
Comparison of Diazotrophic Composition and Distribution in the South China Sea and the Western Pacific Ocean


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Physicochemical Analysis
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Quality Control and Analysis of Sequencing Data
2.5. Quantification of Main Cyanobacterial nifH Phylotypes
2.6. Analysis of Community Composition and Diversity
2.7. Statistical Analysis of Community Composition, Environmental Variables, and Geographical Distance
3. Results
3.1. Environmental Conditions
3.2. Sequencing Statistics and Diversity Estimates
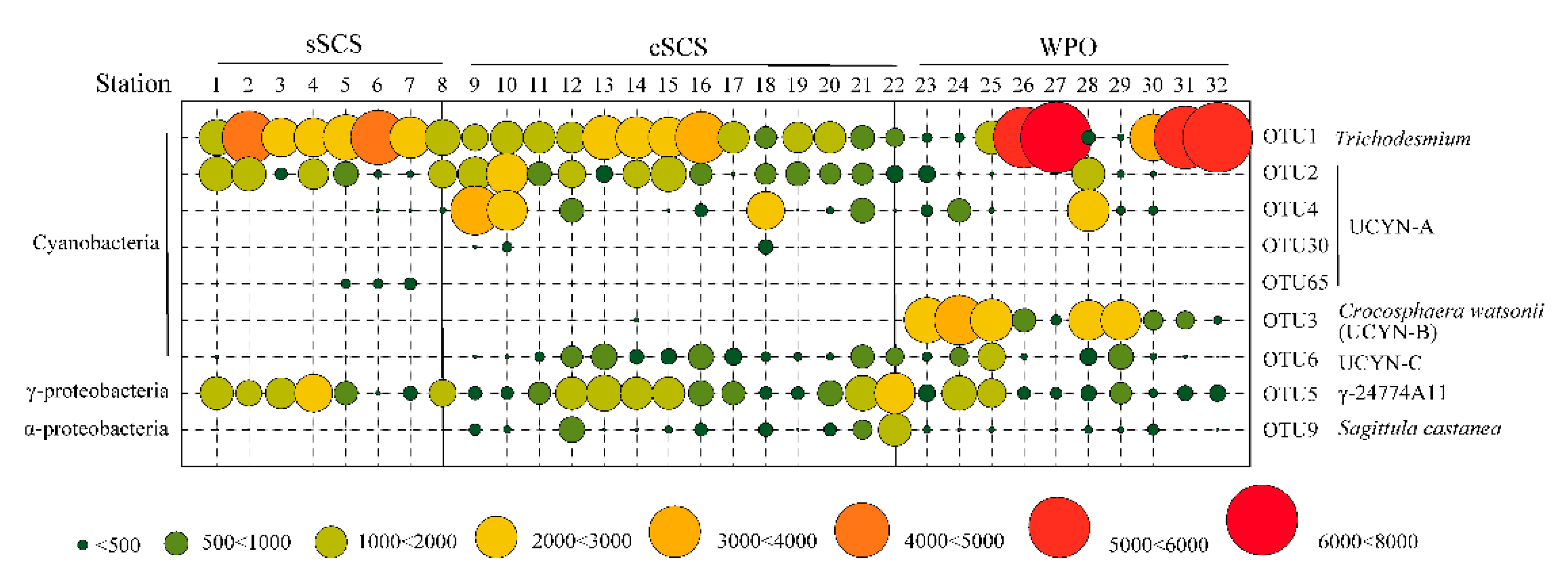
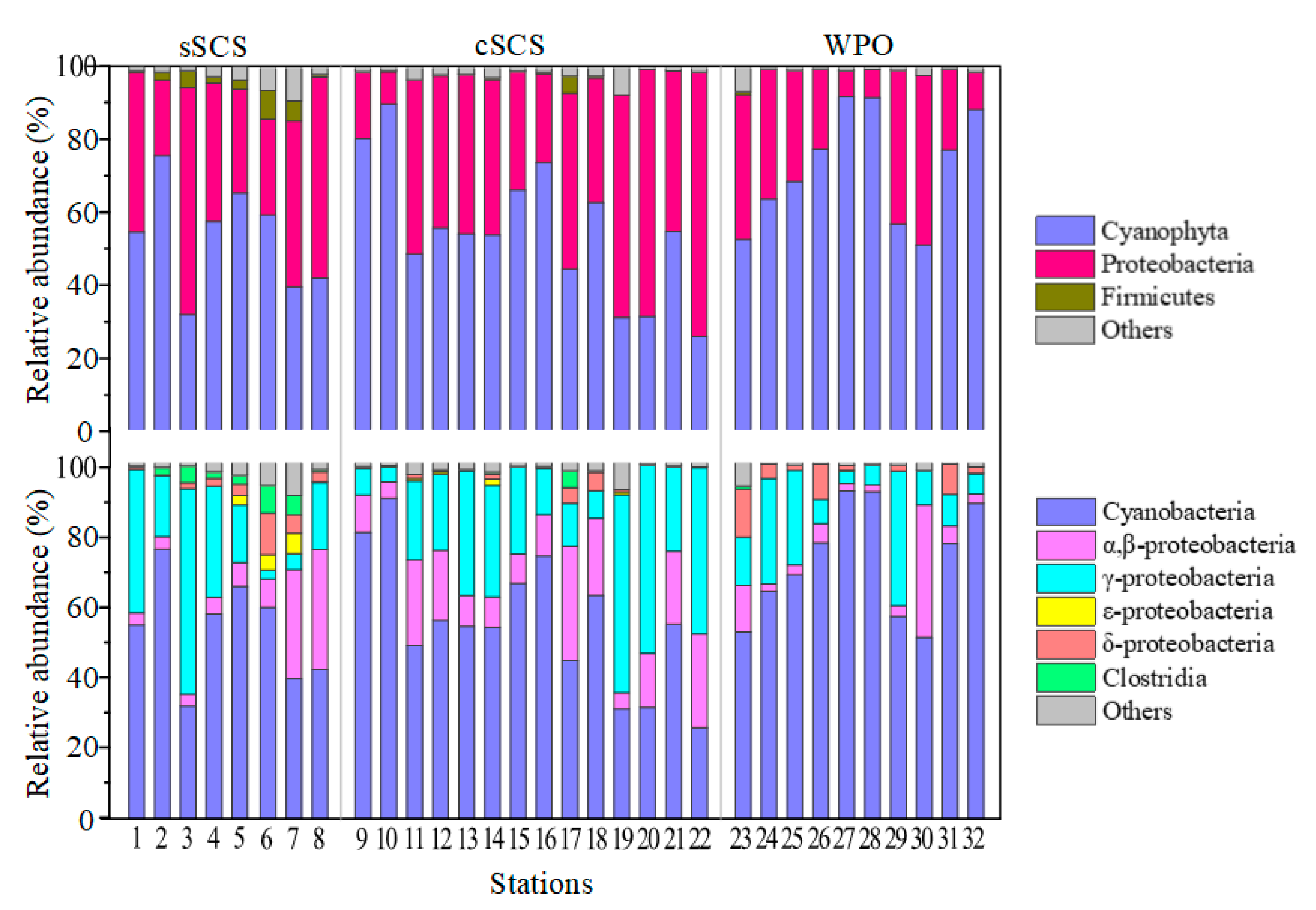
3.3. Phylogeny and Composition of Diazotrophic Communities
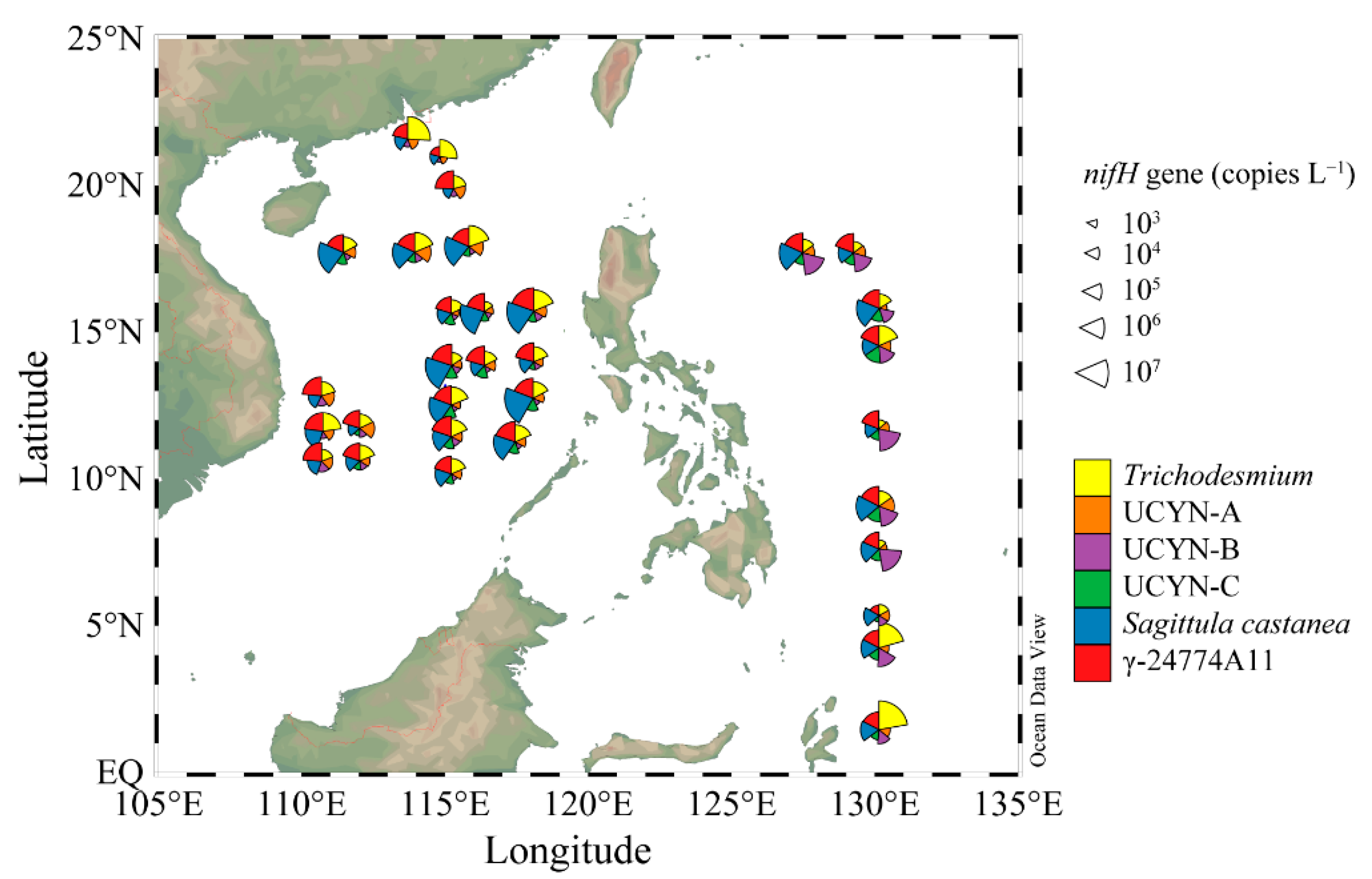
3.4. Quantification of Representative Diazotrophs
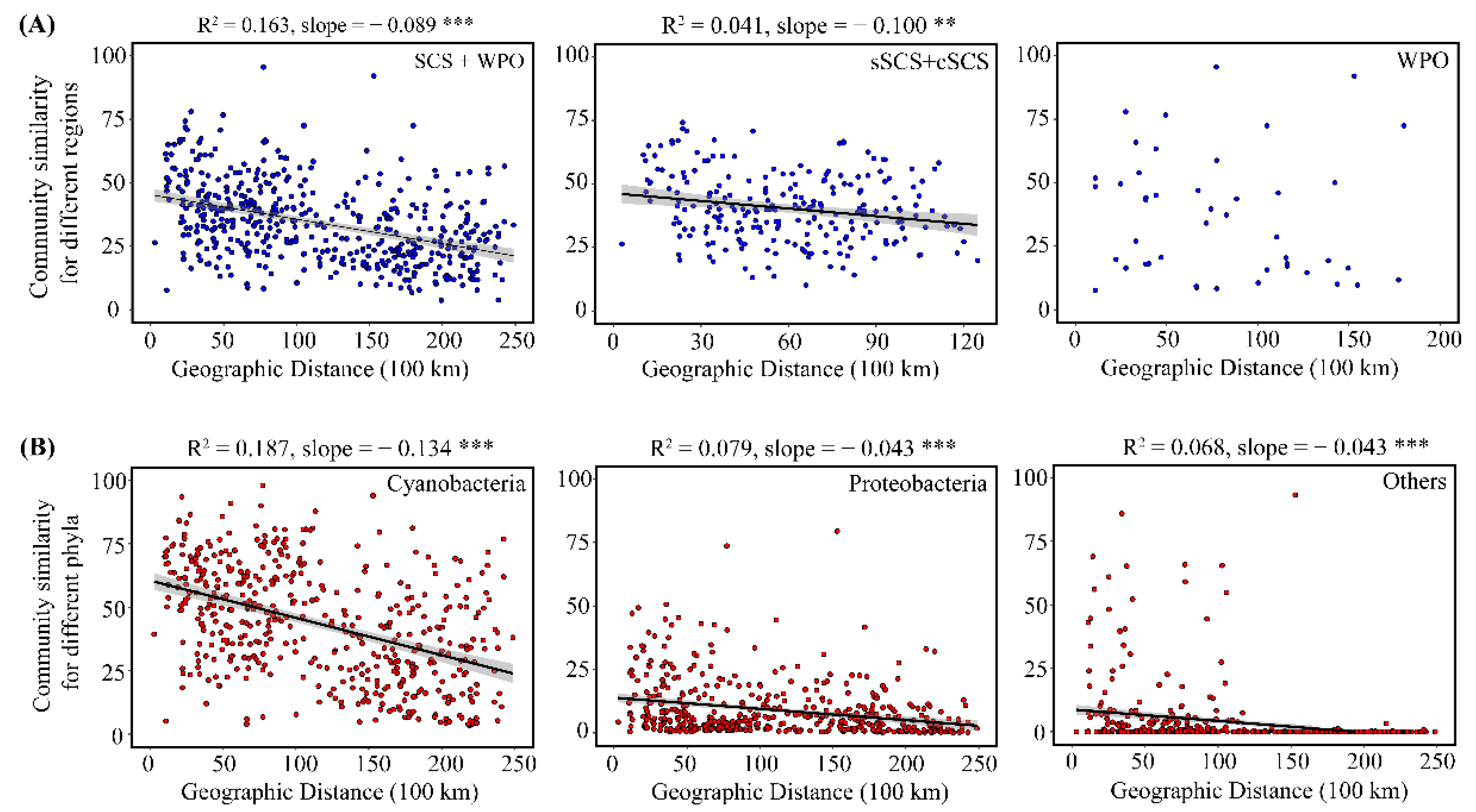
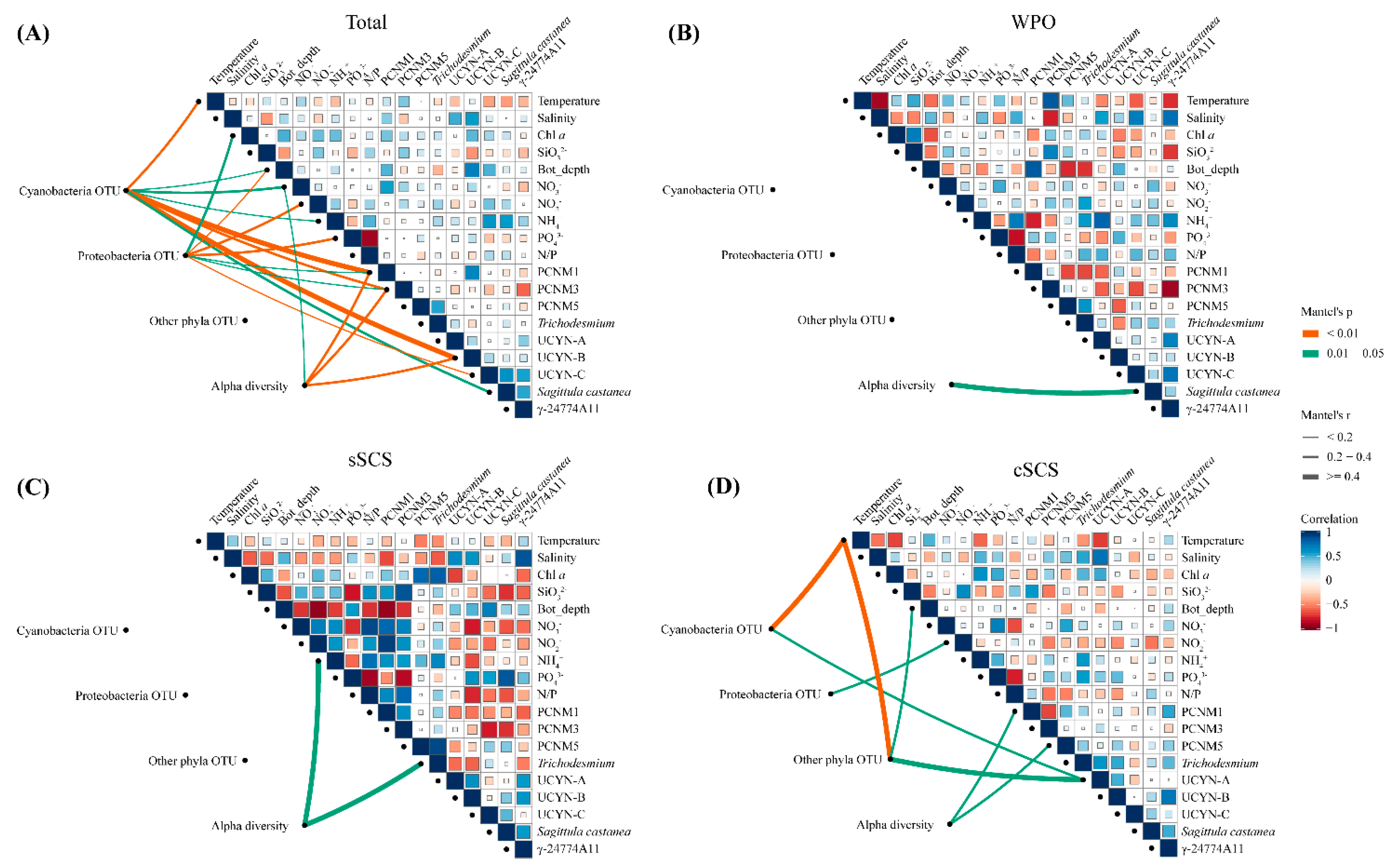
3.5. Diazotrophic Communities in Relation to Spatial and Environmental Variables
4. Discussion
4.1. Role of Diazotrophs in the Nutrient-Replenished Marginal SCS
4.2. Significance of Diazotrophs in the Oligotrophic Oceanic SCS
4.3. Potential Contribution of Diazotrophs in the Pelagic WPO
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





References
- Cao, Z.M.; Yang, W.; Zhao, Y.Y.; Guo, X.H.; Yin, Z.Q.; Du, C.J.; Zhao, H.D.; Dai, M.H. Diagnosis of CO2 dynamics and fluxes in global coastal oceans. Natl. Sci. Rev. 2019, 7, nwz105. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.J.; Liu, Q.Y.; Liu, Z.Y.; Wang, D.X.; Liu, X.B. A general circulation model study of the dynamics of the upper ocean circulation of the South China Sea. J. Geophys. Res. Ocean. 2002, 107, C7. [Google Scholar] [CrossRef]
- Guo, M.X.; Chai, F.; Xiu, P.; Li, S.Y.; Rao, S. Impacts of mesoscale eddies in the South China Sea on biogeochemical cycles. Ocean Dynam. 2015, 65, 1335–1352. [Google Scholar] [CrossRef]
- Cai, W.J.; Dai, M.H.; Wang, Y.C.; Zhai, W.D.; Huang, T.; Chen, S.T.; Zhang, F.; Chen, Z.Z.; Wang, Z.H. The biogeochemistry of inorganic carbon and nutrients in the Pearl River estuary and the adjacent Northern South China Sea. Cont. Shelf. Res. 2004, 24, 1301–1319. [Google Scholar] [CrossRef]
- Voss, M.; Bombar, D.; Loick, N.; Dippner, J.W. Riverine influence on nitrogen fixation in the upwelling region off Vietnam, South China Sea. Geophys. Res. Lett. 2006, 33, L07604. [Google Scholar] [CrossRef]
- Yang, Q.X.; Zhou, L.; Tian, J.W.; Zhao, W. The Roles of Kuroshio Intrusion and Mesoscale Eddy in Upper Mixing in the Northern South China Sea. J. Coast. Res. 2013, 30, 192–198. [Google Scholar]
- Hu, D.; Wu, L.; Cai, W.; Gupta, A.S.; Ganachaud, A.; Qiu, B.; Gordon, A.L.; Lin, X.; Chen, Z.; Hu, S.; et al. Pacific western boundary currents and their roles in climate. Nature 2015, 522, 299–308. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.F.; Chung, S.W.; Wen, L.S.; Liu, K.K.; Chen, Y.L.L.; Chen, H.Y.; Karl, D.M. Dissolved inorganic phosphorus, dissolved iron, and Trichodesmium in the oligotrophic South China Sea. Glob. Biogeochem. Cycles 2003, 17, 1008. [Google Scholar] [CrossRef]
- Karl, D.; Letelier, R.; Tupas, L.; Dore, J.; Christian, J.; Hebel, D. The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean. Nature 1997, 388, 533–538. [Google Scholar] [CrossRef]
- Zehr, J.P.; Capone, D.G. Changing perspectives in marine nitrogen fixation. Science 2020, 368, eaay9514. [Google Scholar] [CrossRef] [PubMed]
- Sohm, J.A.; Subramaniam, A.; Gunderson, T.E.; Carpenter, E.J.; Capone, D.G. Nitrogen fixation by Trichodesmium spp. and unicellular diazotrophs in the North Pacific Subtropical Gyre. J. Geophys. Res. 2011, 116, G03002. [Google Scholar]
- Shiozaki, T.; Bombar, D.; Riemann, L.; Hashihama, F.; Takeda, S.; Yamaguchi, T.; Ehama, M.; Hamasaki, K.; Furuya, K. Basin scale variability of active diazotrophs and nitrogen fixation in the North Pacific, from the tropics to the subarctic Bering Sea. Glob. Biogeochem. Cycles 2017, 31, 996–1009. [Google Scholar] [CrossRef]
- Chen, L.Y.L.; Chen, H.Y.; Lin, Y.H. Distribution and downward flux of Trichodesmium in the South China Sea as influenced by the transport from the Kuroshio Curren. Mar. Ecol. Prog. Ser. 2003, 259, 47–57. [Google Scholar] [CrossRef]
- Chen, L.Y.L.; Chen, H.Y.; Tuo, S.H.; Ohki, K. Seasonal dynamics of new production from Trichodesmium N2 fixation and nitrate uptake in the upstream Kuroshio and South China Sea basin. Limnol. Oceanogr. 2008, 53, 1705–1721. [Google Scholar] [CrossRef] [Green Version]
- Tuo, S.H.; Chen, Y.L.L.; Chen, H.Y. Low nitrate availability promotes diatom diazotroph associations in the marginal seas of the western Pacific. Aquat. Microb. Ecol. 2014, 73, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Farnelid, H.; Andersson, A.F.; Bertilsson, S.; Al-Soud, W.A.; Hansen, L.H.; Sorensen, S.; Steward, G.F.; Hagstrom, A.; Riemann, L. Nitrogenase gene amplicons from global marine surface waters are dominated by genes of non-cyanobacteria. PLoS ONE 2011, 6, e19223. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Mellon, M.T.; Zani, S. New Nitrogen-Fixing Microorganisms Detected in Oligotrophic Oceans by Amplification of Nitrogenase (nifH) Genes. Appl. Environ. Microbiol. 1998, 64, 3444. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Yang, K.; Li, H.; Zhang, H.; Su, J.; Paraskevaidi, M.; Martin, F.; Ren, B.; Zhu, Y. Functional single-cell approach to probing nitrogen-fixing bacteria in soil communities by resonance Raman spectroscopy with 15N2 labeling. Anal. Chem. 2018, 90, 5082–5089. [Google Scholar] [CrossRef]
- Bonnet, S.; Caffin, M.; Berthelot, H.; Grosso, O.; Benavides, M.; Helias-Nunige, S.; Guieu, C.; Stenegren, M.; Foster, R.A. In-depth characterization of diazotroph activity across the western tropical South Pacific hotspot of N2 fixation (OUTPACE cruise). Biogeosciences 2018, 15, 4215–4232. [Google Scholar] [CrossRef] [Green Version]
- Goebel, N.L.; Turk, K.A.; Achilles, K.M.; Paerl, R.; Hewson, I.; Morrison, A.E.; Montoya, J.P.; Edwards, C.A.; Zehr, J.P. Abundance and distribution of major groups of diazotrophic cyanobacteria and their potential contribution to N2 fixation in the tropical Atlantic Ocean. Environ. Microbiol. 2010, 12, 3272–3289. [Google Scholar] [CrossRef]
- Moisander, P.H.; Beinart, R.A.; Hewson, I.; White, A.E.; Johnson, K.S.; Carlson, C.A.; Montoya, J.P.; Zehr, J.P. Unicellular cyanobacterial distributions broaden the oceanic N2 fixation domain. Science 2010, 327, 1512–1514. [Google Scholar] [CrossRef]
- Stenegren, M.; Caputo, A.; Berg, C.; Bonnet, S.; Foster, R.A. Distribution and drivers of symbiotic and free-living diazotrophic cyanobacteria in the western tropical South Pacific. Biogeosciences 2018, 15, 1559–1578. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Kan, J.; Liu, H.; Pujari, L.; Guo, C.; Wang, X.; Sun, J. Heterotrophic Bacteria Dominate the Diazotrophic Community in the Eastern Indian Ocean (EIO) during Pre-Southwest Monsoon. Microb. Ecol. 2019, 78, 804–819. [Google Scholar] [CrossRef]
- Zehr, J.P.; Jenkins, B.D.; Short, S.M.; Steward, G.F. Nitrogenase gene diversity and microbial community structure: A cross-system comparison. Environ. Microbiol. 2003, 5, 539–554. [Google Scholar] [CrossRef]
- Luo, Y.W.; Doney, S.C.; Anderson, L.A.; Benavides, M.; Berman-Frank, I.; Bode, A.; Bonnet, S.; Bostrom, K.H.; Bottjer, D.; Capone, D.G.; et al. Database of diazotrophs in global ocean: Abundance, biomass and nitrogen fixation rates. Earth Syst. Sci. Data 2012, 4, 47–73. [Google Scholar] [CrossRef] [Green Version]
- Xiao, P.; Jiang, Y.G.; Liu, Y.; Tan, W.H.; Li, W.H.; Li, R.H. Re-Evaluation of the diversity and distribution of diazotrophs in the South China Sea by pyrosequencing the nifH gene. Mar. Freshw. Res. 2015, 66, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Grosse, J.; Bombar, D.; Hai, N.D.; Lam, N.N.; Voss, M. The Mekong River plume fuels nitrogen fixation and determines phytoplankton species distribution in the South China Sea during low- and high-discharge season. Limnol. Oceanogr. 2010, 55, 1668–1680. [Google Scholar] [CrossRef]
- Yang, Q.S.; Dong, J.D.; Zhang, Y.Y.; Ling, J.; Wang, D.X.; Wu, M.L.; Jiang, Y.F.; Zhang, Y.Z. Diversity analysis of diazotrophs associated with corals from Xisha and Sanya, South China Sea. Aquat. Ecosyst. Health 2015, 18, 433–442. [Google Scholar] [CrossRef]
- Kong, L.L.; Jing, H.M.; Kataoka, T.; Sun, J.; Liu, H.B. Phylogenetic diversity and spatio-temporal distribution of nitrogenase genes (nifH) in the northern South China Sea. Aquat. Microb. Ecol. 2011, 65, 15–27. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Wen, Z.Z.; Shi, D.L.; Lin, W.F.; Bonnet, S.; Dai, M.H.; Kao, S.J. Biogeography of N2 Fixation Influenced by the Western Boundary Current Intrusion in the South China Sea. J. Geophys. Res. Oceans 2019, 124, 6983–6996. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, Z.; Sun, J.; Jiao, N. Diversity and distribution of diazotrophic communities in the South China Sea deep basin with mesoscale cyclonic eddy perturbations. FEMS Microbiol. Ecol. 2011, 78, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Moisander, P.H.; Beinart, R.A.; Voss, M.; Zehr, J.P. Diversity and abundance of diazotrophic microorganisms in the South China Sea during intermonsoon. ISME J. 2008, 2, 954–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.Y.; Chen, L.Y.L.; Sheu, D.; Chen, H.Y.; Lin, Y.H.; Shiozaki, T. Community and abundance of heterotrophic diazotrophs in the northern South China Sea: Revealing the potential importance of a new alphaproteobacterium in N2 fixation. Deep Sea Res. Part I Oceanogr. Res. Pap. 2019, 143, 104–114. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, H.Y.; Lin, Y.H.; Yong, T.C.; Taniuchi, Y.; Tuo, S.H. The relative contributions of unicellular and filamentous diazotrophs to N2 fixation in the South China Sea and the upstream Kuroshio. Deep Sea Res. Part I Oceanogr. Res. Pap. 2014, 85, 56–71. [Google Scholar] [CrossRef]
- Chen, M.M.; Lu, Y.Y.; Jiao, N.Z.; Tian, J.W.; Kao, S.J.; Zhang, Y. Biogeographic drivers of diazotrophs in the Western Pacific Ocean. Limnol. Oceanogr. 2019, 64, 1403–1421. [Google Scholar] [CrossRef]
- Zehr, J.P. Nitrogen fixation by marine cyanobacteria. Trends Microbiol. 2011, 19, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Bombar, D.; Paerl, R.W.; Riemann, L. Marine Non-cyanobacterial diazotrophs: Moving beyond molecular detection. Trends Microbiol. 2016, 24, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Dippner, J.W.; Nguyen, K.V.; Hein, H.; Ohde, T.; Loick, N. Monsoon-induced upwelling off the Vietnamese coast. Ocean Dynam. 2006, 57, 46–62. [Google Scholar] [CrossRef]
- Du, C.; Liu, Z.; Dai, M.; Kao, S.J.; Cao, Z.; Zhang, Y.; Huang, T.; Wang, L.; Li, Y. Impact of the Kuroshio intrusion on the nutrient inventory in the upper northern South China Sea: Insights from an isopycnal mixing model. Biogeosciences 2013, 10, 6419–6432. [Google Scholar] [CrossRef] [Green Version]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. (Eds.) Methods of Seawater Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 3-527-29589-5. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Sudhir, K.; Glen, S.; Michael, L.; Christina, K.; Koichiro, T. Mega X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eric, G.; Graybill, V. R Development Core Team (2008). In R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; ISBN 3-900051-07-0. [Google Scholar]
- RCRTeam. R: A Language and Environment for Statistical Computing, Version 3.6.1. Vienna, Austria. 2019. Available online: http://www.R-project.org/ (accessed on 5 July 2019).
- Schlitzer, R. Ocean Data View. Available online: https://odv.awi.de (accessed on 2 March 2018).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagne, R.H. Vegan: Community Ecology Package. In R. Package Version p. 2.0-9; Lorenzo Cachón Rodríguez; Available online: http://CRAN.R-project.org/package=vegan (accessed on 1 January 2013).
- Wang, Z.B.; Sun, Y.Y.; Li, Y.; Chen, X.L.; Wang, P.; Ding, H.T.; Chen, B.; Zhang, X.Y.; Song, X.Y.; Wang, M.; et al. Significant Bacterial Distance-Decay Relationship in Continuous, Well-Connected Southern Ocean Surface Water. Microb. Ecol. 2020, 80, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Legendre, P.; Peres-Neto, P.R. Spatial modelling: A comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecol. Model. 2006, 196, 483–493. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, B.; Sun, J.; Li, T.T. Phytoplankton community structure of northern South China Sea in summer of 2014. Haiyangxuebao 2016, 38, 54–65. [Google Scholar]
- Weber, S.C.; Carpenter, E.J.; Coles, V.J.; Yager, P.L.; Goes, J.; Montoya, J.P. Amazon River influence on nitrogen fixation and export production in the western tropical North Atlantic. Limnol. Oceanogr. 2017, 62, 618–631. [Google Scholar] [CrossRef]
- Bombar, D.; Moisander, P.H.; Dippner, J.W.; Foster, R.A.; Voss, M.; Karfeld, B.; Zehr, J.P. Distribution of diazotrophic microorganisms and nifH gene expression in the Mekong River plume during intermonsoon. Mar. Ecol. Prog. Ser. 2011, 424, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Cheung, S.; Suzuki, K.; Saito, H.; Umezawa, Y.; Xia, X.; Liu, H. Highly heterogeneous diazotroph communities in the Kuroshio Current and the Tokara Strait, Japan. PLoS ONE 2017, 12, e0186875. [Google Scholar] [CrossRef] [Green Version]
- Farnelid, H.; Bentzon-Tilia, M.; Andersson, A.F.; Bertilsson, S.; Jost, G.; Labrenz, M.; Jurgens, K.; Riemann, L. Active nitrogen-fixing heterotrophic bacteria at and below the chemocline of the central Baltic Sea. ISME J. 2013, 7, 1413–1423. [Google Scholar] [CrossRef]
- Jing, Z.Y.; Qi, Y.Q.; Du, Y. Upwelling in the continental shelf of northern South China Sea associated with 1997–1998 El Niño. J. Geophys. Res. Oceans 2011, 116, C02033. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, M.; Yang, Q.; Lin, Y.S.; Mao, H.B.; Qiu, Y.S.; Tong, J.L.; Lv, E.; Yang, Z.; Yang, W.F.; et al. Physical-Biological coupling of N2 fixation in the northwestern South China Sea coastal upwelling during summer. Limnol. Oceanogr. 2015, 60, 1411–1425. [Google Scholar] [CrossRef]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Trochodesmium, a Globally Significant Marine Cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Bombar, D.; Dippner, J.W.; Hai, N.D.; Lam, N.N.; Liskow, I.; Loick-Wilde, N.; Voss, M. Sources of new nitrogen in the Vietnamese upwelling region of the South China Sea. J. Geophys. Res. Oceans 2010, 115, C06018. [Google Scholar] [CrossRef] [Green Version]
- Loick, N.; Dippner, J.; Doan, H.N.; Liskow, I.; Voss, M. Pelagic nitrogen dynamics in the Vietnamese upwelling area according to stable nitrogen and carbon isotope data. Deep Sea Res. Pt. I 2007, 54, 596–607. [Google Scholar] [CrossRef]
- Wu, C.; Fu, F.X.; Sun, J.; Thangaraj, S.; Pujari, L. Nitrogen fixation by Trichodesmium and unicellular diazotrophs in the northern South China Sea and the Kuroshio in summer. Sci. Rep. 2018, 8, 2415. [Google Scholar] [CrossRef] [Green Version]
- Bar-Zeev, E.; Avishay, I.; Bidle, K.D.; Berman-Frank, I. Programmed cell death in the marine cyanobacterium Trichodesmium mediates carbon and nitrogen export. ISME J. 2013, 7, 2340–2348. [Google Scholar] [CrossRef]
- Rees, A.P.; Tait, K.; Widdicombe, C.E.; Quartly, G.D.; McEvoy, A.J.; Al-Moosawi, L. Metabolically active, non-nitrogen fixing, Trichodesmium in UK coastal waters during winter. J. Plankton Res. 2016, 38, 673–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inomura, K.; Bragg, J.; Riemann, L.; Follows, M.J. A quantitative model of nitrogen fixation in the presence of ammonium. PLoS ONE 2018, 13, e0208282. [Google Scholar] [CrossRef]
- Subramaniam, A.; Mahaffey, C.; Johns, W.; Mahowald, N. Equatorial upwelling enhances nitrogen fixation in the Atlantic Ocean. Geophys. Res. Lett. 2013, 40, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Bird, C.; Wyman, M. Transcriptionally active heterotrophic diazotrophs are widespread in the upper water column of the Arabian Sea. FEMS Microbiol. Ecol. 2013, 84, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakumar, A.; Al-Rshaidat, M.M.; Ward, B.B.; Mulholland, M.R. Diversity, distribution, and expression of diazotroph nifH genes in oxygen-deficient waters of the Arabian Sea. FEMS Microbiol. Ecol. 2012, 82, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiu, P.; Chai, F. Modeled biogeochemical responses to mesoscale eddies in the South China Sea. J. Geophys. Res. Oceans 2011, 116, C10006. [Google Scholar] [CrossRef] [Green Version]
- Moisander, P.H.; Serros, T.; Paerl, R.W.; Beinart, R.A.; Zehr, J.P. GammaProteobacterial diazotrophs and nifH gene expression in surface waters of the South Pacific Ocean. ISME J. 2014, 8, 1962–1973. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pérez, C.; Mohr, W.; Schwedt, A.; Dürschlag, J.; Callbeck, C.M.; Schunck, H.; Dekaezemacker, J.; Buckner, C.R.T.; Lavik, G.; Fuchs, B.M. Metabolic versatility of a novel N2 -fixing Alphaproteobacterium isolated from a marine oxygen minimum zone. Environ. Microbiol. 2018, 20, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Andersen, K.; Visser, A.; Inomura, K.; Follows, M.; Riemann, L. Quantifying nitrogen fixation by heterotrophic bacteria in sinking marine particles. Nat. Commun. 2020. preprint (Version 1). [Google Scholar] [CrossRef]
- Church, M.J.; Mahaffey, C.; Letelier, R.M.; Lukas, R.; Zehr, J.P.; Karl, D.M. Physical forcing of nitrogen fixation and diazotrophic community structure in the North Pacific subtropical gyre. Glob. Biogeochem. Cycles 2009, 23, GB2020. [Google Scholar] [CrossRef]
- Shiozaki, T.; Kondo, Y.; Yuasa, D.; Takeda, S. Distribution of major diazotrophs in the surface water of the Kuroshio from northeastern Taiwan to south of mainland Japan. J. Plankton Res. 2018, 40, 407–419. [Google Scholar] [CrossRef]
- Bonnet, S.; Rodier, M.; Turk-Kubo, K.A.; Germineaud, C.; Menkes, C.; Ganachaud, A.; Cravatte, S.; Raimbault, P.; Campbell, E.; Quéroué, F.; et al. Contrasted geographical distribution of N2 fixation rates and nifH phylotypes in the Coral and Solomon Seas (southwestern Pacific) during austral winter conditions. Glob. Biogeochem. Cycles 2015, 29, 1874–1892. [Google Scholar] [CrossRef] [Green Version]
- Shiozaki, T.; Kodama, T.; Kitajima, S.; Sato, M.; Furuya, K. Advective transport of diazotrophs and importance of their nitrogen fixation on new and primary production in the western Pacific warm pool. Limnol. Oceanogr. 2013, 58, 49–60. [Google Scholar] [CrossRef]
- Inomura, K.; Wilson, S.T.; Deutsch, C. Mechanistic model for the coexistence of nitrogen fixation and photosynthesis in marine Trichodesmium. Systems 2019, 4, e00210-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | Forward Primer (5′–3′) | Probe | Reverse Primer (5′–3′) | Reference |
|---|---|---|---|---|
| Trichodesmium | GACGAAGTATTGAAGCCAGGTTTC | CATTAAGTGTGTTGAATCTGGTGGTCCTGAGC | ACGGCCAGCGCAACCTA | 21 |
| UCYN-A | GGTTACAACAACGTTTTATGTGTTGA | TCTGGTGGTCCTGAGCCCGGA | GCAGTAATAATACCACGACCAGCAC | 21 |
| UCYN-B | TGGTCCTGAGCCTGGAGTTG | TGTGCTGGTCGTGGTAT | CTTCTTCTAGGAAGTTGATGGAGGTG | 21 |
| UCYN-C | ATACCAAGGAATCAAGTGTGTTGAGT | CGGTGGTCCCGAGCCTGGAG | ATACCAAGGAATCAAGTGTGTTGAGT | 21 |
| γ-24774A11 | TCCACACGTCTTATTCTGCACT | AAGTGCTTAAGGTTGGCTTTGGCGACA | AGAGCAAACAATGTAGATTTCCTG | 22 |
| Sagittula castanea | ATCACCGCCATCAACTTCCT | CGCCTACGATGACGTGGATTACGTGTCC | AGACCACGTCGCCCAGAAC | 32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, C.; Wu, C.; Li, L.; Pujari, L.; Zhang, G.; Sun, J. Comparison of Diazotrophic Composition and Distribution in the South China Sea and the Western Pacific Ocean. Biology 2021, 10, 555. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060555
Ding C, Wu C, Li L, Pujari L, Zhang G, Sun J. Comparison of Diazotrophic Composition and Distribution in the South China Sea and the Western Pacific Ocean. Biology. 2021; 10(6):555. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060555
Chicago/Turabian StyleDing, Changling, Chao Wu, Liuyang Li, Laxman Pujari, Guicheng Zhang, and Jun Sun. 2021. "Comparison of Diazotrophic Composition and Distribution in the South China Sea and the Western Pacific Ocean" Biology 10, no. 6: 555. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10060555