


Memory Modulation by Exercise in Young Adults Is Related to Lactate and Not Affected by Sex or BDNF Polymorphism



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
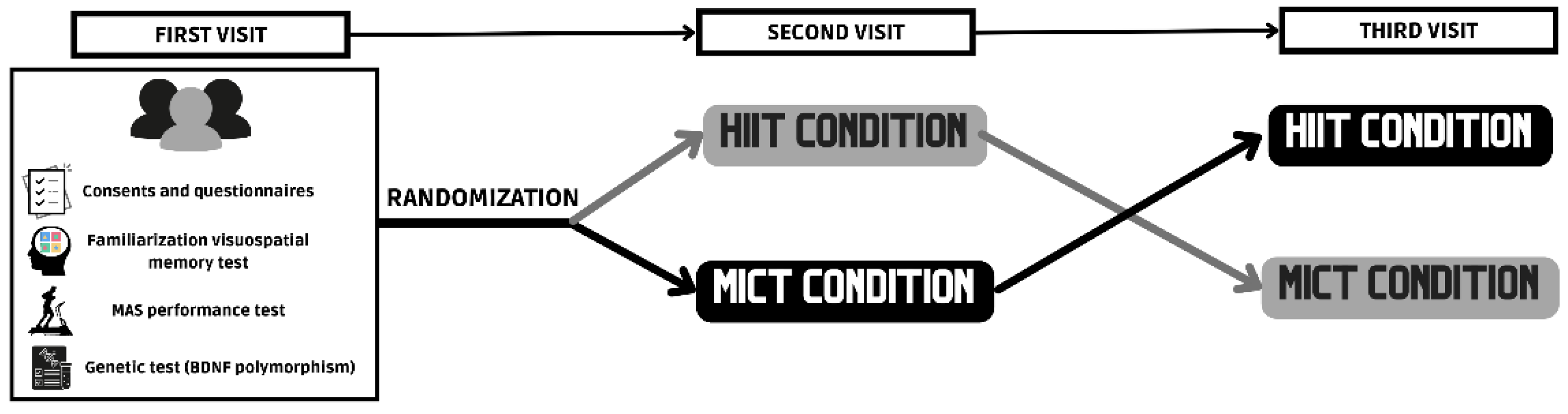
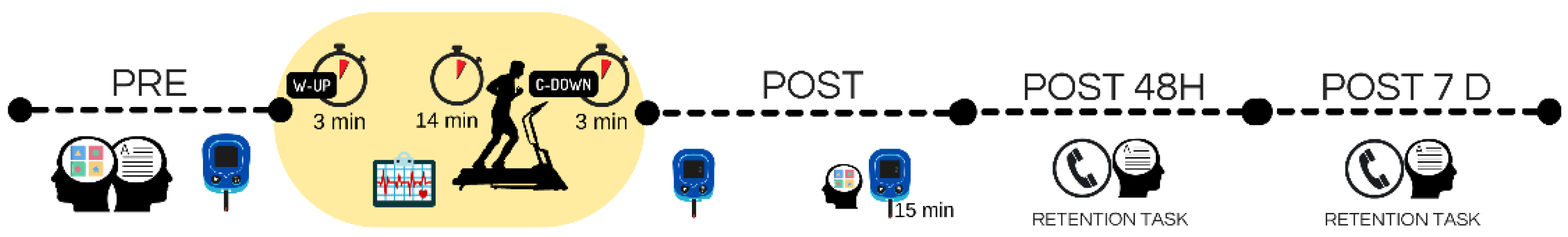
2.2. Experimental Procedure
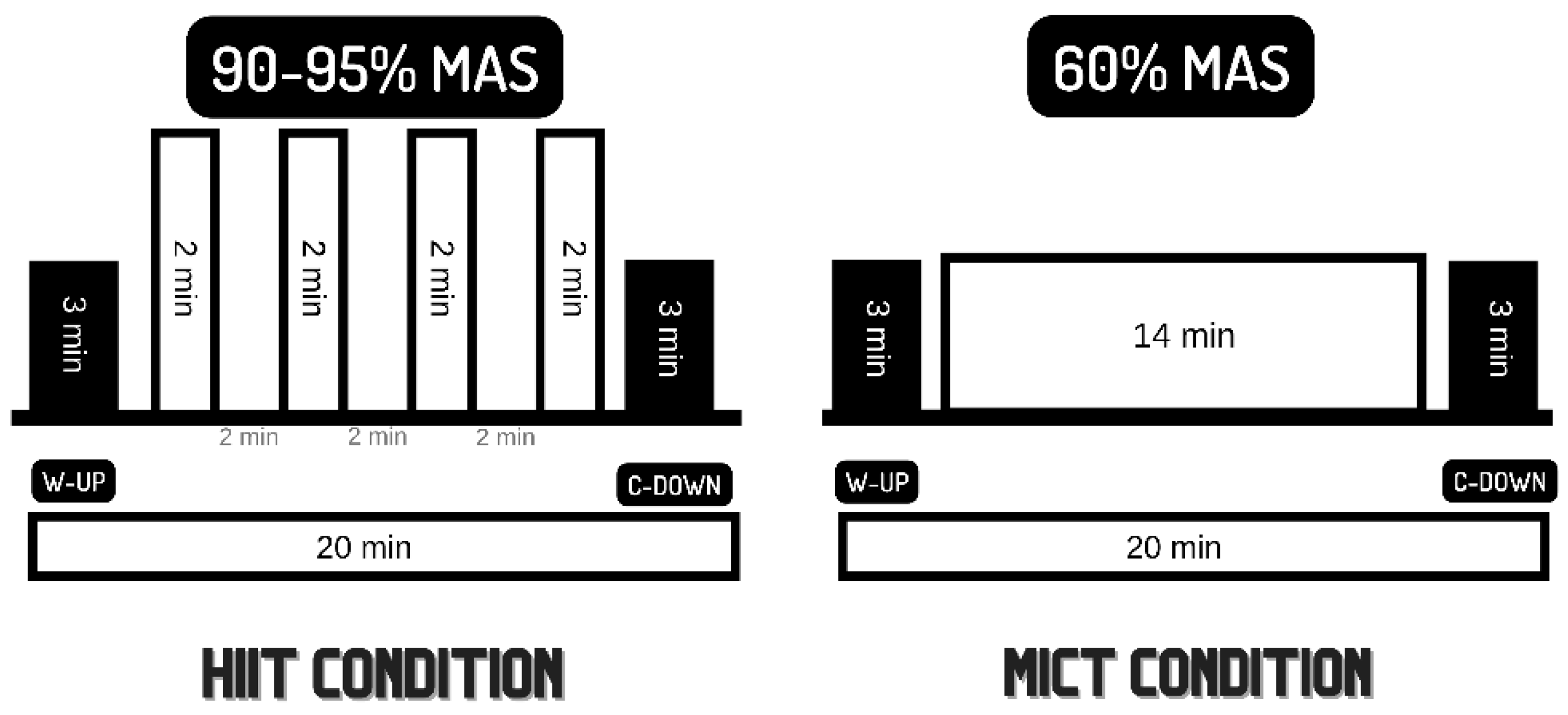
2.3. Experimental Conditions
2.4. Measurements
2.4.1. Graded Exercise Testing
2.4.2. Lactate Blood Sample
2.4.3. Genotype Analysis
2.4.4. Cognitive Function
Short-Term Visuospatial Memory Task
Long-Term Formal Memory Task
2.5. Statistical Analysis
3. Results
3.1. Exercise Characteristics
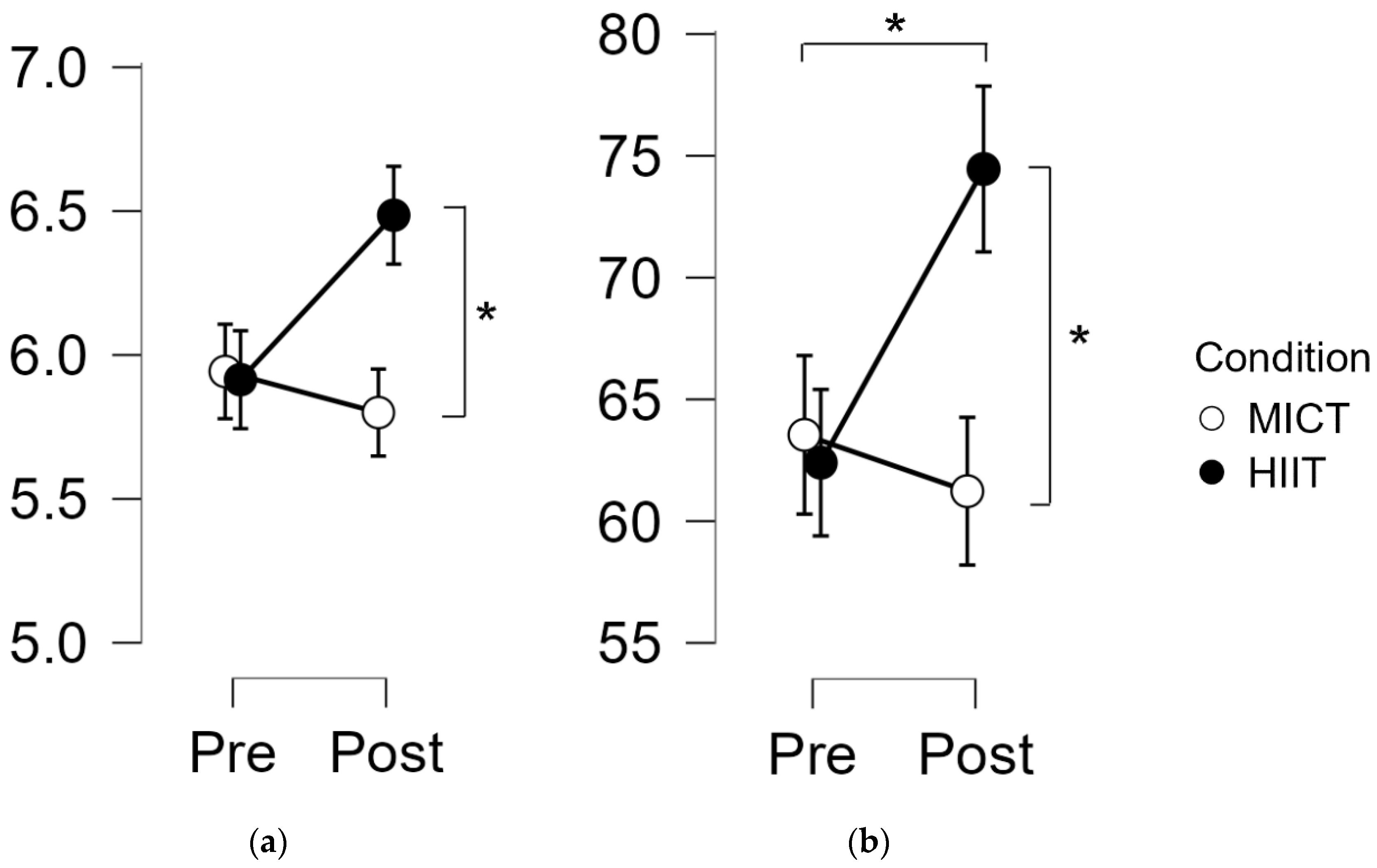
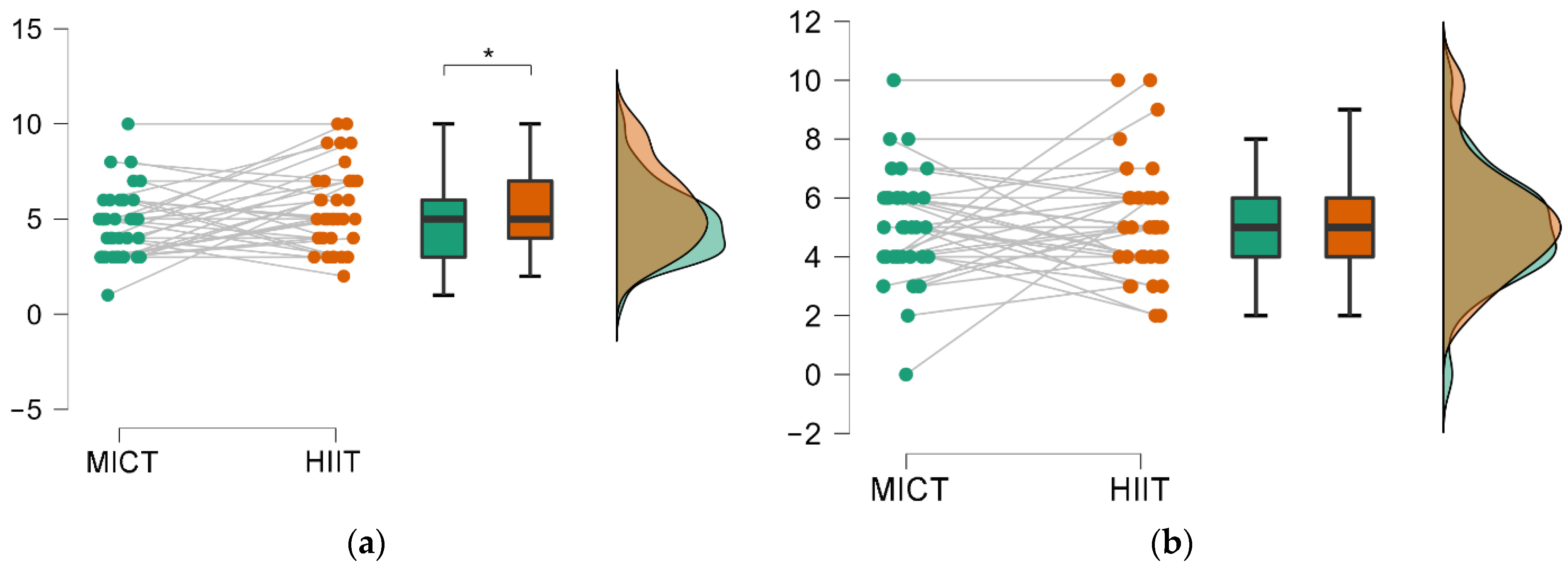
3.2. Intensity Impact on Memory
3.3. Interaction of Response with Sex and BDNF Polymorphism
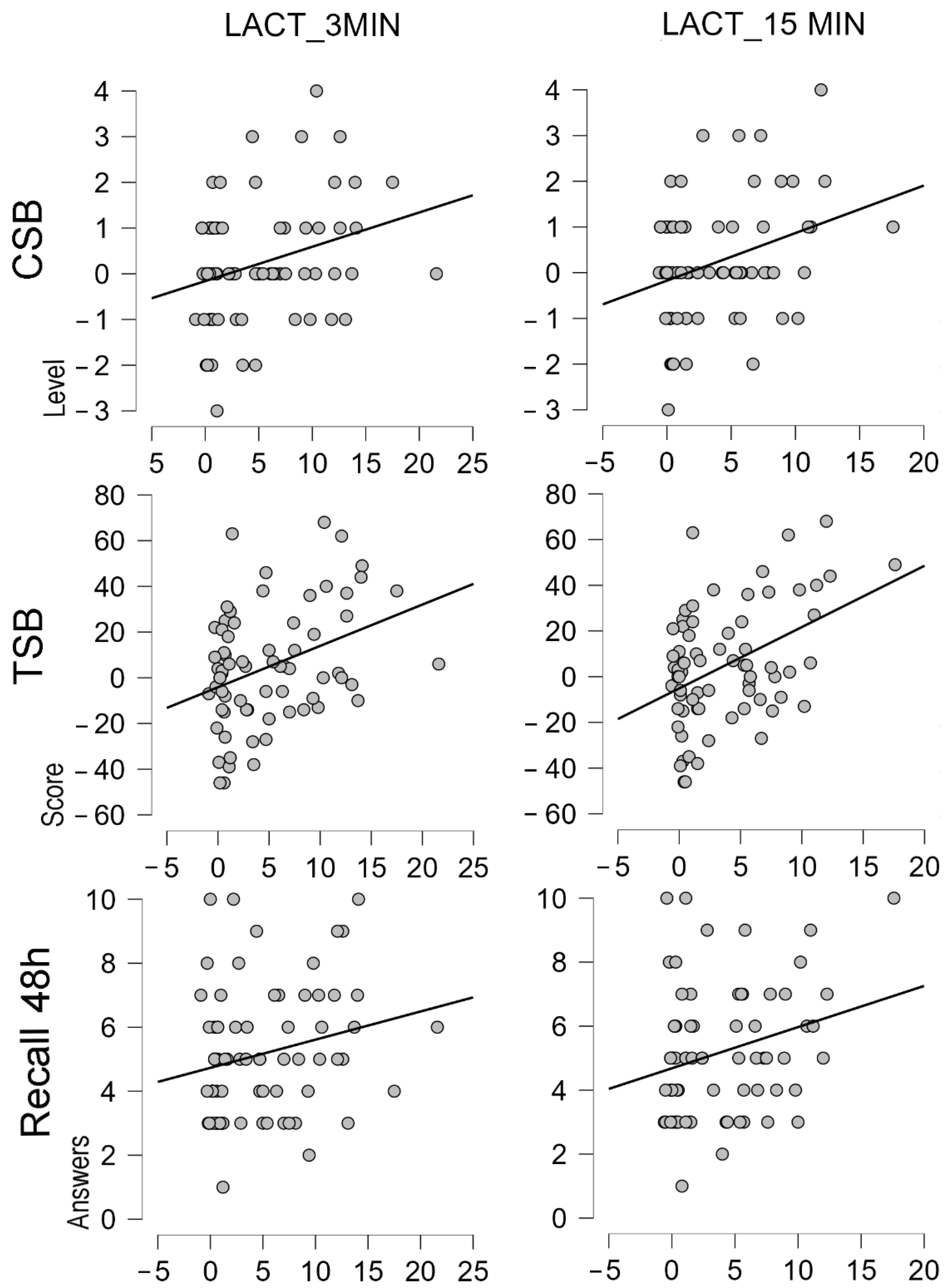
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, Y.K.; Labban, J.D.; Gapin, J.I.; Etnier, J.L. The effects of acute exercise on cognitive performance: A meta-analysis. Brain Res. 2012, 1453, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Herold, F.; Törpel, A.; Schega, L.; Müller, N.G. Functional and/or structural brain changes in response to resistance exercises and resistance training lead to cognitive improvements—A systematic review. Eur. Rev. Aging Phys. Act. Off. J. Eur. Group Res. Into Elder. Phys. Act. 2019, 16, 10. [Google Scholar] [CrossRef]
- Oberste, M.; Javelle, F.; Sharma, S.; Joisten, N.; Walzik, D.; Bloch, W.; Zimmer, P. Effects and Moderators of Acute Aerobic Exercise on Subsequent Interference Control: A Systematic Review and Meta-Analysis. Front. Psychol. 2019, 10, 2616. [Google Scholar] [CrossRef] [Green Version]
- Pontifex, M.B.; McGowan, A.L.; Chandler, M.C.; Gwizdala, K.L.; Parks, A.C.; Fenn, K.; Kamijo, K. A primer on investigating the after effects of acute bouts of physical activity on cognition. Psychol. Sport Exerc. 2019, 40, 1–22. [Google Scholar] [CrossRef]
- Soga, K.; Masaki, H.; Gerber, M.; Ludyga, S. Acute and Long-term Effects of Resistance Training on Executive Function. J. Cogn. Enhanc. 2018, 2, 200–207. [Google Scholar] [CrossRef]
- Stillman, C.M.; Cohen, J.; Lehman, M.E.; Erickson, K.I. Mediators of Physical Activity on Neurocognitive Function: A Review at Multiple Levels of Analysis. Front. Hum. Neurosci. 2016, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stillman, C.M.; Esteban-Cornejo, I.; Brown, B.; Bender, C.M.; Erickson, K.I. Effects of Exercise on Brain and Cognition Across Age Groups and Health States. Trends Neurosci. 2020, 43, 533–543. [Google Scholar] [CrossRef]
- Landrigan, J.F.; Bell, T.; Crowe, M.; Clay, O.J.; Mirman, D. Lifting cognition: A meta-analysis of effects of resistance exercise on cognition. Psychol. Res. 2020, 84, 1167–1183. [Google Scholar] [CrossRef] [Green Version]
- Ludyga, S.; Gerber, M.; Pühse, U.; Looser, V.N.; Kamijo, K. Systematic review and meta-analysis investigating moderators of long-term effects of exercise on cognition in healthy individuals. Nat. Hum. Behav. 2020, 4, 603–612. [Google Scholar] [CrossRef]
- Roig, M.; Nordbrandt, S.; Geertsen, S.S.; Nielsen, J.B. The effects of cardiovascular exercise on human memory: A review with meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 1645–1666. [Google Scholar] [CrossRef]
- de Las Heras, B.; Rodrigues, L.; Cristini, J.; Weiss, M.; Prats-Puig, A.; Roig, M. Does the Brain-Derived Neurotrophic Factor Val66Met Polymorphism Modulate the Effects of Physical Activity and Exercise on Cognition? Neurosci. A Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2022, 28, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Herold, F.; Müller, P.; Gronwald, T.; Müller, N.G. Dose-Response Matters!—A Perspective on the Exercise Prescription in Exercise-Cognition Research. Front. Psychol. 2019, 10, 2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Tsukamoto, H.; Ando, S.; Ogoh, S. Effect of Exercise on Brain Health: The Potential Role of Lactate as a Myokine. Metabolites 2021, 11, 813. [Google Scholar] [CrossRef] [PubMed]
- El Hayek, L.; Khalifeh, M.; Zibara, V.; Abi Assaad, R.; Emmanuel, N.; Karnib, N.; El-Ghandour, R.; Nasrallah, P.; Bilen, M.; Ibrahim, P.; et al. Lactate Mediates the Effects of Exercise on Learning and Memory through SIRT1-Dependent Activation of Hippocampal Brain-Derived Neurotrophic Factor (BDNF). J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 2369–2382. [Google Scholar] [CrossRef] [Green Version]
- Siebenmann, C.; Sørensen, H.; Bonne, T.C.; Zaar, M.; Aachmann-Andersen, N.J.; Nordsborg, N.B.; Nielsen, H.B.; Secher, N.H.; Lundby, C.; Rasmussen, P. Cerebral lactate uptake during exercise is driven by the increased arterial lactate concentration. J. Appl. Physiol. 2021, 131, 1824–1830. [Google Scholar] [CrossRef]
- An, L.; Li, X.; Tang, C.; Xu, N.; Sun, W. Hippocampal proBDNF facilitates place learning strategy associated with neural activity in rats. Brain Struct. Funct. 2018, 223, 4099–4113. [Google Scholar] [CrossRef]
- Bekinschtein, P.; Cammarota, M.; Medina, J.H. BDNF and memory processing. Neuropharmacology 2014, 76 Pt C, 677–683. [Google Scholar] [CrossRef]
- Morland, C.; Andersson, K.A.; Haugen, Ø.P.; Hadzic, A.; Kleppa, L.; Gille, A.; Rinholm, J.E.; Palibrk, V.; Diget, E.H.; Kennedy, L.H.; et al. Exercise induces cerebral VEGF and angiogenesis via the lactate receptor HCAR1. Nat. Commun. 2017, 8, 15557. [Google Scholar] [CrossRef] [Green Version]
- Lev-Vachnish, Y.; Cadury, S.; Rotter-Maskowitz, A.; Feldman, N.; Roichman, A.; Illouz, T.; Varvak, A.; Nicola, R.; Madar, R.; Okun, E. L-Lactate Promotes Adult Hippocampal Neurogenesis. Front. Neurosci. 2019, 13, 403. [Google Scholar] [CrossRef]
- Roig, M.; Thomas, R.; Mang, C.S.; Snow, N.J.; Ostadan, F.; Boyd, L.A.; Lundbye-Jensen, J. Time-Dependent Effects of Cardiovascular Exercise on Memory. Exerc. Sport Sci. Rev. 2016, 44, 81–88. [Google Scholar] [CrossRef]
- Loprinzi, P.D.; Roig, M.; Etnier, J.L.; Tomporowski, P.D.; Voss, M. Acute and Chronic Exercise Effects on Human Memory: What We Know and Where to Go from Here. J. Clin. Med. 2021, 10, 4812. [Google Scholar] [CrossRef] [PubMed]
- Skriver, K.; Roig, M.; Lundbye-Jensen, J.; Pingel, J.; Helge, J.W.; Kiens, B.; Nielsen, J.B. Acute exercise improves motor memory: Exploring potential biomarkers. Neurobiol. Learn. Mem. 2014, 116, 46–58. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Canivet, A.; Albinet, C.T.; André, N.; Pylouster, J.; Rodríguez-Ballesteros, M.; Kitzis, A.; Audiffren, M. Effects of BDNF polymorphism and physical activity on episodic memory in the elderly: A cross sectional study. Eur. Rev. Aging Phys. Act. 2015, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Barha, C.K.; Hsu, C.L.; Ten Brinke, L.; Liu-Ambrose, T. Biological Sex: A Potential Moderator of Physical Activity Efficacy on Brain Health. Front. Aging Neurosci. 2019, 11, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barha, C.K.; Liu-Ambrose, T. Sex differences in exercise efficacy: Is midlife a critical window for promoting healthy cognitive aging? FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 11329–11336. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-C.; Wang, S.-R.; Xu, X.-H. Sex differences in brain-derived neurotrophic factor signaling: Functions and implications. J. Neurosci. Res. 2017, 95, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 2015, 60, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, A.; Andrews, S.J.; Anstey, K.J. Sex Differences in the Impact of BDNF Genotype on the Longitudinal Relationship between Physical Activity and Cognitive Performance. Gerontology 2018, 64, 361–372. [Google Scholar] [CrossRef]
- Barha, C.K.; Davis, J.C.; Falck, R.S.; Nagamatsu, L.S.; Liu-Ambrose, T. Sex differences in exercise efficacy to improve cognition: A systematic review and meta-analysis of randomized controlled trials in older humans. Front. Neuroendocrinol. 2017, 46, 71–85. [Google Scholar] [CrossRef]
- Colcombe, S.; Kramer, A.F. Fitness effects on the cognitive function of older adults: A meta-analytic study. Psychol. Sci. 2003, 14, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romero, M.A.; Dorado, P.; Guarino, E.; Llerena, A. Development of a new genotyping assay for detection of the BDNF Val66Met polymorphism using melting-curve analysis. Pharmacogenomics 2009, 10, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Kessels, R.P.; van Zandvoort, M.J.; Postma, A.; Kappelle, L.J.; de Haan, E.H. The Corsi Block-Tapping Task: Standardization and normative data. Appl. Neuropsychol. 2000, 7, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, I.; Bevilaqua, L.R.; Rossato, J.I.; Lima, R.H.; Medina, J.H.; Cammarota, M. Age-dependent and age-independent human memory persistence is enhanced by delayed posttraining methylphenidate administration. Proc. Natl. Acad. Sci. USA 2008, 105, 19504–19507. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis. Curr. Dir. Psychol. Sci. 1992, 1, 98–101. [Google Scholar] [CrossRef]
- Vandierendonck, A.; Kemps, E.; Fastame, M.C.; Szmalec, A. Working memory components of the Corsi blocks task. Br. J. Psychol. 2004, 95, 57–79. [Google Scholar] [CrossRef]
- Brunetti, R.; Del Gatto, C.; Delogu, F. eCorsi: Implementation and testing of the Corsi block-tapping task for digital tablets. Front. Psychol. 2014, 5, 939. [Google Scholar] [CrossRef]
- Tomporowski, P.D.; Ellis, N.R.J.P.B. Effects of exercise on cognitive processes: A review. Psychol. Bull. 1986, 99, 338. [Google Scholar] [CrossRef]
- Etnier, J.L.; Salazar, W.; Landers, D.M.; Petruzzello, S.J.; Han, M.; Nowell, P. The Influence of Physical Fitness and Exercise upon Cognitive Functioning: A Meta-Analysis. J. Sport Exerc. Psychol. 1997, 19, 249–277. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, H.; Suga, T.; Takenaka, S.; Tanaka, D.; Takeuchi, T.; Hamaoka, T.; Isaka, T.; Hashimoto, T. Greater impact of acute high-intensity interval exercise on post-exercise executive function compared to moderate-intensity continuous exercise. Physiol. Behav. 2016, 155, 224–230. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Takenaka, S.; Suga, T.; Tanaka, D.; Takeuchi, T.; Hamaoka, T.; Isaka, T.; Hashimoto, T. Impact of Exercise Intensity and Duration on Postexercise Executive Function. Med. Sci. Sport. Exerc. 2017, 49, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Tsukamoto, H.; Takenaka, S.; Olesen, N.D.; Petersen, L.G.; Sørensen, H.; Nielsen, H.B.; Secher, N.H.; Ogoh, S. Maintained exercise-enhanced brain executive function related to cerebral lactate metabolism in men. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin Bosch, B.; Bringard, A.; Logrieco, M.G.; Lauer, E.; Imobersteg, N.; Thomas, A.; Ferretti, G.; Schwartz, S.; Igloi, K. A single session of moderate intensity exercise influences memory, endocannabinoids and brain derived neurotrophic factor levels in men. Sci. Rep. 2021, 11, 14371. [Google Scholar] [CrossRef]
- Saucedo Marquez, C.M.; Vanaudenaerde, B.; Troosters, T.; Wenderoth, N. High-intensity interval training evokes larger serum BDNF levels compared with intense continuous exercise. J. Appl. Physiol. 2015, 119, 1363–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMorris, T. Developing the catecholamines hypothesis for the acute exercise-cognition interaction in humans: Lessons from animal studies. Physiol. Behav. 2016, 165, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.C.; Chu, C.H.; Chu, I.H.; Chan, K.H.; Chang, Y.K. Executive function during acute exercise: The role of exercise intensity. J. Sport Exerc. Psychol. 2013, 35, 358–367. [Google Scholar] [CrossRef]
- Proia, P.; Di Liegro, C.M.; Schiera, G.; Fricano, A.; Di Liegro, I. Lactate as a Metabolite and a Regulator in the Central Nervous System. Int. J. Mol. Sci. 2016, 17, 1450. [Google Scholar] [CrossRef] [Green Version]
- van Hall, G.; Strømstad, M.; Rasmussen, P.; Jans, O.; Zaar, M.; Gam, C.; Quistorff, B.; Secher, N.H.; Nielsen, H.B. Blood lactate is an important energy source for the human brain. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2009, 29, 1121–1129. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, P.; Wyss, M.T.; Lundby, C. Cerebral glucose and lactate consumption during cerebral activation by physical activity in humans. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 2865–2873. [Google Scholar] [CrossRef]
- Schurr, A.; West, C.A.; Rigor, B.M. Lactate-supported synaptic function in the rat hippocampal slice preparation. Science 1988, 240, 1326–1328. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Ruchti, E.; Petit, J.M.; Jourdain, P.; Grenningloh, G.; Allaman, I.; Magistretti, P.J. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc. Natl. Acad. Sci. USA 2014, 111, 12228–12233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, R.; Moriguchi, Y.; Yamashita, F.; Mori, T.; Nemoto, K.; Okada, T.; Hori, H.; Noguchi, H.; Kunugi, H.; Ohnishi, T. Dose-dependent effect of the Val66Met polymorphism of the brain-derived neurotrophic factor gene on memory-related hippocampal activity. Neurosci. Res. 2008, 61, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.M.; Pereira, J.R.; Pires de Andrade, L.; Garuffi, M.; Ayan, C.; Kerr, D.S.; Talib, L.L.; Cominetti, M.R.; Stella, F. Physical exercise improves peripheral BDNF levels and cognitive functions in mild cognitive impairment elderly with different bdnf Val66Met genotypes. J. Alzheimer’s Dis. JAD 2015, 43, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; Campbell, K.L.; Ross, C.J.; Boyd, L.A. Promoting neuroplasticity for motor rehabilitation after stroke: Considering the effects of aerobic exercise and genetic variation on brain-derived neurotrophic factor. Phys. Ther. 2013, 93, 1707–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, D.; Kirk, I.J.; Waldie, K.E. High-intensity training enhances executive function in children in a randomized, placebo-controlled trial. eLife 2017, 6, e25062. [Google Scholar] [CrossRef]
- Iuliano, E.; di Cagno, A.; Aquino, G.; Fiorilli, G.; Mignogna, P.; Calcagno, G.; Di Costanzo, A. Effects of different types of physical activity on the cognitive functions and attention in older people: A randomized controlled study. Exp. Gerontol. 2015, 70, 105–110. [Google Scholar] [CrossRef]
- Chang, Y.K.; Chu, C.H.; Wang, C.C.; Wang, Y.C.; Song, T.F.; Tsai, C.L.; Etnier, J.L. Dose-response relation between exercise duration and cognition. Med. Sci. Sport. Exerc. 2015, 47, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.K.; Chen, F.T.; Kuan, G.; Wei, G.X.; Chu, C.H.; Yan, J.; Chen, A.G.; Hung, T.M. Effects of Acute Exercise Duration on the Inhibition Aspect of Executive Function in Late Middle-Aged Adults. Front. Aging Neurosci. 2019, 11, 227. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballester-Ferrer, J.A.; Roldan, A.; Cervelló, E.; Pastor, D. Memory Modulation by Exercise in Young Adults Is Related to Lactate and Not Affected by Sex or BDNF Polymorphism. Biology 2022, 11, 1541. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101541
Ballester-Ferrer JA, Roldan A, Cervelló E, Pastor D. Memory Modulation by Exercise in Young Adults Is Related to Lactate and Not Affected by Sex or BDNF Polymorphism. Biology. 2022; 11(10):1541. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101541
Chicago/Turabian StyleBallester-Ferrer, Juan Arturo, Alba Roldan, Eduardo Cervelló, and Diego Pastor. 2022. "Memory Modulation by Exercise in Young Adults Is Related to Lactate and Not Affected by Sex or BDNF Polymorphism" Biology 11, no. 10: 1541. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101541