
MicroRNA Sequences Modulated by Beta Cell Lipid Metabolism: Implications for Type 2 Diabetes Mellitus


Department of Biological and Biomedical Sciences, School of Health and Life Sciences, Glasgow Caledonian University, Glasgow G4 0BA, UK
*
Author to whom correspondence should be addressed.
Biology 2021, 10(6), 534; https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060534
Submission received: 15 May 2021
/
Revised: 8 June 2021
/
Accepted: 9 June 2021
/
Published: 15 June 2021
Abstract
:Simple Summary
At present, more than 450 million adults worldwide are living with diabetes, with a further 370 million individuals at risk of developing this condition. Diabetes is caused by loss of production of, or sensitivity to, insulin, the hormone which controls blood sugar levels. One key factor contributing to loss of insulin output from beta cells in pancreatic islets is the damaging effects of sugars and fats in the bloodstream. This review article sought to identify the changes in expression of small pieces of RNA (microRNA) which are reported to be caused in beta cells and islets by exposure to sugars and fats. These small RNA sequences alter the expression of networks of genes which can promote, or protect, against beta cell damage, and their levels in the bloodstream have also been used as markers of diabetes. The combined effects of these microRNA sequences in beta cells were predicted, and may help to inform drug discovery strategies.
Abstract
Alterations in lipid metabolism within beta cells and islets contributes to dysfunction and apoptosis of beta cells, leading to loss of insulin secretion and the onset of type 2 diabetes. Over the last decade, there has been an explosion of interest in understanding the landscape of gene expression which influences beta cell function, including the importance of small non-coding microRNA sequences in this context. This review sought to identify the microRNA sequences regulated by metabolic challenges in beta cells and islets, their targets, highlight their function and assess their possible relevance as biomarkers of disease progression in diabetic individuals. Predictive analysis was used to explore networks of genes targeted by these microRNA sequences, which may offer new therapeutic strategies to protect beta cell function and delay the onset of type 2 diabetes.
1. Introduction
The International Diabetes Federation (IDF) Atlas (2019) indicates that there are currently 463 million adults (29–79 y) living with diabetes, predicted to rise to 700 million by 2045, with a further ~374 million people at increased risk of developing type 2 diabetes mellitus [1]. Type 2 diabetes (T2D) is caused by a lack of insulin sensitivity in hepatic and peripheral tissues [2], combined with loss of insulin secretion due to decreased beta cell function and/or mass. Compensatory increases in insulin secretion precede T2D, and loss of this mechanism due to beta cell dysfunction or loss of beta cell mass, is a key factor in triggering frank manifestation of this disease [2,3].
1.1. Glucose-Stimulated Insulin Secretion from Pancreatic Beta Cells
Secretion of insulin from pancreatic beta cells is central in maintenance of whole-body glucose homeostasis (Figure 1). Glucose-stimulated insulin secretion (GSIS) occurs through a sequence of strictly controlled events, following a rise in blood glucose concentrations. Glucose is transported from the plasma across the cell membrane, via glucose transporter 1 (GLUT1) [4], where it becomes available for phosphorylation by free glucokinase (GCK) [5]. Glucokinase is activated by release from insulin granules following monomerisation of neuronal nitric oxide synthase (nNOS) [6,7,8]. The rise in glucose-6-phosphate increases glycolysis and aerobic respiration, and the resultant increase in ATP/ADP ratio causes closure of ATP-dependent K+ channels (KATP) and membrane depolarisation, facilitating opening of voltage-gated calcium channels (CaV). Influx of Ca2+ triggers exocytosis of insulin granules by interaction of insulin secretory granules with soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) complexes located at the cell membrane, comprised of vesicle-associated membrane protein 2 (VAMP2), synaptosomal-associated protein 25 (SNAP25) and syntaxin-1A (STX1A) [9,10,11]. Following restoration of glucose homeostasis, the pancreatic beta cell membrane potential is corrected by voltage-gated potassium channels (KV2.1 and KV3.2 in human beta cells) [12].
1.2. Loss of Insulin Secretion and Beta Cell Mass in Diabetes
Insulin production by pancreatic beta cells exhibits a remarkable degree of plasticity (reviewed in [13]), responding acutely to differing environmental conditions including starvation and over-nutrition. However, the chronic, persistently high, demand for insulin, which occurs during insulin resistance, can lead to progressive dysfunction, and eventual loss, of beta cells [14,15]. The evaluation of GSIS ex vivo in pancreatic islets from T2D donors shows uncoupling between glucose concentration and insulin secretion: insulin secretion does not change at basal glucose concentrations, but exhibits reduced capacity to respond appropriately to rising glucose levels in patients with T2D [16,17,18,19,20]. Xenotransplantation of human T2D islets into immunodeficient diabetic mice was unable to restore normoglycaemia, unlike the implantation of non-diabetic islets [17], demonstrating that T2D limits beta cell function.
Multiple mechanisms contribute to dysfunctional insulin secretion in T2D beta cells. The expression of glucose transporters and glucokinase (GCK) is lower in human T2D islets than in islets of healthy controls, indicating impaired glucose sensing and metabolism [21]. Type 2 diabetes alters the expression of genes encoding proteins with a wide range of functions affecting insulin secretion, such as Ca2+ trafficking (TMEM37, SUR1), mitochondrial metabolism (ALDOB, GPD2, FXYD2 and PCK1), cell cycling (P21/CIP, TTC39C) and fatty acid (FFAR4, TMEM97), insulin (IR), IGF-1 (IGF1R) and TNF (TNFRSF11A) receptor signalling [22,23,24,25,26,27]. There is also evidence of compromised mitochondrial function in T2D diabetic beta cells as a result of reduced enzymatic activity [16,28,29]. This can limit insulin secretion as it leads to a lower ATP/ADP ratio [30], and compromised Ca2+ influx. Protein and/or gene levels of SNARE complex, SNARE-modulating proteins syntaxin-1A, SNAP-25, VAMP-2, nSec1 (Munc), Munc 13-1, synaptotagmin V and synaptophysin, and components of the KATP channel (Kir6.2 and SUR1) were also lower in isolated pancreatic islets from diabetic patients, compared with controls, reflecting impaired insulin secretion in these individuals [31].
Loss of beta cell mass can occur during prediabetes [32]: at diagnosis, patients with T2D have often lost ~50% of their beta cell mass [33], from apoptosis and dedifferentiation [15,34,35]. Hyperinsulinaemia increases the production of hydrogen peroxide, activation of caspases and induces expression of inducible nitric oxide synthase (iNOS) and Tribbles homolog 3 (Trib3) in beta cells [36,37]. Prolonged exposure of rat islets or INS1-E cells to high concentrations of insulin leads to reduced phosphorylation of AktS473, reductions in phosphorylation of P70S6 kinase and ERK-1/2 kinase and increased apoptosis [38]. Hyperglycaemia increases glyceraldehyde-derived advanced glycation end products (AGE) [39], while signalling via the receptor for AGE (RAGE) results in increased cytochrome release and caspase activation [40]. Exposure to elevated levels of glucose in combination with high levels of free fatty acids (glucolipotoxicity) is also thought to be a significant contributor to increased apoptosis and loss of GSIS in beta cells [41,42] (below).
De-differentiation is another factor contributing to loss of beta cell mass in T2D [43,44,45]. Transition of beta cells to a progenitor-like state or α cell [46,47] is associated with downregulation of beta cell identity genes, upregulation of beta cell ‘forbidden’ genes and upregulation of stem-cell genes [15,46]. John et al. (2018) observed downregulation of beta cell identity genes, FoxO1, MafA and Nkx6.1, in the db/db murine model of T2D [48], while genetic deletion of FoxO1 in murine beta cells renders these cells more sensitive to metabolic stress, and is associated with upregulation of pluripotency genes such as Ngn3, Oct4, Nanog and L-Myc [49]. Similar findings were observed in islets from T2D patients, compared with healthy controls [43]. A larger proportion of T2D islets, compared with controls, showed a subpopulation of glucagon-positive cells that expressed cytoplasmic (inactive) FOXO1 and α-cell Aristaless-related homeobox transcription factor (ARX) [50], and a subpopulation of somatostatin-positive cells expressed cytoplasmic (inactive) homeobox protein NKX6.1 [43], suggesting dedifferentiation of beta cells and transition towards α- or δ-like cell physiology. Expression of the progenitor cell marker aldehyde dehydrogenase 1 family, member A3 (ALDH1A3) [51], is also observed in islets from T2D patients [43].
2. Lipid Accumulation and Beta Cell Dysfunction
It is established that over-accumulation of lipids and associated over-activation of lipid signalling pathways (lipotoxicity) contribute to loss of insulin secretion, beta cell toxicity and dysfunction, providing a link between obesity and T2D (reviewed in [41]). Glucolipotoxicity (GLT) describes the synergistic damaging effects of increased concentrations of free fatty acids (FFA) in the presence of high glucose concentrations (reviewed in [42]). Multiple outcomes are triggered in beta cells by GLT, including mitochondrial dysfunction and oxidative stress, endoplasmic reticulum (ER) stress and the protein unfolding response, inflammation and impaired autophagy, and loss of GSIS [42]. These changes reflect altered cell signalling pathways, increased expression of inflammatory cytokines, lipogenic and pro-apoptotic genes and proteins, and the accumulation of lipids, including diacylglycerols and triacylglycerols, ceramides, cholesterol and cholesteryl esters [42,52].
2.1. Fatty Acids, Diacylglycerols, Triacylglycerols and Beta Cell Dysfunction
The biosynthesis of triacylglycerol droplets, via intermediate diacylglycerols, is an important feature in many cell types, storing excess caloric intake against future need, and preventing the build-up of potentially toxic fatty acid derivatives [53]. Triacyglycerol synthesis occurs at the endoplasmic reticulum (ER), primarily from glycerophosphate and fatty acyl CoA. Diacylglycerol acyltransferase-1 (DGAT-1) plays an important role in esterifying (and thereby detoxifying) excess lipids entering the cell, while DGAT-2 esterifies fatty acids arising via de novo lipogenesis from glucose [53].
Fatty acid signalling plays an established (nutritional) role in stimulating insulin secretion by beta cells [54]: Jezek et al. (2018) recently reviewed the physiological roles of fatty acids in amplifying GSIS, inducing insulin granule exocytosis, and interacting with free fatty acid (FA) receptors [55]. The majority of in vitro studies examining the pathological impact of saturated FFA, such as palmitate and stearate, do so in the context of high glucose (GLT), in order to replicate diabetic conditions, although it is a difficult task to define the concentrations to which islets are exposed in vivo, as these depend on circulating levels, and factors influencing both delivery, uptake and release of FFA by islet cells [42]. Certainly, saturated fatty acids, in the presence of glucose, reduce insulin transcription by decreasing the expression of the transcription factor MafA, and translocation of pancreatic and duodenal homeobox 1 (PDX1), but these findings cannot be dissociated from the impact of GLT conditions on generation of ceramide (below) [42]. The molecular mechanisms by which palmitic acid induces apoptosis in beta cells are not completely understood, but may involve activation of kinases, including c-Jun N-terminal kinase (JNK), protein kinase C (PCK), p38 mitogen-activated protein kinase (p38MAPK), extracellular signal-regulated kinase (ERK) and Akt kinase pathways [56].
In contrast to saturated palmitic (C16:0) or stearic acids (C18:0), monounsaturated oleic acid (C18:1) is thought to improve beta cell survival and prevent loss of insulin signalling [57]; Cho et al. (2012) also demonstrated that arachidonic acid can protect against the damaging effects of palmitic acid in HIT-T15 pancreatic cells (loss of GSIS, DNA fragmentation and decreased cell viability) [58]. Notably, this protective mechanism was lost in the presence of a DGAT inhibitor, suggesting that the presence of the polyunsaturated fatty promoted sequestration of toxic palmitic acid into triacylglycerol [58]. Knockdown of fatty acid synthase (FAS), which decreases phospholipid and neutral lipid pools in INS-1 832.13 insulinoma cells, inhibits GSIS, suggesting that efficient storage of newly synthesised lipids is also important in sustaining insulin secretion [59].
Diacylglycerol, as a lipid signal messenger, has a physiological role in beta cells: its primary function is to activate protein kinase C (PKC0, but also triggers other pathways, such as the Munc-13-dependent pathway: the cellular level of diacylglycerol (DAG), which is tightly regulated by DAG kinases (DGK), acts as a positive regulator of insulin secretion [60]. However, diacylglycerol has also been shown to inhibit insulin release via a PKC-independent mechanism in HIT T-15 islet cells, via modulation of Ca2+ flux [61,62]. Sawatini et al. (2019) demonstrated a biphasic response to type I DGK inhibitor, R59949, in MIN6 β cells: while low concentrations of the type I DGK inhibitor, R59949, increase PKC-dependent insulin secretion, higher concentrations (>10 μM), which trigger higher levels of diacylglycerol, suppress this process, possibly via loss of voltage-dependent Ca2+ channel activity [63,64]. Esterification of both fatty acids and diacylglycerol into the relatively inert triacylglycerol pool protect against the accumulation of bioactive (and potentially toxic lipids) (above) [53,54,55]. Exposure of rat islets to elevated levels of glucose stimulates the formation of glycerol and fatty acids, and diversion of glucose carbons into triacylglcyerols and cholesteryl esters [65]. By contrast, elevated plasma concentrations of triacylglycerol-rich lipoproteins reflect increased fatty acid flux from adipose tissue, and are linked with diminution of insulin secretion and induction of insulin resistance in patient cohorts [66].
2.2. Ceramides and Sphingolipid Signalling in Beta Cell Dysfunction
A series of complex interactions, requiring both active synthesis and degradation, determine the cellular sphingolipid content (reviewed in [67]). Biosynthesis is initiated at the cytosolic face of the endoplasmic reticulum (ER), starting with the condensation of L-serine and palmitoyl CoA; reduction, acetylation and desaturation reactions result in the generation of ceramide, which acts as the central substrate for the production of other sphingolipid intermediates (reviewed in [68]). Hydrolysis of ceramide at the ER (neutral ceramidase), plasma membrane (alkaline ceramidase) and in the lysosome (acid ceramidase) generate sphingosine, which can be phosphorylated to sphingosine-1-phosphate by sphingosine kinase. Ceramide is transported from the ER to the Golgi, where is can be used to synthesise sphingomyelin and glucosylceramides; at the plasma membrane, ceramide kinase generates ceramide-1-phosphate (C1P), which can be hydrolysed back to ceramide by C1P phosphatase [68].
Dysregulated ceramide and sphingolipid metabolism has been linked with dysregulation of insulin secretion, and apoptosis of beta cells, in response to glucolipotoxicity and/or inflammatory cytokines. Veluthakal et al. (2009) demonstrated that the impact of palmitic acid under glucolipotoxic conditions can be mimicked by a cell-permeable ceramide analog which reduces the expression of nucleotide diphosphate kinase in INS832/13 cells [69], a feature which may contribute to abnormal G protein activation and impaired insulin secretion. Indeed, recent evidence implicates cross-talk between Ras-related C3 botulinum toxin substrate 1 (Rac1) and the ceramide signalling pathway in the onset of beta cell dysfunction [70]. Exposure to palmitic acid, in the presence of glucose, also impairs transcription of the insulin gene in MIN-6 cells, via activation of Per-Arnt-Sim kinase (PASK) and extracellular regulated kinases-1/2 (ERK1/2) [71].
Incubation with 0.4 mM palmitic acid under normoglycaemic conditions increases de novo synthesis of dihydrosphingosine and dihydroceramides in beta cells without inducing apoptosis; however, increasing the glucose concentration to 30 mM induced apoptosis, and amplified formation of C18:0, C22:0 and C24:1 (dihydro)ceramide species via upregulation of ceramide synthase 4 levels [72]. Activation of the extrinsic apoptotic pathway under glucolipotoxic conditions, mostly via initiator caspase 8, promotes apoptosis by cleavage and activation of downstream effector caspases like caspase 3 (reviewed in [73]). The lack of caspase 8 can protect against ceramide-induced beta cell death, and knockout of caspase-3 can protect mice against the development of autoimmune diabetes [74,75]. Other sphingolipid metabolites, including glycosphingolipids, sphingosine-1-phosphate and gangliosides, can affect beta cell signalling pathways, including apoptosis, cytokine release, ER to Golgi vesicular trafficking and insulin gene expression; the activity of neutral sphingomyelinases, which regulate the composition of the plasma membrane, can also affect beta cell excitability and insulin [76]).
2.3. Cholesterol Accumulation and Beta Cell Dysfunction
Effective cholesterol homeostasis in beta cells is an important factor in maintaining insulin secretion (reviewed in [77]). The uptake, synthesis and removal of cholesterol is tightly controlled by the functional opposition between the activities of sterol regulatory element-binding proteins (SREBPs) and liver X receptor (LXR α/β) transcription factors, while storage is facilitated by esterification to cytosolic droplets of cholesterol esters by acyl CoA: cholesterol acyl transferase (ACAT-1). The primary route for cholesterol uptake is via members of the low-density lipoprotein receptor (LDL-R) [78] and scavenger receptor families [79]. As the intracellular cholesterol content rises, SREBP-2 is sequestered (and inactive) at the endoplasmic reticulum, leading to loss of expression of genes encoding the LDL-R and the enzymes responsible for endogenous synthesis of cholesterol [80]. Instead, (oxy)sterol-mediated activation of nuclear LXR transcription factors, which form obligate heterodimers with retinoid X receptors (RXR), leads to induction of expression of genes encoding proteins involved in the ‘reverse’ cholesterol transport process, including ATP-binding cassette (ABC) transporters A1 (ABCA1) and ABCG1/G4 [81,82]). These transporters work in concert to remove excess cholesterol from cells, via efflux to (apo)lipoprotein acceptors such as apoA-I and high-density lipoprotein, respectively [82].
Naturally, the presence of excess cholesterol regulates the physical properties (fluidity, curvature and lipid raft content) of membranes that influence function and locale of membrane proteins such as receptors, ion channels and transporters, and vesicle formation and fusion, affecting several steps of the insulin secretory pathway [77] (Figure 2). Notably, glucose-stimulated insulin release is reduced by decreased glucose transporter activity [83], and stabilisation of the neuronal nitric oxide synthase (nNOS) dimer, which prevents the movement of glucokinase from insulin granules to the cytosol [77]. An increase in cellular cholesterol level can also increase plasma membrane-associated phosphatidylinositol 4,5 bisphosphate (PIP2) [84]: PIP2 dissociated from the plasma membrane is hydrolysed by phospholipase C leading to Ca2+ release from intracellular stores and may sensitise KATP channels leading to an influx of Ca2+ by CaV channels [85,86]. Alterations in the density of voltage-gated Ca2+ channels lead to reduced flux of Ca2+ into the beta cell, and decreased insulin secretion [87]. In addition, increased production of PIP2 activates dynamin, which acts to reduce full fusion events of granules at the plasma membrane [88,89], while accumulation of excess cholesterol in insulin granules causes dysfunctional retrieval of exocytosis proteins, such as clathrin, syntaxin 6 and vesicle-associated membrane protein 4 (VAMP4) [90]. These factors contribute to incomplete granule–membrane fusion, evidenced by longer duration and reduced lateral spreading of insulin granules [91].
Accumulation of cholesterol at the endoplasmic reticulum not only depletes calcium stores needed for insulin release [92,93] but can trigger protein unfolding by activation of the protein kinase RNA-like endoplasmic reticulum kinase (PERK)–phosphorylated eukaryotic initiation factor 2 alpha (eIF2α) [94] pathway, which results in global inhibition of protein synthesis (including preproinsulin) and translation of activating transcription factor 4 (ATF4) [95]. Build-up of sterol within the trans-Golgi network inhibits granule formation [90] while disruption of lipid rafts alters the sorting of granins, a key constituent of secretory granules, and of endoproteases, needed for the processing and maturation of the insulin hormone [96].
Conversely, reductions in cholesterol biosynthesis caused by ‘statin’ drugs, or the depletion of the plasma membrane cholesterol pool using methyl β-cyclodextrin (MCD), also inhibits GSIS and lowers insulin content in β cells and islets [97,98]. Depletion of cholesterol also affects the formation of insulin granules, while disruption of cholesterol-rich lipid rafts impairs insulin secretion by redistribution of SNARE (syntaxin and SNAP25) and K+ATP and voltage-gated Ca2+ channels [97,98]. High levels of glucose inhibit cholesterol biosynthesis, resulting in disruption of lipid rafts, redistribution of plasma membrane syntaxin 1A, loss of this protein from granule-docking sites, fewer docked granules and reduced insulin secretion [99]. Moreover, recent studies have suggested that use of statin drugs in dyslipidaemia can actually provoke new-onset diabetes in ‘prediabetic’ patients [77], and genetic variants in HMGCR have also been linked with predisposition to diabetes, again positing cholesterol biosynthesis as important in sustaining beta cell function [100].
3. Mechanisms Contributing to Changes in Gene Expression and Beta Cell (dys)function: microRNA
Over the last decade, mechanisms resulting in changes in gene expression, including chromatin modifications, DNA methylation, post-translational modifications of histones, and altered expression of non-coding RNA sequences, such as long non-coding RNA (lncRNA) and microRNA (miRNA/miR) have been implicated in regulation, and loss, of beta cell function and diabetes: a number of excellent reviews have recently covered these areas in depth [101,102,103,104,105,106].
MicroRNA sequences are small (~22 nucleotide) non-coding RNA sequences which regulate the expression of networks of genes in beta cells, in response to environmental factors such as caloric excess, obesity and diabetes [101,102,103,104,105,106]. These sequences can be isolated or clustered within the human genome, either between genes or within the intron–exon regions of genes encoding proteins [107,108]. Transcription of microRNA (miR) sequences is dependent on the expression and activity of RNA polymerase II/III [109,110], can be dependent or independent of mRNA expression [108,111,112] and occur via both canonical and non-canonical pathways [109,113]. In the canonical pathway, a hairpin-containing primary miRNA (pri-miRNA) transcript with a 5’-methylated cap and a 3’-polyadenylated tail is generated, which is then processed via a complex containing double-stranded RNA-binding protein DiGeorge syndrome critical region gene 8 (DCGR8) which recognises methyl motifs present in the pri-miRNA [114,115,116]. This interaction anchors Drosha, a ribonuclease III which generates precursor miRNA (pre-miRNA) by cleaving the hairpin structure from the pri-miRNA transcript [117,118,119]. The pre-miRNA (~70 nucleotides) are exported from the nucleus: exportin-5 interacts with the 3’ overhanging sequence of pre-miRNA, while RanGTP remains bound to the hairpin structure until hydrolysis of GTP to GDP in the cytosol results in release of pre-miRNA [120]. Cytosolic pre-miRNA is processed by Dicer (RNase III), which removes the stem–loop structure to generate a mature miRNA duplex (19–25 nucleotides in length) [121,122]. The guide strand is loaded onto the active RNA-induced silencing complex (RISC), made up of Dicer, TAR RNA-binding protein (TRBP) and argonaute (1–4) proteins; miRNA base pairs with their complementary mRNA molecules are guided by their miRNA recognition element [123,124].
A perfect (exact) or near-perfect complementary match between miRNA and the conserved 3’-UTR region of the target mRNA results in degradation of mRNA; if the complementarity is imperfect (partial), then moderate reductions in mRNA and translational repression occur [125,126,127,128]. The end result is decreased protein output from the target gene, albeit often quite modest in its magnitude [125,126,127,128], reflecting the role of microRNA in ‘fine-tuning’ gene and protein expression. Additional factors can reduce translational efficiency or induce mRNA destabilisation, including AU-rich regions near the ‘seed’-binding sites, auxiliary binding of miRNA to the target transcript, or mRNA deadenylation [127,128]. Each miRNA sequence can have target sites in hundreds of different genes, exhibiting differing degrees of complementarity: computational prediction suggests that >60% of all protein-coding genes are miRNA targets [125,129,130]. Tissue-specific and concentration-dependent effects are also noted, particularly in healthy tissues compared with pathological conditions [131,132,133,134]. Finally, some miRNA sequences exist in the extracellular environment, in microvesicles, like exosomes and ectosomes, complexed with proteins, or transported in lipoproteins such as HDL, and have been widely employed as biomarkers of health and disease [135,136,137,138].
3.1. MicroRNA Sequences Linked with Lipid Accumulation in Beta Cells
Over the last decade, it has become clear that the network of genes encoding proteins involved in lipid metabolism and cholesterol homeostasis also lies under the control of microRNA sequences, such as miR-33 [139]. Table 1 lists some of the microRNA sequences, derived from interrogation of the NCBI/PubMed database which are altered by changes in metabolism induced in beta cells and islets. It is clear that multiple miRNA sequences are regulated in beta cells by exposure to metabolic challenges, targeting an array of genes and processes involved in beta cell function. In particular, induction of miR-34a is strongly linked with beta cell lipotoxicity associated with exposure to saturated fatty acids in vitro and in vivo, via multiple mechanisms [140,141,142,143,144] (Table 1), which may also reflect increased flux of fatty acids through the diacylglycerol/triacylglycerol, ceramide/sphingolipid and cholesterol esterification pathways. These include targeting sirtuin 1 (SIRT1), an NAD+-dependent deacetylase, which activates expression of tumour-suppressor protein p53, DNA repair factor Ku70, nuclear factor κB (NF-κB), STAT3 and the FOXO family of forkhead transcription factors [145]. Sirtuin 1 aids suppression of cellular senescence, delays age-related telomere attrition, promotes DNA damage repair and cell survival, and reduces apoptosis [145]; loss of this protein, due to elevation of miR-34a after exposure to saturated fatty acids, is therefore entirely consistent with enhanced lipotoxicity in β cells. MiR-34a also directly targets lactate hydrogenase, thereby repressing the increased glycolysis observed in proliferating cancer cells (reviewed in [146]), and targets peroxisome proliferator activator receptor α (PPARα) in liver cells [reviewed in 147], both of which may impact utilisation of fatty acids; whether these factors contribute to toxicity in beta cells remains unknown. Certainly miR-34a, itself a target of p53, is an established tumour suppressor, and repression or dysregulation of this sequence is noted in a number of human cancers, leading to the development of MRX34, a liposomal miR-34a mimic, as a putative therapeutic (discussed further below) [146,147,148].
Other microRNA sequences altered in beta cells by exposure to saturated fatty acids, and linked with lipotoxicity, include miR-146a [140,141], miR-182-5p [149], miR-297b-5p [150,151], miR-374c-5p [151], miR-375 [152] and miR-3074-5p [153] (Table 1). MicroRNA-146 exists in two forms (miR-146a/b), often not distinguished despite their distinct chromosomal locations [154], but which share a seed region and target some of same genes involved in the host immune response and inflammation, such as Toll-like receptors. The role of ‘mirR-146’ in promoting apoptosis appears context dependent: mir-146a-5p promotes the apoptosis of chrondrocytes via activation of the NF-κB pathway [155], while miR-146b enhances apoptosis of gastric cancer cells by targeting protein tyrosine phosphatase 1B (PTP1B) [156]; by contrast, ‘miR-146’ protects against cardiomyocyte apoptosis by inhibiting NF-κB [157], and blocks the pro-apoptotic and inflammatory effects of lipopolysaccharide (LPS) in lung cancer cell lines [158]. Fred et al. (2010) also demonstrated that in human islets, the level of ‘miR-146’ increases after exposure to pro-inflammatory cytokines, decreases after culture in media containing high glucose, but was not changed by exposure to palmitate [159].
Notably, both miR-146b and miR-182-5p have been linked with protection against high-fat diet-induced non-alcoholic steatohepatitis in mice: exposure to miR-146b reduces the expression of IL-1 receptor-associated kinase (IRAK1) and tumour necrosis factor (TNF) receptor-associated factor 6 (TRAF6) after exposure to oleic acid, reducing inflammation and lipid accumulation in vitro and in vivo [160]. In the same models, miR-182-5p reduced oleic acid-induced hepatic expression of TNFα, IL-6 and TLR4 [161]; this sequence also prevents apoptosis, and reduces the levels of cluster of differentiation (CD) 36, total cholesterol and triglyceride in macrophages after exposure to oxidised LDL, again by targeting TLR4 [162]. In β cells, miR-182-5p directly targets thrombospondin-1 (TSP-1) [149], a CD36 ligand, which, in human hepatic cells, regulates lipid metabolism by inhibiting the proteolytic cleavage of SREBP-1, reducing lipogenesis and triglyceride accumulation [163]. However, genetic deletion of TSP-1 in mice is associated with reduced plasma lipid levels and hepatic inflammation, and activation of PPARα [164], and decreased obesity-induced microvascular complications in apoE−/− mice [165], findings which resonate with the impact of miR-182-5p in beta cells (Table 1).
One other study remarks the impact of miR-374c-5p on apoptosis, controlling the proliferation, migration, epithelial-mesenchymal transition and apoptosis of human breast cancer cells, via repression of TATA box-binding protein associated 7 (TAF7) and expression of DEP domain containing 1 (DEPDC1), a transcriptional co-repressor involved in the promotion of carcinogenesis [166]. The tumour suppressive sequence miR-3074-5p has also been linked with increased apoptosis in both trophoblasts and breast cancer [167,168], but no direct reports link this sequence with altered lipid metabolism. By contrast, a number of recent studies cite both positive and negatives roles for miR-375 in apoptosis, of chrondocytes, breast, colon, gastric and hepatic cancer cells and cardiomyocytes [169,170,171,172,173,174,175,176]. In mice (C57BL/6), miR-375 blocks high-fat diet-induced insulin resistance and obesity, by inhibiting over-activation of the aryl hydrocarbon receptor and promoting hepatic expression of genes involved in responses to insulin [175], providing protection against the high-fat diet. Notably, this sequence is thought to play an established role in beta cell function and in diabetes: (reviewed in [176]). During the development of the pancreas, the increased expression of miR-375 parallels increased expression of the insulin gene, and proliferation of β cells, while loss-of-function (LOF) knockdown of this sequence in zebrafish and mice suggests a key role in determining the balance between β cells (↓) and α cells (↑). MicroRNA-375 is thought to inhibit GSIS via a number of mechanisms, including targeting myotrophin (Table 1) [152], pyruvate dehydrogenase kinase-1 (PDK1), PI3-kinase and interactions with cAMP-directed pathways [176].
Finally, exposure to saturated fatty acids lowers the expression of miR-297-5p in vitro and ex vivo [150,151], which targets large-tumour-suppressor kinase 2 (LATS-2) [150]. This kinase is a central regulator of cell fate, influencing the function of a host of oncogenic or tumour suppression factors, and is a core component of the canonical Hippo pathway [177], so increased expression of LATS-2 is consistent with the lipid-induced apoptosis seen in beta cells (Table 1). Notably, LATS-2 inhibits the activation of SREBP-2, and suppresses cholesterol accumulation in hepatic cells [178]: if replicated in beta cells, enhanced LATS-2 expression could therefore additionally promote cholesterol deposition. MiR-7222-3p, which is also elevated by exposure to palmitate, targets ACAT-1: loss of this protein would enhance the level of potentially toxic-free cholesterol in beta cells, by abrogating storage as neutral cytosolic droplets of cholesteryl ester [179].
Of the sequences directly moderated by cholesterol exposure in beta cells and islets, elevations in two (miR-27a and miR-33a) [180,181] are linked with repression of ABCA1; it is well established that this cholesterol efflux transporter, its apolipoprotein acceptors (e.g., apoA-I, apoE) and its product, HDL itself, can provide protection to beta cells and pancreatic islets [182,183,184,185,186,187,188] and in experimental models [187,188]. Some, but not all, clinical studies also provide support for this concept [189,190,191]. These protective functions have been linked with the removal of excess cholesterol from beta cells, while others cite sterol- and/or transporter independent effects of apoA-I and HDL [192,193,194,195,196,197]. Certainly, miR-33a is one of the most intensively studied miR sequences in lipid metabolism and is integral to these responses [182,198,199]. Mir-33a is encoded in an intronic region of SREBF2 and thus forms a regulatory link between the active expression of this gene, and repression of ABCA1 and ABCG1: knockout of miR-33a in mice promotes cholesterol efflux via ABCA1 and ABCG1, increases circulating levels of HDL and hepatic excretion of cholesterol in bile (reviewed in [197]), and this sequence is currently under exploration as a possible clinical target [198,199]. Notably, ABCA1 is also regulated by exposure to elevated levels of glucose: miR-145 increases the total level of islet cholesterol [200], while miR-383 targets the anti-inflammatory Toll-like receptor 4 [201], which has also been linked with altered expression of ABCA1 [182].
More complex relationships between miRs and HDL emerge from Table 1: Tarlton et al. (2021) showed that miR-21-5p could mimic the effects of HDL on targets STAT3 and SMAD7, but could not provide equivalent protection against glucolipotoxicity in human PANC hybrid 1.1B4 cells [202], while HDL increases the export of miR-375-3p, a feature which correlates inversely with insulin secretion in murine MIN6 cells [203]. Cholesterol exposure also enhances the expression of miR-24a, which impacts on the transcription factor Sp1 to alter the expression of secretagogin and reduce insulin secretion [204]. This sequence has been closely linked with changes in lipid metabolism in other cells and tissues: obesity induces overexpression of miR-24 [205], a sequence which associates with HDL, and that regulates cholesterol uptake by targeting scavenger receptor (SR) B-1. The same sequence enhances atherosclerosis by reducing lipid uptake from HDL via repression of SR-B1 [206] and, intriguingly, can control triacylglycerol biosynthesis by targeting fatty acid synthase [207]; inhibition of miR-24 can also help to limit hepatic lipid accumulation and hyperlipidaemia [208], suggesting this sequence may integrate neutral lipid metabolism in beta cells.
3.2. MicroRNA Biomarkers: Associations with Changes in Lipid Metabolism in Beta Cells
Some of the sequences altered by metabolic challenges in beta cells have also emerged as biomarkers of diabetes in the circulation (Table 2), although the epigenomic landscape in the bloodstream, as in cells and tissues, is obviously much more complex. Approximately 10% of miRNAs are thought to be secreted encapsulated in exosomes, with the remainder stably complexed with proteins such as argonaute 2 and nucleophosmin 1, and with HDL, under vesicle-free conditions; all of these forms protect miRNA against RNase degradation, allowing their delivery to recipient cells and tissues, and promoting intercellular communication [209]. MicroRNA sequences are thought to be selectively secreted into extracellular vesicles, just as the proteomic profile of secreted exosomes differs from parental cells [210], although routine analysis of the RNA content of extracellular vesicles in liquid biopsies is a challenging proposition [211]. Intriguingly, inhibition of neutral sphingomyelinase-2, which is rate limiting for synthesis of ceramides (above) decreases the amount of miRNA in exosomes (but not parent cells) [212]; ceramide synthesis is also thought to be involved in the functionally distinct, and possibly opposing, pathway mediated by HDL [212].
The data in Table 2 describe the outcome of searches for circulating miRNA sequences in studies relating to diabetic patients [213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271]; of these, 21 studies identify at least one of the sequences described in Table 1. It is clear that the directions of change of such biomarkers are not always consistent between differing studies (Table 2) or indeed, when comparing biomarker studies with outcomes from cells and tissues (Table 1 vs. Table 2). The sequences identified as biomarkers in Table 2, which have also been linked with changes in lipid metabolism in beta cells and islets, include miR-21 [216,244,256,257,261,270], miR-24 [216,229,230,257,261], miR-27a [218], miR-34a [213,229,261,270], miR-145 [247], miR-146a [218,219,228,239,241,244,245,265,266], miR-182 [210,218,237] and miR-375 [217,218,228,243,253]. Hsa-miR-146a emerges as one of the most common regulated sequences linked with diabetes, with decreased levels seen in samples derived from the blood of patients with diabetes [218,219,241,245,265,266] and those with diabetic foot and nephropathy [228], although Mensa et al. (2019) observed increased levels seen in diabetic women, compared with diabetic men [219]. Elevated levels of this sequence were also found in gingival crevicular fluid [239] and corneal samples [244] of type 2 diabetic patients, while in murine beta cells, and db/db islets, levels of miR-146a are increased in response to exposure to palmitate [140], predicating apoptosis (Table 1). Levels of hsa-miR-34 decreased in the bloodstream in two studies of diabetic patients [213,229], but increased in the report from Seyhan et al. (2016) [270]; levels also increased in murine cells exposed to palmitate [141,142,143,144]. These differing outcomes may reflect the progression of the disease, or the selective retention of this sequence under pro-apoptotic conditions. Clearly, there are challenges remaining in relating complex outcomes in cells and tissues with the epigenetic profile found in fluid biopsies.
3.3. Predictive Analysis (DIANA/KEGG) of Pathways Implicated in Beta Cell Dysfunction in the Face of Metabolic Challenges
Bioinformatic analysis was carried out on microRNA sequences identified as associated with lipid metabolism in pancreatic beta cells using miRPath v3.0 [272]. Full details of the list of miRs can be found in the Mendeley dataset deposit (doi: 10.17632/jnz8h974gc.1). In brief, all miRNAs named in Table 1 were used, sequences described in Table 1 were verified in miRbase before inclusion: where 5p or 3p sequences were not specified, both were included in the search list (full search list: hsa-miR-21-5p, hsa-miR-24-1-5p, hsa-miR-24-2 5p, hsa-miR-27a-3p, hsa-miR-33a-3p, hsa-miR-33a-5p, hsa-miR-34a-3p, hsa-miR-34a-5p, hsa-miR-124-3p, hsa-miR-124-5p, hsa-miR-145-3p, hsa-miR-145-5p, hsa-miR-146a-3p, hsa-miR-146a-5p, hsa-miR-182-5p, hsa-miR-297-5p, hsa-miR-374c-5p, hsa-miR-375, hsa-miR-383-3p, hsa-miR-383-5p and hsa-miR-3074-5p). Figure 3A demonstrates the Kyoto Encyclopaedia of Genes and Genomes (KEGG) pathways with a p < 0.01 association with the miRNA sequence involved in lipid metabolism in pancreatic beta cells. This recognises several lipid pathways (fatty acid biosynthesis, fatty acid metabolism, fatty acid elongation, glycosphingolipid biosynthesis and biosynthesis of unsaturated fatty acids) which validates the principle underlying the search. The additional pathways are indicative of the pleiotropic nature of miRNAs which have multiple gene targets, and highlight the importance of lipid homeostasis to beta cell function and health, with pathways implicated that affect pancreatic islet architecture and morphology (ECM-receptor interactions, Hippo signalling pathway, adherens junctions, thyroid hormone signalling pathway), mitosis (cell cycle), cellular energy homeostasis (FoxO signalling pathway) and cell survival (p53 signalling pathway); miRNA-pathway interactions in Figure 3B were mapped in Cytoscape v3.8.0 [273].
3.4. Therapeutic Applications of microRNA (targets) in Beta Cells
MicroRNA pathways can be regulated pharmacologically, and treatments involving miRNA focus on influencing dysregulated levels of miRNA in disease, including suppression or enhancement of key sequences [133]. Gene silencing can be achieved using artificial, double-stranded RNA fragments (mimics) that bind to target mRNA, resulting in activation of the RISC complex, downregulation of specific mRNA, and gene suppression. Synthetic oligonucleotides can bind to mature miRNA targets, leading to reduced RISC activity and upregulation of specific mRNA and protein expression; target ‘mimicry’ can also employ miRNA sponges, masking or erasers [133].
The practical utility of these approaches is exemplified by Miraversen (miR-122) which effectively reduced the expression of hepatitis C virus in Phase II clinical trials (2017) without significant side-effects [274,275,276], and by MRX34, a lipsomal miR-34a mimic, which entered Phase I clinical trials for treatment of advanced liver cancer [277,278]. However, the trial of the latter was halted by the Food and Drug Administration (FDA) in 2016, due to severe immune-mediated toxicity and four patient deaths (reviewed in [141]). Contributing factors may include the packaging vehicle, which was not designed to specifically target the miRNA to cancer cells, or the dose or dose schedule: at present, the reasons for the immune-related adverse events are not understood, and were not predicted by preclinical studies in animals, including non-human primates [141].
The packaging vehicle is key to efficient gene regulation as they facilitate passage through many physiological barriers before reaching the target tissue; options include metal, polymer or lipid nanoparticles, liposomes and hydrogels [279]. Packaging vehicles can also be designed to improve delivery of miRNAs to target tissues: some examples include the modification of polyamidoamine (PAMAM) nanocarriers with folic acid to direct them to cancers that overexpress folate receptors, and an amino acid sequence on cationic liposomes which increased delivery of siRNA to osteogenic cells [280,281]. Ensuring a targeted approach is important to limit off target effects. For example, one plausible target to protect beta cell survival is miR-34a (Table 1); however, this sequence is also a tumour suppressor. Thus, any therapeutic based on targeting miR-34a would require a delivery system directed specifically to beta cells, to avoid global targeting that could lead to uncontrolled growth in healthy tissue leading to cancer [282,283,284]. Several therapeutics are currently being developed that target miRNAs associated with lipid metabolism in beta cells (Table 1) including miR-21, miR-145 and miR-146a; however, none of them are designed to deliver specifically to the pancreas, or indeed, as T2D treatments [285,286,287]. Delivery of miR-216a to the pancreas using nanoparticles has been achieved in vivo [288]; while the nanoparticles were not designed to target the pancreas and improve delivery, the study demonstrates that nanoparticles can enter the pancreas and accumulate therein.
Thus, a number of factors must be considered in developing miRNA-based therapeutics, not least the number of target genes and cell signalling networks affected by these sequences [289], but also effective (tissue-specific) vectors and delivery processes [279,280,281,282,283,284]. Consideration of miRNA networks may also be useful when examining how to make effective therapies from miRNAs [202,289]. Designing treatments that comprehensively alter miRNA networks may achieve improved outcomes while retaining specificity by targeting a specific network. Another issue is that beta cells can export miRNAs which can then modulate gene expression in recipient tissues, which may complicate attempts to limit effects to the pancreas [138,203]. Careful scrutiny of these factors may facilitate the development of new drugs that can provide new treatment options for T2D patients that are more specific and safer than currently available options.
4. Conclusions
The explosion of interest in factors regulating gene expression, and beta cell function, over the last decade has revealed networks of genes regulated by multiple microRNA sequences, and the discovery of new pathways contributing to type 2 diabetes. This review has focused on miRNA sequences which are altered by changes in lipid metabolism in beta cells and islets, and highlighted the pleiotropic roles of these sequences in protecting against apoptosis or exacerbating lipid accumulation in these cells and tissues. Ultimately, the development of research in this field may lead to RNA-based therapeutics capable of sustaining beta cell function and preventing progression to type 2 diabetes.
Author Contributions
Conceptualization, J.M.R.T., S.P. and A.G.; writing—review and editing, J.M.R.T., S.P. and A.G.; supervision, A.G. and S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in thus study are openly available in Mendeley Data at doi:10.17632/jnz8h974gc.1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IDF Diabetes Atlas. Available online: https://diabetesatlas.org/upload/resources/material/20200302_133351_IDFATLAS9e-final-web.pdf (accessed on 11 June 2021).
- Van Haeften, T.; Pimenta, W.; Mitrakou, A.; Korytkowski, M.; Jenssen, T.; Yki-Järvinen, H.; Gerich, J. Relative contributions of β-cell function and tissue insulin sensitivity to fasting and postglucose-load glycemia. Metabolism 2000, 49, 1318–1325. [Google Scholar] [CrossRef]
- Mezza, T.; Cinti, F.; Cefalo, C.M.A.; Pontecorvi, A.; Kulkarni, R.N.; Giaccari, A. β-Cell Fate in Human Insulin Resistance and Type 2 Diabetes: A Perspective on Islet Plasticity. Diabetes 2019, 68, 1121–1129. [Google Scholar] [CrossRef]
- McCulloch, L.J.; van de Bunt, M.; Braun, M.; Frayn, K.N.; Clark, A.; Gloyn, A.L. GLUT2 (SLC2A2) is not the principal glucose transporter in human pancreatic beta cells: Implications for understanding genetic association signals at this locus. Mol. Genet. Metab. 2011, 104, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Matschinsky, F.M. Regulation of pancreatic β-cell glucokinase: From basics to therapeutics. Diabetes 2002, 51, S394–S404. [Google Scholar] [CrossRef] [Green Version]
- Lajoix, A.-D.; Reggio, H.; Chardès, T.; Péraldi-Roux, S.; Tribillac, F.; Roye, M.; Dietz, S.; Broca, C.; Manteghetti, M.; Ribes, G.; et al. A Neuronal Isoform of Nitric Oxide Synthase Expressed in Pancreatic β-Cells Controls Insulin Secretion. Diabetes 2001, 50, 1311–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, M.A.; Piston, D.W. Regulation of β cell glucokinase by S-nitrosylation and association with nitric oxide synthase. J. Cell Biol. 2003, 161, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, M.; Head, W.S.; Gunawardana, S.C.; Hasty, A.; Piston, D.W. Direct Effect of Cholesterol on Insulin Secretion: A Novel Mechanism for Pancreatic β-Cell Dysfunction. Diabetes 2007, 56, 2328–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Hatakeyama, H.; Okado, H.; Miwa, A.; Kishimoto, T.; Kojima, T.; Abe, T.; Kasai, H. Sequential exocytosis of insulin granules is associated with redistribution of SNAP25. J. Cell Biol. 2004, 165, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Nevins, A.K.; Thurmond, D.C. A Direct Interaction between Cdc42 and Vesicle-associated Membrane Protein 2 Regulates SNARE-dependent Insulin Exocytosis. J. Biol. Chem. 2005, 280, 1944–1952. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Qin, T.; Xie, L.; Dolai, S.; Zhu, D.; Prentice, K.J.; Wheeler, M.; Kang, Y.; Osborne, L.; Gaisano, H.Y. New Roles of Syntaxin-1A in Insulin Granule Exocytosis and Replenishment. J. Biol. Chem. 2017, 292, 2203–2216. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Figueroa, D.J.; Austin, C.P.; Liu, Y.; Bugianesi, R.M.; Slaughter, R.S.; Kaczorowski, G.J.; Kohler, M.G. Expression of voltage-gated potassium channels in human and rhesus pancreatic islets. Diabetes 2004, 53, 597–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, B.B.; Rhodes, C.J.; Grimsby, J.S. The dynamic plasticity of insulin production in β-cells. Mol. Metab. 2017, 6, 958–973. [Google Scholar] [CrossRef]
- Marchetti, P.; Suleiman, M.; De Luca, C.; Baronti, W.; Bosi, E.; Tesi, M.; Marselli, L. A direct look at the dysfunction and pathology of the β cells in human type 2 diabetes. Semin. Cell Dev. Biol. 2020, 103, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Han, X. Death versus dedifferentiation: The molecular bases of beta cell mass reduction in type 2 diabetes. Semin. Cell Dev. Biol. 2020, 103, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Alvarez, J.; Conget, I.; Rasschaert, J.; Sener, A.; Gomis, R.; Malaisse, W.J. Enzymatic, metabolic and secretory patterns in human islets of Type 2 (non-insulin-dependent) diabetic patients. Diabetologica 1994, 37, 177–181. [Google Scholar] [CrossRef]
- Deng, S.; Vatamaniuk, M.; Huang, X.; Doliba, N.; Lian, M.-M.; Frank, A.; Velidedeoglu, E.; Desai, N.M.; Koeberlein, B.; Wolf, B.; et al. Structural and Functional Abnormalities in the Islets Isolated from Type 2 Diabetic Subjects. Diabetes 2004, 53, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Guerra, S.; Lupi, R.; Marselli, L.; Masini, M.; Bugliani, M.; Sbrana, S.; Torri, S.; Pollera, M.; Boggi, U.; Mosca, F.; et al. Functional and molecular defects of pancreatic islets in human type 2 diabetes. Diabetes 2005, 54, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Lyon, J.; Fox, J.E.M.; Spigelman, A.F.; Kim, R.; Smith, N.; O’Gorman, D.; Kin, T.; Shapiro, A.M.J.; Rajotte, R.V.; MacDonald, P. Research-Focused Isolation of Human Islets from Donors with and without Diabetes at the Alberta Diabetes Institute IsletCore. Endocrinology 2016, 157, 560–569. [Google Scholar] [CrossRef] [Green Version]
- Solimena, M.; Schulte, A.M.; Marselli, L.; Ehehalt, F.; Richter, D.; Kleeberg, M.; Mziaut, H.; Knoch, K.-P.; Parnis, J.; Bugliani, M.; et al. Systems biology of the IMIDIA biobank from organ donors and pancreatectomised patients defines a novel transcriptomic signature of islets from individuals with type 2 diabetes. Diabetologia 2018, 61, 641–657. [Google Scholar] [CrossRef] [Green Version]
- Gunton, J.E.; Kulkarni, R.N.; Yim, S.; Okada, T.; Hawthorne, W.J.; Tseng, Y.-H.; Roberson, R.S.; Ricordi, C.; O’Connell, P.J.; Gonzalez, F.J.; et al. Loss of ARNT/HIF1β Mediates Altered Gene Expression and Pancreatic-Islet Dysfunction in Human Type 2 Diabetes. Cell 2005, 122, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Marselli, L.; Thorne, J.; Dahiya, S.; Sgroi, D.C.; Sharma, A.; Bonner-Weir, S.; Marchetti, P.; Weir, G.C. Gene Expression Profiles of Beta-Cell Enriched Tissue Obtained by Laser Capture Microdissection from Subjects with Type 2 Diabetes. PLoS ONE 2010, 5, e11499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marselli, L.; Piron, A.; Suleiman, M.; Colli, M.L.; Yi, X.; Khamis, A.; Carrat, G.R.; Rutter, G.A.; Bugliani, M.; Giusti, L.; et al. Persistent or Transient Human β Cell Dysfunction Induced by Metabolic Stress: Specific Signatures and Shared Gene Expression with Type 2 Diabetes. Cell Rep. 2020, 33, 108466. [Google Scholar] [CrossRef] [PubMed]
- Taneera, J.; Lang, S.; Sharma, A.; Fadista, J.; Zhou, Y.; Ahlqvist, E.; Jonsson, A.; Lyssenko, V.; Vikman, P.; Hansson, O.; et al. A Systems Genetics Approach Identifies Genes and Pathways for Type 2 Diabetes in Human Islets. Cell Metab. 2012, 16, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.-M.; Andréasson, A.-C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-Cell Transcriptome Profiling of Human Pancreatic Islets in Health and Type 2 Diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottosson-Laakso, E.; Krus, U.; Storm, P.; Prasad, R.B.; Oskolkov, N.; Ahlqvist, E.; Fadista, J.; Hansson, O.; Groop, L.; Vikman, P. Glucose-Induced Changes in Gene Expression in Human Pancreatic Islets: Causes or Consequences of Chronic Hyperglycemia. Diabetes 2017, 66, 3013–3028. [Google Scholar] [CrossRef] [Green Version]
- Gerst, F.; Jaghutriz, B.A.; Staiger, H.; Schulte, A.M.; Lorza-Gil, E.; Kaiser, G.; Panse, M.; Haug, S.; Heni, M.; Schütz, M.; et al. The Expression of Aldolase B in Islets Is Negatively Associated with Insulin Secretion in Humans. J. Clin. Endocrinol. Metab. 2018, 103, 4373–4383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, M.J.; Longacre, M.J.; Langberg, E.-C.; Tibell, A.; Kendrick, M.A.; Fukao, T.; Ostenson, C.-G. Decreased levels of metabolic enzymes in pancreatic islets of patients with type 2 diabetes. Diabetologia 2009, 52, 1087–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, H.; Ling, C. Mitochondrial dysfunction in pancreatic β-cells in Type 2 Diabetes. Mol. Cell. Endocrinol. 2009, 297, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Anello, M.; Lupi, R.; Spampinato, D.; Piro, S.; Masini, M.; Boggi, U.; Del Prato, S.; Rabuazzo, A.M.; Purrello, F.; Marchetti, P. Functional and morphological alterations of mitochondria in pancreatic beta cells from type 2 diabetic patients. Diabetologia 2005, 48, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Ostenson, C.-G.; Gaisano, H.; Sheu, L.; Tibell, A.; Bartfai, T. Impaired Gene and Protein Expression of Exocytotic Soluble N-Ethylmaleimide Attachment Protein Receptor Complex Proteins in Pancreatic Islets of Type 2 Diabetic Patients. Diabetes 2006, 55, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Weir, G.C.; Bonner-Weir, S. Five stages of evolving β-cell dysfunction during progression to diabetes. Diabetes 2004, 53, S16–S21. [Google Scholar] [CrossRef] [Green Version]
- Marrif, H.I.; Al-Sunousi, S.I. Pancreatic β Cell Mass Death. Front. Pharmacol. 2016, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Cunha, D.A.; Hekerman, P.; Ladrière, L.; Bazarra-Castro, A.; Ortis, F.; Wakeham, M.C.; Moore, F.; Rasschaert, J.; Cardozo, A.K.; Bellomo, E.; et al. Initiation and execution of lipotoxic ER stress in pancreatic β-cells. J. Cell Sci. 2008, 121, 2308–2318. [Google Scholar] [CrossRef] [Green Version]
- Ghavami, S.; Hashemi, M.; Ande, S.R.; Yeganeh, B.; Xiao, W.; Eshraghi, M.; Bus, C.J.; Kadkhoda, K.; Wiechec, E.; Halayko, A.J.; et al. Apoptosis and cancer: Mutations within caspase genes. J. Med. Genet. 2009, 46, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Sampson, S.R.; Bucris, E.; Horovitz-Fried, M.; Parnas, A.; Kahana, S.; Abitbol, G.; Chetboun, M.; Rosenzweig, T.; Brodie, C.; Frankel, S. Insulin increases H2O2-induced pancreatic beta cell death. Apoptosis 2010, 15, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Bucris, E.; Beck, A.; Boura-Halfon, S.; Isaac, R.; Vinik, Y.; Rosenzweig, T.; Sampson, S.; Zick, Y. Prolonged insulin treatment sensitizes apoptosis pathways in pancreatic β cells. J. Endocrinol. 2016, 230, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Rachdaoui, N.; Polo-Parada, L.; Ismail-Beigi, F. Prolonged Exposure to Insulin Inactivates Akt and Erk1/2 and Increases Pancreatic Islet and INS1E β-Cell Apoptosis. J. Endocr. Soc. 2019, 3, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitahara, Y.; Takeuchi, M.; Miura, K.; Mine, T.; Matsui, T.; Yamagishi, S. Glyceraldehyde-derived advanced glycation end products (AGEs). A novel biomarker of postprandial hyperglycaemia in diabetic rats. Clin. Exp. Med. 2008, 8, 175–177. [Google Scholar] [CrossRef]
- Zhu, Y.; Shu, T.; Lin, Y.; Wang, H.; Yang, J.; Shi, Y.; Han, X. Inhibition of the receptor for advanced glycation endproducts (RAGE) protects pancreatic β-cells. Biochem. Biophys. Res. Commun. 2011, 404, 159–165. [Google Scholar] [CrossRef]
- Imai, Y.; Cousins, R.S.; Liu, S.; Phelps, B.M.; Promes, J.A. Connecting pancreatic islet lipid metabolism with insulin secretion and the development of type 2 diabetes. Ann. N. Y. Acad. Sci. 2019, 1461, 53–72. [Google Scholar] [CrossRef]
- Lytrivi, M.; Castell, A.-L.; Poitout, V.; Cnop, M. Recent Insights into Mechanisms of β-Cell Lipo- and Glucolipotoxicity in Type 2 Diabetes. J. Mol. Biol. 2020, 432, 1514–1534. [Google Scholar] [CrossRef]
- Cinti, F.; Bouchi, R.; Kim-Muller, J.Y.; Ohmura, Y.; Sandoval, P.R.; Masini, M.; Marselli, L.; Suleiman, M.; Ratner, L.E.; Marchetti, P.; et al. Evidence of β-Cell Dedifferentiation in Human Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.S.; Stein, R.W. Evidence for Loss in Identity, De-Differentiation, and Trans-Differentiation of Islet β-Cells in Type 2 Diabetes. Front. Genet. 2017, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Bensellam, M.; Jonas, J.-C.; Laybutt, D.R. Mechanisms of β-cell dedifferentiation in diabetes: Recent findings and future research directions. J. Endocrinol. 2018, 236, R109–R143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brereton, M.F.; Iberl, M.; Shimomura, K.; Zhang, Q.; Adriaenssens, A.E.; Proks, P.; Spiliotis, I.I.; Dace, W.; Mattis, K.K.; Ramracheya, R.; et al. Reversible changes in pancreatic islet structure and function produced by elevated blood glucose. Nat. Commun. 2014, 5, 4639. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; York, N.W.; Nichols, C.G.; Remedi, M.S. Pancreatic β Cell Dedifferentiation in Diabetes and Redifferentiation following Insulin Therapy. Cell Metab. 2014, 19, 872–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, A.N.; Ram, R.; Jiang, F.-X. RNA-Seq Analysis of Islets to Characterise the Dedifferentiation in Type 2 Diabetes Model Mice db/db. Endocr. Pathol. 2018, 29, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Talchai, C.; Xuan, S.; Lin, H.V.; Sussel, L.; Accili, D. Pancreatic β Cell Dedifferentiation as a Mechanism of Diabetic β Cell Failure. Cell 2012, 150, 1223–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spijker, H.S.; Ravelli, R.B.; Mommaas-Kienhuis, A.M.; Van Apeldoorn, A.A.; Engelse, M.A.; Zaldumbide, A.; Bonner-Weir, S.; Rabelink, T.J.; Hoeben, R.C.; Clevers, H.; et al. Conversion of Mature Human β-Cells into Glucagon-Producing α-Cells. Diabetes 2013, 62, 2471–2480. [Google Scholar] [CrossRef] [Green Version]
- Marcato, P.; Dean, C.A.; Giacomantonio, C.A.; Lee, P.W. Aldehyde dehydrogenase: Its role as a cancer stem cell marker comes down to the specific isoform. Cell Cycle 2011, 10, 1378–1384. [Google Scholar] [CrossRef]
- Stratford, S.; Hoehn, K.; Liu, F.; Summers, S.A. Regulation of Insulin Action by Ceramide. J. Biol. Chem. 2004, 279, 36608–36615. [Google Scholar] [CrossRef] [Green Version]
- Walther, T.C.; Chung, J.; Farese, R.V. Lipid Droplet Biogenesis. Annu. Rev. Cell Dev. Biol. 2017, 33, 491–510. [Google Scholar] [CrossRef] [Green Version]
- Moullé, V.S.; Ghislain, J.; Poitout, V. Nutrient regulation of pancreatic β-cell proliferation. Biochimie 2017, 143, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ježek, P.; Jabůrek, M.; Holendová, B.; Plecitá-Hlavatá, L. Fatty Acid-Stimulated Insulin Secretion vs. Lipotoxicity. Molecules 2018, 23, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šrámek, J.; Němcová-Fürstová, V.; Kovář, J. Kinase Signaling in Apoptosis Induced by Saturated Fatty Acids in Pancreatic β-Cells. Int. J. Mol. Sci. 2016, 17, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vázquez-Carrera, M. Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2018, 29, 178–190. [Google Scholar] [CrossRef]
- Cho, Y.S.; Kim, C.H.; Kim, K.Y.; Cheon, H.G. Protective effects of arachidonic acid against palmitic acid-mediated lipotoxicity in HIT-T15 cells. Mol. Cell. Biochem. 2011, 364, 19–28. [Google Scholar] [CrossRef]
- MacDonald, M.; Hasan, N.M.; Dobryzn, A.; Stoker, S.W.; Ntabmi, J.M.; Liu, X.; Sampath, H. Knockdown of pyruvate carboxylase or fatty acid synthase lowers numerous lipids and glucose-stimulated insulin in insulinoma cells. Arch. Biochem. Biophys. 2013, 532, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, Y.K.; Ishikawa, T. Diacylglycerol Signaling Pathway in Pancreatic β-Cells: An Essential Role of Diacylglycerol Kinase in the Regulation of Insulin Secretion. Biol. Pharm. Bull. 2015, 38, 669–673. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.P.; Pek, S.B. Diacylglycerol inhibits potassium-induced calcium influx and insulin release by a protein kinase-C-independent mechanism in HIT T-15 islet cells. Endocrinology 1992, 131, 1985–1992. [Google Scholar] [CrossRef]
- Thomas, T.P.; Martin, D.B.; Pek, S.B. Dioctanoylglycerol Regulation of Cytosolic Ca2+ by Protein Kinase C-Independent Mechanism in HIT T-15 Islet Cells. Diabetes 1991, 40, 621–627. [Google Scholar] [CrossRef]
- Sawatani, T.; Kaneko, Y.; Ishikawa, T. Dual effect of reduced type I diacylglycerol kinase activity on insulin secretion from MIN6 β-cells. J. Pharmacol. Sci. 2019, 140, 178–186. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kobayashi, Y.; Motoki, K.; Nakata, K.; Miyagawa, S.; Yamamoto, M.; Hayashi, D.; Shirai, Y.; Sakane, F.; Ishikawa, T. Depression of Type I Diacylglycerol Kinases in Pancreatic β-Cells from Male Mice Results in Impaired Insulin Secretion. Endocrinology 2013, 154, 4089–4098. [Google Scholar] [CrossRef] [Green Version]
- Mugabo, Y.; Zhao, S.; Lamontagne, J.; Al-Mass, A.; Peyot, M.-L.; Corkey, B.E.; Joly, E.; Madiraju, S.R.M.; Prentki, M. Metabolic fate of glucose and candidate signaling and excess-fuel detoxification pathways in pancreatic β-cells. J. Biol. Chem. 2017, 292, 7407–7422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T. Pathophysiology of Diabetic Dyslipidemia. J. Atheroscler. Thromb. 2018, 25, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, S91–S96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veluthakal, R.; Suresh, M.V.; Kowluru, A. Down-regulation of expression and function of nucleoside diphosphate kinase in insulin-secreting β-cells under in vitro conditions of glucolipotoxicity. Mol. Cell. Biochem. 2009, 329, 121–129. [Google Scholar] [CrossRef]
- Kowluru, A.; Kowluru, R.A. RACking up ceramide-induced islet β-cell dysfunction. Biochem. Pharmacol. 2018, 154, 161–169. [Google Scholar] [CrossRef]
- Fontés, G.; Semache, M.; Hagman, D.K.; Tremblay, C.; Shah, R.; Rhodes, C.J.; Rutter, J.; Poitout, V. Involvement of Per-Arnt-Sim Kinase and Extracellular-Regulated Kinases-1/2 in Palmitate Inhibition of Insulin Gene Expression in Pancreatic β-Cells. Diabetes 2009, 58, 2048–2058. [Google Scholar] [CrossRef] [Green Version]
- Véret, J.; Coant, N.; Berdyshev, E.V.; Skobeleva, A.; Therville, N.; Bailbé, D.; Gorshkova, I.; Natarajan, V.; Portha, B.; Le Stunff, H. Ceramide synthase 4 and de novo production of ceramides with specific N-acyl chain lengths are involved in glucolipotoxicity-induced apoptosis of INS-1 β-cells. Biochem. J. 2011, 438, 177–189. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Liadis, N.; Murakami, K.; Eweida, M.; Elford, A.R.; Sheu, L.; Gaisano, H.Y.; Hakem, R.; Ohashi, P.S.; Woo, M. Caspase-3-Dependent β-Cell Apoptosis in the Initiation of Autoimmune Diabetes Mellitus. Mol. Cell. Biol. 2005, 25, 3620–3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liadis, N.; Salmena, L.; Kwan, E.; Tajmir, P.; Schroer, S.A.; Radziszweska, A.; Li, X.; Sheu, L.; Eweida, M.; Xu, S.; et al. Distinct in vivo roles of caspase-8 in β-cells in physiological and diabetes models. Diabetes 2007, 56, 2302–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boslem, E.; Meikle, P.J.; Biden, T.J. Roles of ceramide and sphingolipids in pancreatic β-cell function and dysfunction. Islets 2012, 4, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perego, C.; Da Dalt, L.; Pirillo, A.; Galli, A.; Catapano, A.L.; Norata, G.D. Cholesterol metabolism, pancreatic β-cell function and diabetes. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2149–2156. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, M.; Dietrich, M.F.; Herz, J. Lipoprotein receptors—An evolutionarily ancient multifunctional receptor family. Biol. Chem. 2010, 391, 1341–1363. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Ketelhuth, D.; Wirström, T.; Ohki, T.; Forteza, M.J.; Wang, H.; Grill, V.; Wollheim, C.B.; Björklund, A. Increased uptake of oxLDL does not exert lipotoxic effects in insulin-secreting cells. J. Mol. Endocrinol. 2019, 62, 159–168. [Google Scholar] [CrossRef]
- Ishikawa, M.; Iwasaki, S.; Yatoh, S.; Kato, T.; Kumadaki, S.; Inoue, N.; Yamamoto, T.; Matsuzuka, T.; Nakagawa, Y.; Yahagi, N.; et al. Cholesterol accumulation and diabetes in pancreatic β-cell specific SREBP-2 transgenic mice: A new model for lipotoxicity. J. Lipid Res. 2008, 49, 2524–2534. [Google Scholar] [CrossRef] [Green Version]
- Yvan-Charvet, L.; Wang, N.; Tall, A.R. Role of HDL, ABCA1, and ABCG1 Transporters in Cholesterol Efflux and Immune Responses. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.-J.; Yin, K.; Fu, Y.-C.; Tang, C.-K. The Interaction of ApoA-I and ABCA1 Triggers Signal Transduction Pathways to Mediate Efflux of Cellular Lipids. Mol. Med. 2011, 18, 149–158. [Google Scholar] [CrossRef]
- Lee, A.K.; Yeung-Yam-Wah, V.; Tse, F.W.; Tse, A. Cholesterol Elevation Impairs Glucose-Stimulated Ca2+ Signaling in Mouse Pancreatic β-Cells. Endocrinology 2011, 152, 3351–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, M.; Bogan, J. Cholesterol Regulates Glucose-stimulated Insulin Secretion through Phosphatidylinositol 4,5-Bisphosphate. J. Biol. Chem. 2009, 284, 29489–29498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thore, S.; Wuttke, A.; Tengholm, A. Rapid Turnover of Phosphatidylinositol-4,5-Bisphosphate in Insulin-Secreting Cells Mediated by Ca2+ and the ATP-to-ADP Ratio. Diabetes 2007, 56, 818–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Cruz, L.; Puente, E.I.; Reyes-Vaca, A.; Arenas, I.; Garduño, J.; Bravo-Martínez, J.; Garcia, D.E. PIP2 in pancreatic β-cells regulates voltage-gated calcium channels by a voltage-independent pathway. Am. J. Physiol. Physiol. 2016, 311, C630–C640. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Choi, A.H.; Ham, M.; Kim, J.-W.; Choe, S.S.; Park, J.; Lee, G.Y.; Yoon, K.-H.; Kim, J.B. G6PD Up-Regulation Promotes Pancreatic β-Cell Dysfunction. Endocrinology 2011, 152, 793–803. [Google Scholar] [CrossRef]
- Stephens, L.; Eguinoa, A.; Erdjument-Bromage, H.; Lui, M.; Cooke, F.; Coadwell, J.; Smrcka, A.; Thelen, M.; Cadwallader, K.; Tempst, P.; et al. The Gβγ Sensitivity of a PI3K Is Dependent upon a Tightly Associated Adaptor, p101. Cell 1997, 89, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, J.K.; Rivera, V.M.; Simon, S.M. Exocytosis of Post-Golgi Vesicles Is Regulated by Components of the Endocytic Machinery. Cell 2009, 137, 1308–1319. [Google Scholar] [CrossRef] [Green Version]
- Bogan, J.; Xu, Y.; Hao, M. Cholesterol Accumulation Increases Insulin Granule Size and Impairs Membrane Trafficking. Traffic 2012, 13, 1466–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Toomre, D.K.; Bogan, J.; Hao, M. Excess cholesterol inhibits glucose-stimulated fusion pore dynamics in insulin exocytosis. J. Cell. Mol. Med. 2017, 21, 2950–2962. [Google Scholar] [CrossRef] [Green Version]
- Cnop, M.; Hannaert, J.C.; Grupping, A.Y.; Pipeleers, D.G. Low Density Lipoprotein Can Cause Death of Islet β-Cells by Its Cellular Uptake and Oxidative Modification. Endocrinology 2002, 143, 3449–3453. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Liu, J.; Hou, F.; Liu, Z.; Cao, X.; Seo, H.; Gao, B. Cholesterol induces pancreatic β cell apoptosis through oxidative stress pathway. Cell Stress Chaperones 2011, 16, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Kouroku, Y.; Fujita, E.; Tanida, I.; Ueno, T.; Isoai, A.; Kumagai, H.; Ogawa, S.; Kaufman, R.J.; Kominami, E.; Momoi, T. ER stress (PERK/eIF2α phosphorylation) mediates the polyglutamine-induced LC3 conversion, an essential step for autophagy formation. Cell Death Differ. 2007, 14, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Rozpedek, W.; Pytel, D.; Mucha, B.; Leszczynska, H.; Diehl, J.A.; Majsterek, I. The Role of the PERK/eIF2α/ATF4/CHOP Signaling Pathway in Tumor Progression During Endoplasmic Reticulum Stress. Curr. Mol. Med. 2016, 16, 533–544. [Google Scholar] [CrossRef]
- Gondré-Lewis, M.C.; Petrache, H.I.; Wassif, C.; Harries, D.; Parsegian, A.; Porter, F.D.; Loh, Y.P. Abnormal sterols in cholesterol-deficiency diseases cause secretory granule malformation and decreased membrane curvature. J. Cell Sci. 2006, 119, 1876–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, F.; Xie, L.; Mihic, A.; Gao, X.; Chen, Y.; Gaisano, H.Y.; Tsushima, R.G. Inhibition of Cholesterol Biosynthesis Impairs Insulin Secretion and Voltage-Gated Calcium Channel Function in Pancreatic β-Cells. Endocrinology 2008, 149, 5136–5145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertz, J.P.Z.; Rebelato, E.; Kassan, A.; Khalifa, A.M.; Ali, S.S.; Patel, H.H.; Abdulkader, F. Distinct pathways of cholesterol biosynthesis impact on insulin secretion. J. Endocrinol. 2014, 224, 75–84. [Google Scholar] [CrossRef]
- Somanath, S.; Barg, S.; Marshall, C.; Silwood, C.J.; Turner, M. High extracellular glucose inhibits exocytosis through disruption of syntaxin 1A-containing lipid rafts. Biochem. Biophys. Res. Commun. 2009, 389, 241–246. [Google Scholar] [CrossRef]
- Swerdlow, D.I.; Preiss, D.; Kuchenbaecker, K.B.; Holmes, M.V.; Engmann, J.E.L.; Shah, T.; Sofat, R.; Stender, S.; Johnson, P.C.D.; Scott, R.A.; et al. HMG-coenzyme A reductase inhibition, type 2 diabetes, and bodyweight: Evidence from genetic analysis and randomised trials. Lancet 2015, 385, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Astro, V.; Adamo, A. Epigenetic Control of Endocrine Pancreas Differentiation in vitro: Current Knowledge and Future Perspectives. Front. Cell Dev. Biol. 2018, 6, 141. [Google Scholar] [CrossRef]
- Campbell, S.A.; Hoffman, B.G. Chromatin Regulators in Pancreas Development and Diabetes. Trends Endocrinol. Metab. 2016, 27, 142–152. [Google Scholar] [CrossRef]
- Dayeh, T.; Ling, C. Does epigenetic dysregulation of pancreatic islets contribute to impaired insulin secretion and type 2 diabetes? Biochem. Cell Biol. 2015, 93, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Spaeth, J.; Walker, E.M.; Stein, R. Impact of Pdx1-associated chromatin modifiers on islet β-cells. Diabetes Obes. Metab. 2016, 18, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Haumaitre, C. Epigenetic Regulation of Pancreatic Islets. Curr. Diabetes Rep. 2013, 13, 624–632. [Google Scholar] [CrossRef]
- Schuit, F. Epigenetic programming of glucose-regulated insulin release. J. Clin. Investig. 2015, 125, 2565–2568. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian microRNA Host Genes and Transcription Units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskerville, S. Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. RNA 2005, 11, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Borchert, G.; Lanier, W.; Davidson, B.L. RNA polymerase III transcribes human microRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Monteys, A.M.; Spengler, R.M.; Wan, J.; Tecedor, L.; Lennox, K.A.; Xing, Y.; Davidson, B.L. Structure and activity of putative intronic miRNA promoters. RNA 2010, 16, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, P.; Palanichamy, J.K.; Singh, A.; Das, P.; Bhagat, M.; Kassab, M.A.; Sinha, S.; Chattopadhyay, P. Biogenesis of intronic miRNAs located in clusters by independent transcription and alternative splicing. RNA 2013, 20, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelfattah, A.M.; Park, C.; Choi, M.Y. Update on non-canonical microRNAs. Biomol. Concepts 2014, 5, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nat. Cell Biol. 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Yeom, K.-H.; Lee, Y.; Han, J.; Suh, M.R.; Kim, V.N. Characterization of DGCR8/Pasha, the essential cofactor for Drosha in primary miRNA processing. Nucleic Acids Res. 2006, 34, 4622–4629. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, Y.; Yeom, K.-H.; Nam, J.-W.; Heo, I.; Rhee, J.-K.; Sohn, S.Y.; Cho, Y.; Zhang, B.-T.; Kim, V.N. Molecular Basis for the Recognition of Primary microRNAs by the Drosha-DGCR8 Complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nat. Cell Biol. 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Bohnsack, M.T.; Czaplinski, K.; Görlich, D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Hutvagner, G.; McLachlan, J.; Pasquinelli, A.E.; Bálint, É.; Tuschl, T.; Zamore, P.D. A Cellular Function for the RNA-Interference Enzyme Dicer in the Maturation of the let-7 Small Temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Kolb, F.A.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single Processing Center Models for Human Dicer and Bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.; Iwasaki, S.; Liu, Q.; Tomari, Y. ATP-dependent human RISC assembly pathways. Nat. Struct. Mol. Biol. 2009, 17, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Chendrimada, T.P.; Cooch, N.; Shiekhattar, R. Human RISC Couples MicroRNA Biogenesis and Posttranscriptional Gene Silencing. Cell 2005, 123, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Shih, I.-H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimson, A.; Farh, K.K.-H.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA Targeting Specificity in Mammals: Determinants beyond Seed Pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.J.; Scheel, T.; Luna, J.; Park, C.Y.; Fak, J.J.; Nishiuchi, E.; Rice, C.M.; Darnell, R. miRNA-target chimeras reveal miRNA 3′-end pairing as a major determinant of Argonaute target specificity. Nat. Commun. 2015, 6, 8864. [Google Scholar] [CrossRef]
- Friedman, R.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2008, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Bushati, N.; Cohen, S.M. microRNA Functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef]
- Deng, Q.; Hu, H.; Yu, X.; Liu, S.; Wang, L.; Chen, W.; Zhang, C.; Zeng, Z.; Cao, Y.; Xu-Monette, Z.Y.; et al. Tissue-specific microRNA expression alters cancer susceptibility conferred by a TP53 noncoding variant. Nat. Commun. 2019, 10, 5061. [Google Scholar] [CrossRef] [Green Version]
- Schulte, C.; Zeller, T. microRNA-based diagnostics and therapy in cardiovascular disease—Summing up the facts. Cardiovasc. Diagn. Ther. 2015, 5, 17–36. [Google Scholar] [CrossRef]
- Genemaras, A.A.; Ennis, H.; Kaplan, L.; Huang, C.-Y.; Huang, C.-Y.C. Inflammatory cytokines induce specific time- and concentration-dependent MicroRNA release by chondrocytes, synoviocytes, and meniscus cells. J. Orthop. Res. 2015, 34, 779–790. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, R.; De Rosa, S.; Leistner, D.; Boeckel, J.-N.; Keller, T.; Fichtlscherer, S.; Dimmeler, S.; Zeiher, A.M. Transcoronary Concentration Gradient of microRNA-133a and Outcome in Patients with Coronary Artery Disease. Am. J. Cardiol. 2017, 120, 15–24. [Google Scholar] [CrossRef]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhu, D.; Huang, L.; Zhang, J.; Bian, Z.; Chen, X.; Liu, Y.; Zhang, C.-Y.; Zen, K. Argonaute 2 Complexes Selectively Protect the Circulating MicroRNAs in Cell-Secreted Microvesicles. PLoS ONE 2012, 7, e46957. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafi-Shoushtari, H.; Kristo, F.; Li, Y.; Shioda, T.; Cohen, D.E.; Gerszten, R.E.; Näär, A.M. MicroRNA-33 and the SREBP Host Genes Cooperate to Control Cholesterol Homeostasis. Science 2010, 328, 1566–1569. [Google Scholar] [CrossRef] [Green Version]
- Lovis, P.; Roggli, E.; Laybutt, D.R.; Gattesco, S.; Yang, J.-Y.; Widmann, C.; Abderrahmani, A.; Regazzi, R. Alterations in microRNA expression contribute to fatty acid-induced pancreatic β-cell dysfunction. Diabetes 2008, 57, 2728–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.-B.; Wang, M.-N.; Li, Q.; Guo, L.; Yang, Y.-M.; Li, P.-J.; Wang, W.; Zhang, J.-C. MicroRNA-34a contributes to the protective effects of glucagon-like peptide-1 against lipotoxicity in INS-1 cells. Chin. Med. J. 2012, 125, 4202–4208. [Google Scholar]
- Lin, X.; Guan, H.; Huang, Z.; Liu, J.; Li, H.; Wei, G.; Cao, X.; Li, Y. Downregulation of Bcl-2 Expression by miR-34a Mediates Palmitate-Induced Min6 Cells Apoptosis. J. Diabetes Res. 2014, 2014, 258695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Hao, L.; Li, S.; Lin, S.; Lv, L.; Chen, Y.; Cui, H.; Zi, T.; Chu, X.; Na, L.; et al. Elevated circulating stearic acid leads to a major lipotoxic effect on mouse pancreatic beta cells in hyperlipidaemia via a miR-34a-5p-mediated PERK/p53-dependent pathway. Diabetologia 2016, 59, 1247–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Liu, C.-X.; Wang, G.; Yang, H.; Yao, X.-M.; Hua, Q.; Li, X.-Y.; Zhang, H.-M.; Ma, M.-Z.; Su, Q.; et al. LncRNA LEGLTBC Functions as a ceRNA to Antagonize the Effects of miR-34a on the Downregulation of SIRT1 in Glucolipotoxicity-Induced INS-1 Beta Cell Oxidative Stress and Apoptosis. Oxid. Med. Cell. Longev. 2019, 2019, 4010764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Zhang, H.; Wang, B.; Zhang, Y.; Zheng, X.; Shao, B.; Zhuge, Q.; Jin, K. Key Signaling Pathways in Aging and Potential Interventions for Healthy Aging. Cells 2021, 10, 660. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Liu, R.; Kasinski, A.L.; Shen, H.; Slack, F.J.; Tang, D.G. MicroRNA-34a: Potent Tumor Suppressor, Cancer Stem Cell Inhibitor, and Potential Anticancer Therapeutic. Front. Cell Dev. Biol. 2021, 9, 640587. [Google Scholar] [CrossRef]
- Agbu, P.; Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef]
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dong, J.; Ren, B. MicroRNA-182-5p contributes to the protective effects of thrombospondin 1 against lipotoxicity in INS-1 cells. Exp. Ther. Med. 2018, 16, 5272–5279. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Yu, Y.; Zhang, Y.; Li, Y.; Chu, X.; Lu, H.; Sun, C. Overexpression of miR-297b-5p protects against stearic acid-induced pancreatic β-cell apoptosis by targeting LATS2. Am. J. Physiol. Metab. 2020, 318, E430–E439. [Google Scholar] [CrossRef]
- Yu, Y.; Guo, R.; Zhang, Y.; Shi, H.; Sun, H.; Chu, X.; Wu, X.; Lu, H.; Sun, C. miRNA-mRNA profile and regulatory network in stearic acid-treated β-cell dysfunction. J. Endocrinol. 2020, 246, 13–27. [Google Scholar] [CrossRef]
- Li, Y.; Xu, X.; Liang, Y.; Liu, S.; Xiao, H.; Li, F.; Cheng, H.; Fu, Z. miR-375 enhances palmitate-induced lipoapoptosis in insulin-secreting NIT-1 cells by repressing myotrophin (V1) protein expression. Int. J. Clin. Exp. Pathol. 2010, 3, 254–264. [Google Scholar]
- Wang, J.; Lin, Z.; Yang, Z.; Liu, X. lncRNA Eif4g2 improves palmitate-induced dysfunction of mouse β-cells via modulation of Nrf2 activation. Exp. Cell Res. 2020, 396, 112291. [Google Scholar] [CrossRef] [PubMed]
- Paterson, M.R.; Kriegel, A.J. MiR-146a/b: A family with shared seeds and different roots. Physiol. Genom. 2017, 49, 243–252. [Google Scholar] [CrossRef]
- Shao, J.; Ding, Z.; Peng, J.; Zhou, R.; Li, L.; Qian, Q.; Chen, Y. MiR-146a-5p promotes IL-1β-induced chondrocyte apoptosis through the TRAF6-mediated NF-kB pathway. Inflamm. Res. 2020, 69, 619–630. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Z.; Chen, Q.; Yang, L.; Yin, J. miR-146b Regulates Cell Proliferation and Apoptosis in Gastric Cancer by Targeting PTP1B. Dig. Dis. Sci. 2020, 65, 457–463. [Google Scholar] [CrossRef]
- Zhang, W.; Shao, M.; He, X.; Wang, B.; Li, Y.; Guo, X. Overexpression of microRNA-146 protects against oxygen-glucose deprivation/recovery-induced cardiomyocyte apoptosis by inhibiting the NF-κB/TNF-α signaling pathway. Mol. Med. Rep. 2017, 17, 1913–1918. [Google Scholar] [CrossRef]
- Wang, Q.; Li, D.; Han, Y.; Ding, X.; Xu, T.; Tang, B. MicroRNA-146 protects A549 and H1975 cells from LPS-induced apoptosis and inflammation injury. J. Biosci. 2017, 42, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Fred, R.G.; Bang-Berthelsen, C.H.; Mandrup-Poulsen, T.; Grunnet, L.G.; Welsh, N. High Glucose Suppresses Human Islet Insulin Biosynthesis by Inducing miR-133a Leading to Decreased Polypyrimidine Tract Binding Protein-Expression. PLoS ONE 2010, 5, e10843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Liu, J.; Dai, Y.; Zhou, N.; Ji, C.; Li, X. MiR-146b attenuates high-fat diet-induced non-alcoholic steatohepatitis in mice. J. Gastroenterol. Hepatol. 2015, 30, 933–943. [Google Scholar] [CrossRef]
- Liang, Q.; Chen, H.; Xu, X.; Jiang, W. miR-182-5p Attenuates High-Fat-Diet-Induced Nonalcoholic Steatohepatitis in Mice. Ann. Hepatol. 2019, 18, 116–125. [Google Scholar] [CrossRef]
- Qin, S.-B.; Peng, D.-Y.; Lu, J.-M.; Ke, Z.-P. MiR-182-5p inhibited oxidative stress and apoptosis triggered by oxidized low-density lipoprotein via targeting toll-like receptor 4. J. Cell. Physiol. 2018, 233, 6630–6637. [Google Scholar] [CrossRef]
- Bai, J.; Xia, M.; Xue, Y.; Ma, F.; Cui, A.; Sun, Y.; Han, Y.; Xu, X.; Zhang, F.; Hu, Z.; et al. Thrombospondin 1 improves hepatic steatosis in diet-induced insulin-resistant mice and is associated with hepatic fat content in humans. EBioMedicine 2020, 57, 102849. [Google Scholar] [CrossRef]
- Min-Debartolo, J.; Schlerman, F.; Akare, S.; Wang, J.; McMahon, J.; Zhan, Y.; Syed, J.; He, W.; Zhang, B.; Martinez, R.V. Thrombospondin-I is a critical modulator in non-alcoholic steatohepatitis (NASH). PLoS ONE 2019, 14, e0226854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maimaitiyiming, H.; Clemons, K.; Zhou, Q.; Norman, H.; Wang, S. Thrombospondin1 Deficiency Attenuates Obesity-Associated Microvascular Complications in ApoE−/− Mice. PLoS ONE 2015, 10, e0121403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Tian, W.; Chen, Y.; Wang, L.; Jiang, Y.; Gao, B.; Luo, D. MicroRNA-374c-5p inhibits the development of breast cancer through TATA-box binding protein associated factor 7-mediated transcriptional regulation of DEP domain containing 1. J. Cell. Biochem. 2019, 120, 15360–15368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Shi, Y.; Yang, Q.; Gu, W.-W.; He, Y.-P.; Zheng, H.-J.; Zhang, X.; Wang, J.-M.; Wang, J. miR-3074-5p Promotes the Apoptosis but Inhibits the Invasiveness of Human Extravillous Trophoblast-Derived HTR8/SVneo Cells In Vitro. Reprod. Sci. 2017, 25, 690–699. [Google Scholar] [CrossRef]
- Lou, S.; Gao, H.; Hong, H.; Zhu, Z.; Zhao, H. Inhibition of retinoic acid receptor α phosphorylation represses the progression of triple-negative breast cancer via transactivating miR-3074-5p to target DHRS3. J. Exp. Clin. Cancer Res. 2021, 40, 141. [Google Scholar] [CrossRef]
- Zhang, M.; Mou, L.; Liu, S.; Sun, F.; Gong, M. Circ_0001103 alleviates IL-1β-induced chondrocyte cell injuries by upregulating SIRT1 via targeting miR-375. Clin. Immunol. 2021, 227, 108718. [Google Scholar] [CrossRef]
- Guan, X.; Shi, A.; Zou, Y.; Sun, M.; Zhan, Y.; Dong, Y.; Fan, Z. EZH2-Mediated microRNA-375 Upregulation Promotes Progression of Breast Cancer via the Inhibition of FOXO1 and the p53 Signaling Pathway. Front. Genet. 2021, 12, 633756. [Google Scholar] [CrossRef]
- Wei, Z.; Tian, Z.; Zhang, L. CircPPP1R12A promotes the progression of colon cancer through regulating CTNNB1 via sponging miR-375. Anticancer Drugs 2021, 32, 635–646. [Google Scholar] [CrossRef]
- Yan, X.-L.; Luo, Q.-Y.; Zhou, S.-N.; Pan, W.-T.; Zhang, L.; Yang, D.-J.; Qui, M.-Z. MicroRNA-375 reverses the expression of PD-L1 by inactivating the JAK2/STAT3 signaling pathways in gastric cancer. Clin. Res. Hepatol. Gastroenterol. 2020, 101574. [Google Scholar] [CrossRef]
- Xu, R.; Yin, S.; Zheng, M.; Pei, X.; Ji, X. Circular RNA circZFR Promotes Hepatocellular Carcinoma Progression by Regulating miR-375/HMGA2 Axis. Dig. Dis. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Wang, L.; Han, N.; Yin, G. miR-375-3p contributes to hypoxia-induced apoptosis by targeting forkhead box P1 (FOXP1) and Bcl2 like protein 2 (Bcl2l2) in rat cardiomyocyte h9c2 cells. Biotechnol. Lett. 2021, 43, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ren, Y.; Sundaram, K.; Mu, J.; Sriwastva, M.K.; Dryden, G.W.; Lei, C.; Zhang, L.; Yan, J.; Zhang, X.; et al. miR-375 prevents high-fat diet-induced insulin resistance and obesity by targeting the aryl hydrocarbon receptor and bacterial tryptophanase (tnaA) gene. Theranostics 2021, 11, 4061–4077. [Google Scholar] [CrossRef] [PubMed]
- Li, X. miR-375, a microRNA related to diabetes. Gene 2014, 533, 1–4. [Google Scholar] [CrossRef]
- Furth, N.; Aylon, Y. The LATS1 and LATS2 tumor suppressors: Beyond the Hippo pathway. Cell Death Differ. 2017, 24, 1488–1501. [Google Scholar] [CrossRef] [Green Version]
- Aylon, Y.; Gershoni, A.; Rotkopf, R.; Biton, I.E.; Porat, Z.; Koh, A.P.; Sun, X.; Lee, Y.; Fiel, M.-I.; Hoshida, Y.; et al. The LATS2 tumor suppressor inhibits SREBP and suppresses hepatic cholesterol accumulation. Genes Dev. 2016, 30, 786–797. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Xiong, L.; Li, J.; Peng, Z.; Zhang, L.; Shi, P.; Gong, Y.; Xiao, H. Circ-Tulp4 promotes β-cell adaptation to lipotoxicity by regulating soat1 expression. J. Mol. Endocrinol. 2020, 65, 149–161. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, Y.; Wang, W.; Zhang, J.; Li, Q. Glucagon-like peptide-1 improves β-cell dysfunction by suppressing the miR-27a-induced downregulation of ATP-binding cassette transporter A1. Biomed. Pharmacother. 2017, 96, 497–502. [Google Scholar] [CrossRef]
- Wijesekara, N.; Kaur, A.; Westwell-Roper, C.; Nackiewicz, M.; Soukhatcheva, G.; Hayden, M.R.; Verchere, C.B. ABCA1 deficiency and cellular cholesterol accumulation increases islet amyloidogenesis in mice. Diabetologia 2016, 59, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Von Eckardstein, A.; Widmann, C. High-density lipoprotein, beta cells, and diabetes. Cardiovasc. Res. 2014, 103, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutti, S.; Ehses, J.A.; Sibler, R.A.; Prazak, R.; Rohrer, L.; Georgopoulos, S.; Meier, D.T.; Niclauss, N.; Berney, T.; Donath, M.Y.; et al. Low- and High-Density Lipoproteins Modulate Function, Apoptosis, and Proliferation of Primary Human and Murine Pancreatic β-Cells. Endocrinology 2009, 150, 4521–4530. [Google Scholar] [CrossRef] [Green Version]
- Fryirs, M.A.; Barter, P.J.; Appavoo, M.; Tuch, B.E.; Tabet, F.; Heather, A.K.; Rye, K.-A. Effects of High-Density Lipoproteins on Pancreatic β-Cell Insulin Secretion. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1642–1648. [Google Scholar] [CrossRef] [Green Version]
- Cochran, B.J.; Bisoendial, R.J.; Hou, L.; Glaros, E.N.; Rossy, J.; Thomas, S.R.; Barter, P.J.; Rye, K.-A. Apolipoprotein A-I Increases Insulin Secretion and Production From Pancreatic β-Cells via a G-Protein-cAMP-PKA-FoxO1–Dependent Mechanism. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2261–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Tang, S.; Wu, B.J.; Ong, K.-L.; Westerterp, M.; Barter, P.J.; Cochran, B.J.; Tabet, F.; Rye, K.-A. Apolipoprotein A-l improves pancreatic β-cell function independent of the ATP-binding cassette transporters ABCA1 and ABCG1. FASEB J. 2019, 33, 8479–8489. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Del Giudice, R.; Nagao, M.; Grönberg, C.; Eliasson, L.; Lagerstedt, J.O. Apolipoprotein A-I primes beta cells to increase glucose stimulated insulin secretion. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165613. [Google Scholar] [CrossRef]
- Brunham, L.R.; Kruit, J.K.; Pape, T.D.; Timmins, J.M.; Reuwer, A.Q.; Vasanji, Z.; Marsh, B.J.; Rodrigues, B.; Johnson, J.D.; Parks, J.S.; et al. β-cell ABCA1 influences insulin secretion, glucose homeostasis and response to thiazolidinedione treatment. Nat. Med. 2007, 13, 340–347. [Google Scholar] [CrossRef]
- Kruit, J.K.; Kremer, P.H.C.; Dai, L.; Tang, R.; Ruddle, P.; De Haan, W.; Brunham, L.R.; Verchere, C.B.; Hayden, M.R. Cholesterol efflux via ATP-binding cassette transporter A1 (ABCA1) and cholesterol uptake via the LDL receptor influences cholesterol-induced impairment of beta cell function in mice. Diabetologia 2010, 53, 1110–1119. [Google Scholar] [CrossRef]
- Kruit, J.K.; Wijesekara, N.; Fox, J.E.M.; Dai, X.-Q.; Brunham, L.R.; Searle, G.J.; Morgan, G.P.; Costin, A.J.; Tang, R.; Bhattacharjee, A.; et al. Islet Cholesterol Accumulation Due to Loss of ABCA1 Leads to Impaired Exocytosis of Insulin Granules. Diabetes 2011, 60, 3186–3196. [Google Scholar] [CrossRef] [Green Version]
- Dullaart, R.P.; Annema, W.; de Boer, J.F.; Tietge, U.J. Pancreatic β-cell function relates positively to HDL functionality in well-controlled Type 2 diabetes mellitus. Atherosclerosis 2012, 222, 567–573. [Google Scholar] [CrossRef]
- Bardini, G.; Dicembrini, I.; Rotella, C.M.; Giannini, S. Correlation between HDL cholesterol levels and beta-cell function in subjects with various degree of glucose tolerance. Acta Diabetol. 2013, 50, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Xu, H.; Zhou, H.; Ren, X.; Han, T.; Chen, Y.; Qiu, H.; Wu, P.; Zheng, J.; Wang, L.; et al. Associations of lipid profiles with insulin resistance and β cell function in adults with normal glucose tolerance and different categories of impaired glucose regulation. PLoS ONE 2017, 12, e0172221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, H.; Mishra, M.; Bajpai, S.; Pokhria, D.; Arya, A.K.; Singh, R.K.; Tripathi, K. Correlation of insulin resistance, beta cell function and insulin sensitivity with serum sFas and sFasL in newly diagnosed type 2 diabetes. Acta Diabetol. 2011, 50, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, T.V.; Succurro, E.; Marini, M.A.; Pedace, E.; Andreozzi, F.; Perticone, M.; Sciacqua, A.; Perticone, F.; Sesti, G. HDL cholesterol is an independent predictor of β-cell function decline and incident type 2 diabetes: A longitudinal study. Diabetes Metab. Res. Rev. 2020, 36, e3289. [Google Scholar] [CrossRef] [PubMed]
- Vergeer, M.; Brunham, L.R.; Koetsveld, J.; Kruit, J.K.; Verchere, C.B.; Kastelein, J.J.; Hayden, M.R.; Stroes, E.S. Carriers of Loss-of-Function Mutations in ABCA1 Display Pancreatic β-Cell Dysfunction. Diabetes Care 2010, 33, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Rickels, M.R.; Goeser, E.S.; Fuller, C.; Lord, C.; Bowler, A.M.; Doliba, N.M.; Hegele, R.A.; Cuchel, M. Loss-of-Function Mutations in ABCA1 and Enhanced β-Cell Secretory Capacity in Young Adults. Diabetes 2014, 64, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Ono, K. Functions of microRNA-33a/b and microRNA therapeutics. J. Cardiol. 2016, 67, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Ono, K.; Horie, T.; Nishino, T.; Baba, O.; Kuwabara, Y.; Yokode, M.; Kita, T.; Kimura, T. MicroRNA-33a/b in lipid metabolism—novel ‘thrifty’ models. Circ. J. 2015, 79, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Zhang, L.-H.; Wijesekara, N.; De Haan, W.; Butland, S.; Bhattacharjee, A.; Hayden, M.R. Regulation of ABCA1 Protein Expression and Function in Hepatic and Pancreatic Islet Cells by miR-145. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2724–2732. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Huang, C.; Yang, P.; Bu, L. miR-383 ameliorates high glucose-induced β-cells apoptosis and hyperglycaemia in high-fat induced diabetic mice. Life Sci. 2020, 263, 118571. [Google Scholar] [CrossRef]
- Tarlton, J.M.; Lightbody, R.J.; Patterson, S.; Graham, A. Protection against Glucolipotoxicity by High Density Lipoprotein in Human PANC-1 Hybrid 1.1B4 Pancreatic Beta Cells: The Role of microRNA. Biology 2021, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Sedgeman, L.R.; Beysen, C.; Solano, M.A.R.; Michell, D.L.; Sheng, Q.; Zhao, S.; Turner, S.; Linton, M.F.; Vickers, K.C. Beta cell secretion of miR-375 to HDL is inversely associated with insulin secretion. Sci. Rep. 2019, 9, 3803. [Google Scholar] [CrossRef]
- Yang, J.; Lv, Y.; Zhao, Z.; Li, W.; Xiang, S.; Zhou, L.; Gao, A.; Yan, B.; Ou, L.; Ling, H.; et al. A microRNA-24-to-secretagogin regulatory pathway mediates cholesterol-induced inhibition of insulin secretion. Int. J. Mol. Med. 2019, 44, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, L.; Liu, R.; Song, Y.; Zhang, X.; Niu, W.; Kumar, A.K.; Guo, Z.; Hu, Z. Obesity-induced overexpression of miRNA-24 regulates cholesterol uptake and lipid metabolism by targeting SR-B1. Gene 2018, 668, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Zhu, X.; Zheng, Z.; Mo, Z.-C.; Peng, X.-S.; Zeng, Y.-Z.; Ou, H.-X.; Zhang, Q.-H.; Qi, H.-Z.; Zhao, G.-J.; et al. MicroRNA-24 aggravates atherosclerosis by inhibiting selective lipid uptake from HDL cholesterol via the post-transcriptional repression of scavenger receptor class B type I. Atherosclerosis 2018, 270, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, J.; Chen, Z.; Cao, W.; Xu, H.; Gou, D.; Zhu, J. MicroRNA-24 can control triacylglycerol synthesis in goat mammary epithelial cells by targeting the fatty acid synthase gene. J. Dairy Sci. 2015, 98, 9001–9014. [Google Scholar] [CrossRef]
- Ng, R.; Wu, H.; Xiao, H.; Chen, X.; Willenbring, H.; Steer, C.J.; Song, G. Inhibition of microRNA-24 expression in liver prevents hepatic lipid accumulation and hyperlipidaemia. Hepatology 2014, 60, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Kai, K.; Dittmar, R.; Sen, S. Secretory microRNAs as biomarkers of cancer. Semin. Cell Dev. Biol. 2018, 78, 22–36. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]
- Mateescu, B.; Kowal, E.J.K.; Van Balkom, B.W.M.; Bartel, S.; Bhattacharyya, S.N.; Buzás, E.I.; Buck, A.; De Candia, P.; Chow, W.-N.; Das, S.; et al. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA—An ISEV position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral Sphingomyelinase 2 (nSMase2)-dependent Exosomal Transfer of Angiogenic MicroRNAs Regulate Cancer Cell Metastasis. J. Biol. Chem. 2013, 288, 10849–10859. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.; Danielson, K.M.; Benton, M.C.; Ziegler, O.; Shah, R.; Stubbs, R.S.; Das, S.; Macartney-Coxson, D. miRNA Signatures of Insulin Resistance in Obesity. Obesity 2017, 25, 1734–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Li, K.; Zheng, P.; Wang, Y.; Lv, Y.; Shen, L.; Chen, Y.; Xue, Z.; Li, B.; Jin, L.; et al. Weighted Gene Coexpression Network Analysis Identified MicroRNA Coexpression Modules and Related Pathways in Type 2 Diabetes Mellitus. Oxid. Med. Cell. Longev. 2019, 2019, 9567641. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Wiklander, O.P.; Fritz, T.; Caidahl, K.; El-Andaloussi, S.; Zierath, J.R.; Krook, A. Circulating Exosomal miR-20b-5p is Elevated in Type 2 Diabetes and Could Impair Insulin Action in Human Skeletal Muscle. Diabetes 2018, 68, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma MicroRNA Profiling Reveals Loss of Endothelial MiR-126 and Other MicroRNAs in Type 2 Diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; Van Ommen, B.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zúñiga, O.A.; et al. A plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: From the CORDIOPREV study. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Karolina, D.S.; Armugam, A.; Tavintharan, S.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. MicroRNA 144 Impairs Insulin Signaling by Inhibiting the Expression of Insulin Receptor Substrate 1 in Type 2 Diabetes Mellitus. PLoS ONE 2011, 6, e22839. [Google Scholar] [CrossRef]
- Mensà, E.; Giuliani, A.; Matacchione, G.; Gurău, F.; Bonfigli, A.R.; Romagnoli, F.; De Luca, M.; Sabbatinelli, J.; Olivieri, F. Circulating miR-146a in healthy aging and type 2 diabetes: Age- and gender-specific trajectories. Mech. Ageing Dev. 2019, 180, 1–10. [Google Scholar] [CrossRef]
- Monfared, Y.K.; Honardoost, M.; Sarookhani, M.R.; Farzam, S.A. Circulating miR-135 May Serve as a Novel Co-biomarker of HbA1c in Type 2 Diabetes. Appl. Biochem. Biotechnol. 2020, 191, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Wang, G.; Xu, C.; Zeng, M.; Lin, F.; Wu, J.; Wan, Q. Circulating miR-30c as a predictive biomarker of type 2 diabetes mellitus with coronary heart disease by regulating PAI-1/VN interactions. Life Sci. 2019, 239, 117092. [Google Scholar] [CrossRef]
- Luo, M.; Xu, C.; Luo, Y.; Wang, G.; Wu, J.; Wan, Q. Circulating miR-103 family as potential biomarkers for type 2 diabetes through targeting CAV-1 and SFRP4. Acta Diabetol. 2020, 57, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Parrizas, M.; Mundet, X.; Castaño, C.; Canivell, S.; Cos, X.; Brugnara, L.; Giráldez-García, C.; Regidor, E.; Mata-Cases, M.; Franch-Nadal, J.; et al. miR-10b and miR-223-3p in serum microvesicles signal progression from prediabetes to type 2 diabetes. J. Endocrinol. Investig. 2019, 43, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Massaro, J.D.; Polli, C.D.; Costa e Silva, M.; Alves, C.C.; Passos, G.A.; Sakamoto-Hojo, E.T.; Miranda, W.R.D.H.; Cezar, N.J.B.; Rassi, D.M.; Crispim, F.; et al. Post-transcriptional markers associated with clinical complications in Type 1 and Type 2 diabetes mellitus. Mol. Cell. Endocrinol. 2019, 490, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Prabu, P.; Poongothai, S.; Shanthirani, C.S.; Anjana, R.M.; Mohan, V.; Balasubramanyam, M. Altered circulatory levels of miR-128, BDNF, cortisol and shortened telomeres in patients with type 2 diabetes and depression. Acta Diabetol. 2020, 57, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.; Liang, Y.; Zhou, X. Expression Profiling and Clinical Significance of Plasma MicroRNAs in Diabetic Nephropathy. J. Diabetes Res. 2019, 2019, 5204394. [Google Scholar] [CrossRef]
- Lareyre, F.; Clément, M.; Moratal, C.; Loyer, X.; Jean-Baptiste, E.; Hassen-Khodja, R.; Chinetti, G.; Mallat, Z.; Raffort, J. Differential micro-RNA expression in diabetic patients with abdominal aortic aneurysm. Biochimie 2019, 162, 1–7. [Google Scholar] [CrossRef]
- García-Jacobo, R.E.; Uresti-Rivera, E.E.; Portales-Pérez, D.P.; González-Amaro, R.; Lara-Ramírez, E.E.; Enciso-Moreno, J.A.; García-Hernández, M.H. Circulating miR-146a, miR-34a and miR-375 in type 2 diabetes patients, pre-diabetic and normal-glycaemic individuals in relation to β-cell function, insulin resistance and metabolic parameters. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1092–1100. [Google Scholar] [CrossRef]
- Kokkinopoulou, I.; Maratou, E.; Mitrou, P.; Boutati, E.; Sideris, D.C.; Fragoulis, E.G.; Christodoulou, M.-I. Decreased expression of microRNAs targeting type-2 diabetes susceptibility genes in peripheral blood of patients and predisposed individuals. Endocrine 2019, 66, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Avgeris, M.; Kokkinopoulou, I.; Maratou, E.; Mitrou, P.; Boutati, E.; Scorilas, A.; Fragoulis, E.G.; Christodoulou, M.-I. Blood-based analysis of 84 microRNAs identifies molecules deregulated in individuals with type-2 diabetes, risk factors for the disease or metabolic syndrome. Diabetes Res. Clin. Pract. 2020, 164, 108187. [Google Scholar] [CrossRef]
- Ghai, V.; Baxter, D.; Wu, X.; Kim, T.; Kuusisto, J.; Laakso, M.; Connolly, T.; Li, Y.; Andrade-Gordon, P.; Wang, K. Circulating RNAs as predictive markers for the progression of type 2 diabetes. J. Cell. Mol. Med. 2019, 23, 2753–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Candia, P.; Spinetti, G.; Specchia, C.; Sangalli, E.; La Sala, L.; Uccellatore, A.; Lupini, S.; Genovese, S.; Matarese, G.; Ceriello, A. A unique plasma microRNA profile defines type 2 diabetes progression. PLoS ONE 2017, 12, e0188980. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.-M.; Chen, L.-H.; Hong, M.; Chen, Y.-Y.; Yang, X.-R.; Tang, S.-M.; Yuan, Q.-F.; Chen, W.-W. Serum microRNA profiling and bioinformatics analysis of patients with type 2 diabetes mellitus in a Chinese population. Mol. Med. Rep. 2017, 15, 2143–2153. [Google Scholar] [CrossRef] [Green Version]
- Aljaibeji, H.; Elemam, N.M.; Mohammed, A.K.; Hasswan, H.; Al Thahyabat, M.; Alkhayyal, N.; Sulaiman, N.; Taneera, J. Let7b-5p is Upregulated in the Serum of Emirati Patients with Type 2 Diabetes and Regulates Insulin Secretion in INS-1 Cells. Exp. Clin. Endocrinol. Diabetes 2020. [Google Scholar] [CrossRef]
- Shaker, O.G.; Abdelaleem, O.O.; Mahmoud, R.H.; Abdelghaffar, N.K.; Ahmed, T.I.; Said, O.M.; Zaki, O.M. Diagnostic and prognostic role of serum miR-20b, miR-17-3p, HOTAIR, and MALAT1 in diabetic retinopathy. IUBMB Life 2019, 71, 310–320. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, G.; Yang, C.; Zhou, K.; Shen, B.; Liang, H.; Jiang, X. The Role of Circulating MicroRNA-126 (miR-126): A Novel Biomarker for Screening Prediabetes and Newly Diagnosed Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2014, 15, 10567–10577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wander, P.L.; Enquobahrie, D.A.; Bammler, T.K.; Srinouanprachanh, S.; Macdonald, J.; Kahn, S.E.; Leonetti, D.; Fujimoto, W.Y.; Boyko, E.J. Short Report: Circulating microRNAs are associated with incident diabetes over 10 years in Japanese Americans. Sci. Rep. 2020, 10, 6509. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Wang, J.; Wu, J.; Song, J.; Zhang, C.-Y.; Zhang, C.; Wang, C.; Wang, J.-J. Increased serum miR-7 is a promising biomarker for type 2 diabetes mellitus and its microvascular complications. Diabetes Res. Clin. Pract. 2017, 130, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Radović, N.; Jakoba, N.N.; Petrovic, N.; Milosavljević, A.; Brković, B.; Roganović, J. MicroRNA-146a and microRNA-155 as novel crevicular fluid biomarkers for periodontitis in non-diabetic and type 2 diabetic patients. J. Clin. Periodontol. 2018, 45, 663–671. [Google Scholar] [CrossRef]
- Jaeger, A.; Zollinger, L.; Saely, C.H.; Muendlein, A.; Evangelakos, I.; Nasias, D.; Charizopoulou, N.; Schofield, J.D.; Othman, A.; Soran, H.; et al. Circulating microRNAs -192 and -194 are associated with the presence and incidence of diabetes mellitus. Sci. Rep. 2018, 8, 14274. [Google Scholar] [CrossRef] [PubMed]
- Alipoor, B.; Ghaedi, H.; Meshkani, R.; Omrani, M.D.; Sharifi, Z.; Golmohammadi, T. The rs2910164 variant is associated with reduced miR-146a expression but not cytokine levels in patients with type 2 diabetes. J. Endocrinol. Investig. 2018, 41, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Motawae, T.M.; Ismail, M.F.; Shabayek, M.I.; Seleem, M.M. MicroRNAs 9 and 370 Association with Biochemical Markers in T2D and CAD Complication of T2D. PLoS ONE 2015, 10, e0126957. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, T.; Wei, Y.; Cai, W.-J.; Feng, G.; Chang, X.-Y.; Sun, K. Epigenetic regulation of microRNA-375 and its role as DNA epigenetic marker of type 2 diabetes mellitus in Chinese Han population. Int. J. Clin. Exp. Pathol. 2017, 10, 11986–11994. [Google Scholar]
- Kulkarni, M.; Leszczynska, A.; Wei, G.; Winkler, M.A.; Tang, J.; Funari, V.A.; Deng, N.; Liu, Z.; Punj, V.; Deng, S.X.; et al. Genome-wide analysis suggests a differential microRNA signature associated with normal and diabetic human corneal limbus. Sci. Rep. 2017, 7, 3448. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, H.; Si, H.; Li, X.; Ding, X.; Sheng, Q.; Chen, P.; Zhang, H. Serum miR-23a, a potential biomarker for diagnosis of pre-diabetes and type 2 diabetes. Acta Diabetol. 2014, 51, 823–831. [Google Scholar] [CrossRef]
- Jiménez-Lucena, R.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; López-Moreno, J.; Roncero-Ramos, I.; Molina-Abril, H.; Yubero-Serrano, E.M.; Caballero-Villarraso, J.; Delgado-Lista, J.; Castaño, J.P.; et al. Circulating miRNAs as Predictive Biomarkers of Type 2 Diabetes Mellitus Development in Coronary Heart Disease Patients from the CORDIOPREV Study. Mol. Ther. Nucleic Acids 2018, 12, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lucena, R.; Alcala-Diaz, J.F.; Roncero-Ramos, I.; Lopez-Moreno, J.; Camargo, A.; Gomez-Delgado, F.; Quintana-Navarro, G.M.; Vals-Delgado, C.; Rodriguez-Cantalejo, F.; Luque, R.M.; et al. MiRNAs profile as biomarkers of nutritional therapy for the prevention of type 2 diabetes mellitus: From the CORDIOPREV study. Clin. Nutr. 2021, 40, 1028–1038. [Google Scholar] [CrossRef]
- Luo, M.; Li, R.; Deng, X.; Ren, M.; Chen, N.; Zeng, M.; Yan, Y.; Xia, J.; Liu, F.; Ma, W.; et al. Platelet-derived miR-103b as a novel biomarker for the early diagnosis of type 2 diabetes. Acta Diabetol. 2015, 52, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Al-Kafaji, G.; Al-Mahroos, G.; Alsayed, N.A.; Hasan, Z.A.; Nawaz, S.; Bakhiet, M. Peripheral blood microRNA-15a is a potential biomarker for type 2 diabetes mellitus and pre-diabetes. Mol. Med. Rep. 2015, 12, 7485–7490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Dai, X.; Zhan, J.; Zhang, Y.; Zhang, H.; Zeng, S.; Xi, W.; Zhang, H. Profiling peripheral microRNAs in obesity and type 2 diabetes mellitus. APMIS 2015, 123, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Li, Y.-Q.; Liang, Y.-Z.; Dong, J.; He, Y.; Zhang, L.; Yan, Y.-X. Expression of miR-18a and miR-34c in circulating monocytes associated with vulnerability to type 2 diabetes mellitus and insulin resistance. J. Cell. Mol. Med. 2017, 21, 3372–3380. [Google Scholar] [CrossRef]
- Gallo, W.; Esguerra, J.L.S.; Eliasson, L.; Melander, O. miR-483-5p associates with obesity and insulin resistance and independently associates with new onset diabetes mellitus and cardiovascular disease. PLoS ONE 2018, 13, e0206974. [Google Scholar] [CrossRef]
- Sun, K.; Chang, X.; Yin, L.; Li, J.; Zhou, T.; Zhang, C.; Chen, X. Expression and DNA methylation status of microRNA-375 in patients with type 2 diabetes mellitus. Mol. Med. Rep. 2013, 9, 967–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, R.; Ghorbani, S.; Alipoor, B.; Panahi, G.; Khodabandehloo, H.; Esfahani, E.; Razi, F.; Meshkani, R. Decreased Serum Level of miR-155 is Associated with Obesity and its Related Metabolic Traits. Clin. Lab. 2018, 64, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lv, C.; Li, L.; Chen, S.; Liu, S.; Wang, C.; Su, B. Plasma miR-126 Is a Potential Biomarker for Early Prediction of Type 2 Diabetes Mellitus in Susceptible Individuals. BioMed Res. Int. 2013, 2013, 761617. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Spazzafumo, L.; Bonafè, M.; Recchioni, R.; Prattichizzo, F.; Marcheselli, F.; Micolucci, L.; Mensà, E.; Giuliani, A.; Santini, G.; et al. MiR-21-5p and miR-126a-3p levels in plasma and circulating angiogenic cells: Relationship with type 2 diabetes complications. Oncotarget 2015, 6, 35372–35382. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sundquist, J.; Zöller, B.; Memon, A.A.; Palmér, K.; Sundquist, K.; Bennet, L. Determination of 14 Circulating microRNAs in Swedes and Iraqis with and without Diabetes Mellitus Type 2. PLoS ONE 2014, 9, e86792. [Google Scholar] [CrossRef]
- Giannella, A.; Radu, C.M.; Franco, L.; Campello, E.; Simioni, P.; Avogaro, A.; De Kreutzenberg, S.V.; Ceolotto, G. Circulating levels and characterization of microparticles in patients with different degrees of glucose tolerance. Cardiovasc. Diabetol. 2017, 16, 118. [Google Scholar] [CrossRef]
- Yan, S.; Wang, T.; Huang, S.; Di, Y.; Huang, Y.; Liu, X.; Luo, Z.; Han, W.; Tianqiong, W. Differential expression of microRNAs in plasma of patients with prediabetes and newly diagnosed type 2 diabetes. Acta Diabetol. 2016, 53, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.-T.; Li, C.-L.; Tian, H.; Li, J.; Pei, Y.; Liu, Y.; Gong, Y.-P.; Fang, F.-S.; Sun, B.-R. MiR-199a is overexpressed in plasma of type 2 diabetes patients which contributes to type 2 diabetes by targeting GLUT4. Mol. Cell. Biochem. 2014, 397, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lopez, Y.O.N.; Garufi, G.; Seyhan, A.A. Altered levels of circulating cytokines and microRNAs in lean and obese individuals with prediabetes and type 2 diabetes. Mol. BioSyst. 2016, 13, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhao, J.; Chen, Y.; Lei, M. Biomarkers Associated with Ischemic Stroke in Diabetes Mellitus Patients. Cardiovasc. Toxicol. 2015, 16, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, L.; Shang, Q.; Lv, C.; Wang, C.; Su, B. Circulating miR-126 is a potential biomarker to predict the onset of type 2 diabetes mellitus in susceptible individuals. Biochem. Biophys. Res. Commun. 2015, 463, 60–63. [Google Scholar] [CrossRef]
- Prabu, P.; Rome, S.; Sathishkumar, C.; Aravind, S.; Mahalingam, B.; Shanthirani, C.S.; Gastebois, C.; Villard, A.; Mohan, V.; Balasubramanyam, M. Circulating MiRNAs of ‘Asian Indian Phenotype’ Identified in Subjects with Impaired Glucose Tolerance and Patients with Type 2 Diabetes. PLoS ONE 2015, 10, e0128372. [Google Scholar] [CrossRef] [Green Version]
- Baldeón, L.; Weigelt, K.; de Wit, H.; Özcan, B.; van Oudenaren, A.; Sempértegui, F.; Sijbrands, E.; Grosse, L.; Freire, W.; Drexhage, H.; et al. Decreased Serum Level of miR-146a as Sign of Chronic Inflammation in Type 2 Diabetic Patients. PLoS ONE 2014, 9, e115209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenin, R.; Sankaramoorthy, A.; Mohan, V.; Balasubramanyam, M. Altered immunometabolism at the interface of increased endoplasmic reticulum (ER) stress in patients with type 2 diabetes. J. Leukoc. Biol. 2015, 98, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Rezk, N.A.; Sabbah, N.A.; Saad, M.S.S. Role of MicroRNA 126 in screening, diagnosis, and prognosis of diabetic patients in Egypt. IUBMB Life 2016, 68, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.J.; Mercader, J.M.; Moreno-Navarrete, J.M.; Rovira, O.; Guerra, E.; Esteve, E.; Xifra, G.; Martínez, C.; Ricart, W.; Rieusset, J.; et al. Profiling of Circulating MicroRNAs Reveals Common MicroRNAs Linked to Type 2 Diabetes That Change with Insulin Sensitization. Diabetes Care 2014, 37, 1375–1383. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Wang, H.; Przybilla, D.; Franklin, B.S.; Dolf, A.; Pfeifer, P.; Schmitz, T.; Flender, A.; Endl, E.; Nickenig, G.; et al. Vascular endothelial microparticles-incorporated microRNAs are altered in patients with diabetes mellitus. Cardiovasc. Diabetol. 2016, 15, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyhan, A.A.; Lopez, Y.N.; Xie, H.; Yi, F.; Mathews, C.; Pasarica, M.; Pratley, R.E. Pancreas-enriched miRNAs are altered in the circulation of subjects with diabetes: A pilot cross-sectional study. Sci. Rep. 2016, 6, 31479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wan, S.; Yang, T.; Niu, D.; Zhang, A.; Yang, C.; Cai, J.; Wu, J.; Song, J.; Zhang, C.-Y.; et al. Increased serum microRNAs are closely associated with the presence of microvascular complications in type 2 diabetes mellitus. Sci. Rep. 2016, 6, 20032. [Google Scholar] [CrossRef] [Green Version]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chang, J.; Guo, J.-T.; Jiang, D.; Guo, H.; Taylor, J.M.; Block, T.M. Liver-Specific MicroRNA miR-122 Enhances the Replication of Hepatitis C Virus in Nonhepatic Cells. J. Virol. 2008, 82, 8215–8223. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.; Rebhan, M.A.E.; Crivelli, S.E.M.; Denzler, R.; Stoffel, M.; Hall, J. Miravirsen (SPC3649) can inhibit the biogenesis of miR-122. Nucleic Acids Res. 2014, 42, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Janssen, H.L.A.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; Van Der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV Infection by Targeting MicroRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.-K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Kapadia, C.H.; Luo, B.; Dang, M.N.; Irvin-Choy, N.; Valcourt, D.M.; Day, E.S. Polymer nanocarriers for MicroRNA delivery. J. Appl. Polym. Sci. 2020, 137, 48651. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, G.; Su, Z.; Jiang, Z.; Chen, L.; Wang, J.; Yu, S.; Liu, Z. Poly(amido amine) is an ideal carrier of miR-7 for enhancing gene silencing effects on the EGFR pathway in U251 glioma cells. Oncol. Rep. 2013, 29, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Guo, B.; Wu, H.; Tang, T.; Zhang, B.-T.; Zheng, L.; He, Y.; Yang, Z.; Pan, X.; Chow, H.; et al. A delivery system targeting bone formation surfaces to facilitate RNAi-based anabolic therapy. Nat. Med. 2012, 18, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Inoue, A.; Seers, T.; Hato, Y.; Igarashi, A.; Toyama, T.; Taganov, K.D.; Boldin, M.P.; Asahara, H. Identification of targets of tumor suppressor microRNA-34a using a reporter library system. Proc. Natl. Acad. Sci. USA 2017, 114, 3927–3932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, L.G.T.; Chan, T.A. Therapeutic targeting of tumor suppressor genes. Cancer 2015, 121, 1357–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hu, X.; Han, C.; Wang, L.; Zhang, X.; He, X.; Lu, X. Targeting tumor suppressor genes for cancer therapy. BioEssays 2015, 37, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.-E.; Suh, H.-W.; Bahal, R.; Josowitz, A.; Zhang, J.; Song, E.; Cui, J.; Noorbakhsh, S.; Jackson, C.; Bu, T.; et al. Nanoparticle-mediated intratumoral inhibition of miR-21 for improved survival in glioblastoma. Biomaterials 2019, 201, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Nishio, H.; Masumoto, H.; Sakamoto, K.; Yamazaki, K.; Ikeda, T.; Minatoya, K. MicroRNA-145-loaded poly(lactic-co-glycolic acid) nanoparticles attenuate venous intimal hyperplasia in a rabbit model. J. Thorac. Cardiovasc. Surg. 2019, 157, 2242–2251. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Feng, Y.; Xie, Y.; Shao, Y.; Wu, M.; Deng, X.; Yuan, W.-E.; Chen, Y.; Shi, X. Nanoparticle-microRNA-146a-5p polyplexes ameliorate diabetic peripheral neuropathy by modulating inflammation and apoptosis. Nanomed. Nanotechnol. Biol. Med. 2019, 17, 188–197. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Q.; Zhao, H.; Bishop, J.O.; Zhou, G.; Olson, L.K.; Moore, A. miR-216a-targeting theranostic nanoparticles promote proliferation of insulin-secreting cells in type 1 diabetes animal model. Sci. Rep. 2020, 10, 5302. [Google Scholar] [CrossRef] [Green Version]
- Cora, D.; Re, A.; Caselle, M.; Bussolino, F. MicroRNA-mediated regulatory circuits: Outlook and perspectives. Phys. Biol. 2017, 14, 045001. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Glucose-stimulated insulin secretion from pancreatic beta cells. Glucose is imported via GLUT1 and phosphorylated by GCK. The increase in cellular respiration leads to an increase in the ATP/ADP ratio and the closure of KATP channels and opening of CaV channels. The entry of Ca2+ ions prompts the fusion of insulin granules with the cell membrane, and this process is facilitated by SNARE complexes. Feedback signals after the return to glucose homeostasis leads to restoration of the membrane potential through the influx of K+ ions through KV channels.
Figure 1.
Glucose-stimulated insulin secretion from pancreatic beta cells. Glucose is imported via GLUT1 and phosphorylated by GCK. The increase in cellular respiration leads to an increase in the ATP/ADP ratio and the closure of KATP channels and opening of CaV channels. The entry of Ca2+ ions prompts the fusion of insulin granules with the cell membrane, and this process is facilitated by SNARE complexes. Feedback signals after the return to glucose homeostasis leads to restoration of the membrane potential through the influx of K+ ions through KV channels.

Figure 2.
Detrimental effects of cholesterol accumulation on GSIS. High intracellular cholesterol has been shown to negatively affect GSIS through many processes including decreased glucose transporter activity, reduced active GCK leading to decreased change in ATP/ADP ratio, less insulin granule fusion partially due to an increased dynamin activity.
Figure 2.
Detrimental effects of cholesterol accumulation on GSIS. High intracellular cholesterol has been shown to negatively affect GSIS through many processes including decreased glucose transporter activity, reduced active GCK leading to decreased change in ATP/ADP ratio, less insulin granule fusion partially due to an increased dynamin activity.

Figure 3.
(A) The KEGG pathways identified through miRPath v3.0 strongly associated with miRNAs involved in lipid metabolism in pancreatic beta cells. All miRNAs included in the analysis were conserved between species with two caveats: mature rno-miR-182 shares 95% sequence similarity with hsa-miR-182-5p and mmu-miR-297b-5p shares 71% sequence similarity with mature hsa-miR-297b. Disease-specific pathways were excluded from the results. Pathways are ranked in order of the most closely linked pathways. (B) Interaction network between miRNAs involved in lipid metabolism in pancreatic beta cells mapped in Cytoscape v3.8.0. The network is annotated so that the size of the pathways labels is proportional to the number of genes in the network associated with the relevant miRNAs and the thickness of the edges shows the contribution of that miRNA as a proportion of the total miRNA–gene interactions in each pathway. Pathways associated with <4 miRNAs were excluded from the analysis to improve clarity of the map.
Figure 3.
(A) The KEGG pathways identified through miRPath v3.0 strongly associated with miRNAs involved in lipid metabolism in pancreatic beta cells. All miRNAs included in the analysis were conserved between species with two caveats: mature rno-miR-182 shares 95% sequence similarity with hsa-miR-182-5p and mmu-miR-297b-5p shares 71% sequence similarity with mature hsa-miR-297b. Disease-specific pathways were excluded from the results. Pathways are ranked in order of the most closely linked pathways. (B) Interaction network between miRNAs involved in lipid metabolism in pancreatic beta cells mapped in Cytoscape v3.8.0. The network is annotated so that the size of the pathways labels is proportional to the number of genes in the network associated with the relevant miRNAs and the thickness of the edges shows the contribution of that miRNA as a proportion of the total miRNA–gene interactions in each pathway. Pathways associated with <4 miRNAs were excluded from the analysis to improve clarity of the map.


{kind=link}
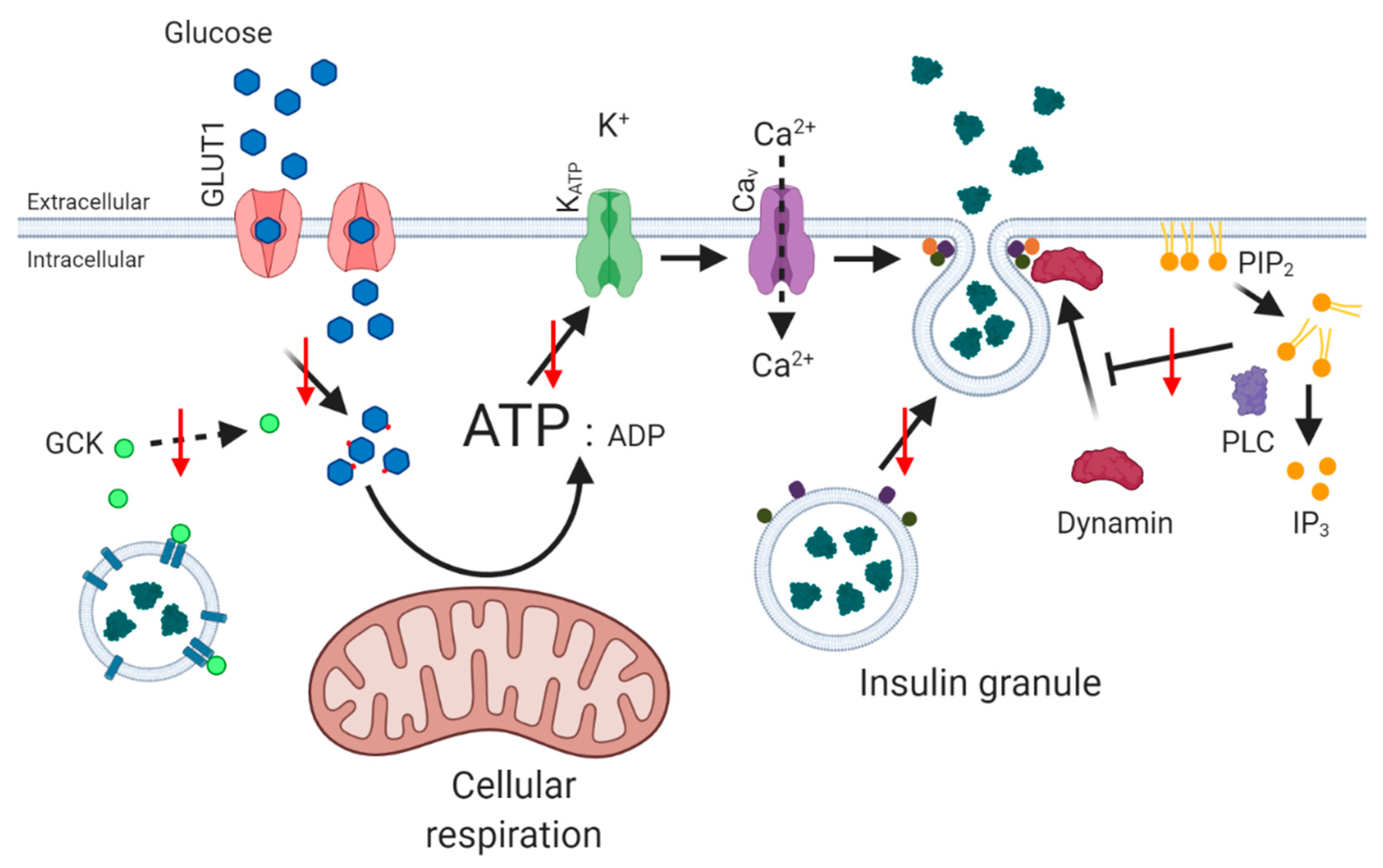{kind=link}
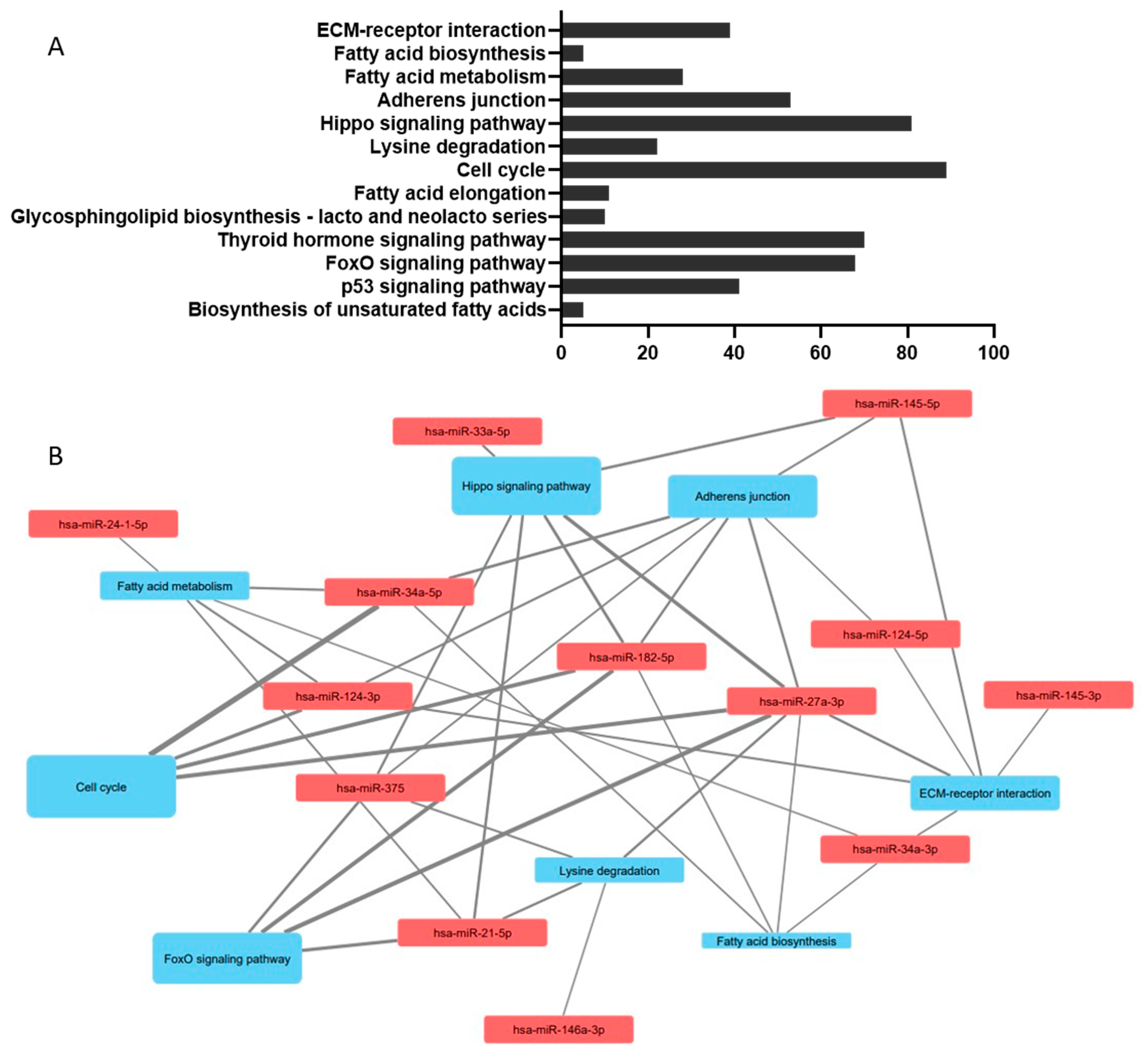{kind=link}
Table 1.
MicroRNA sequences altered in response to metabolic challenges in islets and beta cells. Upregulation (↑) or downregulation (↓) is noted next to the microRNAs.
Table 1.
MicroRNA sequences altered in response to metabolic challenges in islets and beta cells. Upregulation (↑) or downregulation (↓) is noted next to the microRNAs.
| MicroRNA (↑↓) | Cell/Tissue | Test Condition | Target | Outcomes | Reference |
|---|---|---|---|---|---|
| miR-21-5p (↑) | Human PANC hybrid 1.1B4 pancreatic beta cell line | High-density lipoprotein (HDL) | Signal transducer and activator of transcription 3 (STAT3) and decapentapegic protein 3 (SMAD7) | HDL protects against glucolipotoxicity (GLT); miR-21-5p mimic, replicates HDL repression of SMAD7 and STAT3, but does not protect against GLT. | Tarlton et al. (2021) [202] |
| miR-24 (↑) | Murine MIN6 pancreatic beta cell line | Cholesterol exposure | Transcription factor specificity factor 1 (Sp1) and Ca2+ sensor, secretagogin (Scgn) | Impaired Scgn-mediated phosphorylation of focal adhesion kinase and paxillin; reductions in focal adhesions in insulin granules and insulin secretion. | Yang et al. (2019) [204] |
| mir-27a (↑) | Rat INS-1 beta cell line | Cholesterol exposure | ATP-binding cassette transporter A1 (ABCA1) | GLP-1 ↓miR-27a, and increases expression of ABCA1: protects against cholesterol-induced lipotoxicity. | Yao et al. (2017) [180] |
| miR-33a (↑) | Islets isolated from mice with beta cell expression of human islet amyloid polypeptide (IAPP) | Cholesterol exposure | ATP-binding cassette transporter A1 (ABCA1) | ↑miR-33 increases IAPP deposition; knockout of beta cell ABCA1 in hIAPP +/− mice impairs glucose tolerance, insulin secretion, induces hyperglycaemia. | Wijesekara et al. (2016) [181] |
| miR-34a (↑) | Murine MIN6B1 beta cell line Pancreatic islets (db/db) mice | Palmitate exposure | Tumour protein p53 Vesicle-associated membrane protein 2 (VAMP-2) | ↑ miR-34a causes sensitisation to apoptosis, and reduces insulin secretion. Anti-miR-34a oligonucleotide partially protection against apoptosis. | Lovis et al. (2008) [140] |
| miR-34a (↑↓) | Rat INS-1 islet beta cell line | Glucagon-like peptide-1 (GLP-1) and palmitate exposure | Sirtuin 1 | Palmitate exposure ↑ miR-34a; GLP-1 ↓ miR-34a; miR-34a mimics enhances palmitate lipotoxicity; inhibitors achieve the reverse. | Han et al. (2012) [141] |
| miR-34a (↑) | Murine MIN6B1 pancreatic beta cell line | Palmitate exposure | B-cell lymphoma cell-2 (Bcl-2) | MiR-34a interacts directly with Bcl-2; mir-34a mimic promotes lipoapoptosis; inhibitor achieves the reverse. | Lin et al. (2014) [142] |
| miR-34a-5p (↑) | Murine islets and rat INS-1 insulinoma cells | Stearic acid | B-cell lymphoma cell-2 (Bcl-2) | Lipotoxicity reduced by inhibitors of miR-34a-5p; protein kinase-like endoplasmic reticulum kinase (PERK) and p53 mediate stearic acid elevation of miR-34a-5p. | Lu et al. (2016) [143] |
| miR-34a (↑) | Rat INS-1 islet beta cell line | Exposure to ‘high’ glucose and palmitate (GLT) | Sirtuin 1 (SIRT1) | Long non-coding (lnc)RNA, NONRATT003679.2, ‘sponges’ miR-34a and reduces cell injury due to GLT. | Kong et al. (2019) [144] |
| miR-145 (↓) | Murine pancreatic islets | Elevated glucose | ATP-binding cassette transporter A1 (ABCA1) | ↑miR-145 increases total islet cholesterol, and decreases GSIS; inhibitors achieve the reverse. | Kang et al. (2013) [200] |
| miR-146a (↑) | Murine MIN6B1 pancreatic beta cell line Pancreatic islets (db/db) mice | Palmitate exposure | - | ↑ miR-146a induces sensitisation to apoptosis. Anti-mir-146a oligonucleotide provides partial protection against apoptosis. | Lovis et al. (2008) [140] |
| miR-182-5p (↑) | Rat INS-1 beta cell line | Palmitate exposure | Thrombospondin 1 (THBS-1) | Mimic of mir-182-5p decreased viability and increases lipotoxicity due to palmitate; inhibitor achieves the reverse. | Liu et al. (2018) [149] |
| miR-297b-5p (↓) | Murine TC6 beta cell line. Islets derived from C57BL/6 mice | Stearic acid or palmitic acid exposure | Large-tumour-suppressor kinase 2 (LATS2) | Upregulation of miR-297b-5p protects against stearic acid-induced apoptosis and loss of insulin secretion. | Guo et al. (2020) [150] |
| miR-297b-5p (↓) | Murine TC6 beta cell line | Stearic acid exposure | - | - | Yu et al. (2020) [151] |
| miR-375 (↑) | Murine TC6 beta cell line | Stearic acid exposure | Yu et al. (2020) [151] | ||
| miR-375 (↑) | Murine NIT-1 cells | Palmitate exposure | Myotrophin (V1) protein | ↑ miR-375 increases susceptibility to palmitate-induced lipoapoptosis; knockdown of endogenous pri-miR-375 protects against lipoapoptosis | Li et al. (2010) [152] |
| miR-375-3p | Human islets, INS-1 and MIN6 bet cell lines | HDL | - | Export of miR-375-3p to HDL correlates inversely with insulin secretion. | Sedgeman et al. (2019) [203] |
| miR-383 | Murine MIN6 beta cells | Exposure to ‘high’ glucose | Toll-like receptor 4 (TLR4) ApoC3 | Overexpression of miR-383 inhibits glucose-dependent apoptosis and oxidative stress. | Cheng et al. (2020) [201] |
| miR-3074-5p (↓) | Rat INS-2 cells and murine islets | Palmitate exposure | cAMP-responsible element-binding protein (CREB) | Long non-coding RNA lncEif4g2 ↓miR-3074-5p, decreases CREB, enhances nuclear factor erythroid 2-related factor 2 (Nrf2) and protects against lipotoxicity. | Wang et al. (2020) [153] |
| miR-7222-3p (↑) | Murine MIN6 pancreatic beta cell lines | Palmitate exposure | Acyl CoA: Cholesterol Acyltransferase (SOAT1) | Circular RNA circ-Tulp4 promotes beta cell function by sponging miR-7222-3p and regulating SOAT1. | Wu et al. (2020) [179] |
Table 2.
Circulating biomarkers of diabetes, cross-referenced to microRNA sequences modified by metabolic challenges in beta cells. Upregulation (↑) or downregulation (↓) is noted next to the microRNAs.
Table 2.
Circulating biomarkers of diabetes, cross-referenced to microRNA sequences modified by metabolic challenges in beta cells. Upregulation (↑) or downregulation (↓) is noted next to the microRNAs.
| MicroRNA in T2D (↑↓) | Study Design | Subjects | Groups | Specific Association Tested | Reference |
|---|---|---|---|---|---|
| 39 regulated miRs identified: hsa-miR-34a (↓) hsa-miR-182 (↑) | Measured levels of 170 miRNAs in plasma | Women with obesity and sex-matched controls | T2D (15), insulin resistance (IR) (19), insulin sensitivity (IS) (12) and controls (12) | T2D patients with obesity compared to controls | Jones et al. (2017) [213] |
| 9 regulated miRs identified: no matches with Table 1 | Whole peripheral blood: RNA sequencing prior to weighted gene coexpression network analysis (WGCNA) | 40–60-year-old men | Newly diagnosed T2D (3) and control (3) | T2D patients compared to controls | Feng et al. (2019) [214] |
| 6 regulated miRs identified: no matches with Table 1 | Measured 179 microRNA sequences in exosomes isolated from serum | Men matched for age and BMI | Control (NGT) (4) and T2D (4) | T2D patients compared to controls | Katayama et al. (2018) [215] |
| 30 regulated miRs identified, including hsa-miR-21 hsa-miR-24 | Measured 754 small noncoding RNAs in pooled samples isolated from serum | Subjects selected from the Bruneck study | Diabetes patients (10) and age, sex and risk factor profile-matched controls (30) | T2D patients compared to controls | Zampetaki et al. (2010) [216] |
| 4 regulated miRs identified, including hsa-miR-375 (↓) | Four miRNAs measured | Subjects initially non-T2D at start of CORDIOPREV (cardiovascular disease) study | Controls (78), prediabetic (223), incident prediabetic (30) and incident T2D (107) | T2D patients at risk of CVD compared to controls at risk of CVD | Jiménez-Lucena, Carmago et al. (2018) [217] |
| 82 regulated miRs identified, including hsa-miR-27a (↑) hsa-miR-146a (↓) hsa-miR-182 (↓) hsa-miR-375 (↑) | MicroRNAs from miRbase 11.0 were measured from whole blood samples | Men with minimal differences in their clinical characteristics between the groups | Controls (7), impaired fasting glucose (IFG) (6) and T2D (8) | T2D patients compared to Controls | Karolina et al. (2011) [218] |
| hsa-miR-146a (↓) in males hsa-miR-146a (↑) in females | Circulating miR-146a levels in serum | Subjects recruited from the Italian National Research Center on Aging (INRCA) | Controls (188) and T2D patients (144) | T2D patients compared to controls | Mensà et al. 2019 [219] |
| No match with Table 1 | Circulating miR-135a levels in plasma | Case–control study; T2D patients were treatment naive | Controls (40), prediabetic patients (40) and T2D patients (40) | T2D patients compared to controls | Monfared et al. (2020) [220] |
| No match with Table 1 | Circulating miR-30c | Patients that had previously undergone coronary artery angiography | Control (32), coronary heart disease (CHD) (34), non-complicated T2D (47) and CHD + T2D (27) | T2D patients compared to controls | Luo et al. (2019) [221] |
| No match with Table 1 | Circulating miR-103a and miR-103b | Patients that had previously been assessed at the Department of Endocrinology, Alliliated Hospital of Southwest Medical University, Luzhou, Sichuan, China | Controls (50), prediabetes (47) and T2D (48). | T2D patients compared to controls | Luo et al. (2020) [222] |
| No match with Table 1 | Measured 179 miRNAs most highly abundant in human serum/ plasma in microvesicle isolations | Patients selected from primary health care on the Evolution of Patients with Prediabetes (PREDAPS) study. No patients had T2D at the start of the study | Control (8), fatty liver (8), prediabetes (8) and T2D (7) | T2D patients compared to controls | Parrizas et al. (2020) [223] |
| Four regulated miRs identified: no matches with Table 1 | PBMC sample RNA sequencing for bioinformatic analysis | Recruited from Outpatient Clinics of Division of Endocrinology, Ribeirão Preto Medical School, University of São Paulo and of the São Paulo Federal University, Brazil. Controls were matched to patients | Controls (40), T1D (31) and T2D (32) | T2D patients compared to controls | Massaro et al. (2019) [224] |
| No match with Table 1 | Abundance of miR-128 was measured in serum | Recruited from the Chennai Urban Rural Epidemiology Study (CURES) | Control (40), depression patients (40), T2D (40) and T2D with depression (40) | T2D patients compared to controls | Prabu et al. (2020) [225] |
| No match with Table 1 | Measured 325 miRNAs expression in plasma samples | Patients admitted in Lanzhou University Second Hospital. Controls were matched to patients | Control (40), T2D patients with norm-albuminuria (40) and T2D patients with albuminuria (66) | T2D patients compared to controls | Wang et al. (2019) [226] |
| Four regulated miRs identified: No matches with Table 1 | Profiled 752 miRs isolated from PBMCs | Patients recruited with abdominal aortic aneurysm (AAA) | Control (3) and T2D (3) | T2D patients with AAA compared to controls with AAA | Lareyre et al. (2019) [227] |
| Two regulated miRs identified: hsa-miR-146a (↓) hsa-miR-375 (↓) | Circulating miR-146a, miR-34a and miR-375 in serum | Cross-sectional study conducted in Mexico | Control (35), prediabetes (16), T2D (54), T2D with nephropathy (18), T2D with diabetic foot (3) and T2D with CVD | T2D patients with diabetic foot or nephropathy compared to controls | Garciá-Jacobo et al. (2019) [228] |
| Six regulated miRs identified, including hsa-miR-24-3p (↓) hsa-miR-34a-5p (↓) | Measured 14 miRNAs predicted to target diabetes susceptibility genes from peripheral blood | Study conducted in Greece | Controls (37) and T2D (40) | T2D patients compared to controls | Kokkinopoulou et al, (2019) [229] |
| Three regulated miRs identified, including hsa-miR-24-3p (↓) | Measured 84 T2D-related miRNAs in peripheral blood | Study conducted in Greece | Controls (37) and T2D (40) | T2D patients compared to controls | Avgeris et al. (2020) [230] |
| Eight regulated miRs identified: no matches to Table 1 | Circulating microRNA sequenced from plasma samples | Prediabetic subjects selected from METabolic Syndrome In Men (METSIM) study. Subjects were matched between groups | Prediabetic patients that did not develop T2D (145). Prediabetic patients that developed T2D (145) | Prediabetic patients that developed T2D compared to prediabetic patients that did not develop T2D | Ghai et al. (2019) [231] |
| Three regulated miRs identified: no matches to Table 1 | Measured 179 miRNAs from plasma samples | All subjects selected from Diabetes Prediction and Screening Observational Study (DIAPASON) | Control (9), impaired glucose tolerance (IGT) (9) and T2D (9) | T2D patients compared to controls | De Candia et al. (2017) [232] |
| Seven regulated miRs identified: no matches with Table 1 | Measured 372 mature miRNAs from serum samples | Subjects recruited from First Affiliated Hospital of Guangzhou University of Chinese Medicine | Control (5) and T2D (10) | T2D patients compared to Controls | Yang et al. (2017) [233] |
| No match with Table 1 | Measured let-7b-5p in serum samples | Subjects were recruited from the All-New Diabetics in harjah and Ajman (ANDISA) study | Control (25) and T2D without complications (29) and T2D patients with complications (27) | T2D patients without complications compared to controls | Aljaibeji et al. (2020) [234] |
| No match with Table 1 | Expression of miR-20b and miR-17-3p were measured in serum samples | Subjects were selected from Internal Medicine and Opthalmology departments of Fayoum University, Fayoum, Egypt | Control (81), non-diabetic retinopathy (30), diabetic retinopathy (50) | Diabetic retinopathy patients compared to non-diabetic retinopathy patients | Shaker et al. (2019) [235] |
| No match with Table 1 | MicroRNA-126 was measured in serum samples | Subjects recruited from Department of Endocrinology in the Hospital of Harbin Medical University, Harbin, China | Control (138), IGT (82), IFG (75) and T2D (60) | T2D patients compared to controls | Liu et al. (2014) [236] |
| 36 regulated miRs identified, including hsa-miR-182-5p (↓) | Circulating miRNAs in plasma samples were sequenced | Participants selected from the Japanese American Community Diabetes Study (JACDS). Controls were matched to patients | Control (5), T2D patients (5) | T2D patients compared to controls | Wander et al. (2020) [237] |
| No match with Table 1 | Measured miR-7 in serum samples | T2D patients that were newly diagnosed or during drug withdrawal. Controls were matched to patients | Controls (74) and T2D (152) | T2D patients compared to controls | Wan et al. (2017) [238] |
| Match with Table 1: hsa-miR-146a (↑) | Measured miR-146a and miR-155 in gingival crevicular fluid | Study conducted in Belgrade | Control (24), chronic periodontitis (24), peridontally healthy with T2D (24) and T2D patients with chronic periodontitis (24) | T2D patients compared to controls | Radović et al. (2018) [239] |
| Seven regulated miRs identified: no match with Table 1 | Measured 372 miRNAs in serum samples | Subjects were selected from Vorarlberg Institute for Vascular Investigation and Treatment (VIVIT) cohort | Controls (43), IFG (43) and T2D (43) | T2D patients compared to controls | Jaeger et al. (2018) [240] |
| Match with Table 1: hsa-miR-146a (↓) | Measured miR-146a in PBMC and plasma samples | Study conducted in Tehran | Controls (30) and T2D (30) | T2D patients compared to controls | Alipoor et al. (2018) [241] |
| Two regulated miRs identified: no match with Table 1 | Measured miRNA-9 and miRNA-370 | Subjects were recruited from Al-Qasr AlEiny Teaching hospitals, Cairo, Egypt | Controls (50), T2D (50), CVD (50) and T2D with CVD (50) | T2D patients compared to controls | Motawae et al. (2015) [242] |
| hsa-miR-375 (↑) | Measured mature miRNA-375 | Subjects were recruited from Departments of Endocrinology and Metabolism Shihezi University School of Medicine, China | Control (100) and T2D (100) | T2D patients compared to controls | Yin et al.(2017) [243] |
| 34 regulated miRs identified, including hsa-miR-21-5p (↑) hsa-miR-146a-5p (↑) | All small RNA sequenced in corneas | Subject tissues were obtained from the National Disease Research Interchange (NDRI) | Control (10) and T2D (12) | T2D patients compared to controls | Kulkarni et al. (2017) [244] |
| Ten regulated miRs identified, including hsa-miR-146a (↓) | Circulating RNA sequenced from serum samples | Subjects recruited from the Diabetes Specialities Centre, Department of Endocrinology, Zhejiang Provincial Hospital or TCM, Hangzhou, China | Control (5) and T2D (5) | T2D patients compared to controls | Yang et al. (2014) [245] |
| Six regulated miRs identified: no matches with Table 1 | Measured 24 miNAs selected through literature | Subjects selected from CORDIOPREV study | Control (355) and T2D (107) | T2D patients compared to Controls | Jiménez-Lucena, Rangel-Zúñiga et al. (2018) [246] |
| Four regulated miRs identified, including hsa-miR-145 (↓) | Measured miRNAs previously identified with a link to diet and developing T2D | Subjects selected from CORDIOPREV study comparing low-fat high-complex-carbohydrate (LFHCC) diet and Mediterranean diet | Control (355) and T2D (107) | T2D patients on LFHCC diet compared to controls on LFHCC diet | Jiménez-Lucena et al. (2021) [247] |
| No match with Table 1 | Measured miR-103b in serum samples | Patients that had previously been assessed at the Department of Endocrinology, Alliliated Hospital of Southwest Medical University, Luzhou, Sichuan, China | Control (46), prediabetes (48), non-complicated T2D (43) and T2D and coronary heart disease (CHD) (36) | Non-complicated T2D patients compared to controls | Luo et al. (2015) [248] |
| No match with Table 1 | Measured miR-15a in peripheral blood samples | Subjects recruited from King Abdullah University Medical Centre, Bahrain | Control (24), prediabetes (22) and T2D (24) | T2D patients compared to controls | Al-Kafaji et al. (2015) [249] |
| No match with Table 1 | Exiqon qPCR panels used on serum samples | Study was conducted in Guangzhou, China | Control (25), T2D (25), obese (25) and T2D and obese (25) | T2D patients compared to controls | Wu et al. (2015) [250] |
| No match with Table 1 | Measured miR-18a and miR-34c in PBMC samples | Subjects recruited from Beijing Xuanwu Hospital, Capital Medical University | Control (105), IFG (74) and T2D (117) | T2D patients compared to controls | Wang et al. (2017) [251] |
| One regulated miR detected: no match with Table 1 | Measured 47 circulating miRNAs selected from pilot study in serum samples | Subjects were selected from the fasting cohort of the Malmö Diet and Cancer cardiovascular cohort (MDC-CC) | Control (259), CVD (169) and T2D (140) | T2D patients compared to Controls | Gallo et al. (2018) [252] |
| Regulated miR matches with Table 1: hsa-miR-375 (↑) | Measured miR-375 in serum samples | Subjects were selected from the First Affiliated Hospital of Shihezi University School of Medicine, Shihezi, Xinjiang, China | Controls (100) and T2D (100) | T2D patients compared to controls | Sun et al. (2014) [253] |
| No match to Table 1 | Measured miR-155 in serum samples | Subjects were recruited from the Diabetes clinic of the Diabetes Research Centre, Endocrinology and Metabolism Research Institute, Tehran University of Medical Sciences | Controls (42) and T2D (45) | T2D patients compared to controls | Mahdavi et al. (2018) [254] |
| One regulated miR identified: no match to Table 1 | Measured five miRNAs associated with T2D in serum samples | Subjects recruited from Outpatient Department if Laboratory Medicine, Chronic Disease Hospital of Nanshan District, Shenzhen, China | Control (3), prediabetes (30) and T2D (30) | T2D patients compared to controls | Zhang et al. (2013) [255] |
| Two regulated miRs identified, including hsa-miR-21-5p (↓) | Measured miR-21-5p and mR-126-3p in serum samples | All subjects reported a Mediterranean diet | Controls (107), T2D with no Complications (76) and T2D with complications (117) | T2D patients compared to controls | Olivieri et al. (2015) [256] |
| Nine regulated miRs identified, including hsa-miR-21 (↑) hsa-miR-24 (↑) | Measured miRNAs commonly found in plasma | Subjects selected from the impact of Migration and Ethnicity on Diabetes in Malmö (MEDIM) study | Control (119), T2D (33) | T2D patients compared to controls | Wang et al. (2014) [257] |
| No match with Table 1 | Measured microvesicle miR-126 in serum samples | Subjects recruited from the Division of Metabolic Diseases of the University of Padua | Control (53), prediabetes (39) and T2D (68) | T2D patients compared to controls | Gianella et al. (2017) [258] |
| Four regulated miRs identified: no matches with Table 1 | Four miRNAs selected following pilot study. Measured in plasma samples | Subjects recruited from People’s Hospital of Guizhou Privince, Guiyang, China | Control (50), prediabetes (50) and T2D (50) | T2D patients compared to controls | Yan et al. (2016) [259] |
| No match with Table 1 | Measured miR-31, miR-93, miR0199a and miR-146a in plasma samples | Subjects recruited from the Geriatric Endocrinology Department of PLA General Hospital | Control (64), prediabetes (64) and T2D (64) | T2D patients compared to controls | Yan et al. (2014) [260] |
| Four regulated miRs detected including: hsa-miR-21 (↑) hsa-miR-24 (↑) hsa-miR-34a (↑) | Assessed a Diabetes-related human miRNA panel in serum samples | Subjects selected from the Outcome Reduction with Initial Glargine Intervention (ORIGIN) trial | Control (20), prediabetes (21) and T2D (17) | T2D patients compared to controls | Nunez Lopez et al. (2017) [261] |
| Two regulated miRs detected: no match with Table 1 | Measured miR-144 and miR-223 in platelets and plasma samples | Subjects recruited at the Department of Endocrinology, Xiangya Hospital | Controls (30), T2D (56) and T2D with acute ischemic stroke | T2D patients compared to controls | Yang et al. (2016) [262] |
| No match with Table 1 | Measured miRNAs that had been previously associated with T2D in plasma samples | Subjects recruited at the Outpatient Department of Laboratory Medicine, Chronic Disease Hospital of NanShan District, Shenzhen, China | Controls (20) and T2D (20) | T2D patients compared to controls | Zhang et al. (2015) [263] |
| Four regulated miRs identified: no match with Table 1 | Serum miRNAs included in Exiqon Human panel 1 | Subjects were recruited from Diabetes Specialities Centre, Chennai, India | Control (49), IGT (47) and T2D (49) | T2D patients compared to controls | Prabu et al. (2015) [264] |
| One regulated miR identified: hsa-miR-146a (↓) | Measured miR-155 and miR-146a in serum samples | Subjects were recruited in Quito, Ecuador | Control (40) and T2D (56) | T2D patients compared to controls | Baldeón et al. (2014) [265] |
| Match with Table 1: hsa-miR-146a (↓) | Measured miR-146a in PBMC samples | Subjects were recruited from Diabetes Specialities Centre, Chennai, India | Controls (35) and T2D (35) | T2D patients compared to controls | Lenin et al. (2015) [266] |
| No match with Table 1 | Measured miR-126 in serum samples | Subjects recruited from Medical Biochemistry and Internal Medicine Departments of Zagazig University Hospitals, Egypt | Controls (100), IGT (86) and T2D (100) | T2D patients compared to controls | Rezk et al. (2016) [267] |
| Ten regulated miRs identified: no matches with Table 1 | Assessed miRNAs in TaqMan Low-Density Arrays using serum samples | Subjects recruited in the Endocrinology Service of the Hospital Universitari Dr. Josep Trueta, Girona, Spain | Controls (45) and T2D (48) | T2D patients compared to controls | Ortega et al. (2014) [268] |
| Two regulated miRs detected: no matches with Table 1 | Assessed miRNAs in circulating microparticles | Study conducted in Germany | Control (80) and T2D (55) | T2D patients compared to controls | Jansen et al. (2016) [269] |
| Four regulated miRs detected including: hsa-miR-21 (↑) hsa-miR-34a (↑) | Measured 28 pancreas-enriched miRNAs in plasma samples | Subjects were recruited from Florida, USA | Control (27), prediabetes (12), T2D (31), latent autoimmune diabetes in adults (LADA) (6) and T1D (16) | T2D patients compared to controls | Seyhan et al. (2016) [270] |
| Five regulated miRs identified: no matches with Table 1 | Assessed miRNAs in TaqMan Low-Density Arrays using serum samples | Subjects were recruited from the Department of Endocrinology Jinling Hospital, Nanjing, China | Control (92) and T2D (184) | T2D patients compared to controls | Wang et al. (2016) [271] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tarlton, J.M.R.; Patterson, S.; Graham, A. MicroRNA Sequences Modulated by Beta Cell Lipid Metabolism: Implications for Type 2 Diabetes Mellitus. Biology 2021, 10, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060534
AMA Style
Tarlton JMR, Patterson S, Graham A. MicroRNA Sequences Modulated by Beta Cell Lipid Metabolism: Implications for Type 2 Diabetes Mellitus. Biology. 2021; 10(6):534. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060534
Chicago/Turabian StyleTarlton, Jamie M. R., Steven Patterson, and Annette Graham. 2021. "MicroRNA Sequences Modulated by Beta Cell Lipid Metabolism: Implications for Type 2 Diabetes Mellitus" Biology 10, no. 6: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060534
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.