
Response to Climate Change: Evaluation of Methane Emissions in Northern Australian Beef Cattle on a High Quality Diet Supplemented with Desmanthus Using Open-Circuit Respiration Chambers and GreenFeed Emission Monitoring Systems

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Procedures
2.2. Desmanthus Cultivars
2.3. Experiment 1
2.3.1. Animals and Treatments
2.3.2. Measurement of Methane Emissions
2.4. Experiment 2
2.4.1. Animals and Treatments
2.4.2. Measurement of Methane Emissions
2.5. Feed Chemical Composition
2.6. Plant Extraction Procedure and Analysis of Secondary Compounds
2.7. Dry Matter Intake and Liveweight
2.8. Statistical Analyses
3. Results
3.1. Experiment 1
3.1.1. Chemical Composition of the Diets
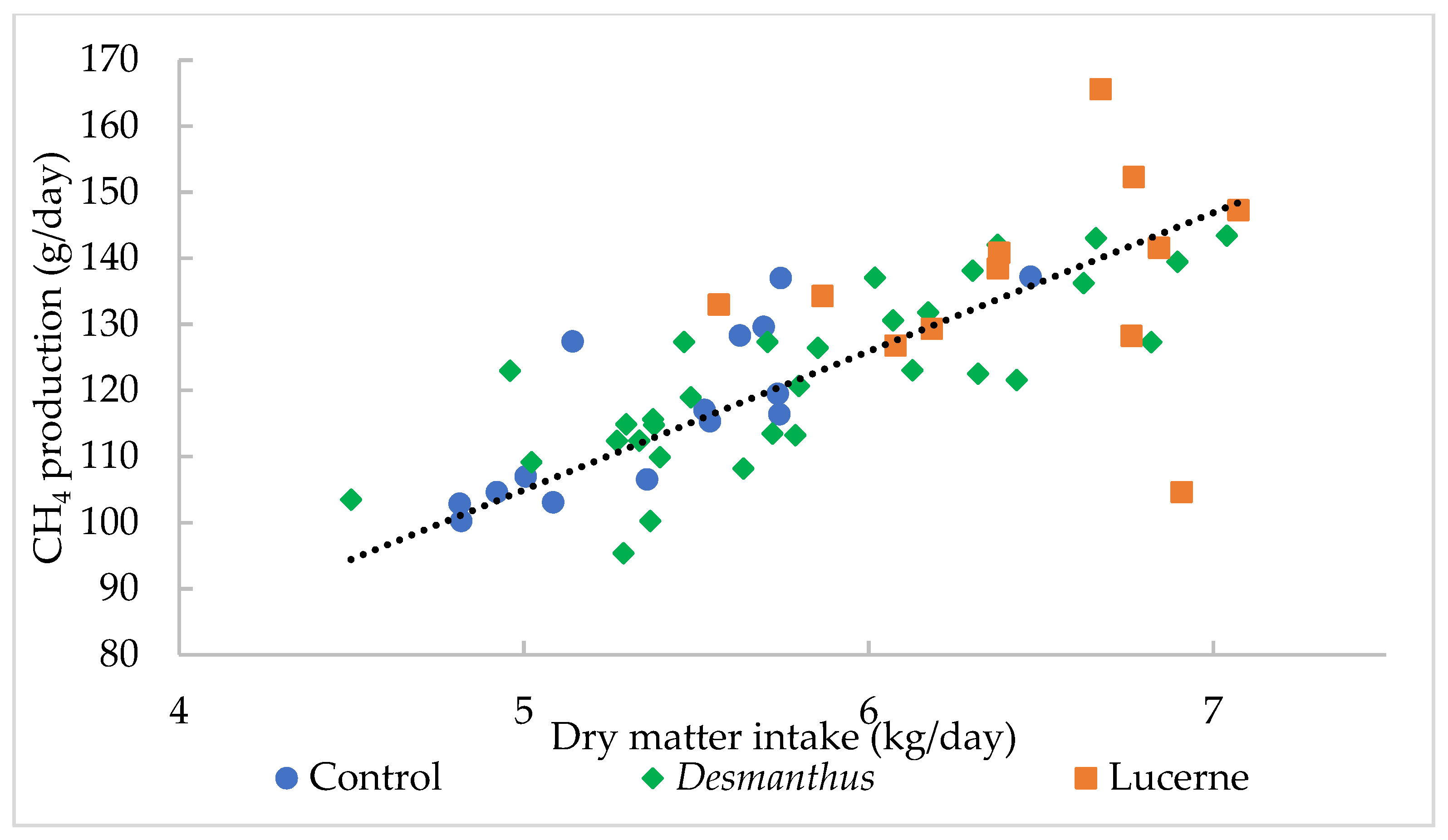
3.1.2. Impact of Diet on Methane Emissions
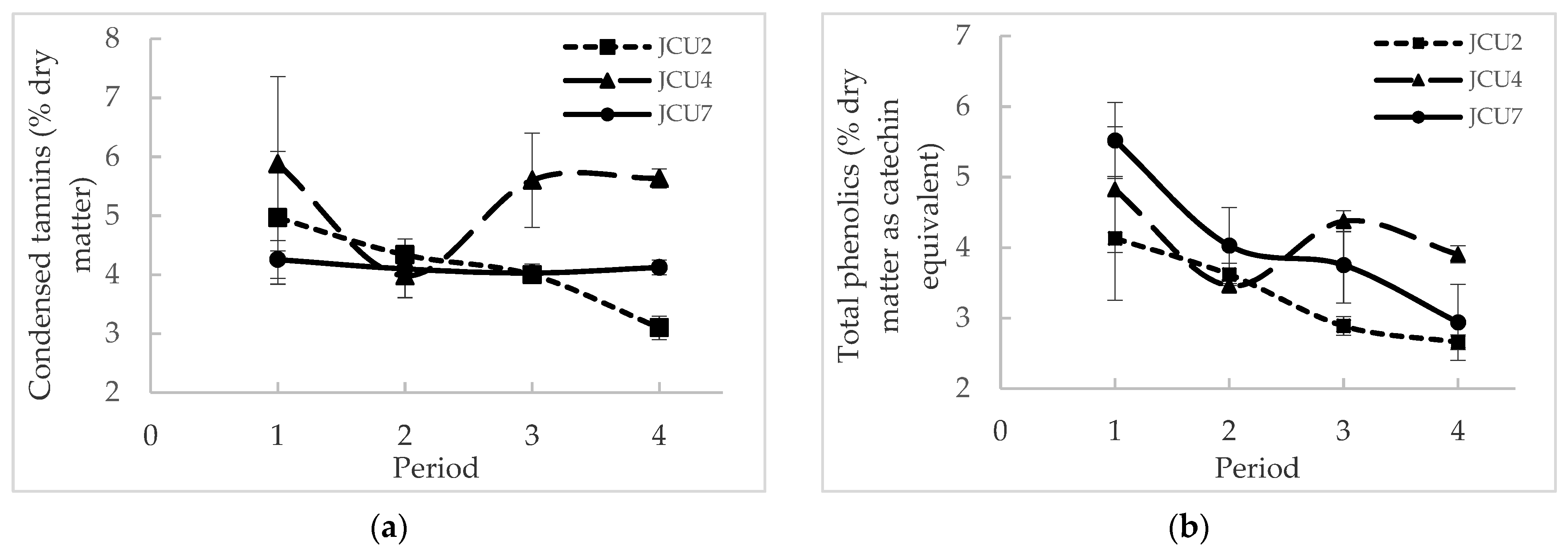
3.1.3. Effect of Polyethylene Glycol on Nutritive Intakes, Phenolics, Tannins Concentrations and Methane Emissions
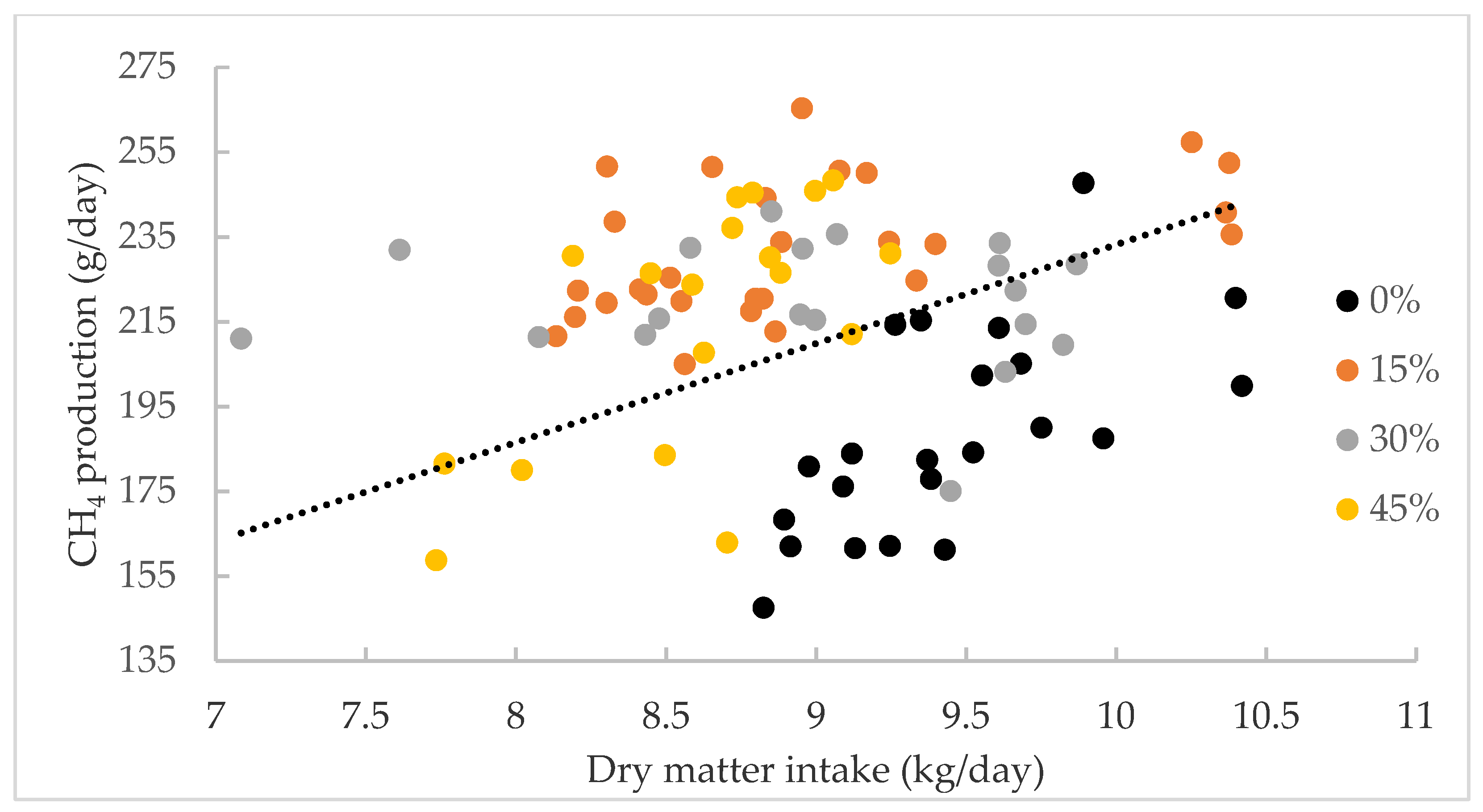
3.2. Experiment 2
4. Discussion
4.1. Experiment 1
4.1.1. Comparison of Methane Emissions between the Desmanthus Cultivars
4.1.2. Comparison of Methane Emissions between the Backgrounding and Desmanthus Diets
4.1.3. Comparison of the Methane Emissions between the Lucerne and Desmanthus Diets
4.1.4. Effect of Polyethylene Glycol on Methane Emissions
4.2. Experiment 2—Effect of Level of Inclusion of Desmanthus Cultivars on Intake, Methane Production and Yield
4.3. Comparison between Open-Circuit and GreenFeed Emission Monitoring Systems
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagundes, G.M.; Benetel, G.; Carriero, M.M.; Sousa, R.L.M.; Muir, J.P.; Macedo, R.O.; Bueno, I.C.S. Tannin-rich forage as a methane mitigation strategy for cattle and the implications for rumen microbiota. Anim. Prod. Sci. 2020, 61, 26–37. [Google Scholar] [CrossRef]
- Fagundes, G.M.; Benetel, G.; Santos, K.C.; Welter, K.C.; Melo, F.A.; Muir, J.P.; Bueno, I.C.S. Tannin-Rich Plants as Natural Manipulators of Rumen Fermentation in the Livestock Industry. Molecules 2020, 25, 2943. [Google Scholar] [CrossRef]
- Vandermeulen, S.; Singh, S.; Ramírez-Restrepo, C.A.; Kinley, R.D.; Gardiner, C.P.; Holtum, J.A.; Hannah, I.; Bindelle, J. In vitro assessment of ruminal fermentation, digestibility and methane production of three species of Desmanthus for application in northern Australian grazing systems. Crop Pasture Sci. 2018, 69, 797–807. [Google Scholar] [CrossRef]
- Durmic, Z.; Ramírez-Restrepo, C.A.; Gardiner, C.; O’Neill, C.J.; Hussein, E.; Vercoe, P.E. Differences in the nutrient concentrations, in vitro methanogenic potential and other fermentative traits of tropical grasses and legumes for beef production systems in northern Australia. J. Sci. Food Agric. 2017, 97, 4075–4086. [Google Scholar] [CrossRef]
- Suybeng, B.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Malau-Aduli, A.E. Supplementing northern Australian beef cattle with Desmanthus tropical legume reduces in-vivo methane emissions. Animals 2020, 10, 2097. [Google Scholar] [CrossRef]
- Terranova, M.; Wang, S.; Eggerschwiler, L.; Braun, U.; Kreuzer, M.; Schwarm, A. Dose-response effects of woody and herbaceous forage plants on in vitro ruminal methane and ammonia formation, and their short-term palatability in lactating cows. Animal 2020, 14, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef] [Green Version]
- Gaviria-Uribe, X.; Bolivar, D.M.; Rosenstock, T.S.; Molina-Botero, I.C.; Chirinda, N.; Barahona, R.; Arango, J. Nutritional quality, voluntary intake and enteric methane emissions of diets based on novel cayman grass and its associations with two leucaena shrub legumes. Front. Vet. Sci. 2020, 7, 764. [Google Scholar] [CrossRef] [PubMed]
- Boadi, D.A.; Wittenberg, K.M. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. Can. J. Anim. Sci. 2002, 82, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Kanani, J.; Lukefahr, S.; Stanko, R. Evaluation of tropical forage legumes (Medicago sativa, Dolichos lablab, Leucaena leucocephala and Desmanthus bicornutus) for growing goats. Small Rumin. Res. 2006, 65, 1–7. [Google Scholar] [CrossRef]
- McDonnell, R.P.; Staines, M.V.; Douglas, M.L.; Auldist, M.J.; Jacobs, J.L.; Wales, W.J. Rumen degradability characteristics of five starch-based concentrate supplements used on Australian dairy farms. Anim. Prod. Sci. 2017, 57, 1512–1519. [Google Scholar] [CrossRef]
- Charmley, E.; Eady, S.; McSweeney, C. Strategies for measuring and reducing methane emissions from beef cattle in northern Australia. In Proceedings of the Northern Beef Research Update Conference, Darwin, Australia, 3–4 August 2011; pp. 73–80. [Google Scholar]
- Goopy, J.P.; Chang, C.; Tomkins, N. A Comparison of Methodologies for Measuring Methane Emissions from Ruminants. In Methods for Measuring Greenhouse Gas Balances and Evaluating Mitigation Options in Smallholder Agricultur; Rosenstock, T.S., Rufino, M.C., ButterbachBahl, K., Wollenberg, E., Richards, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–117. [Google Scholar]
- Thompson, L.R.; Rowntree, J.E. INVITED REVIEW: Methane sources, quantification, and mitigation in grazing beef systems. Appl. Anim. Sci. 2020, 36, 556–573. [Google Scholar] [CrossRef]
- Zhao, Y.G.; Nan, X.M.; Yang, L.; Zheng, S.S.; Jiang, L.S.; Xiong, B.H. A review of enteric methane emission measurement techniques in ruminants. Animals 2020, 10, 1004. [Google Scholar] [CrossRef] [PubMed]
- Llonch, P.; Troy, S.M.; Duthie, C.-A.; Somarriba, M.; Rooke, J.; Haskell, M.J.; Roehe, R.; Turner, S.P. Changes in feed intake during isolation stress in respiration chambers may impact methane emissions assessment. Anim. Prod. Sci. 2018, 58, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Llonch, P.; Somarriba, M.; Duthie, C.-A.; Haskell, M.J.; Rooke, J.A.; Troy, S.; Roehe, R.; Turner, S.P. Association of temperament and acute stress responsiveness with productivity, feed efficiency, and methane emissions in beef cattle: An observational study. Front. Vet. Sci. 2016, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Huhtanen, P.; Ramin, M.; Hristov, A.N. Enteric methane emission can be reliably measured by the GreenFeed monitoring unit. Livest. Sci. 2019, 222, 31–40. [Google Scholar] [CrossRef]
- Waghorn, G.C.; Jonker, A.; Macdonald, K.A. Measuring methane from grazing dairy cows using GreenFeed. Anim. Prod. Sci. 2016, 56, 252–257. [Google Scholar] [CrossRef]
- Doreau, M.; Arbre, M.; Rochette, Y.; Lascoux, C.; Eugene, M.; Martin, C. Comparison of 3 methods for estimating enteric methane and carbon dioxide emission in nonlactating cows. J. Anim. Sci. 2018, 96, 1559–1569. [Google Scholar] [CrossRef] [Green Version]
- Cook, B.; Pengelly, B.; Schultze-Kraft, R.; Taylor, M.; Burkart, S.; Cardoso Arango, J.A.; Gonzalez Guzman, J.J.; Cox, K.; Jones, C.; Peters, M. Tropical Forages: An Interactive Selection Tool—Digital. Available online: https://apps.lucidcentral.org/tropical_forages/text/intro/index.html (accessed on 23 September 2020).
- Loch, D.S. Desmanthus varieties JCU1–5 descriptors. Plant Var. J. 2015, 28, 59–192. [Google Scholar]
- Gardiner, C.P. Developing and commercializing new pasture legumes for clay soils in the semi-arid rangelands of northern Australia: The new Desmanthus cultivars JCU 1–5 and the Progardes story. In Tropical Forage Legumes: Harnessing the Potential of Desmanthus and Other Genera for Heavy Clay Soils; CABI: Wallingford, UK, 2016; pp. 1–20. [Google Scholar]
- Suybeng, B.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O. Plasma metabolites, productive performance and rumen volatile fatty acid profiles of northern Australian Bos indicus steers supplemented with Desmanthus and lucerne. Metabolites 2021, 11, 356. [Google Scholar] [CrossRef]
- Martinez Fernandez, G.; Duval, S.M.; Kindermann, M.; Schirra, H.J.; Denman, S.E.; McSweeney, C.S. 3-NOP vs Halogenated compound: Methane production, ruminal fermentation and microbial community response in forage fed cattle. Front. Microbiol. 2018, 9, 1582. [Google Scholar] [CrossRef]
- Williams, Y.; Klein, L.; Wright, A.-D. A protocol for the operation of open-circuit chambers for measuring methane output in sheep. In Measuring Methane Production from Ruminants; Springer: Dordrecht, The Netherlands, 2007; pp. 111–123. [Google Scholar]
- Hammond, K.J.; Humphries, D.J.; Crompton, L.A.; Green, C.; Reynolds, C.K. Methane emissions from cattle: Estimates from short-term measurements using a GreenFeed system compared with measurements obtained using respiration chambers or sulphur hexafluoride tracer. Anim. Feed Sci. Technol. 2015, 203, 41–52. [Google Scholar] [CrossRef]
- Coates, D.B.; Dixon, R.M. Developing robust faecal near infrared spectroscopy calibrations to predict diet dry matter digestibility in cattle consuming tropical forages. J. Near Infrared Spectrosc. 2011, 19, 507–519. [Google Scholar] [CrossRef]
- CSIRO. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Collingwood, VIC, Australia, 2007. [Google Scholar]
- Terrill, T.; Rowan, A.; Douglas, G.; Barry, T. Determination of extractable and bound condensed tannin concentrations in forage plants, protein concentrate meals and cereal grains. J. Sci. Food Agric. 1992, 58, 321–329. [Google Scholar] [CrossRef]
- Makkar, H.P. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.M.; Terrill, T.H.; Muir, J.P. Drying method and origin of standard affect condensed tannin (CT) concentrations in perennial herbaceous legumes using simplified butanol-HCl CT analysis. J. Sci. Food Agric. 2008, 88, 1060–1067. [Google Scholar] [CrossRef]
- Wickham, H.; Francois, R.; Henry, L.; Muller, K.; RStudio. Ddplyr: A Grammar of Data Manipulation. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 7 May 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. Available online: https://CRAN.R-project.org/package=nlme (accessed on 7 May 2021).
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications Inc.: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-V, E.; Hussey, M.; Ortega-S, J. Nutritive value of Desmanthus associated with Kleingrass during the establishment year. Rangel. Ecol. Manag. 2005, 58, 308–314. [Google Scholar] [CrossRef]
- McMahon, L.R.; McAllister, T.A.; Berg, B.P.; Majak, W.; Acharya, S.N.; Popp, J.D.; Coulman, B.E.; Wang, Y.; Cheng, K.-J. A review of the effects of forage condensed tannins on ruminal fermentation and bloat in grazing cattle. Can. J. Plant Sci. 2000, 80, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Top, S.M.; Preston, C.M.; Dukes, J.S.; Tharayil, N. Climate influences the content and chemical composition of foliar tannins in green and senesced tissues of Quercus rubra. Front. Plant Sci. 2017, 8, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatta, R.; Tajima, K.; Takusari, N.; Higuchi, K.; Enishi, O.; Kurihara, M. Comparison of in vivo and in vitro techniques for methane production from ruminant diets. Asian-Australas. J. Anim. Sci. 2007, 20, 1049–1056. [Google Scholar] [CrossRef]
- Charmley, E.; Williams, S.R.O.; Moate, P.J.; Hegarty, R.S.; Herd, R.M.; Oddy, V.H.; Reyenga, P.; Staunton, K.M.; Anderson, A.; Hannah, M.C. A universal equation to predict methane production of forage-fed cattle in Australia. Anim. Prod. Sci. 2016, 56, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Benaouda, M.; Gonzalez-Ronquillo, M.; Appuhamy, J.; Kebreab, E.; Molina, L.T.; Herrera-Camacho, J.; Ku-Vera, J.C.; Angeles-Hernandez, J.C.; Castelan-Ortega, O.A. Development of mathematical models to predict enteric methane emission by cattle in Latin America. Livest. Sci. 2020, 241, 104177. [Google Scholar] [CrossRef]
- van Lingen, H.J.; Niu, M.T.; Kebreab, E.; Valadares, S.C.; Rooke, J.A.; Duthie, C.A.; Schwarm, A.; Kreuzer, M.; Hynd, P.I.; Caetano, M.; et al. Prediction of enteric methane production, yield and intensity of beef cattle using an intercontinental database. Agric. Ecosyst. Environ. 2019, 283, 106575. [Google Scholar] [CrossRef]
- Kennedy, P.M.; Charmley, E. Methane yields from Brahman cattle fed tropical grasses and legumes. Anim. Prod. Sci. 2012, 52, 225–239. [Google Scholar] [CrossRef]
- Mertens, D.R. Regulation of Forage Intake; Soil Science Society of America: Madison, WI, USA, 1994; pp. 450–493. [Google Scholar]
- Popp, J.D.; McCaughey, W.P.; Cohen, R.D.H.; McAllister, T.A.; Majak, W. Enhancing pasture productivity with alfalfa: A review. Can. J. Plant Sci. 2000, 80, 513–519. [Google Scholar] [CrossRef]
- Benchaar, C.; Pomar, C.; Chiquette, J. Evaluation of dietary strategies to reduce methane production in ruminants: A modelling approach. Can. J. Anim. Sci. 2001, 81, 563–574. [Google Scholar] [CrossRef]
- Tafaj, M.; Zebeli, Q.; Baes, C.; Steingass, H.; Drochner, W. A meta-analysis examining effects of particle size of total mixed rations on intake, rumen digestion and milk production in high-yielding dairy cows in early lactation. Anim. Feed Sci. Technol. 2007, 138, 137–161. [Google Scholar] [CrossRef]
- Naumann, H.D.; Tedeschi, L.O.; Zeller, W.E.; Huntley, N.F. The role of condensed tannins in ruminant animal production: Advances, limitations and future directions. Rev. Bras. Zootec. 2017, 46, 929–949. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, K.; McGinn, S.; Martinez, T.; McAllister, T. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirumeignanam, D.; Chellapandian, M.; Arulnathan, N. Effect of Supplementation of Graded Levels of Tanniferous Feeds on the Nutritional Quality of Hedge Lucerne Silage for Goats. Anim. Nutr. Feed Technol. 2020, 20, 51–60. [Google Scholar] [CrossRef]
- Animut, G.; Puchala, R.; Goetsch, A.L.; Patra, A.K.; Sahlu, T.; Varel, V.H.; Wells, J. Methane emission by goats consuming different sources of condensed tannins. Anim. Feed Sci. Technol. 2008, 144, 228–241. [Google Scholar] [CrossRef]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Enishi, O.; Kurihara, M. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef] [Green Version]
- Bueno, I.C.S.; Vitti, D.; Louvandini, H.; Abdalla, A.L. A new approach for in vitro bioassay to measure tannin biological effects based on a gas production technique. Anim. Feed Sci. Technol. 2008, 141, 153–170. [Google Scholar] [CrossRef]
- Ku-Vera, J.C.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gómez-Bravo, C.A.; Aguilar-Pérez, C.F.; Solorio-Sánchez, F.J. Role of Secondary Plant Metabolites on Enteric Methane Mitigation in Ruminants. Front. Vet. Sci. 2020, 7, 584. [Google Scholar] [CrossRef]
- Arbre, M.; Rochette, Y.; Guyader, J.; Lascoux, C.; Gomez, L.M.; Eugene, M.; Morgavi, D.P.; Renand, G.; Doreau, M.; Martin, C. Repeatability of enteric methane determinations from cattle using either the SF6 tracer technique or the GreenFeed system. Anim. Prod. Sci. 2016, 56, 238–243. [Google Scholar] [CrossRef]
- Manafiazar, G.; Zimmerman, S.; Basarab, J.A. Repeatability and variability of short-term spot measurement of methane and carbon dioxide emissions from beef cattle using GreenFeed emissions monitoring system. Can. J. Anim. Sci. 2017, 97, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Coppa, M.; Jurquet, J.; Eugene, M.; Dechaux, T.; Rochette, Y.; Lamy, J.M.; Ferlay, A.; Martin, C. Repeatability and ranking of long-term enteric methane emissions measurement on dairy cows across diets and time using GreenFeed system in farm-conditions. Methods 2021, 186, 59–67. [Google Scholar] [CrossRef]
- Denninger, T.M.; Dohme-Meier, F.; Eggerschwiler, L.; Vanlierde, A.; Grandl, F.; Gredler, B.; Kreuzer, M.; Schwarm, A.; Munger, A. Persistence of differences between dairy cows categorized as low or high methane emitters, as estimated from milk mid-infrared spectra and measured by GreenFeed. J. Dairy Sci. 2019, 102, 11751–11765. [Google Scholar] [CrossRef] [Green Version]
- Arthur, P.; Barchia, I.; Weber, C.; Bird-Gardiner, T.; Donoghue, K.; Herd, R.; Hegarty, R. Optimizing test procedures for estimating daily methane and carbon dioxide emissions in cattle using short-term breath measures. J. Anim. Sci. 2017, 95, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Melgar, A. Short communication: Relationship of dry matter intake with enteric methane emission measured with the GreenFeed system in dairy cows receiving a diet without or with 3-nitrooxypropanol. Animal 2020, 14, S484–S490. [Google Scholar] [CrossRef] [PubMed]
- Alemu, A.W.; Vyas, D.; Manafiazar, G.; Basarab, J.A.; Beauchemin, K.A. Enteric methane emissions from low- and high-residual feed intake beef heifers measured using GreenFeed and respiration chamber techniques. J. Anim. Sci. 2017, 95, 3727–3737. [Google Scholar] [CrossRef] [PubMed]
- McGinn, S.; Beauchemin, K.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Hammond, K.J.; Crompton, L.A.; Bannink, A.; Dijkstra, J.; Yáñez-Ruiz, D.R.; O’Kiely, P.; Kebreab, E.; Eugène, M.A.; Yu, Z.; Shingfield, K.J.; et al. Review of current in vivo measurement techniques for quantifying enteric methane emission from ruminants. Anim. Feed Sci. Technol. 2016, 219, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Velazco, J.I.; Mayer, D.G.; Zimmerman, S.; Hegarty, R.S. Use of short-term breath measures to estimate daily methane production by cattle. Animal 2016, 10, 25–33. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variable | Lucerne | JCU2 | JCU4 | JCU7 | Desmanthus spp. 2 | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| CP (% DM) | 10.2 | 9.2 | 10.1 | 9.4 | 9.6 | 0.17 | 0.08 |
| ADF (% DM) | 41.3 a | 41.9 b | 41.9 b | 42.8 b | 42.6 | 0.62 | 0.01 |
| NDF (% DM) | 68.4 | 68.7 | 67.2 | 68.8 | 68.3 | 0.74 | 0.13 |
| ME (MJ/kg DM) 1 | 7.6 a | 7.1 b | 7.1 b | 7.0 b | 7.0 | 0.06 | 0.01 |
| Variable | Backgrounding | Lucerne | JCU2 (D. virgatus) | JCU4 (D. bicornutus) | JCU7 (D. Leptophyllus) | Des. spp. 1 | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Backgrounding vs. Des spp. | Luc. vs. Des. spp. | Des. spp. | |||||||
| DMI (kg/day) | 5.47 ± 0.10 | 6.57 ± 0.10 | 5.96 ± 0.16 | 5.85 ± 0.24 | 5.90 ± 0.21 | 5.90 ± 0.12 | 0.01 | 0.02 | 0.96 |
| DMI/kg LW (%) | 1.99 ± 0.04 | 2.02 ± 0.07 | 2.04 ± 0.06 | 2.03 ± 0.05 | 1.96 ± 0.04 | 2.01 ± 0.03 | 0.70 | 0.98 | 0.44 |
| CP intake (kg/day) | 0.509 ± 0.01 | 0.705 ± 0.04 | 0.568 ± 0.03 | 0.602 ± 0.02 | 0.572 ± 0.03 | 0.582 ± 0.02 | 0.01 | 0.01 | 0.64 |
| ADF intake (kg/day) | 2.39 ± 0.04 | 2.68 ± 0.05 | 2.51 ± 0.06 | 2.40 ± 0.11 | 2.48 ± 0.09 | 2.46 ± 0.05 | 0.30 | 0.02 | 0.66 |
| NDF intake (kg/day) | 4.06 ± 0.08 | 4.37 ± 0.07 | 4.05 ± 0.10 | 3.90 ± 0.16 | 4.02 ± 0.13 | 3.99 ± 0.08 | 0.25 | 0.01 | 0.68 |
| ME intake (MJ) | 38.3 ± 1.24 | 52.1 ± 1.23 | 42.5 ± 1.40 | 43.1 ± 1.79 | 42.6 ± 1.62 | 42.8 ± 0.91 | 0.01 | 0.01 | 0.92 |
| Total phenolics in diet (% DM as catechin equivalent) | 0.191 | 0.313 ± 0.01 | 1.23 ± 0.10 | 1.42 ± 0.12 | 1.50 ± 0.13 | 1.38 ± 0.07 | 0.01 | 0.01 | 0.21 |
| Condensed tannins in diet (% DM) | 0.0799 | 0.685 ± 0.01 | 1.44 ± 0.13 | 1.64 ± 0.21 | 1.37 ± 0.04 | 1.49 ± 0.87 | 0.01 | 0.01 | 0.08 |
| Variable | Backgrounding | Lucerne | JCU2 (D. virgatus) | JCU4 (D. bicornutus) | JCU7 (D. leptophyllus) | Des. spp. 1 | SEM | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Backgrounding vs. Des spp. | Luc. vs. Des. spp. | Des. spp. | ||||||||
| CH4 production (g/day) | 117 | 137 | 122 | 121 | 123 | 122 | 1.93 | 0.01 | 0.01 | 0.92 |
| CH4 yield (g/kg DMI) | 21.6 | 21.1 | 21.0 | 20.8 | 21.1 | 21.0 | 0.21 | 0.93 | 0.36 | 0.93 |
| CH4 (g/kg LW) | 0.428 | 0.448 | 0.415 | 0.414 | 0.411 | 0.414 | 0.01 | 0.24 | 0.01 | 0.83 |
| Variable | Desmanthus spp. | SEM | p-Value | |
|---|---|---|---|---|
| No PEG | PEG | |||
| DMI (kg/day) | 5.6 | 6.2 | 0.21 | 0.20 |
| DMI/kg LW (%) | 1.8 | 1.9 | 0.04 | 0.37 |
| CP intake (kg/day) | 0.62 | 0.67 | 0.04 | 0.51 |
| ADF intake (kg/day) | 2.6 | 2.6 | 0.12 | 0.88 |
| NDF intake (kg/day) | 4.3 | 4.3 | 0.17 | 0.78 |
| ME intake (MJ) | 44.8 | 43.6 | 2.19 | 0.91 |
| Total phenolics in diet (%DM as catechin equivalent) | 1.1 | 1.1 | 0.08 | 0.86 |
| Condensed tannins in diet (% DM) | 1.4 | 1.4 | 0.12 | 0.86 |
| CH4 production (g/day) | 125 | 145 | 4.16 | 0.03 |
| CH4 yield (g/kg DMI) | 22.7 | 23.4 | 0.56 | 0.80 |
| CH4 (g/kg LW) | 0.41 | 0.44 | 0.01 | 0.32 |
| Desmanthus in the Diet | SEM | p-Value | ||||
|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | |||
| DMI (kg/day) | 9.4 | 8.9 | 9.0 | 8.6 | 2.34 | 0.16 |
| DMI/kg LW (%) | 2.26 | 2.19 | 2.23 | 2.11 | 0.02 | 0.77 |
| CP intake (kg/day) | 1.05 | 0.987 | 0.952 | 0.927 | 0.01 | 0.61 |
| ADF intake (kg/day) | 3.51 a | 3.33 ab | 3.17 ab | 3.01 b | 0.03 | 0.01 |
| NDF intake (kg/day) | 5.78 a | 5.45 ab | 5.13 b | 4.87 b | 0.06 | 0.01 |
| ME intake (MJ) | 72.7 a | 69.3 ab | 65.0 bc | 62.1 c | 0.64 | 0.01 |
| CH4 production (g/day) | 188 a | 232 b | 219 b | 215 ab | 2.88 | 0.01 |
| CH4 yield (g/kg DMI) | 20.6 a | 26.2 b | 24.2 c | 25.1 bc | 0.33 | 0.01 |
| CH4 (g/kg LW) | 0.450 a | 0.570 b | 0.545 ab | 0.528 ab | 0.01 | 0.01 |
| JCU2 (D. virgatus) | JCU4 (D. bicornutus) | |||||
|---|---|---|---|---|---|---|
| Durmic et al. [6] | Vandermeulen et al. [5] | Current Study | Durmic et al. [6] | Vandermeulen et al. [5] | Current Study | |
| CP (% DM) | 16.0 | 12.3 | 10.3 | 17.5 | 16.5 | 13.0 |
| NDF (% DM) | 50.0 | 52.2 | 57.5 | 48.1 | 42.6 | 53.1 |
| ADF (% DM) | 27.2 | 31.2 | 44.5 | 27.1 | 20.5 | 40.4 |
| ME (MJ/kg DM) | 8.68 | - | 6.5 | 8.56 | - | 7.2 |
| TP | - | 4.25% DM as tannic acid equivalent | 3.32% DM as catechin equivalent | - | 6.90% DM as tannic acid equivalent | 4.14% DM as catechin equivalent |
| CT | - | 2.09% DM as leucocyanidin equivalent | 4.10% DM | - | 3.81% DM as leucocyanidin equivalent | 7.27% DM |
| CH4 | 29.2 mL/g DMI | 23.1 mL/g OM | 21.0 g/kg DMI | 29.7 mL/g DMI | 21.9 mL/g OM | 20.8 g/kg DMI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suybeng, B.; Mwangi, F.W.; McSweeney, C.S.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O. Response to Climate Change: Evaluation of Methane Emissions in Northern Australian Beef Cattle on a High Quality Diet Supplemented with Desmanthus Using Open-Circuit Respiration Chambers and GreenFeed Emission Monitoring Systems. Biology 2021, 10, 943. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090943
Suybeng B, Mwangi FW, McSweeney CS, Charmley E, Gardiner CP, Malau-Aduli BS, Malau-Aduli AEO. Response to Climate Change: Evaluation of Methane Emissions in Northern Australian Beef Cattle on a High Quality Diet Supplemented with Desmanthus Using Open-Circuit Respiration Chambers and GreenFeed Emission Monitoring Systems. Biology. 2021; 10(9):943. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090943
Chicago/Turabian StyleSuybeng, Bénédicte, Felista W. Mwangi, Christopher S. McSweeney, Edward Charmley, Christopher P. Gardiner, Bunmi S. Malau-Aduli, and Aduli E. O. Malau-Aduli. 2021. "Response to Climate Change: Evaluation of Methane Emissions in Northern Australian Beef Cattle on a High Quality Diet Supplemented with Desmanthus Using Open-Circuit Respiration Chambers and GreenFeed Emission Monitoring Systems" Biology 10, no. 9: 943. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090943