
Contrasting Effects of Forest Type and Stand Age on Soil Microbial Activities: An Analysis of Local Scale Variability

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
- (1)
- Higher temperatures in summer and spring;
- (2)
- Higher water availability in spring than in autumn and winter (while drought conditions in summer may reduce microbial activity);
- (3)
- Higher water storage and nutrient (N, P, C) availability in heavy vs. light soils;
- (4)
- Lower water and higher nutrient demand of younger trees;
- (5)
- The presence of leaf litter, which is most abundant in mature deciduous forests, and which could maintain a higher soil temperature in winter than in sites without litter and protect against excessive water loss in summer.
2. Materials and Methods
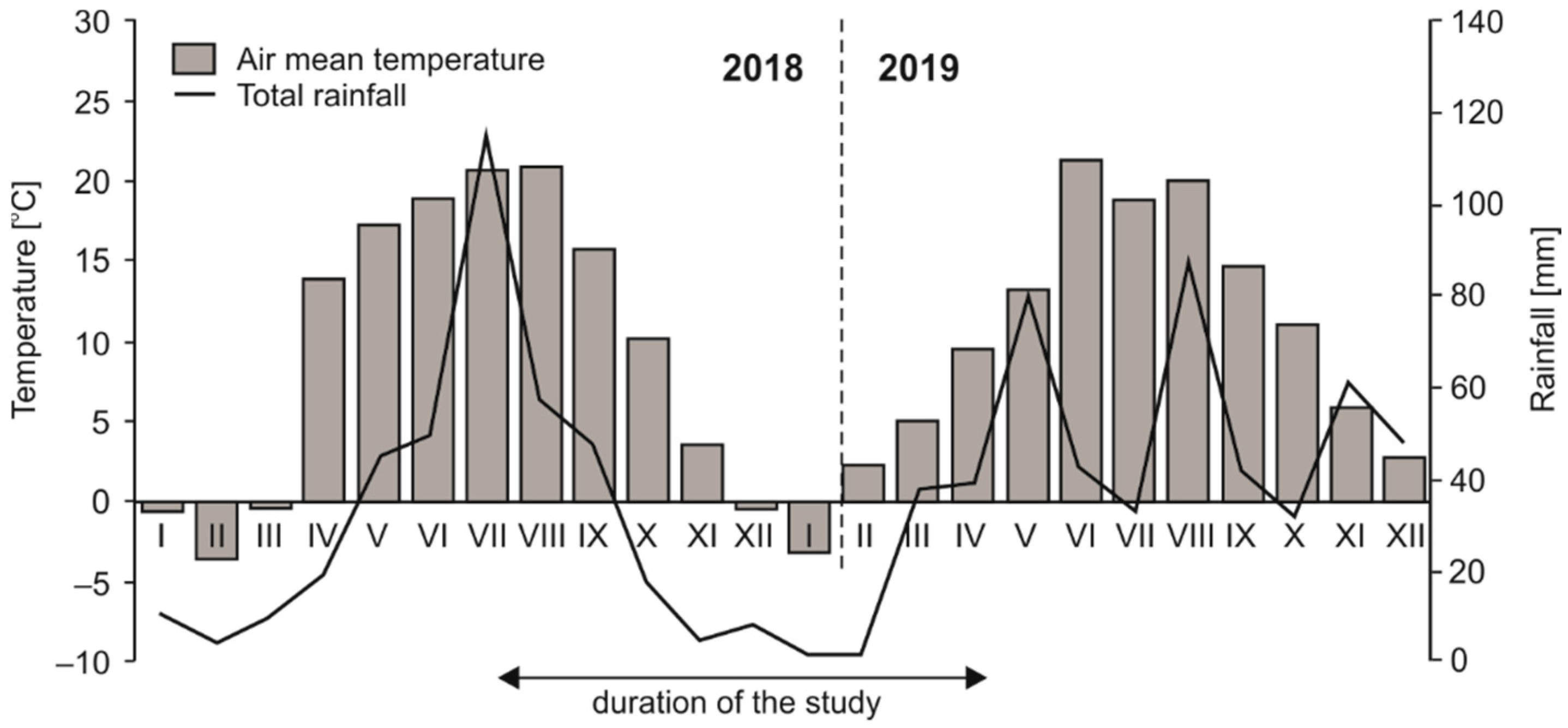
2.1. Site and Soil Characteristics
2.2. Experimental Design
2.3. Analysis of Microbial Parameters
2.4. Calculation and Statistical Analysis
3. Results
3.1. Annual Mean of Microbiological Parameters of Forest Soils
3.2. Seasonal Changes in Soil Basal Respiration (BR)
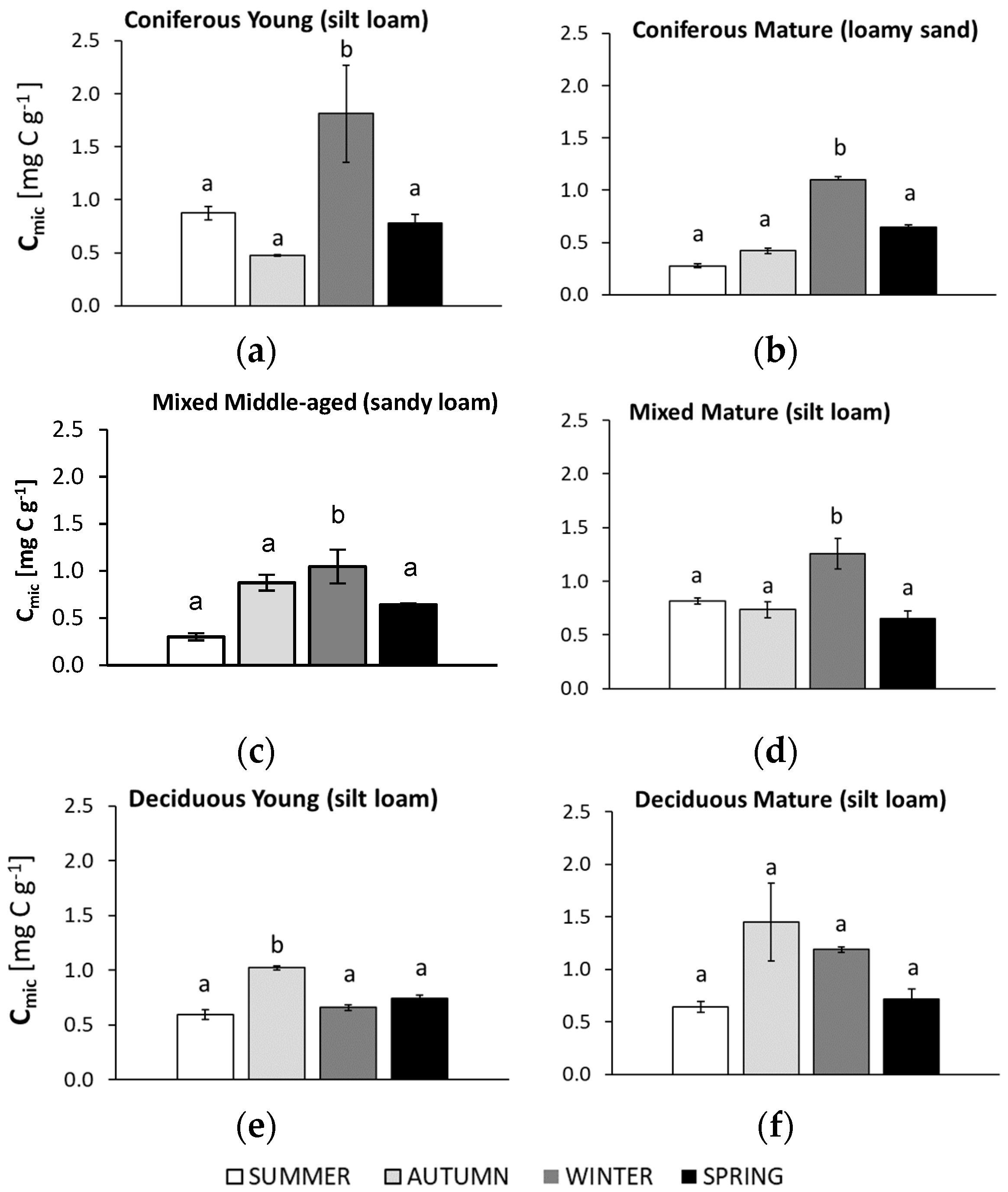
3.3. Seasonal Changes in Microbial Biomass (Cmic)
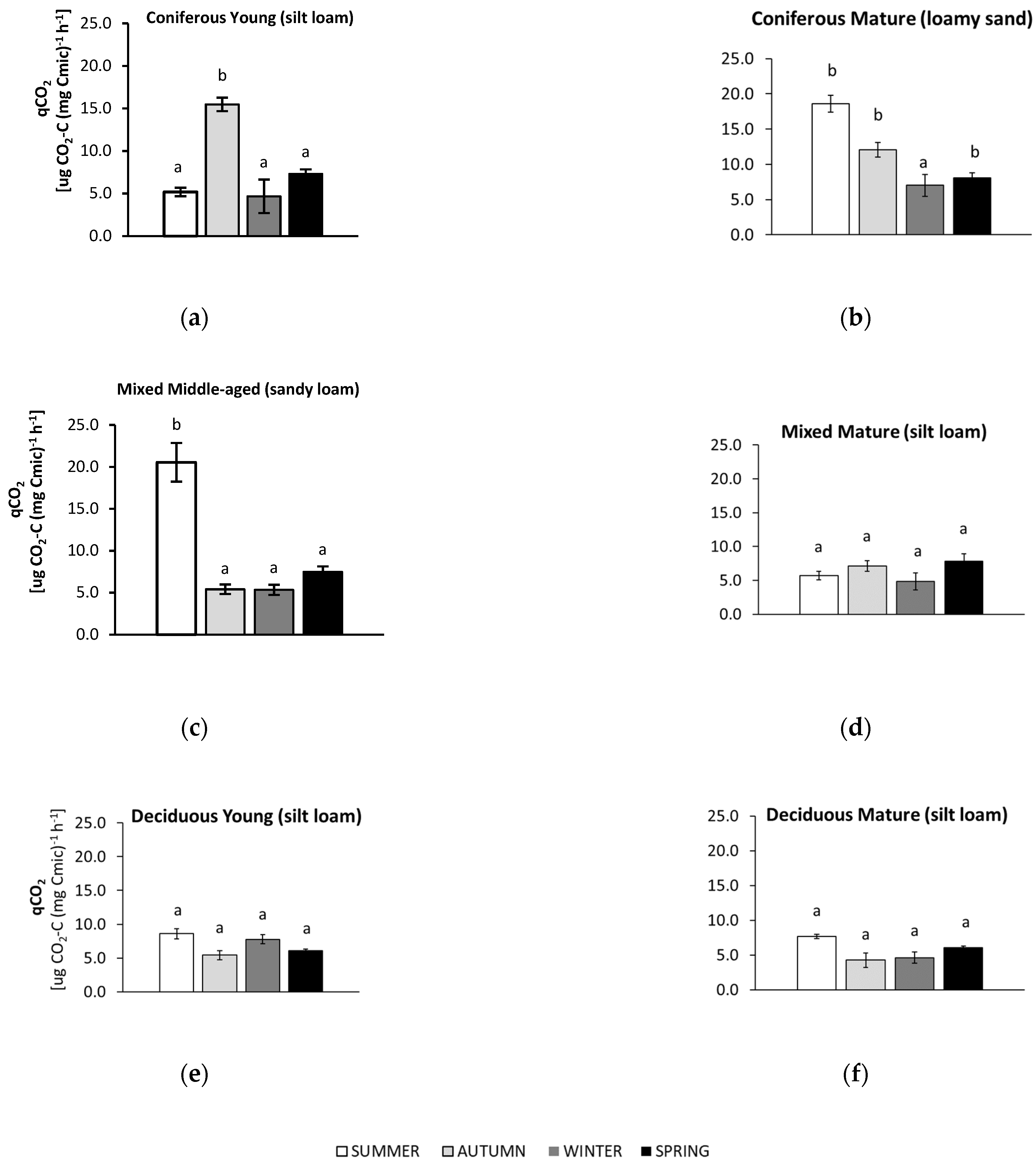
3.4. Seasonal Changes in Soil Metabolic Quotient (qCO2)
3.5. Seasonal Changes in Soil Dehydrogenase Activity (DHA)
3.6. Seasonal Changes in Soil Cmic:Corg Ratio
3.7. Principal Component Analysis
4. Discussion
4.1. Soil Basal Respiration (BR)
4.2. Soil Microbial Biomass (Cmic)
4.3. Microbial Metabolic Quotient (qCO2)
4.4. Soil Dehydrogenase Activity (DHA)
4.5. Soil Cmic:Corg
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Summer | Autumn | Winter | Spring |
|---|---|---|---|---|
| Coniferous Mature | 2.4% | 1.9% | 11.4% | 10.6% |
| Coniferous Young | 10.6% | 4.9% | 26.7% | 4.9% |
| Deciduous Mature | 4.3% | 6.7% | 22.0% | 20.5% |
| Deciduous Young | 14.8% | 25.6% | 24.3% | 32.2% |
| Mixed Mature | 11.0% | 5.6% | 32.8% | 11.5% |
| Mixed Middle-aged | 2.7% | 12.9% | 14.4% | 8.7% |

References
- Babur, E.; Dindaroglu, T. Seasonal changes of soil organic carbon and microbial biomass carbon in different forest ecosystems. In Environmental Factors Affecting Human Health; Uher, I., Ed.; IntechOpen Limited: London, UK, 2020; pp. 1–21. [Google Scholar]
- FAO; UNEP. The State of the World’s Forests 2020. In Forests, Biodiversity and People; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Feng, H.; Guo, J.; Han, M.; Wang, W.; Peng, C.; Jin, J.; Song, X.; Yu, S. A review of the mechanisms and controlling factors of methane dynamics in forest ecosystems. For. Ecol. Manag. 2020, 455, 117702. [Google Scholar] [CrossRef]
- Forest Europe. 2020: State of Europe’s Forests; Forest Europe: Bratislava, Slovakia, 2020. [Google Scholar]
- Forest Europe. 2015: State of Europe’s Forests; Forest Europe: Madrid, Spain, 2015. [Google Scholar]
- EEA. Technical Report No 9/2006 European Forest Types Categories and Types for Sustainable Forest Management Reporting and Policy, 2nd ed.; European Environment Agency: Copenhagen, Denmark, 2007.
- Binkley, D.; Fisher, R.F. Ecology and Management of Forest Soils, 5th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020. [Google Scholar]
- Lu, J.Z.; Scheu, S. Response of soil microbial communities to mixed beech-conifer forests varies with site conditions. Soil Biol. Biochem. 2021, 155, 108155. [Google Scholar] [CrossRef]
- Muscolo, A.; Panuccio, M.R.; Mallamaci, C.; Sidari, M. Biological indicators to assess short-term soil quality changes in forest ecosystems. Ecol. Indic. 2014, 45, 416–426. [Google Scholar] [CrossRef]
- Nielsen, M.N.; Winding, A. Microorganisms as indicators of soil health. In NERI Technical Report No. 388; National Environmental Research Institute: Silkeborg, Denmark, 2002; p. 84. [Google Scholar]
- Goberna, M.; Sánchez, J.; Pascual, J.A.; García, C. Surface and subsurface organic carbon, microbial biomass and activity in a forest soil sequence. Soil Biol. Biochem. 2006, 38, 2233–2243. [Google Scholar] [CrossRef]
- Morales, D.; Vargas, M.M.; de Oliveira, M.P.; Taffe, B.L.; Comin, J.; Soares, C.R.; Lovato, P. Response of soil microbiota to nine-year application of swine manure and urea. Cienc. Rural 2016, 46, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Salazar, S.; Sánchez, L.E.; Alvarez, J.; Valverde, A.; Galindo, P.; Igual, J.M.; Peix, A.; Santa-Regina, I. Correlation among soil enzyme activities under different forest system management practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- Luo, P.; Han, X.; Wang, Y.; Han, M.; Shi, H.; Liu, N.; Bai, H. Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Ann. Microbiol. 2015, 65, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.A.; Krawulski, C.C.; Bini, D.; Filho, T.G.; Knob, A.; Medina, C.C.; Filho, G.A.; Nogueira, M.A. Reclamation status of a degraded pasture based on soil health indicators. Sci. Agric. 2015, 72, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Susyan, E.A.; Wirth, S.; Ananyeva, N.D.; Stolnikova, E.V. Forest succession on abandoned arable soils in European Russia—Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity. Eur. J. Soil Biol. 2011, 47, 169–174. [Google Scholar] [CrossRef]
- Błońska, E.; Lasota, J.; Zwydak, M. The relationship between soil properties, enzyme activity and land use. For. Res. Pap. 2017, 78, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Januszek, K.; Błońska, E.; Długa, J.; Socha, J. Dehydrogenase activity of forest soils depends on the assay used. Int. Agrophys. 2015, 29, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Quilchano, C.; Marañon, T. Dehydrogenase activity in Mediterranean forest soils. Biol. Fertil. Soils 2002, 35, 102–107. [Google Scholar] [CrossRef]
- Fterich, A.; Mahdhi, M.; Mars, M. Seasonal changes of microbiological properties in steppe soils from degraded arid area in Tunisia. Arid Land Res. Manag. 2014, 28, 49–58. [Google Scholar] [CrossRef]
- Deng, S.P.; Parham, J.A.; Hattey, J.A.; Babu, D. Animal manure and anhydrous ammonia amendment alter microbial carbon use efficiency, microbial biomass, and activities of dehydrogenase and amidohydrolases in semiarid agroecosystems. Appl. Soil Ecol. 2006, 33, 258–268. [Google Scholar] [CrossRef]
- Błońska, E. 2010 Enzyme Activity in Forest Peat Soils. Folia For. Pol. Ser. A For. 2010, 52, 20–25. [Google Scholar]
- Błońska, E.; Lasota, J.; Gruba, P. Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation to other properties of soil derived from loess and glaciofluvial sand. Ecol. Res. 2016, 31, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Peng, X.; Zhao, P.; Yuan, J.; Zhong, C.; Cheng, Y.; Cui, C.; Zhang, S. Soil microbial biomass, basal respiration and enzyme activity of main forest types in the Qinling Mountains. PLoS ONE 2013, 8, e67353. [Google Scholar] [CrossRef]
- Condron, L.M.; Turner, B.L.; Cade-Menun, B.J. Chemistry and dynamics of soil organic phosphorus. In Phosphorus: Agriculture and the Environment; Sims, J.T., Sharpley, A.N., Eds.; ASA, CSSA, and SSSA: Madison, WI, USA, 2005; pp. 87–121. [Google Scholar]
- Hamarashid, N.H.; Othman, M.A.; Hussain, M.A.H. Effect of soil texture on chemical compositions, microbial populations and carbon mineralization in soil. Egypt. J. Exp. Biol. 2010, 6, 59–64. [Google Scholar]
- Hemkemeyer, M.; Dohrmann, A.B.; Christensen, B.T.; Tebbe, C.C. Bacterial preferences for specific soil particle size fractions revealed by community analyses. Front. Microbiol. 2018, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Moeskops, B.; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z. Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; Canuto, R.A., Ed.; IntechOpen Limited: London, UK, 2012; pp. 183–210. [Google Scholar]
- Zavišić, A.; Yang, N.; Marhan, S.; Kandeler, E.; Polle, A. Forest soil phosphorus resources and fertilization affect ectomycorrhizal community composition, beech P uptake efficiency, and photosynthesis. Front. Plant Sci. 2018, 9, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinistö, S.M.; Silvola, J.; Kellomäki, S. Soil CO2 efflux in a boreal pine forest under atmospheric CO2 enrichment and air warming. Glob. Chang. Biol. 2004, 10, 1363–1376. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; King, J.S.; Burton, A.J.; Brown, S.E. Responses of tree fine roots to temperature. New Phytol. 2000, 147, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Li, M.H.; Cheng, X.B.; Wang, C.G.; Fan, A.N.; Shi, L.X.; Wang, X.X.; Han, S. Soil respiration in relation to photosynthesis of Quercus mongolica trees at elevated CO2. PLoS ONE 2010, 5, e15134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackl, E.; Buchmann, G.; Zechmeister-Boltenstern, S. Soil microbial biomass and rhizosphere effects in natural forest stands. Phyton 2000, 40, 83–90. [Google Scholar]
- Bolat, İ.; Kara, Ö.; Tunay, M. Effects of seasonal changes on microbial biomass and respiration of forest floor and topsoil under Bornmullerian firstand. Eurasian J. For. Sci. 2015, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Thoms, C.; Gleixner, G. Seasonal differences in tree species’ influence on soil microbial communities. Soil Biol. Biochem. 2013, 66, 239–248. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroğlu, T.; Solaiman, Z.M.; Battaglia, M.L. Microbial respiration, microbial biomass and activity are highly sensitive to forest tree species and seasonal patterns in the Eastern Mediterranean Karst Ecosystems. Sci. Tot. Environ. 2021, 775, 145868. [Google Scholar] [CrossRef]
- Błońska, E.; Piaszczyk, W.; Staszel, K.; Lasota, J. Enzymatic activity of soils and soil organic matter stabilization as an effect of components released from the decomposition of litter. Appl. Soil Ecol. 2021, 157, 103723. [Google Scholar] [CrossRef]
- Sessitsch, A.; Weilharter, A.; Gerzabek, M.H.; Kirchmann, H.; Kandeler, E. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment. Appl. Environ. Microbiol. 2001, 67, 4215–4224. [Google Scholar] [CrossRef] [Green Version]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C:N ratios in European forest soils. For. Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weathers, K.C.; Arthur, M.A. Control of nitrogen loss from forested watersheds by soil carbon:nitrogen ratio and tree species composition. Ecosystems 2002, 5, 712–718. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weathers, K.C.; Arthur, M.A.; Schultz, J.C. Nitrogen cycling in a northern hardwood forest: Do species matter? Biogeochemistry 2004, 67, 289–308. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Hedo, J.; Cerdá, A.; Candel-Pérez, D.; Viñegla, B. Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine (Pinus nigra Ar. ssp. salzmannii) forest. Sci. Total Environ. 2016, 562, 145–154. [Google Scholar] [CrossRef]
- Schultz, H.; Sigrid, H.; Stange, C.F. Species-specific differences in nitrogen uptake and utilization by six European tree species. J. Plant Nutr. Soil Sci. 2011, 174, 28–37. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.; Xia, H.; Zou, B.; Li, N.; Liu, J.; Zhu, W. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci. Plant Nutr. 2010, 56, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Xiang, W.; Chen, L.; Ouyang, S.; Xiao, W.; Li, S.; Forrester, D.I.; Lei, P.; Zeng, Y.; Deng, X.; et al. Soil phosphorus bioavailability and recycling increased with stand age in Chinese fir plantations. Ecosystems 2020, 23, 973–988. [Google Scholar] [CrossRef]
- Zhou, Y.; Boutton, T.W.; Wu, X.B. Soil C:N:P stoichiometry responds to vegetation change from grassland to woodland. Biogeochemistry 2018, 140, 341–357. [Google Scholar] [CrossRef]
- Scheibe, A.; Steffens, C.; Seven, J.; Jacob, A.; Hertel, D.; Leuschner, C.; Gleixner, G. Effects of tree identity dominate over tree diversity on the soil microbial community structure. Soil Biol. Biochem. 2015, 8, 219–227. [Google Scholar] [CrossRef]
- Khlifa, R.; Paquette, A.; Messier, C.; Reich, P.B.; Munson, A.D. Do temperate tree species diversity and identity influence soil microbial community function and composition? Ecol. Evol. 2017, 7, 7965–7974. [Google Scholar] [CrossRef] [Green Version]
- Buragohain, P.; Nath, D.J.; Phonglosa, A. Role of microbes on carbon sequestration. Intern. J. Microb. Res. 2019, 1, 1464–1468. [Google Scholar]
- Bieganowski, A.; Ryżak, M.; Sochan, A.; Makó, A.; Barna, G.; Hernádi, H.; Beczek, M.; Polakowski, C. Laser diffractometry in the measurements of soil and sediment particle size distribution. Adv. Agron. 2018, 151, 215–279. [Google Scholar]
- Kitowski, I.; Sujak, A.; Wiacek, D.; Strobel, W.; Rymarz, M. Trace element residues in eggshells of Grey Heron (Ardea cinerea) from colonies of East Poland. North-West. J. Zool. 2014, 10, 346–354. [Google Scholar]
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Šimek, M.; Kalčík, J. Carbon and nitrate utilization in soils: The effect of long-term fertilization on potential denitrification. Geoderma 1998, 83, 269–280. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil Dehydrogenase Activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Agnelli, A.; Ascher, J.; Corti, G.; Ceccherini, M.T.; Nannipieri, P.; Pietramellara, G. Distribution of microbial communities in a forest soil profile investigated by microbial biomass, soil respiration and DGGE of total and extracellular DNA. Soil Biol. Biochem. 2004, 36, 859–868. [Google Scholar] [CrossRef]
- Meisner, A.; Rousk, J.; Bååth, E. Prolonged drought changes the bacterial growth response to rewetting. Soil Biol. Biochem. 2015, 88, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Koponen, H.T.; Bååth, E. Soil bacterial growth after a freezing/thawing event. Soil Biol. Biochem. 2016, 100, 229–232. [Google Scholar] [CrossRef]
- Schmiel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Malchair, S.; Carnol, M. Microbial biomass and C and N transformations in forest floors under European beech, sessile oak, Norway spruce and Douglas-fir at four temperate forest sites. Soil Biol. Biochem. 2009, 41, 831–839. [Google Scholar] [CrossRef]
- Bérard, A.; Sassi, M.B.; Kaisermann, A.; Renault, P. Soil microbial community responses to heat wave components: Drought and high temperature. Clim. Res. 2015, 66, 243–264. [Google Scholar] [CrossRef]
- Gillespie, L.M.; Fromin, N.; Milcu, A.; Buatois, B.; Pontoizeau, C.; Hättenschwiler, S. Higher tree diversity increases soil microbial resistance to drought. Commun. Biol. 2020, 3, 377. [Google Scholar] [CrossRef]
- Prevost-Boure, N.C.; Maron, P.A.; Ranjard, L.; Nowak, V.; Dufrene, E.; Damesin, C.; Soudani, K.; Lata, J.C. Seasonal dynamics of the bacterial community in forest soils under different quantities of leaf litter. Appl. Soil Ecol. 2011, 47, 14–23. [Google Scholar] [CrossRef]
- Ananyeva, N.D.; Susyan, E.A.; Gavrilenko, E.G. Determination of the soil microbial biomass carbon using the method of substrate-induced respiration. Eurasian Soil Sci. 2011, 44, 1215–1221. [Google Scholar] [CrossRef]
- Thoms, C.; Gattinger, A.; Jacob, M.; Thomas, F.M.; Gleixner, G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biol. Biochem. 2010, 42, 1558–1565. [Google Scholar] [CrossRef]
- Henry, H.A.L. Soil freeze-thaw cycle experiments: Trends, methodological weaknesses and suggested improvements. Soil Biol. Biochem. 2007, 39, 977–986. [Google Scholar] [CrossRef]
- Koponen, H.T.; Jaakkola, T.; Keinänen-Toivola, M.M.; Kaipainen, S.; Tuomainen, J.; Servomaa, K.; Martikainen, P.J. Microbial communities, biomass, and activities in soil as affected by freeze thaw cycles. Soil Biol. Biochem. 2006, 38, 1861–1871. [Google Scholar] [CrossRef]
- Alvarez, E.; Torrado, V.M.; Marcos, M.L.F.; Diaz-Ravia, M. Microbial biomass and activity in a forest soil under different tree species. Electron. J. Environ. Agric. Food Chem. 2009, 8, 878–887. [Google Scholar]
- Díaz-Raviña, M.; Acea, M.J.; Carballas, T. Seasonal changes in microbial biomass and nutrient flush in forest soils. Biol. Fertil. Soils 1995, 19, 220–226. [Google Scholar] [CrossRef]
- Voříšková, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Bauhus, J.; Paré, D.; Côté, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Graham, M.H.; Haynes, R.J. Organic matter status and the size, activity and metabolic diversity of the soil microflora as indicators of the success of rehabilitation of mined sand dunes. Biol. Fertil. Soils 2004, 39, 429–437. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, D.; Zhou, G.; Liu, J. Soil respiration associated with forest succession in subtropical forests in Dinghushan Biosphere Reserve. Soil Biol. Biochem. 2009, 41, 991–999. [Google Scholar] [CrossRef]
- Chodak, M.; Niklińska, M. Effect of texture and tree species on microbial properties of mine soils. Appl. Soil Ecol. 2010, 46, 268–275. [Google Scholar] [CrossRef]
- Neto, M.S.; Scopel, E.; Corbeels, M.; Cardoso, A.N.; Douzet, J.M.; Feller, C.; Piccolo, M.C.; Cerri, C.C.; Bernoux, M. Soil carbon stocks under no-tillage mulch-based cropping systems in the Brazilian Cerrado: An on-farm synchronic assessment. Soil Tillage Res. 2010, 110, 187–195. [Google Scholar] [CrossRef]
- Lu, H.; Li, Z.; Fu, S.; Méndez, A.; Gascó, G.; Paz-Ferreiro, J. Combining phytoextraction and biochar addition improves soil biochemical properties in a soil contaminated with Cd. Chemosphere 2015, 119, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.M.; Cao, J.; Wang, C.; Wang, G. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China. For. Ecol. Manag. 2005, 217, 117–125. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Novak, E.; Carvalho, L.A.; Santiago, E.F.; Portilho, I.I.R. Chemical and microbiological attributes under different soil cover. Cerne 2017, 23, 19–30. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate firest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Schimel, J.P.; Janssens, I.A.; Song, X.; Song, C.; Yu, G.; Sinsabaugh, R.L.; Tang, D.; Zhang, X.; Thornton, P.E. Global pattern and controls of soil microbial metabolic quotient. Ecol. Monogr. 2017, 87, 429–441. [Google Scholar] [CrossRef] [Green Version]
- Böhme, L.; Langer, U.; Böhme, F. Microbial biomass, enzyme activities and microbial community structure in two European long-term field experiments. Agric. Ecosyst. Environ. 2005, 109, 141–152. [Google Scholar] [CrossRef]
- Bérard, A.; Sassi, M.B.; Renault, P.; Gros, R. Severe drought-induced community tolerance to heat wave. An experimental study on soil microbial processes. J. Soils Sediments 2012, 12, 513–518. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biol. Biochem. 1990, 22, 251–255. [Google Scholar] [CrossRef]
- Bauhus, J.; Khanna, P.K. The significance of microbial biomass in forest soils. In Going Underground—Ecological Studies in Forest Soils; Rastin, N., Bauhus, J., Eds.; Research Signpost: Trivandrum, India, 1999; pp. 77–110. [Google Scholar]
- Agnelli, A.; Ugolini, F.C.; Corti, P.; Pietramellara, G. Microbial biomass-C and basal respiration of fine earth and highly altered rock fragments of two forest soils. Soil Biol. Biochem. 2001, 33, 613–620. [Google Scholar] [CrossRef]
- Wardle, D.A.; Ghani, A. A critique of the microbial metabolic quotient (qCO2) as a bioindicator of disturbance and ecosystem development. Soil Biol. Biochem. 1995, 27, 1601–1610. [Google Scholar] [CrossRef]
- Priha, O.; Smolander, A. Microbial biomass and activity in soil and litter under Pinus sylvestris, Picea abies and Betula pendula at originally similar field afforestation sites. Biol. Fertil. Soils 1997, 24, 45–51. [Google Scholar] [CrossRef]
- Rahman, M.M.; Tsukamoto, J.; Rahman, M.M.; Yoneyama, A.; Mostafa, K.M. Lignin and its effects on litter decomposition in forest ecosystems. Chem. Ecol. 2013, 29, 540–553. [Google Scholar] [CrossRef]
- Smolander, A.; Loponen, J.; Suominen, K.; Kitunen, V. Organic matter characteristics and C and N transformations in the humus layer under two tree species, Betula pendula and Picea abies. Soil Biol. Biochem. 2005, 37, 1309–1318. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z. Microorganisms abundance and dehydrogenase activity as a consequence of soil reoxidation process. In Soil Tillage & Microbial Activities; Miransari, M., Ed.; Research Singpost: Kerala, India, 2011; pp. 111–143. [Google Scholar]
- Józefowska, A.; Woś, B.; Pietrzykowski, M. Tree species and soil substrate effects on soil biota during early soil forming stages at afforested mine sites. App. Soil Ecol. 2016, 102, 70–79. [Google Scholar] [CrossRef]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Kleja, D.B. Differences in soil properties in adjacent stand of Scots pine, Norway spruce and silver birch in SW Sweden. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]
- Brzezińska, M.; Stępniewski, W.; Stępniewska, Z.; Przywara, G. Effect of oxygen deficiency on soil dehydrogenase activity in a pot experiment with Triticale cv. Jago vegetation. Int. Agrophys. 2001, 15, 145–149. [Google Scholar]
- Ros, M.; Hernandez, M.; Garcia, C. Soil microbial activity after restoration of a semiarid soil by organic amendments. Soil Biol. Biochem. 2003, 35, 463–469. [Google Scholar] [CrossRef]
- Włodarczyk, T.; Stępniewski, W.; Brzezińska, M. Dehydrogenase activity, redox potential, and emissions of carbon dioxide and nitrous oxide from Cambisols under flooding conditions. Biol. Fertil. Soils 2002, 36, 200–206. [Google Scholar] [CrossRef]
- Almeida, D.; Filho, O.K.; Almeida, H.C.; Gebler, L.; Felipe, A.F. Soil microbial biomass under mulch types in an integrated apple orchard from Southern Brazil. Sci. Agric. 2011, 68, 217–222. [Google Scholar] [CrossRef]
- Stolnikova, E.V.; Ananyeva, N.D.; Chernova, O.V. The microbial biomass and its activity and structure in the soils of old forests in the European Russia. Eurasian Soil Sci. 2011, 44, 437–452. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Ratios of microbial biomass carbon to total organic carbon in arable soils. Soil Biol. Biochem. 1989, 21, 471–479. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]










| Forest Type | Coordinates | Tree Species | Contribution of Dominant Tree Species * | Tree Age * | Trees Biomass * |
|---|---|---|---|---|---|
| [%] | [year] | [m3 ha−1] | |||
| Coniferous Mature | 51.109675, 22.455327 | Scot pine Pinus sylvestris | 100 | 100 | 83 |
| Coniferous Young | 51.036295, 22.993784 | Norway spruce Picea abies | 100 | 36 | 68 |
| Deciduous Mature | 51.034831, 22.993789 | Oak Quercus robur Aspen Populus tremula | 70 30 | 71 63 | 193 87 |
| Deciduous Young | 51.217270, 22.670177 | Sweet cherry Prunus avium | 100 | 12 | 26 |
| Mixed Mature | 51.220832, 22.676460 | Scot pine Pinus sylvestris Oak Quercus robur | 80 20 | 85 85 | 306 65 |
| Mixed Middle Aged | 51.111152, 22.453757 | Oak Quercus robur Scot pine Pinus sylvestris Black alder Alnus glutinosa | 80 10 10 | 59 59 59 | 199 35 21 |
| Forest Type | Coniferous Mature | Coniferous Young | Deciduous Mature | Deciduous Young | Mixed Mature | Mixed Middle Aged |
|---|---|---|---|---|---|---|
| Soil Type | Dystric Cambisol | Albic Dystric Luvisol | Leptic Cambisol | Dystric Cambisol | Dystric Cambisol | Albic Luvisol |
| Texture | loamy sand | silt loam | silt loam | silt loam | silt loam | sandy loam |
| Clay Content [%] | 1.6 ± 0.11 | 5.6 ± 0.11 | 5.4 ± 0.13 | 4.8 ± 0.10 | 5.1 ± 0.15 | 2.0 ± 0.08 |
| Silt Content [%] | 22.9 ± 1.29 | 72.4 ± 0.35 | 69.7 ± 0.53 | 64.3 ± 1.40 | 69.4 ± 0.76 | 27.7 ± 0.82 |
| Sand Content [%] | 75.5 ± 1.40 | 22.1 ± 0.38 | 24.9 ± 0.62 | 30.8 ± 1.43 | 25.4 ± 0.66 | 70.3 ± 0.89 |
| Bulk Density [g cm−3] | 1.20 | 0.94 | 1.14 | 1.23 | 1.13 | 1.23 |
| pHH2O | 4.37 ± 0.006 | 4.82 ± 0.015 | 4.80 ± 0.006 | 5.34 ± 0.006 | 4.22 ± 0.021 | 4.79 ± 0.010 |
| N [%] | 0.074 ± 0.016 | 0.083 ± 0.004 | 0.108 ± 0.017 | 0.107 ± 0.002 | 0.163 ± 0.004 | 0.063 ± 0.001 |
| Corg [%] | 1.20 ± 0.363 | 0.91 ± 0.068 | 1.73 ± 0.670 | 1.14 ± 0.065 | 2.25 ± 0.103 | 0.82 ± 0.074 |
| C/N | 16.12 ± 1.39 | 11.04 ± 0.85 | 15.71 ± 3.68 | 10.66 ± 0.43 | 13.81 ± 0.30 | 13.17 ± 0.95 |
| Ptot [mg/kg] | 277.0 ± 6.74 | 273.6 ± 9.41 | 316.7 ± 23.81 | 456.8 ± 159.5 | 458.2 ± 20.36 | 264.2 ± 14.55 |
| C/P | 42.4 ± 12.94 | 32.3 ± 2.51 | 51.2 ± 18.03 | 18.6 ± 0.78 | 47.2 ± 2.29 | 29.7 ± 2.63 |
| N/P | 2.60 ± 0.56 | 2.93 ± 0.16 | 3.21 ± 0.46 | 1.77 ± 0.016 | 3.41 ± 0.16 | 2.25 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walkiewicz, A.; Bieganowski, A.; Rafalska, A.; Khalil, M.I.; Osborne, B. Contrasting Effects of Forest Type and Stand Age on Soil Microbial Activities: An Analysis of Local Scale Variability. Biology 2021, 10, 850. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090850
Walkiewicz A, Bieganowski A, Rafalska A, Khalil MI, Osborne B. Contrasting Effects of Forest Type and Stand Age on Soil Microbial Activities: An Analysis of Local Scale Variability. Biology. 2021; 10(9):850. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090850
Chicago/Turabian StyleWalkiewicz, Anna, Andrzej Bieganowski, Adrianna Rafalska, Mohammad I. Khalil, and Bruce Osborne. 2021. "Contrasting Effects of Forest Type and Stand Age on Soil Microbial Activities: An Analysis of Local Scale Variability" Biology 10, no. 9: 850. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090850