patients with alcoholic liver disease. Clin. Liver Dis. 2013, 2, 68–71. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Sandrin, L. Liver stiffness: A novel parameter for the diagnosis of liver disease. Hepatic Med. Evid. Res. 2010, 2, 49. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.N.; Balis, U.J.; Yarmush, M.L.; Toner, M. Probing heterotypic cell interactions: Hepatocyte function in microfabricated co-cultures. J. Biomater. Sci. Polym. Ed. 1998, 9, 1137–1160. [Google Scholar] [CrossRef] [PubMed]
- Owens, R.B.; Smith, H.S.; Hackett, A.J. Epithelial cell cultures from normal glandular tissue of mice. J. Natl. Cancer Inst. 1974, 53, 261–269. [Google Scholar] [CrossRef]
- Halaban, R.; Alfano, F.D. Selective elimination of fibroblasts from cultures of normal human melanocytes. Vitro 1984, 20, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Pauli, B.U.; Anderson, S.N.; Memoli, V.A.; Kuettner, K.E. The isolation and characterization in vitro of normal epithelial cells, endothelial cells and fibroblasts from rat urinary bladder. Tissue Cell 1980, 12, 419–436. [Google Scholar] [CrossRef]
- Rubio, S.; Cazares, O.; Macias, H.; Hinck, L. Generation of Mosaic Mammary Organoids by Differential Trypsinization. J. Vis. Exp. JoVE 2020. [Google Scholar] [CrossRef]
- O'Callaghan, A.R.; Morgan, L.; Daniels, J.T.; Lewis, M.P. Human-derived feeder fibroblasts for the culture of epithelial cells for clinical use. Regen. Med. 2016, 11, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Tibbitt, M.W.; Basta, L.; Anseth, K.S. Mechanical memory and dosing influence stem cell fate. Nat. Mater 2014, 13, 645–652. Available online: http://0-www-nature-com.brum.beds.ac.uk/nmat/journal/v13/n6/abs/nmat3889.html#supplementary-information (accessed on 3 June 2020). [CrossRef] [PubMed]
- Mak, T.M.; Huang, Y.P.; Zheng, Y.P. Liver fibrosis assessment using transient elastography guided with real-time B-mode ultrasound imaging: A feasibility study. Ultrasound Med. Biol. 2013, 39, 956–966. [Google Scholar] [CrossRef]
- Zhou, K.; Li, C.; Chen, S.; Nabi, G.; Huang, Z. Feasibility study of using the dispersion of surface acoustic wave impulse for viscoelasticity characterization in tissue mimicking phantoms. J. Biophotonics 2019, 12, e201800177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghe, P.V.; Berthiaume, F.; Ezzell, R.M.; Toner, M.; Tompkins, R.G.; Yarmush, M.L. Culture matrix configuration and composition in the maintenance of hepatocyte polarity and function. Biomaterials 1996, 17, 373–385. [Google Scholar] [CrossRef]
- Dunn, J.C.; Yarmush, M.L.; Koebe, H.G.; Tompkins, R.G. Hepatocyte function and extracellular matrix geometry: Long-term culture in a sandwich configuration. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1989, 3, 174–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogoke, O.; Maloy, M.; Parashurama, N. The science and engineering of stem cell-derived organoids-examples from hepatic, biliary, and pancreatic tissues. Biol. Rev. Camb. Philos. Soc. 2021, 96, 179–204. [Google Scholar] [CrossRef]
- Lee, J.Y.; Han, H.J.; Lee, S.J.; Cho, E.H.; Lee, H.B.; Seok, J.H.; Lim, H.S.; Son, W.C. Use of 3D Human Liver Organoids to Predict Drug-Induced Phospholipidosis. Int. J. Mol. Sci. 2020, 21, 2982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, H.; Deng, P.; Chen, W.; Guo, Y.; Tao, T.; Qin, J. In situ differentiation and generation of functional liver organoids from human iPSCs in a 3D perfusable chip system. Lab chip 2018, 18, 3606–3616. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natarajan, V.; Moeun, Y.; Kidambi, S. Exploring Interactions between Primary Hepatocytes and Non-Parenchymal Cells on Physiological and Pathological Liver Stiffness. Biology 2021, 10, 408. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050408
Natarajan V, Moeun Y, Kidambi S. Exploring Interactions between Primary Hepatocytes and Non-Parenchymal Cells on Physiological and Pathological Liver Stiffness. Biology. 2021; 10(5):408. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050408
Chicago/Turabian StyleNatarajan, Vaishaali, Youra Moeun, and Srivatsan Kidambi. 2021. "Exploring Interactions between Primary Hepatocytes and Non-Parenchymal Cells on Physiological and Pathological Liver Stiffness" Biology 10, no. 5: 408. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050408