
Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress


1
Dipartimento di Scienze e Innovazione Tecnologica, Università del Piemonte Orientale, Viale T. Michel 11, 15121 Alessandria, Italy
2
Department of Biology, University of Waterloo, Waterloo, ON N2L 3G1, Canada
*
Author to whom correspondence should be addressed.
Biology 2022, 11(3), 437; https://0-doi-org.brum.beds.ac.uk/10.3390/biology11030437
Submission received: 12 February 2022
/
Revised: 10 March 2022
/
Accepted: 10 March 2022
/
Published: 12 March 2022
(This article belongs to the Collection Plant Growth-Promoting Bacteria: Mechanisms and Applications)
Abstract
:Simple Summary
Salt and drought stress cause enormous crop losses worldwide. Several different approaches may be taken to address this problem, including increased use of irrigation, use of both traditional breeding and genetic engineering to develop salt-tolerant and drought-resistant crop plants, and the directed use of naturally occurring plant growth-promoting bacteria. Here, the mechanisms used by these plant growth-promoting bacteria are summarized and discussed. Moreover, recently reported studies of the effects that these organisms have on the growth of plants in the laboratory, the greenhouse, and the field under high salt and/or drought conditions is discussed in some detail. It is hoped that by understanding the mechanisms that these naturally occurring plant growth-promoting bacteria utilize to overcome damaging environmental stresses, it may be possible to employ these organisms to increase future agricultural productivity.
Abstract
The recent literature indicates that plant growth-promoting bacteria (PGPB) employ a range of mechanisms to augment a plant’s ability to ameliorate salt and drought stress. These mechanisms include synthesis of auxins, especially indoleacetic acid, which directly promotes plant growth; synthesis of antioxidant enzymes such as catalase, superoxide dismutase and peroxidase, which prevents the deleterious effects of reactive oxygen species; synthesis of small molecule osmolytes, e.g., trehalose and proline, which structures the water content within plant and bacterial cells and reduces plant turgor pressure; nitrogen fixation, which directly improves plant growth; synthesis of exopolysaccharides, which protects plant cells from water loss and stabilizes soil aggregates; synthesis of antibiotics, which protects stress-debilitated plants from soil pathogens; and synthesis of the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase, which lowers the level of ACC and ethylene in plants, thereby decreasing stress-induced plant senescence. Many of the reports of overcoming these plant stresses indicate that the most successful PGPB possess several of these mechanisms; however, the involvement of any particular mechanism in plant protection is nearly always inferred and not proven.
1. Drought and Salt Stress
Plants that are grown in the field under natural conditions are typically subjected to a wide range of both biotic and abiotic stresses, any one of which may significantly inhibit plant growth and development [1]. Biotic stress factors that are inhibitory to plant growth and development include viruses, nematodes, insects, phytopathogenic bacteria, and phytopathogenic fungi. In addition, abiotic stress factors that are detrimental to plants include extremes of temperature, excessively high or low light conditions, flooding, drought, high salt, toxic metals, organic contaminants, and excessive radiation. This review focuses on the effects and remediation of drought and salt stress, two of the most problematic abiotic stresses as far as plant growth is concerned.
Soil salinity and drought are enormous global problems for the growth of agricultural plants. Worldwide drought has increased dramatically in recent years because of the increasing impacts of climate change (https://climate.nasa.gov/news/3117/drought-makes-its-home-on-the-range/, accessed on 5 January 2022; https://www.c2es.org/content/drought-and-climate-change/, accessed on 5 January 2022). Soil salinity is not only affected by climate change but is especially problematic for crops that require irrigation. Because of the lack of sufficient water (i.e., drought) in many locations, some farmlands are under-irrigated, causing salt (from the irrigation water) to accumulate in the soil. In this case, the salt remains in the soil when the irrigation water is either utilized by plants or is lost to evaporation. Moreover, the majority of the world’s salt-inhibited soils are found in arid or semi-arid climates (https://www.fao.org/global-soil-partnership/resources/highlights/detail/en/c/1412475/, accessed on 5 January 2022). Compounding the negative effects of drought, the high salt levels that result from irrigation are inhibitory to the growth of a large number of plants. At present, it is estimated that >900 million hectares or ~20% of the world’s cultivated land mass is negatively impacted by high levels of salt. Moreover, a significant fraction of the global land mass used for the growth of irrigated crops is at least somewhat adversely affected by high levels of salinity.
The initial responses of most plants to drought and salinity are similar; both responses are largely due to water stress within the plant [2,3]. When plants are first exposed to high levels of salt, a decrease in their growth rate occurs. This is often followed by a slow recovery to a lower growth rate. Subsequently, following the continued uptake of salt by plants, sodium ions are translocated through the xylem to the plant shoots, with leaves and shoots eventually accumulating high levels of sodium ions. The toxicity of accumulating sodium ions in plants is generally considered to be a function of the excessive amounts of sodium ions that compete with potassium ions for the binding sites that are necessary for plant cellular functioning. This abiotic stress results in the generation of a number of reactive oxygen species within the plant [4], which in turn can cause electrolyte leakage from plant cells, plant cell membrane lipid peroxidation, an increase in photorespiration, a decrease in transpiration, eventual pollen sterility, and a decrease in the plant’s rate of photosynthesis, all of which negatively affect the plant yield and quality.
Plants are able to use a range of mechanisms to deal with salt stress [2,5,6,7,8,9,10,11,12,13,14], including (i) selective accumulation (in vacuoles) or exclusion of sodium ions, (ii) modulation of the root uptake of sodium ions and the subsequent transport of these ions into leaves, (iii) compartmentalization of sodium ions at the plant cellular level, (iv) synthesis of a variety of compatible solutes such as trehalose and proline, (v) modification of plant cell membranes, (vi) synthesis of various antioxidative enzymes, including superoxide dismutase and peroxidase, and (vii) modulation of some plant hormone levels, including auxin, cytokinin, and ethylene. Moreover, the salt tolerance of any particular plant species is often a function of the specific salt tolerance of the cultivar of the plant being examined, the growth phase of the plant, the soil composition, the plant’s health, the presence and nature of plant pathogenic organisms, the presence of specific rhizospheric or endophytic plant growth-promoting bacteria (PGPB), and the presence of mycorrhizae [1].
In considering the different PGPB mechanisms that can be employed (elaborated in the following section), it is necessary to bear in mind that many of these activities and their regulation are interconnected. Moreover, different PGPB contain different sets of genes that enable them to provide a range of varied protective responses against the inhibitory effects of a range of abiotic (and biotic) stresses, including drought and salinity. In addition to the more direct effects of PGPB on plants, many PGPB can modify plant gene expression, thereby increasing the plant’s synthesis of stress protective agents. Thus, for example, some PGPB may promote an increase in the plant’s production of “water-structuring” metabolites (osmolytes), such as betaine, proline, and trehalose, and the synthesis of reactive oxygen detoxifying enzymes, such as superoxide dismutase and catalase [15].
In assessing the scientific literature, it should be pointed out that the ability of PGPB to overcome the effects of salt stress on plant growth has been studied to a much greater extent than the ability of PGPB to ameliorate drought stress. This somewhat disparate focus may reflect the fact that salt stress is technically easier to impose and study in a laboratory setting and is of more universal interest than drought stress. This notwithstanding, numerous studies have reported success in using PGPB to ameliorate some of the deleterious effects of both salt and drought stress.
Interestingly, drought and/or salt stress often increases the sensitivity of many plants to various phytopathogens, often by decreasing the plant’s ability to effectively mount an attack against the pathogen. However, it has been observed that PGPB isolated from salt- or drought-stressed soils, in addition to the previously mentioned ability to protect plants against these abiotic stresses, can frequently effectively protect plants against the damage caused by many different fungal phytopathogens [16,17,18].
In the examples given below, conferring plants with drought and/or salt tolerance is often attributed to one or more well-known PGPB mechanisms; this is despite the fact that drought and/or salt tolerance in nature is probably attributable to multiple mechanisms. In addition, the scientific literature also contains numerous studies in which the mechanism used by a PGPB strain to confer drought and/or salt tolerance to a plant is not known. While these strains may be useful in some limited applications, since other researchers do not know what mechanisms they are using, the isolated strains are only useful to those who have isolated and identified them, since they do not provide any guidelines for selecting other (possibly similar or more efficacious) PGPB that are better at conferring salt and/or drought tolerance to treated plants.
2. Plant Growth-Promoting Bacteria
A very large number of bacteria are typically found in soil (~108 to 109 bacterial cells per gram of soil), although drought- or salt-stressed soils often contain as little as 106 bacterial cells per gram of soil [19]. Moreover, the bacteria are not evenly distributed throughout the soil. That is, much greater numbers of bacteria are usually found around the roots of plants than in the bulk soil. This is a consequence of the fact that plant roots generally exude a large amount sugars, amino acids, and small organic acids, and different types of these small molecules can serve as a nutrient source for many different soil bacteria [20]. The bacteria in the soil are made up of a range of different genera and species, some of which can promote plant growth (i.e., PGPB), some of which can inhibit plant growth (i.e., phytopathogens), and some of which have no discernible effect on plant growth.
PGPB stimulation of plant growth by may occur by a variety of either direct or indirect mechanisms. The direct promotion of plant growth by PGPB may occur by the bacterium, providing a plant with one or more phytohormones, such as auxin, cytokinin, or gibberellin, or by improving the ability of a plant to acquire soil nutrients by providing plants with sequestered iron, solubilized phosphate, zinc and potassium, and fixed nitrogen. In addition, PGPB that express the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase can lower plant ACC and ethylene levels and hence significantly increase a plant’s tolerance to a wide range of environmental ethylene-generating stresses [11,21,22,23,24]. Importantly, PGPB may possess one or more of these activities, with most PGPB possessing a few of these activities.
The indirect promotion of plant growth by PGPB happens when the PGPB acts as a biocontrol agent and can prevent or thwart phytopathogens (usually either fungi or bacteria) from inhibiting plant growth. This may occur in several different ways, including the synthesis of pathogen-inhibiting antibiotics and/or hydrogen cyanide by the PGPB; the synthesis of any one of several different pathogenic fungal cell wall lytic enzymes; the synthesis of siderophores that bind extremely tightly to soil Fe+++, resulting in the pathogen (that has a much lower affinity for iron) being deprived of necessary iron; the PGPB outcompeting the phytopathogen for sites on or near the plant root; the synthesis of plant hormones by the PGPB, thereby promoting plant growth and strengthening the plant’s ability to fight off the attacking phytopathogen; the PGPB decreasing the amount of pathogen-induced stress ethylene within a plant; the synthesis of volatile organic compounds (VOC) that are toxic to a variety of pathogenic organisms; and the PGPB inducing the plant’s own defenses against phytopathogens (i.e., Induced Systemic Resistance) [1].
Several of the direct mechanisms mentioned above have previously been shown to protect plants against both drought and salt stress [10,13,25,26,27,28]. For example, cytokinin, auxin, and ethylene levels have all been directly implicated in playing a role in how plants respond to drought and salt stress [1,2,10,22,29]. In addition, indirect mechanisms of plant growth promotion have been shown to protect a salt- or drought-debilitated plant against the phytopathogens present during periods of salt or drought stress [18].
PGPB and the mechanisms that they employ have been extensively studied over the past 20–25 years, and they have been shown to be effective at facilitating plant growth and development under a wide range of potentially inhibitory conditions, with many different species of plants, and under laboratory, greenhouse, and field conditions. Moreover, several of the isolated and characterized PGPB strains have been commercialized in several different countries [1,30]. Although PGPB presently make up only a very small portion of the global fertilizer market, with each succeeding year, more of these bacteria are being used in sustainable agricultural practice. It is therefore expected that PGPB will eventually (hopefully in the next 5–15 years) be utilized on a global scale and replace many of the environmentally deleterious chemicals that are currently used in agriculture.
2.1. How PGPB Mechanisms Deal with Plant Drought and Salt Stress
2.1.1. Mechanisms: ACC Deaminase, IAA, Cytokinin, and Metabolites Such as Proline and Trehalose
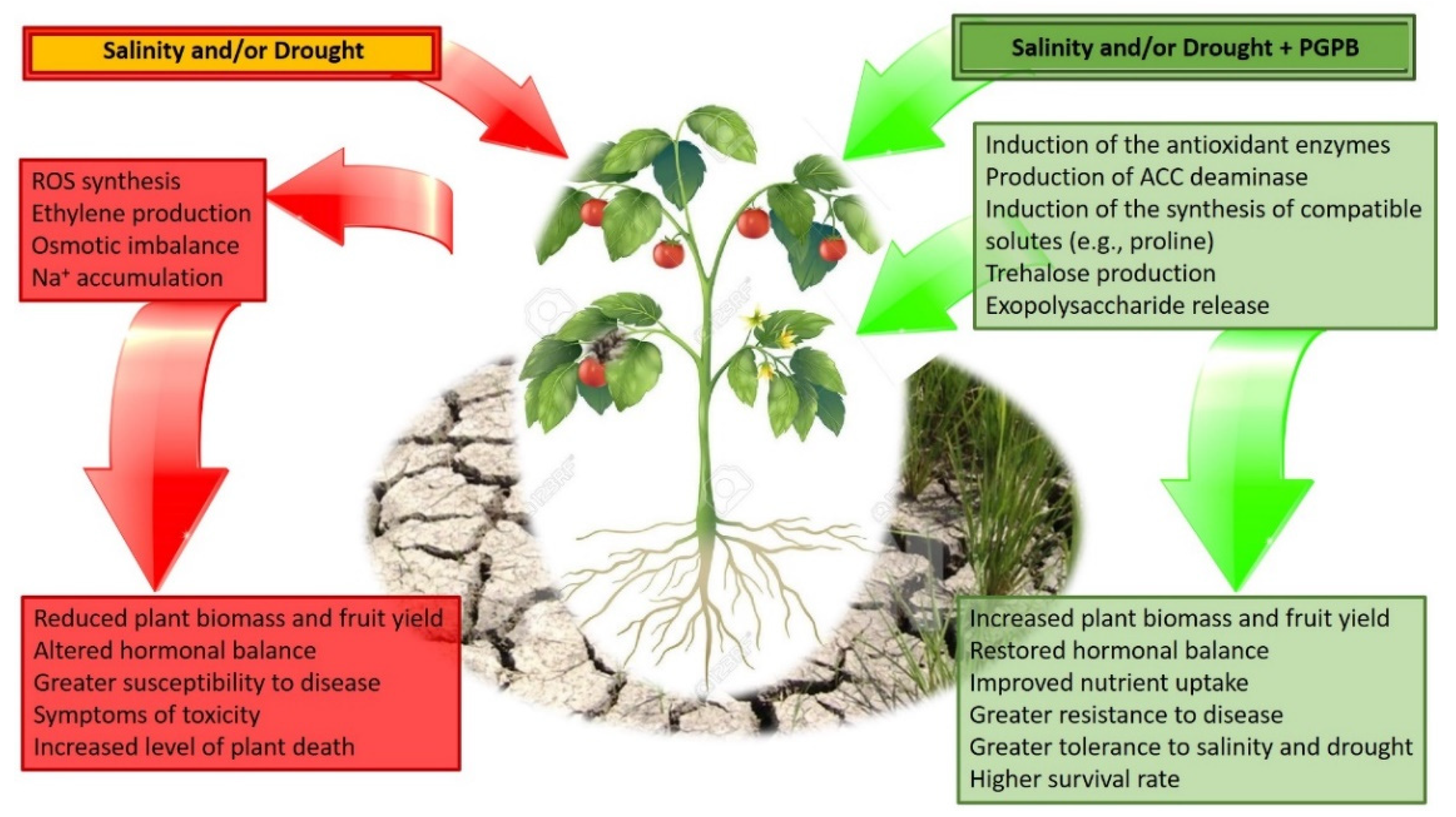
PGPB exhibit a wide range of mechanisms through which they can improve the tolerance of plants to salt and drought stress. After a careful analysis of the existing literature, some of the most common traits involved are (i) the synthesis of the enzyme ACC deaminase, lowering the amount of stress ethylene produced by plants; (ii) the release of auxins and especially IAA, modulating the regulatory responses of plants that have been exposed to various environmental stresses; (iii) the synthesis of metabolites such as proline and trehalose, well known to behave as protectants against various stresses (particularly drought and salt stress; Figure 1). The protective effect of these bacterial mechanisms against abiotic stresses has been extensively tested and validated for a large number of plant and bacterial species. In the following section, a description of the functioning of these mechanisms is presented.
2.1.2. Details of How Each Mechanism Functions
The plant hormone ethylene is involved in several plant biological activities, including germination of seeds, tissue differentiation, the regulation of root branching and root elongation, flowering, fruit ripening, and leaf abscission. However, when plants are subjected to biotic or abiotic environmental stresses, the synthesis of a high amount of ethylene occurs. This synthesis starts from S-Adenosyl-Met; this molecule is converted into ACC by ACC synthase, and the ACC is subsequently converted into ethylene by the enzyme ACC oxidase [31]. ACC deaminase produced by PGPB can lower ACC and ethylene levels in a plant via the degradation of ACC to ammonia and alpha-keto-butyrate. As a consequence, the ethylene level inside a plant will not reach concentrations that are inhibitory to plant growth [32,33]. There is a body of literature demonstrating the efficacy of PGPB in producing ACC deaminase [34], supporting plant growth in the presence of salt or/and drought stresses [1,2] (see Table 1 and Table 2). The clear involvement of ACC deaminase in facilitating plant growth under water deprivation or the excess of salt is typically demonstrated with the use of mutants lacking this enzymatic activity [25,34]. Moreover, ACC deaminase is often associated with the expression of other PGPB plant-beneficial traits, such as IAA synthesis [33], and the production of osmoprotectant molecules [35]. Auxins are phytohormones whose effect is dose-dependent, driving different stages of plant development, such as germination of seeds, xylematic vessel formation, the elongation and proliferation of plant cells, root branching (including the emergence of both lateral and adventitious roots), plant responses to light and gravity, photosynthesis, florescence and fructification, and tolerance to stressful conditions [36]. Soil bacteria able to synthesize IAA are quite common and belong to a wide variety of taxonomic groups. Bacterial biosyntheis of IAA may occur through one or more of seven different biosynthetic pathways that have been identified, with five of these pathways relying on tryptophan as a precursor of IAA [37,38]. Usually, since amino acid synthesis is costly from an energetic point of view, the tryptophan concentration inside a bacterial cell is low and is strictly regulated. For this reason, to synthesize IAA, bacterial cells primarily use the tryptophan released through exudation by the plant roots. IAA produced by bacteria may be taken up by plant roots, augmenting the endogenous plant pool. The increased IAA generally has the effect of stimulating plant growth (or suppressing plant growth if the IAA level becomes too high).
IAA-producing bacteria mediate drought and salt stress tolerance by improving root structure architecture, increasing the permeability of water into the cell and the water uptake in leaves, regulating metabolic homeostasis by increasing osmotic content, ROS detoxification, and inducing the transcription of a plethora of stress-related genes combined with the induction of specific protein synthesis [39,40].
Following several previous experiments using transgenic plants, a strain of the Gram-negative bacterium Ensifer meliloti was engineered with an ipt gene from a strain of Agrobacterium under the control of the E. coli trp promoter, which causes the overproduction of the phytohormone cytokinin [41]. The genetically engineered bacteria were then assessed to determine whether they could protect alfalfa plants against the growth inhibition and senescence that is a consequence of drought stress. Alfalfa plants were first inoculated with engineered E. meliloti and then subjected to several weeks of drought stress. Following this stress, the plants inoculated with the transformed PGPB strain were significantly larger than the plants inoculated with the non-transformed strain. When the plants were later rewatered, those that had been inoculated with the engineered PGPB strain grew to a level similar to plants that had not been subjected to drought stress, suggesting that the high levels of bacterial cytokinin produced by the engineered bacterium improved the ability of alfalfa to withstand drought stress. Unfortunately, the use of genetically engineered bacterial strains in the field is not permitted in most countries; however, it may be possible to alter rhizobial strains to produce high cytokinin levels by either conventional mutagenesis or CRISPR modification of the bacterial genome (which is not considered to be genetic engineering in many countries).
Trehalose (α-D-glucopyranosyl-(1→1)-α-D-glucopyranoside) is a non-reducing disaccharide that consists of two glucose molecules linked by a α,α-1,1-glycosidic bond; it is found in archaea, bacteria, fungi, plants, and in many invertebrates. In bacteria, trehalose synthesis is realized through five different pathways, with only one consisting of a single reaction, e.g., in Pseudomonas putida UW4 and other bacterial species, and consisting of trehalose synthesis from maltose mediated by a transglucosylation reaction [42]. Trehalose in plant and bacterial cells behave as a xeroprotectant, through three hypothesized mechanisms (water replacement, glass formation, and stability theory), enabling them to deal with harsh abiotic stress such as desiccation and or high levels of salinity [43,44,45,46]. The environmental pressure induced by salinity or drought selects and favors tolerant plants and microorganisms. As a consequence, PGPB living on the root and able to tolerate arid or saline conditions can limit the damage caused by drought and/or salinity to the plants [47]. One of the mechanisms responsible for the induced plant tolerance to desiccation is the bacterial modulation of the amount of trehalose in plants. Trehalose behaves as a signal of drying damage due to its ability to trigger the plant defense system to limit the damages caused by drought. This was demonstrated by Vilchez et al. [48], who identified the otsAB genes (which codes for alpha, alpha-trehalose-phosphate synthase, and trehalose-6-phosphate phosphatase) in the drought-tolerant Microbacterium sp. strain 3J1 and inserted them into the drought-sensitive P. putida KT2440 strain. The transformed strain P. putida KT2440 (pUCP22:otsAB) overproduced trehalose under water stress conditions and efficiently supported the growth of pepper plants cultivated under water deficit. Moreover, several examples in the literature, reviewed by Sharma et al. [49], demonstrate that trehalose is a key component of the functioning of the legume-rhizobia-mycorrhizal fungi tripartite symbiosis under drought conditions.
In addition to increasing bacterial and plant tolerance to environmental stresses, trehalose improves the survival of PGPB or other bacterial biocontrol agents during long-term storage of commercial formulations and increases root competence [50]. Finally, trehalose has recently been classified by the US Food and Drug Administration as Generally Regarded As Safe (GRAS) [51], thus becoming useful as a food formulant.
Among the compatible solutes produced by the plant following water deprivation, proline reduces the cell water potential and helps to maintain the turgor pressure, thus ensuring the plant’s full range of plant metabolic activity, development, and yield rate. The amount of proline synthesized by different species of plants increases from 20% to 80% of the total number of free amino acids under optimal conditions and osmotic and salt stress, respectively [52]. Fortunately, PGPB can modulate proline expression in plants. For example, Pseudomonas putida strain GAP-P45 can promote the growth of Arabidopsis thaliana plants that have been exposed to drought, upregulating the expression of genes that are involved in proline production, including OAT (ornithine-Δ-aminotransferase), P5CS1 (Δ1-pyrroline-5-carboxylate synthetase 1), P5CR (Δ1-pyrroline5-carboxylate reductase), and in proline degradation, such as PHD1 (proline dehydrogenase 1) and P5CDH (Δ1–pyrroline 5-carboxylate dehydrogenase). These results, obtained by real-time PCR, are consistent with the improvement of the growth parameters (plant biomass, water content, chlorophyll concentration) observed in plants cultivated under water deficit and inoculated with this bacterial strain [53]. Similarly, salt tolerance in wheat induced by Bacillus sp. strain wp-6 was determined to be related to an increased content of proline, soluble sugar, and soluble protein of 7.48%, 12.34%, and 4.12%, respectively [54].

{kind=link}
Table 1.
Recent studies indicating how PGPB help to ameliorate drought stress.
| Plant | Bacteria | Comments | Reference |
|---|---|---|---|
| Arabidopsis thaliana (thale cress) | Flavobacterium crocinum HYN0056T | Inoculation with HYN0056T enhanced tolerance against drought and salt stress, possibly via induction of stomatal closure. Treatment with strain HYN0056T followed by drought or salt stress caused the upregulation of several drought- and salt-inducible Arabidopsis genes. | [55] |
| Arabidopsis thaliana (thale cress) | Kosakonia cowanii GG1 isolated as an endophyte of seeds of the xerophytic invasive plant Lactuca serriola | Inoculation of A. thaliana with K. cowanii GG1 stimulated plant growth under drought conditions. The bacterial strain reduced soil water loss, indicating that the synthesis of exopolysaccharides contributes to maintaining the soil water content. | [56] |
| C. arietinum L. (chickpea) | Mesorhizobium ciceri Ca181 | Mutants of M. ciceri defective in phosphorous solubilization and drought stress tolerance were selected. The results indicated that the otsA (Trehalose-6-phosphate synthase), Auc (Acetoin utilisation protein), and Usp (Universal stress protein) genes contributed to the mechanism of drought stress tolerance. | [57] |
| Eleusine coracana (L.) Gaertn. (finger millet) | Variovorax paradoxus RAA3 and ACC deaminase-producing bacteria (Ochrobactrum anthropi DPC9, Pseudomonas palleroniana DPB13 and DPB16, and Pseudomonas fluorescens DPB15) | Inoculation of plants with V. paradoxus RAA3 and the consortium of O. anthropi DPC9, P. palleroniana DPB13 and DPB16, and P. fluorescens DPB15 increased plant growth and nutrient levels in leaves. High amounts of ROS-scavenging enzymes, including superoxide dismutase, guaiacol peroxidase, catalase, and ascorbate peroxidase, as well as the cellular osmolytes proline and phenol, leaf chlorophyll, and a reduced level of hydrogen peroxide and malondialdehyde, were observed after inoculation with RAA3 and the consortium of the other four bacterial strains compared to untreated plants. | [58] |
| Eucalyptus grandis (rose gum) | Pseudomonas sp. M25 and N33 | Plants inoculated with strain M25 exposed to gradual water deficit showed a significant increase in plant water content and cell wall elasticity. Rapid water deficit conditions caused partial defoliation in the absence of added bacteria. Both PGPB strains stimulated the formation of new leaves; inoculation with strain M25 reduced the transpiration rate; and co-inoculation with both strains increased both growth and photosynthetic activity. | [59] |
| Glycine max L. Merrill (soybean) | Bradyrhizobium japonicum and Azospirillum brasilense | The inoculation of soybean plants with B. japonicum and/or A. brasilense and then subjected to drought stress yielded increased leaf membrane stability. Co-inoculation with these strains followed by drought stress improved nodulation. Treatment with one or both of these strains reduced the pod abortion rate under moderate drought stress but not severe drought stress. | [60] |
| Juglans regia L. (walnut.) | AM fungi (Funnelliformis mosseae, Claroideoglomus etunicatum), and Azotobacter chroococcum, Azospirillium lipoferum | Drought stress caused a reduction in plant growth and leaf nutrient content, while increased proline, soluble sugar, starch peroxidase enzyme activity, and phenolic content was seen in leaves. Inoculation with consortia alleviated the negative effects of drought stress on seedlings by increasing the phenol, proline, peroxidase activity, soluble sugar, and starch content. C. etunicatum was the most effective AM fungi. | [61] |
| Solanum lycopersicum Mill cv. F144 (tomato) and Capsicum annuum L. cv. Maor (pepper) | Achromobacter piechaudii ARV8 (isolated from the Arava region of the Negev desert, Israel) | Tomato and pepper seedlings exposed to transient water stress and inoculated with strain A. piechaudii ARV8 showed increased biomass. Moreover, ARV8 lowered ethylene synthesis in tomato seedlings during drought stress, favoring the recovery of ARV8-treated plants when watering was resumed. | [62] |
| Solanum lycopersicum L. (tomato) | Various rhizobacteria (12 Bacillus spp. strains, 6 Pseudomonas, 4 Brevibacillus and 1 Paenibacillus strain isolated from Cistanthe longiscapa (a native flowering desert plant from the Atacama desert, Chile) | The Bacillus strains were used to formulate three consortia and to inoculate tomato seeds that were subsequently exposed to different degrees of water limitation. Inoculated seedlings showed higher biomass and recovery rates compared to uninoculated ones. | [63] |
| Solanum tuberosum (cultivars Swift and Nevsky) (potato) | Achromobacter xylosoxidans Cm4, Pseudomonas oryzihabitans Ep4, and Variovorax paradoxus 5C-2 | PGPB inoculation increased tuber yield in field experiments in plants cultivated under water-limited conditions. However, the leaf water concentration both in inoculated and uninoculated plants was similar, suggesting that other mechanisms (such as the modulation of phytohormone levels) might be responsible for plant growth promotion. | [64] |
| Triticum aestivum L. (wheat) | Azospirillum brasilense and Herbaspirillum seropedicae | Inoculation with the bacterial strains induced drought resistance in the wheat cultivar CD-120. The grain index was improved with H. seropedicae under water stress conditions. | [65] |
| Triticum aestivum L. (wheat) | Variovorax paradoxus RAA3; Pseudomonas spp. DPC12, DPB13, DPB15, and DPB16; Achromobacter spp. PSA7 and PSB8; and Ochrobactrum anthropi DPC9 | In drought conditions, inoculation with strain RAA3 and a consortium of DPC9 + DPB13 + DPB15 + DPB16 improved wheat plant growth and foliar nutrient levels and positively modulated antioxidant properties compared to uninoculated plants. | [66] |
| Triticum aestivum L. (wheat) | Azospirillum brasilense NO40 and Stenotrophomonas maltophilia B11 | Seedling inoculation with the two bacterial strains overcame the negative effects of drought stress, including changes to the relative water content of roots, shoots, and leaves; the area of leaves; the contents of chlorophyll a and b and ascorbic acid; and the protein patterns of root extracts. Bacterial inoculation reduced the drought-induced negative changes (i.e., the leakage of electrolytes and accumulation of malondialdehyde and hydrogen peroxide, the production of proline, and the activities of catalase and peroxidase compared to their uninoculated counterparts). | [67] |
| Triticum aestivum (wheat) | Curtobacterium flaccumfaciens Cf D3-2 and Arthrobacter sp. Ar sp. D4-1 | The two bacterial strains, when used separately to inoculate wheat plants, showed the ability to promote growth under drought conditions. | [68] |
| Vigna mungo L. (black gam) and Pisum sativum L. (pea) | Ochrobactrum pseudogrignonense RJ12, Pseudomonas sp. RJ15, Bacillus subtilis RJ46, alone and in consortia | Consortium treatment increased seed germination, root and shoot length, and plant biomass. Under drought conditions, treated plants exhibited elevated ROS and cellular osmolyte synthesis, higher leaf chlorophyll content, and increased relative water content compared to uninoculated plants. Bacterial inoculation reduced ACC accumulation in plants and down-regulated ACC-oxidase gene expression. | [69] |
| Vigna unguiculata (cowpea) | Bacillus aryabhattai strain MoB09 (able to degrade the herbicide paraquat) | B. aryabhattai MoB09 promoted the growth of cowpea plants following drought stress. | [70] |
| Zea mays L. (maize) | 12 drought-tolerant bacterial strains producing ACC deaminase and/or exopolysaccharides | Strains that synthesize both ACC deaminase and exopolysaccharides induced increased photosynthesis rate, stomatal conductance, vapor pressure, water-use efficiency, and transpiration rate. The strain B. velezensis D3 was the best PGPB. | [71] |
| Zea mays L. (maize) | Commercial biostimulant BACSTIMR 100 composed of a consortium of two Bacillus licheniformis strains, two Brevibacillus laterosporus strains, and one Bacillus amyloliquefaciens strain | Plant inoculation conferred increaseed drought resistance in maize by altering several plant metabolic pathways, including pathways encoding redox homeostasis and strengthening of the plant cell wall, osmoregulation, energy production, and membrane remodelling. | [72] |
Table 2.
Recent studies indicating how PGPB help to ameliorate salt stress.
| Plant | Bacteria | Comments | Reference |
|---|---|---|---|
| Arabidopsis thaliana (thale cress) | Paraburkholderia phytofirmans PsJN | A. thaliana plants inoculated with strain PsJN showed higher survival rate when exposed to long-term salinity and reduced Na+ accumulation within leaf tissues compared to uninoculated plants. Mutants defective in ACC deaminase, auxin catabolism, N-acyl-homoserine-lactone production, and flagellin synthesis showed a low relevance of these functions to salinity tolerance. Bacterial release of volatile organic compounds (mainly 2-undecanone, 7-hexanol, 3-methylbutanol and dimethyl disulphide) reproduced the effects of direct bacterial inoculation of roots, increasing plant growth rate and tolerance to salt stress conditions. Exposure of A. thaliana to different amounts of these molecules demonstrated their capability to affect growth, while exposure to a mixture of the first three compounds mimicked the effects of the bacterial strain on plant growth stimulation and salinity tolerance. | [73] |
| Arachis hypogaea L. (peanut) | Klebsiella sp., Pseudomonas sp., Agrobacterium sp., and Ochrobactrum sp. isolated from the halophyte Arthrocnemum indicum | Five diazotrophic salt-resistant strains of Klebsiella, Pseudomonas, Agrobacterium, and Ochrobactrum produced IAA and ACC deaminase, fixed N2, and solubilized phosphate. All of the isolates promoted peanut growth under non-stressful conditions and increased the N content in plants. In plants that were previously inoculated with these bacterial strains and then exposed to salt stress, accumulation of ROS-modulating enzymes and increased biomass was recorded compared to uninoculated ones. | [74] |
| Brassica campestris L. (canola) | Brevibacterium epidermidis RS15 and Bacillus aryabhattai RS341 | 120 mM NaCl reduced the rate of seed germination by 50%. Inoculation with B. epidermidis RS15 and B. aryabhattai RS34, both halotolerant and able to synthesize ACC deaminase, enhanced seed germination under salt stress and reduced the ACC content in seeds. Inoculation with both bacterial strains increased hydrolytic enzyme activities (amylase, invertase, and protease) and decreased ethylene levels compared to uninoculated seeds exposed to salt stress. | [75] |
| Camelina sativa (camelina or false flax) | Pseudomonas putida UW4, two root endophytes Pseudomonas migulae 8R6 and Pseudomonas fluoresces YsS6 (both ACC deaminase producing strains), and the acdS minus mutants 8R6M and YsS6M | Soil inoculation with wild-type strains increased shoot length without salt, and seed yield under moderate salinity. Transgenic plants that expressed the acdS gene, encoding the enzyme ACC deaminase, showed reduced inhibition of root lengthening and biomass development, and increased seed production, better seed quality, and higher levels of seed oil production under salt stress. | [76] |
| Camelina sativa (camelina or false flax) | Pseudomonas migulae 8R6 | Both of the C. sativa plants treated with the ACC deaminase producing endophyte P. migulae 8R6 and transgenic plants expressing acdS demonstrated increased tolerance to salt. Inoculation with strain 8R6 positively impacted ethylene- and abscisic acid-dependent signalling. The expression of acdS in transgenic plants altered auxin, jasmonic acid, and brassinosteroid signalling and/biosynthesis. Expression of genes involved in carbohydrate metabolism were up-regulated, as was the expression of genes modulating the level of ROS released. The expression of the acdS gene also positively effected the expression of photosynthesis genes. | [77] |
| Camelina sativa (camelina or false flax) | Pseudomonas migulae 8R6 | Treatment of C. sativa, grown under salt stress, with the endophyte P. migulae 8R6, able to synthesize ACC deaminase, induced a negative modulation of ethylene signaling as well as auxin and jasmonic acid biosynthesis and signaling, while genes involved in regulation of gibberellin signaling were positively affected. In plants cultivated with salt and inoculated with 8R6, a moderate expression of the acdS gene under the control of the rolD promoter occurred, which was highly efficient in lowering the expression of the genes involved in the synthesis of ethylene and its signaling. | [78] |
| Capsicum annuum (pepper) | Brevibacterium iodinum RS16, Bacillus licheniformis RS656, and Zhihengliuela alba RS111 | Brevibacterium iodinum RS16, Bacillus licheniformis RS656, and Zhihengliuela alba RS111 were identified as both halotolerant and ACC deaminase producers. Single inoculation with the three bacterial strains in red pepper plants grown at three salinity levels induced lower ethylene production. Plant biomass and salt tolerance index (the ratio of the biomass of salt stressed to non-stressed plants) in inoculated plants was higher compared to non-inoculated plants. | [79] |
| Capsicum annuum L. (red pepper) | Pseudomonas frederiksbergensis OS261 | Plants were inoculated with strain OS261 and grown with three levels of salt. Growth parameters (height and plant biomass) of plants were increased by the presence of the bacterial strain compared to uninoculated controls. The amount of ethylene synthesized by plants grown under salinity stress was high, but inoculation with strain OS261 reduced the release of this hormone. The level of antioxidant enzyme activity in leaves of inoculated plants grown in salinity was increased, while the H+ concentration was reduced. | [80] |
| Capsicum annuum L. cv. Bulmat (red pepper) | Pseudomonas frederiksbergensis OB139, Pseudomonas vancouverensis OB155 | Plants were cultivated under four levels of salt concentration and inoculated or not with one or both strains. Salt stress inhibited plant growth through increased ethylene synthesis and the disruption of photosynthetic parameters compared to uninoculated plants. The combination of the two bacterial strains, both able to synthesize ACC deaminase, lowered ethylene levels in plants and increased catalase activity, leading to increased plant growth compared to a single bacterium or the uninoculated control. | [81] |
| Cicer arietinum L. (chickpea) | Mesorhizobium ciceri EE-7 (salt-sensitive) and Mesorhizobium ciceri G-55 (salt-tolerant) | Two isolates of M. ciceri, one that was salt sensitive and another that was salt tolerant, were transformed with an isolated acdS gene encoding ACC deaminase. Salt stress reduced the biomass of plants inoculated with the wild-type strains. The salt-tolerant bacterial strain induced a higher nodulation rate in chickpeas compared to the salt-sensitive strain. The shoot dry weight was increased in plants inoculated with the salt-sensitive transformant strain. In plants inoculated with the salt-sensitive transformant strain, nodulation was found to be comparable to that induced by the salt-tolerant strain. | [82] |
| Coriandrum sativum L. (coriander) | Azospirillum brasiliense and Azotobacter chroococcum | Inoculation of coriander seeds, exposed to four levels of salt stress, with a mixture of A. brasiliense and A. chroococcum enhanced chlorophyll content and increased grain yield and plant biomass compared to uninoculated plants. Combined inoculation and salt stress increased catalase and decreased the level of ascorbate peroxidase and guaiacol peroxidase compared to untreated plants. Inoculation with both PGPB lowered Na and increased the K concentration in coriander leaves compared to untreated plants. The presence of PGPB improved plant growth in both the absence and presence of salt stress conditions. | [83] |
| Cucumis sativus (cucumber) | Pseudomonas fluorescens, Bacillus megaterium, and Variovorax paradoxus | The ability to solubilize phosphates and synthesize ACC deaminase, siderophores, and IAA was assessed in the three PGPB strains grown at two salt concentrations (2 and 5% NaCl w/v). While B. megaterium was the least affected by high salinity, ACC deaminase activity as well as siderophore and IAA production in P. fluorescens remained unaffected under salt stress. On the contrary, V. paradoxus was not tolerant to salt, and its expression of plant beneficial traits was reduced by salinity. When inoculated onto cucumber plants grown at three different salinity levels, P. fluorescens was the most effective of the three strains at decreasing the inhibitory effects of salinity. | [84] |
| Hordeum vulgare L. (barley), Trifolium repens L. (clover), and Pennisetum glaucum L.R. Br. (pearl millet) | Pseudomonas putida UW3 and UW4 | Barley, clover, and pearl millet plants grown in the presence of salt and inoculated with P. putida UW3 and UW4. P. putida UW4 increased barley biomass compared to uninoculated plants. Strain UW3 increased the biomass of the three crops. Shoot and root length and weight were increased in inoculated plants, suggesting a more efficient photosynthetic activity in the presence of the bacterial strains. Data from pulse amplitude modulation fluorometry showed that the reduction of plant photosynthetic activity induced by salt stress was recovered once the strains were applied. | [85] |
| Medicago sativa L. (alfalfa) | Bacillus megaterium NRCB001, Bacillus subtilis subsp. subtilis NRCB002, and Bacillus subtilis NRCB003 | Thirteen bacterial strains were isolated from the rice rhizosphere and characterized for their plant beneficial traits. B. megaterium NRCB001, B. subtilis subsp. subtilis NRCB002, and B. subtilis NRCB003 synthesized auxin, siderophores, NH3, and ACC deaminase and solubilized phosphate and potassium. Strains NRCB001 and NRCB002 tolerated 1750 mM NaCl. The three strains were inoculated onto M. sativa grown under normal conditions and salinity stress. Strains NRCB002 and NRCB003 increased the dry weight of alfalfa compared with non-inoculated seedlings treated with 130 mM NaCl. | [86] |
| Oryza sativa L. (rice) | Streptomyces sp. GMKU 336 and its ACC deaminase-deficient mutant | Plants of Thai jasmine rice cultivar Khao Dok Mali 105 grown under salt stress were inoculated with the endophyte Streptomyces sp. GMKU 336 or with its mutant lacking ACC deaminase activity. Strain GMKU 336 increased plant growth and chlorophyll, proline, K+, Ca+, and water content. The amount of released ethylene was reduced, as was the content of ROS and Na+, and the Na+/K+ ratio, compared to uninoculated plants or to those inoculated with the mutant. Plants treated with the wild type showed down-regulation of genes involved in the ethylene synthesis pathway, ACO1 and EREBP1, while acdS was up-regulated. Genes involved in osmotic balance, Na+ transport, calmodulin, and antioxidant enzymes were upregulated. | [87] |
| Oryza sativa (rice) | Bacillus tequilensis 10b (UPMRB9) | The effect of strains 10b UPMRB9′ on the growth of rice that was grown in the presence of salt was assessed. Strain 10b UPMRB9′ improved osmoprotectant properties such as proline, the soluble sugar concentration, and the levels of the antioxidant enzymes uperoxide dismutase, peroxidase, and catalase. Rice inoculated with strain UPMRB9 accumulated a greater amount of N and Ca in plant tissues, suggesting that this strain could behave as a bio-augmenter to improve biochemical and nutritional features in rice plants under salinity stress. | [88] |
| Panax ginseng (ginseng) | Paenibacillus yonginensis DCY84T | The impact of strain DCY84T, able to synthesize IAA and siderophore and solubilize phosphate, was assessed under short- and long-term salinity stress. Ginseng seedlings inoculated with the bacterial strain, following exposure to salt stress, were protected by the induction of plant defense-related systems (ion transport, ROS scavenging enzymes, proline content, total sugars, and ABA biosynthetic genes), as well as genes involved in root hair formation. The metabolome of the seedlings treated with DCY84T and exposed to salt stress overlapped with that of control plants. | [89] |
| Pisum sativum (pea) | Bacillus marisflavi (CHR JH 203) and Bacillus cereus (BST YS1_42) | Inoculation of pea plants with B. marisflavi CHR JH 203 and B. cereus BST YS1_42, both synthesizing a high amount of ACC deaminase, grown under salinity, improved plant biomass as well as the amount of plant carbohydrates, reducing sugars, proteins, chlorophylls, phenol, flavonoids, and antioxidant enzymes levels. In addition, plant ROS scavenging genes, defense genes, and cell rescue genes were all overexpressed in inoculated plants in the presence of 1% NaCl. | [90] |
| Seidlitzia rosmarinus Ehrenb. ex Boiss (perennial-green desert species of saltwort) | Rothia terrae, Kocuria palustris, Pseudomonas baetica, Pseudomonas fluorescens Staphylococcus warneri, Staphylococcus epidermidis, Staphylococcus succinus, Paenibacillus amylolyticus, Brevibacterium frigoritolerans, Stenotrophomonas pavanii, Halomonas sulfidaeris, Planococcus salinarum, Planomicrobium koreense, Planococcus halocryophilus, Planomicrobium soli | Culturable endophytic bacteria from the halophytic plant Seidlitzia rosmarinus Ehrenb. ex Boiss. were isolated and characterized to evaluate their plant beneficial traits under salt stress. Root endophytes belonged to genera Rothia, Kocuria, Pseudomonas, Staphylococcus, Paenibacillus, and Brevibacterium; shoot isolates belonged to Staphylococcus, Rothia, Stenotrophomonas, Brevibacterium, Halomonas, Planococcus, Planomicrobium, and Pseudomonas genera; Staphylococcus, Rothia, and Brevibacterium occurred in both roots and shoots. Synthesis of IAA and ACC deaminase was higher in bacteria from roots than from shoots. Finally, S. pavanii JST3 and P. fluorescens JST2 improved both shoot and root growth of Lepidium sativuum under salinity conditions. | [91] |
| Solanum lycopersicum (tomato) | Pseudomonas azotoformans CHB 1107 | Strain CHB 1107 wild-type (producing ACC deaminase) lowered ethylene and proline levels in tomato plants exposed to high salt levels, increasing the dry weights of shoots and roots compared with uninoculated plants. Plants that were inoculated with a mutant that lacked ACC deaminase activity showed reduced K, Ca, and Mn uptake compared with plants inoculated with the wild-type strain. The wild-type strain CHB 1107 reduced the uptake of Na by tomato plants compared with the mutant strain under salt stress. Tomato plants inoculated with the wild-type strain yielded a higher K/Na ratio than those that were inoculated with the mutant. | [92] |
| Sorghum vulgare (sorghum) | Pseudomonas migulae SVB3R2, SVB3R3, SVB3R4, Pseudomonas sp. SVB3R5, Pseudomonas brassicacearum SVB6R1 | Sorghum, tomato, and cucumber bacterial endophytes were characterized by 16S rRNA sequence determination and tested for plant beneficial traits. The activity of five endophytes was tested on plants grown with salinity stress. Strains SVB3R3 and SVB3R4 increased plant biomass, and strains SVB3R3 and SVB3R4 and SVB6R1 decreased the symptoms of plant salinity stress. Only strain SVB6R1 could produce ACC deaminase. | [93] |
| Triticum aestivum (wheat) | Bacillus pumilus SU3, Bacillus aquimaris SU8, Bacillus pumilus SU10, Bacillus arsinicus SU13, Arthrobacter sp. SU18, Bacillus cereus SU24, Pseudomonas mendocina SU40, Bacillus aquimaris SU44, and Bacillus subtilis SU47 | Salt-tolerant (ST) PGPB positively influenced the growth and yield of wheat in saline soil. All nine tested strains improved plant growth in saline soil under greenhouse conditions, with strain DU18 being the most efficient. Under field conditions, strains SU44 and SU8 were the best in increasing plant biomass. Plant inoculation with strain SU8 led to higher proline content and total soluble sugar accumulation in wheat, while strain SU44 resulted in a higher accumulation of reducing sugars. The amounts of N, K, and P in wheat leaves increased significantly after inoculation with all the strains; B. subtilis SU47 lowered the sodium (Na) content in wheat leaves. | [94] |
| Triticum aestivum (wheat) | Serratia marcescens CDP-13 | Serratia marcescens CDP-13 is halotolerant, produces ACC deaminase, solubilizes phosphate, synthesizes siderophores and IAA, and fixes N2. Wheat inoculation with strain CDP-13 increased plant biomass under salinity stress, reducing inhibition of plant growth caused by salt and lowering the amount of osmoprotectants (such as proline, malondialdehyde, soluble sugar), protein, and IAA content in plants. | [95] |
| Triticum aestivum L. (wheat). | Bacillus megaterium PN89 | B. megaterium PN89, able to synthesize IAA, induced increased germination rate and root and shoot length in wheat plants exposed to salt stress, compared to non-inoculated controls. | [96] |
| Triticum aestivum (wheat) | Brevibacterium frigoritolerans, Bacillus velezensis, and Bacillus thuringiensis | The expression of plant-beneficial traits of Br. frigoritolerans alone or in combination with B. velezensis and B. thuringiensis under six salinity levels was characterized, and the effects on wheat of these strains alone or in combination under salt stress were assessed. B. frigoritolerans was the most effective, both for physiological trait expression and wheat plant growth promotion. Under salinity stress, the mixed inoculation of the three bacterial strains was more efficient than any single inoculation. | [97] |
| Triticum durum (wheat) | Fourteen strains of the genera Streptomyces and Nocardiopsis | Fourteen Actinomycetes strains were tested for expression of plant-beneficial activities under salinity conditions. The isolates could solubilize inorganic phosphate and synthesize IAA, HCN, and ammonia when grown in the presence of different salt concentrations. The majority of the strains produced ACC deaminase. Plant inoculation with these strains improved biomass and yielded an increased amount of chlorophyll and proline compared to uninoculated plants, both with and without salt. | [98] |
| Vigna radiata L. (mung bean) | Enterobacter cloacae KBPD | Strain KBPD is an ACC deaminase producer, able to solubilize phosphates and synthesize IAA, siderophore, ammonia, hydrogen cyanide, and exopolysaccharide. V. radiata plants exposed to salinity and inoculated with this bacterial strain showed increased shoot length, root length, and fresh and dry weights. Inoculation with strain KBPD also reduced proline content in plants grown with salt stress. | [99] |
| Vigna radiata (L.) R. Wilczekspring (mung bean) | Rhizobium sp. LSMR-32 and Enterococcus mundtii LSMRS-3 | Under salt stress conditions, separate inoculation with the two bacterial strains induced increased seed germination, grain yield, plant height, biomass, chlorophyll content, and nutrient uptake compared to uninoculated plants. Inoculation with both strains increased both symbiotic parameters (nodulation rate, nodule biomass, and leghaemoglobin amount) and soil phosphatase and dehydrogenase levels. The microbial consortium enhanced the level of proline and anti-oxidative enzymes. | [100] |
| Zea mays (maize) | Azospirillum lipoferum or Azotobacter chroococcum | Maize (corn) plants that were exposed to salt stress had reduced growth parameters, pigments, soluble proteins, K+, and a K+/Na+ ratio. Salinity led to increased levels of soluble sugars, proline, Na+, malondialdehyde, and peroxidase and catalase activity, while the activity of plant ascorbate peroxidase remained unaffected. Plants inoculated with A. lipoferum or A. chroococcum increased growth parameters, pigments, K+, osmolytes, K+/Na+ ratio, and the antioxidative enzymes in salt-affected maize. Both bacterial strains also lowered malondialdehyde and Na+ in maize plants. | [101] |
The majority of the papers reported in Table 1 and Table 2 are focused on experiments performed under controlled laboratory or greenhouse conditions. Unfortunately, despite their effectiveness under these conditions, the field application of PGPB covers only a small portion of agriculture at a global level [102]. Although PGPB are more acceptable for deliberate use in the field than transgenic plants in many countries, their exploitation is well below their potentiality, and this is mainly related to the lack of consistency and repeatability of PGPB performance in open field conditions. The survival of PGPB once inoculated onto the plants is one of the main problems. This is related to the compatibility of the selected PGPB with the considered plant species and the soil characteristics. Once introduced in the environment, the PGPB establishes relationships with the resident microbiota, and the results of this interaction may range from positive to negative [103]. Finally, the inoculant formulation has a specific role in maintaining the strain survival, especially after inoculation in open field conditions [104] Regarding stressed soil such as those affected by an excess of salt or water deficiency, it is necessary to consider the adaptability of the selected PGPB to these harsh conditions. When considering a stressed environment, it is always important to keep in mind that the expression of plant-beneficial bacterial physiological traits can be modified by the stress itself [105,106]. Following the analysis of the literature for this manuscript, it is clear that there is a general lack of data on the performance of PGPB in increasing plant growth and yield under salinity and drought stress conditions under open field conditions.
3. Groups of Microorganisms to Lower Plant Drought and Salt Stress
Since bacteria can express different plant-beneficial activities, inoculation of plants with a combination of two or more non-antagonistic microorganisms, belonging to the same or different species, or including both prokaryotic and eukaryotic organisms, can often lead to additive or synergistic effects in the promotion of plant growth. In fact, the lack of one plant beneficial activity in one strain can sometimes be overcome if another microorganism expresses this specific physiological trait [29,107,108,109]. Moreover, the combination of diverse plant growth-promoting microorganisms (PGPM) offers a wide range of positive activity to a plant, including the enhancement of plant growth, reduced susceptibility to soil-borne diseases, increased yield, and improved seed and fruit nutritional value. The commercialization of groups of PGPM as biofertilizers and biocontrol agents represents the bottleneck of the full process [110,111]. However, as demonstrated by the large number of scientific papers recently published on this topic, the use of microbial consortia is becoming a reliable tool to support plant growth in stressed environments, especially those characterized by an excess of salt [112,113,114,115,116] or a deficiency of water [63,117,118,119,120,121].
3.1. Bacterial Consortia
Bacterial consortia may be classified as either simple or complex. In simple consortia, all of the bacterial strains are inoculated in a bioreactor at the same time, while in a complex consortium, the bacterial growth occurs in separate bioreactors [29,122]. Once the strategy of employing a simple consortium has been decided upon, it should be taken into account that the metabolism of each inoculated bacterial strain affects the growth and physiology of the other co-existing strains, leading to a plethora of possible interactions, ranging from neutral, to positive (cooperation, synergism, commensalism), to negative (amensalism, competition, antagonism). Although the occurrence of one or the other type of interactions is usually finely monitored and regulated during the scale-up of the fermentation process, the issue of the strain’s compatibility plays a pivotal role in the final result [123]. For, example, in Pseudomonas koreensis S2CB45 (which was isolated from the surface sterilized spore of a mycorrhizal fungi) and Microbacterium hydrothermale IC37-36 (which was isolated from a rice spermosphere), the expression of plant-beneficial traits such as ACC deaminase, IAA, and cytokinin was higher when these PGPB were co-cultured rather than grown separately. Consequently, when this consortium was used to inoculate red pepper plants grown at two salt stress levels, these bacterial strains reduced ethylene emission by 20% compared to uninoculated plants, lowered ROS accumulation, and increased activity of antioxidant enzymes [112].
Especially for those consortia intended for use as plant growth promoters in saline or arid environments, the sampling site from which the bacterial strains are isolated is key. Since autochtonous plants growing in arid environments have co-evolved with their associated microbiota, it is a common opinion that selecting bacterial strains from these specific environments may help to obtain well-adapted bacterial strains with high tolerance to stress as well as long survival and persistence in open field conditions [124,125].
In a recent study, Mansour et al. [118] examined the effects of either the single or double inoculation of five faba bean cultivars with strains R. leguminosarum bv. viciae USDA 2435 and P. putida RA MTCC5279 on the growth and health of these plants treated to (i) optimal watering, (ii) moderate watering, or (iii) severe drought conditions. While the water deficiency reduced the synthesis of photosynthetic pigments and enhanced the production of antioxidant enzymes and osmoprotectants to a different extent according to the drought sensitivity of the cultivar, plant inoculation with the bacterial strains led to increased seed yield and crop water productivity.
Similarly, among 89 strains originally from the rhizosphere or endosphere of two Algerian autochthonous halophytic plants (Suaeda mollis and Salsola tetrandra), three bacterial strains, i.e., Bacillus atropheus (BR5, OR15, and RB13), were selected based on plant growth-promoting traits (IAA, ACC deaminase, nitrogen fixation, and phosphate solubilization), antifungal activity, and tolerance to environmental conditions (pH, PEG, and NaCl). The bacterial strains were used to inoculate A. thaliana and durum wheat alone or in combination under salt stress conditions. Overall, the data obtained indicate that bacterial inoculation of both plant species increased plant growth under normal conditions. When wheat plants were cultivated in soils characterized by three salinity levels, inoculation with the consortium was the most efficient strategy for increasing growth compared to control plants. Finally, inoculation with the consortium increased chlorophyll and carotenoid contents while lowering proline content, lipid peroxidation, and the activities of antioxidant enzymes in treated plants, suggesting that the bacterial treatments limit the damages induced by salt stress [116].
3.2. Bacteria Plus Fungi (Including AMF)
Consistent with the fact that plant microbiota includes a wide variety of different microorganisms, the effects of consortia involving bacterial strains associated with arbuscular mycorrhizal fungi (AMF) and other non-obligate symbiotic fungi on plant growth under arid conditions is a powerful tool, where each microorganism potentiates the bioprotection against the effects of various abiotic stresses on the plant [29]. AMF are common plant-beneficial fungi that typically form a symbiotic relationship with the roots of ~80–90% of all land plants, colonizing plant roots intracellularly. AMF enter the root cortical cell walls, and once inside these cells, form obligate branched intracellular fungal structures (arbuscules), where an exchange of nutrients between the plant and the beneficial fungus takes place. In this exchange, the plant provides the fungus with both fixed carbon and nitrogen, and in exchange, the fungus provides the plant with an effectively increased root system and a significantly increased ability to take up water and nutrients from the soil.
The work of several researchers has highlighted the occurrence of additive and synergistic effects between AMF and PGPB [29], with PGPB behaving as mycorrhizal helper bacteria and thus favoring the establishment of the mycorrhizal symbiosis. Even more complex is the interrelationship involving legumes, rhizobia, and AMF, where the AMF can favor nodule formation, while rhizobia may reduce mycorrhizal hyphal development [126]. On this latest point, contrasting results have been obtained by Igiehon and Babalola [121], who assessed the effects of a combination of rhizobia species and AMF on the growth of soybean under drought stress. Based on their results, co-inoculation of soybean plants with the bacteria Rhizobium sp. R1 and R. cellulosilyticum R3, together with a consortium of AMF (consisting of Paraglomus occulum, Gigaspora gigantea, Funneliformis mosseae, Claroideoglomus etunicatum and Rhizophagus clarus), improved the growth and yield of soybean plants that were exposed to 40% water stress, leading to enhanced leaf relative water content and proline concentration. Moreover, mycorrhizal plants that were inoculated with rhizobia displayed the greatest number of fungal spores and mycorrhizal colonization intensity of all of the water regimes, revealing that the rhizobia and AMF consortium can positively interact and represent a valuable tool in sustainable agriculture.
A large number of factors can affect the tripartite relationship between the host plant, mycorrhizal fungi, and PGPB strains, ranging from the host plant and AMF species involved to the chemical and physical parameters of the growth substrate(s) [127,128]. For example, Veselaj et al. [129] reported that in legumes the relationships occurring among AMF, PGPB, and rhizobia are dependent both on the species of microorganisms involved and the level of salinity. In this study, inoculated pea plants were grown at two different levels of salt stress, with two AMF (Rhizophagus irregularis and C. claroideum) and/or two PGPB, one of them being a rhizobial strain (Rhizobium leguminosarum) and Burkholderia sp. When grown under optimal conditions, the plant growth parameters were improved by inoculation with R. irregularis and bacteria (R. leguminosarum and Burkholderia sp.) or by the combination of the AMF (R. irregularis + C. claroideum) with each type of bacteria. While the plant growth was reduced by the salt stress, inoculation with R. irregularis led to better vegetative development and higher productivity than with C. claroideum. Moreover, under salt stress, pea plants treated with Burkolderia sp. increased vegetative growth more efficiently than R. leguminosarum. In this experiment, the best results, in terms of plant growth and productivity, were observed with plants inoculated with both AMF the rhizobia strain.
In addition to AMF, mixed bacterial/fungal consortia can include other microfungi, such as isolates of Trichoderma species and the root endophyte Serendipita indica (previously classified as Piriformospora indica). While Trichoderma spp., and especially Trichoderma harzianum, is mainly known for its mycoparasistic behaviour mediated by chitin degradation [130], S. indica is a root endophyte mimicking AMF, able to grow in pure culture and improve nutrient uptake, increasing disease resistance and stress tolerance to environmental conditions [131]. In a recent study [132], lettuce and tomato plants were cultivated in the presence of different salinity levels and inoculated with two commercial biostimulants, one containing Bacillus amyloliquefaciens, B. brevis, B. circulans, B. coagulans, B. firmus, B. halodenitrificans, B. laterosporus, B. licheniformis, B. megaterium, B. mycoides, B. pasteurii, B. subtilis, and Paenibacillus polymyxa, and the other Glomus spp., Agrobacterium radiobacter, Bacillus subtilis, Streptomyces spp., and Thricoderma spp. While uninoculated plants showed symptoms related to salinity, plants inoculated with the two formulations showed increased biomass, leaf number, and leaf area and were less sensitive to salinity stress. Comparing the efficacy of the two commercial formulations, the inoculum with the consortium containing AMF was more effective than the one based exclusively on Bacillus species.
Notwithstanding the good results obtained inoculating plants with S. indica, both under optimal and drought/salt stressed conditions [133,134,135,136,137], combinations of S. indica and PGPB have not always yielded positive results. Thus, the possibility of using this fungus in consortia with PGPB has been shown to be effective only in studies aimed at increasing plant disease resistance [138].
4. Summary and Conclusions
Approximately 20 years ago, scientists began observing and reporting that PGPB could help plants to overcome some of the effects of drought and salt stress. Since that time, there have been a large number of literature reports confirming the ability of PGPB, applied by itself or as part of a consortium, to ameliorate some of the negative effects of the related abiotic stresses of drought and salt stress on plant growth and development. On the one hand, PGPB can directly promote plant growth, thereby helping plants to defend themselves against growth-inhibiting drought and salt stress. On the other hand, PGPB can augment a plant’s defenses against the often deleterious effects of drought and/or salt stress using a number of different mechanisms. These mechanisms include (but are not limited to) the synthesis of the enzyme ACC deaminase, which can decrease plant stress ethylene levels, the synthesis of IAA and cytokinin, which can modulate a plant’s response to stress and promote its growth, and the synthesis of various osmoprotectant molecules, such as proline and trehalose, which can structure plant intracellular water, thereby protecting the cell’s structural integrity.
To date, the majority of the reported experiments using PGPB to ameliorate drought and salt stress include just a single well-characterized PGPB strain. However, scientists have demonstrated that the addition of several bacterial strains or a mixture of bacterial and fungal strains may also be highly effective. However, additional work is required to determine whether some of these consortia will be sufficiently stable and therefore effective throughout a growing season and in the long term. In addition, just how effective these approaches will be in the field (as opposed to in growth chamber and greenhouse experiments) in the presence of severe drought or salt stress remains to be determined. Certainly, the increased use of naturally occurring PGPB in agriculture as a means of addressing various abiotic stresses is an attractive approach that (based on the results that have been published) shows quite a lot of promise. Nevertheless, there are still many scientific and regulatory hurdles to overcome before this technology is widely accepted and used on a large scale.
Author Contributions
E.G. and B.R.G. jointly conceived of, wrote, and edited this article. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Glick, B.R. Beneficial Plant-Bacterial Interactions; Springer: Berlin/Heidelberg, Germany, 2020; 383p. [Google Scholar]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.S.P. Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: Oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Gepstein, S.; Sabehi, G.; Carp, M.-J.; Hajouj, T.; Nesher, M.F.O.; Yariv, I.; Dor, C.; Bassani, M. Large-scale identification of leaf senescence-associated genes. Plant J. 2003, 36, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–28. [Google Scholar] [CrossRef]
- Umezawa, T.; Fujita, M.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Engineering drought tolerance in plants: Discovering and tailoring genes to unlock the future. Curr. Opin. Biotechnol. 2006, 17, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial mediation of plant hormone status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gepstein, S.; Glick, B.R. Strategies to ameliorate abiotic stress-induced plant senescence. Plant Mol. Biol. 2013, 82, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda, M.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.; Cho, Y.-C. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Christakis, C.A.; Daskalogiannis, G.; Chatzaki, A.; Markakis, E.A.; Mermigka, G.; Sagia, A.; Rizzo, G.F.; Catara, V.; Lagkouvardos, I.; Studholme, D.J.; et al. Endophytic bacterial isolates from haplophytes demonstrate phytopathogen biocontrol and plant growth promotion under high salinity. Front. Microbiol. 2021, 12, 681567. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Jabborova, D.; Hashem, A. Pseudomonas induces salinity tolerance in cotton (Gossypium hirsutum) and resistance to Fusarium root rot through the modulation of indole-3-acetic acid. Saudi J. Biol. Sci. 2015, 22, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Mavrodi, O.V.; Parejko, J.A.; Bonsall, R.F.; Kwak, Y.-S.; Paulitz, T.C.; Thomashow, L.S.; Weller, D.M. Accumulation of the antibiotic phenazine-1-carboxylic acid in the rhizosphere of dryland cereals. Appl. Environ. Microbiol. 2012, 78, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Paalme, V.; Pavlicek, T.; Bergquist, J.; Vangala, A.; Danilas, T.; Nevo, E. Bacterial distribution in the rhizosphere of wild barley under contrasting microclimates. PLoS ONE 2011, 6, e17968. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Glick, B.R. Mechanisms used by plant growth-promoting bacteria. In Bacteria in Agrobiology: Plant Nutrient Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 17–46. [Google Scholar]
- Gamalero, E.; Glick, B.R. Plant growth-promoting bacteria in agriculture and stressed environments. In Modern Soil Microbiology, 3rd ed.; van Elsas, J.D., Trevors, J.T., Soares Rosado, A., Nannipieri, P., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 361–380. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1-carboxylate (ACC) deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Berta, G.; Glick, B.R. The use of microorganisms to facilitate the growth of plants in saline soils. In Microbial Strategies for Crop Improvement; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–22. [Google Scholar]
- Kim, Y.-C.; Glick, B.R.; Bashan, Y.; Ryu, C.-M. Enhancement of plant drought tolerance by microbes. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 383–413. [Google Scholar]
- Sergeeva, E.; Shah, S.; Glick, B.R. Tolerance of transgenic canola expressing a bacterial ACC deaminase gene to high concentrations of salt. World J. Microbiol. Biotechnol. 2006, 22, 277–282. [Google Scholar] [CrossRef]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-bacterial amelioration of plant abiotic and biotic stress. Front. Sustain. Food Syst. 2021, 5, 672881. [Google Scholar] [CrossRef]
- Reed, M.L.E.; Glick, B.R. Applications of plant growth-promoting bacteria for plant and soil systems. In Applications of Microbial Engineering; Gupta, V.K., Schmoll, M., Maki, M., Tuohy, M., Mazutti, M.A., Eds.; Taylor and Francis: Enfield, CT, USA, 2013; pp. 181–229. [Google Scholar]
- Pattyn, J.; Vaughan-Hirsh, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex control circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-containing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of damages caused by high salinity stress by plant growth-promoting bacterial endophytes. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.C.; Duan, J.; DiBernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The production of ACC deaminase and trehalose by the plant growth promoting bacterium Pseudomonas sp. UW4 synergistically protect tomato plants against salt stress. Front. Microbiol. 2019, 10, 1392. [Google Scholar] [CrossRef] [Green Version]
- Tsukanova, K.A.; Chebotar, V.K.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Duca, D.R.; Glick, B.R. Indole-3-acetic acid biosynthesis and its regulation in plant-associated bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of Plant Growth-Promoting Rhizobacteria (PGPR) in stimulating salinity stress defense in plants: A review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef]
- Xu, J.; Li, X.-L.; Luo, L. Effects of engineered Sinorhizobium meliloti on cytokinin synthesis and tolerance of alfalfa to extreme drought stress. Appl. Environ. Microbiol. 2012, 78, 8056–8061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Jiang, W.; Cheng, Z.; Heikkila, J.J.; Glick, B.R. The complete genome sequence of the plant growth-promoting bacterium Pseudomonas putida UW4. PLoS ONE 2013, 8, e58640. [Google Scholar]
- Chaplin, M. Do we underestimate the importance of water in cell biology? Nat. Rev. Mol. Cell. Biol. 2006, 7, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Julca, I.; Alaminos, M.; Gonzalez-Lopez, J.; Manzanera, M. Xeroprotectants for the stabilization of biomaterials. Biotechnol. Adv. 2012, 30, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose metabolism: From osmoprotection to signaling. Int. J. Mol. Sci. 2009, 10, 3793–3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, M.M.S.; Abdelgawad, Z.A.; El-Bassiouny, H.M.S. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties. S. Afr. J. Bot. 2016, 103, 275–282. [Google Scholar] [CrossRef]
- Lau, J.A.; Lennon, J.T. Microbe-mediated adaptation to novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vílchez, J.I.; García-Fontana, C.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant drought tolerance enhancement by trehalose production of desiccation-tolerant microorganisms. Front. Microbiol. 2016, 7, 1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.P.; Grover, M.; Chourasiya, D.; Bharti, A.; Agnihotri, R.; Maheshwari, H.S.; Pareek, A.; Buyer, J.S.; Sharma, S.K.; Schütz, L.; et al. Deciphering the role of trehalose in tripartite symbiosis among rhizobia, arbuscular mycorrhizal fungi, and legumes for enhancing abiotic stress tolerance in crop plants. Front. Microbiol. 2020, 17, 509919. [Google Scholar] [CrossRef] [PubMed]
- Onwe, R.O.; Onwosi, C.O.; Ezugworie, F.N.; Ekwealor, C.C.; Okonkwo, C.C. Microbial trehalose boosts the ecological fitness of biocontrol agents, the viability of probiotics during long-term storage and plants tolerance to environmental-driven abiotic stress. Sci. Total Environ. 2022, 1, 150432. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, S.; Song, L.; Yan, H.; Liu, K.; Meng, W.; Wang, T. Trehalose production using recombinant trehalose synthase in Bacillus subtilis by integrating fermentation and biocatalysis. J. Agric. Food Chem. 2019, 67, 9314–9324. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Patel, M.K.; Kumar, N.; Bajpai, A.B.; Siddique, K.H.M. Metabolomics and molecular approaches reveal drought stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 9108. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Sen, S.; Mohapatra, S. Modulation of proline metabolic gene expression in Arabidopsis thaliana under water-stressed conditions by a drought-mitigating Pseudomonas putida strain. Ann. Microbiol. 2017, 67, 655–668. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, F.; Mickan, B.S.; Wang, D.; Wang, W. Physiological, proteomic, and metabolomic analysis provide insights into Bacillus sp.-mediated salt tolerance in wheat. Plant Cell Rep. 2021, 41, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Woo, O.-G.; Bae, Y.; Keum, H.L.; Chung, S.; Sul, W.J.; Lee, J.-H. Enhanced drought and salt stress tolerance in Arabidopsis by Flavobacterium crocinum HYN0056T. J. Plant Biol. 2020, 63, 63–71. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, T.M.; Choi, B.; Kim, Y.; Kim, E. Invasive Lactuca serriola seeds contain endophytic bacteria that contribute to drought tolerance. Sci. Rep. 2021, 11, 13307. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Singh, R.P.; Singh, A.L.; Singh, M. Identification of genes involved in phosphate solubilization and drought stress tolerance in chickpea symbiont Mesorhizobium ciceri Ca181. Arch. Microbiol. 2021, 203, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Chaín, J.M.; Tubert, E.; Graciano, C.; Castagno, L.N.; Recchi, M.; Pieckenstain, F.L.; Estrella, M.J.; Gudesblat, G.; Amodeo, G.; Baroli, I. Growth promotion and protection from drought in Eucalyptus grandis seedlings inoculated with beneficial bacteria embedded in a superabsorbent polymer. Sci. Rep. 2020, 26, 18221. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.). Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Behrooz, A.; Vahdati, K.; Rejali, F.; Lotfi, M.; Sarikhani, S.; Leslie, C. Arbuscular mycorrhiza and plant growth-promoting bacteria alleviate drought stress in walnut. Hort. Sci. 2019, 54, 1087–1092. [Google Scholar] [CrossRef] [Green Version]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria that confer resistance in tomato to salt stress. Plant Physiol. Biochem. 2004, 42, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Astorga-Eló, M.; Gonzalez, S.; Acuña, J.J.; Sadowski, M.J.; Jorquera, M.A. Rhizobacteria from ‘flowering desert’ events contribute to the mitigation of water scarcity stress during tomato seedling germination and growth. Sci. Rep. 2021, 11, 13745. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Davies, W.J.; Tikhonovich, I.A. Rhizobacteria that produce auxins and contain ACC deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Furlan, F.; Saatkamp, K.; Gazolla Volpiano, C.; de Assis Franco, F.; Fonseca dos Santos, M.; Gruszka Vendruscolo, E.C.; Guimarães, V.F.; Torres da Costa, A.C. Plant growth-promoting bacteria effect in withstanding drought in wheat cultivars. Sci. Agrar. 2017, 18, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Srivastava, R.; Gupta, V.V.S.R.; Franco, C.M.M.; Sharma, A.K. Evaluation of ACC-deaminase-producing rhizobacteria to alleviate water-stress impacts in wheat (Triticum aestivum L.) plants. Can. J. Microbiol. 2019, 65, 387–403. [Google Scholar] [CrossRef]
- Kasim, W.A.; Osman, M.E.H.; Omar, M.N.; Salama, S. Enhancement of drought tolerance in Triticum aestivum L. seedlings using Azospirillum brasilense NO40 and Stenotrophomonas maltophilia B11. Bull. Natl. Res. Cent. 2021, 45, 95. [Google Scholar] [CrossRef]
- Hone, H.; Mann, R.; Yang, G.; Kaur, J.; Tannenbaum, I.; Li, T.; Spangenberg, G.; Sawbridge, T. Profiling, isolation and characterisation of beneficial microbes from the seed microbiomes of drought tolerant wheat. Sci. Rep. 2021, 7, 11916. [Google Scholar] [CrossRef] [PubMed]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci. Rep. 2018, 23, 3560. [Google Scholar] [CrossRef] [PubMed]
- Inthama, P.; Pumas, P.; Pekkoh, J.; Pathom-Aree, W.; Pumas, C. Plant growth and drought tolerance-promoting bacterium for bioremediation of paraquat pesticide residues in agriculture soils. Front. Microbiol. 2021, 12, 604662. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Tufail, M.A.; Asghar, H.N.; Nazli, F.; Zahir, Z.A. Appraising the potential of EPS-producing rhizobacteria with ACC-deaminase activity to improve growth and physiology of maize under drought stress. Physiol. Plant. 2021, 172, 463–476. [Google Scholar] [CrossRef]
- Nephali, L.; Moodley, V.; Piater, L.; Steenkamp, P.; Buthelezi, N.; Dubery, I.; Burgess, K.; Huyser, J.; Tugizimana, F. A metabolomic landscape of maize plants treated with a microbial biostimulant under well-watered and drought conditions. Front. Plant Sci. 2021, 12, 676632. [Google Scholar] [CrossRef] [PubMed]
- Ledger, T.; Rojas, S.; Timmermann, T.; Pinedo, I.; Poupin, M.J.; Garrido, T.; Richter, P.; Tamayo, J.; Donoso, R. Volatile-mediated effects predominate in Paraburkholderia phytofirmans growth promotion and salt stress tolerance of Arabidopsis thaliana. Front. Microbiol. 2016, 7, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut. Front. Microbiol. 2016, 7, 1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddikee, M.A.; Sundaram, S.; Chandrasekaran, M.; Kim, K.; Selvakumar, G.; Sa, T. Halotolerant bacteria with ACC deaminase activity alleviate salt stress effect in canola seed germination. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 237–241. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Glick, B.R.; Zhou, R.; Hegedus, D.D. Inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acdS gene in transgenic plants increases salinity tolerance in Camelina sativa. Front. Microbiol. 2016, 7, 1966. [Google Scholar] [CrossRef]
- Heydarian, Z.; Gruber, M.; Glick, B.R.; Hegedus, D.D. Gene expression patterns in roots of Camelina sativa with enhanced salinity tolerance arising from inoculation of soil with Plant Growth Promoting Bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression the corresponding acdS gene. Front. Microbiol. 2018, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Heydarian, Z.; Gruber, M.; Coutu, C.; Glick, B.R.; Hegedus, D.D. Gene expression patterns in shoots of Camelina sativa with enhanced salinity tolerance provided by plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acdS gene. Sci. Rep. 2021, 11, 4260. [Google Scholar] [CrossRef] [PubMed]
- Siddikee, M.A.; Glick, B.R.; Chauhan, P.S.; Yim, W.J.; Sa, T. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1-carboxylic acid deaminase activity. Plant Physiol. Biochem. 2011, 49, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Samaddar, S.; Anandham, R.; Kang, Y.; Kim, K.; Selvakumar, G.; Sa, T. Beneficial soil bacterium Pseudomonas frederiksbergensis OS261 augments salt tolerance and promotes red pepper plant growth. Front. Plant Sci. 2017, 8, 705. [Google Scholar] [CrossRef] [PubMed]
- Samaddar, S.; Chatterjee, P.; Choudhury, A.R.; Ahmed, S.; Sa, T. Interactions between Pseudomonas spp. and their role in improving the red pepper plant growth under salinity stress. Microbiol. Res. 2019, 219, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Brígido, C.; Nascimento, F.X.; Duan, J.; Glick, B.R.; Oliveira, S. Expression of an exogenous 1-aminocyclopropane-1-carboxylate deaminase gene in Mesorhizobium spp. reduces the negative effects of salt stress in chickpea. FEMS Microbiol. Lett. 2013, 349, 46–53. [Google Scholar] [PubMed] [Green Version]
- Rabiei, Z.; Hosseini, S.J.; Pirdashti, H.; Hazrati, S. Physiological and biochemical traits in coriander affected by plant growth-promoting rhizobacteria under salt stress. Heliyon 2020, 6, e05321. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, S.M.; Ahmad, M.; Naveed, M.; Imran, M.; Zahir, Z.A.; Crowley, D.E. Relationship between in vitro characterization and comparative efficacy of plant growth-promoting rhizobacteria for improving cucumber salt tolerance. Arch. Microbiol. 2016, 198, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Alkowni, R.; Jodeh, S.; Hamed, R.; Samhan, S.; Daraghmeh, H. The impact of Pseudomonas putida UW3 and UW4 strains on photosynthetic activities of selected field crops under saline conditions. Int. J. Phytoremediat. 2019, 21, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, H.; Leng, J.; Niu, H.; Chen, X.; Liu, D.; Chen, Y.; Gao, N.; Ying, H. Isolation and characterization of plant growth-promoting rhizobacteria and their effects on the growth of Medicago sativa L. under salinity conditions. Antonie Van Leeuwenhoek 2020, 113, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Jaemsaeng, R.; Jantasuriyarat, C.; Thamchaipenet, A. Molecular interaction of 1-aminocyclopropane-1-carboxylate deaminase (ACCD)-producing endophytic Streptomyces sp. GMKU 336 towards salt-stress resistance of Oryza sativa L. cv. KDML105. Sci. Rep. 2018, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Shultana, R.; Kee Zuan, A.T.; Yusop, M.R.; Saud, H.M.; El-Shehawi, A.M. Bacillus tequilensis strain ‘UPMRB9′ improves biochemical attributes and nutrient accumulation in different rice varieties under salinity stress. PLoS ONE 2021, 13, e0260869. [Google Scholar] [CrossRef] [PubMed]
- Sukweenadhi, J.; Balusamy, S.R.; Kim, Y.J.; Lee, C.H.; Kim, Y.J.; Koh, S.C.; Yang, D.C. A growth-promoting bacteria, Paenibacillus yonginensis DCY84T enhanced salt stress tolerance by activating defense-related systems in Panax ginseng. Front. Plant Sci. 2018, 9, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Bano, A.; Rai, S.; Kumar, M.; Ali, J.; Sharma, S.; Pathak, N. ACC deaminase producing plant growth promoting rhizobacteria enhance salinity stress tolerance in Pisum sativum. 3 Biotech 2021, 11, 514. [Google Scholar] [CrossRef] [PubMed]
- Shurigin, V.; Egamberdieva, D.; Li, L.; Davranov, K.; Panosyan, H.; Birkeland, N.K.; Wirth, S.; Bellingrath-Kimura, S.D. Endophytic bacteria associated with halophyte Seidlitzia rosmarinus Ehrenb. ex Boiss. from saline soil of Uzbekistan and their plant beneficial traits. J. Arid Land 2020, 12, 730–740. [Google Scholar] [CrossRef]
- Liu, C.H.; Siew, W.; Hung, Y.T.; Jiang, Y.T.; Huang, C.H. 1-Aminocyclopropane-1-carboxylate (ACC) deaminase gene in Pseudomonas azotoformans is associated with the amelioration of salinity stress in tomato. J. Agric. Food Chem. 2021, 69, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Favale, N.; Bona, E.; Novello, G.; Cesaro, P.; Massa, N.; Glick, B.R.; Orozco-Mosqueda, M.d.C.; Berta, G.; Lingua, G. Screening of bacterial endophytes able to promote plant growth and increase salinity tolerance. Appl. Sci. 2020, 10, 5767. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P. Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 2015, 17, 288–293. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The Multifarious PGPR Serratia marcescens CDP-13 Augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.G.; Lee, J.M.; Choi, C.G.; Lee, H.; Moon, J.C.; Chung, N. Effect of plant growth-promoting rhizobacterial treatment on growth and physiological characteristics of Triticum aestivum L. under salt stress. Appl. Biol. Chem. 2021, 64, 89. [Google Scholar] [CrossRef]
- Wang, R.; Wang, C.; Feng, Q.; Liou, R.M.; Lin, Y.F. Biological inoculant of salt-tolerant bacteria for plant growth stimulation under different saline soil conditions. J. Microbiol. Biotechnol. 2021, 28, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Djebaili, R.; Pellegrini, M.; Rossi, M.; Forni, C.; Smati, M.; Del Gallo, M.; Kitouni, M. Characterization of Plant Growth-Promoting traits and inoculation effects on Triticum durum of Actinomycetes isolates under salt stress conditions. Soil Syst. 2021, 5, 26. [Google Scholar] [CrossRef]
- Bhise, K.K.; Bhagwat, P.K.; Dandge, P.B. Plant growth-promoting characteristics of salt tolerant Enterobacter cloacae strain KBPD and its efficacy in amelioration of salt stress in Vigna radiata L. J. Plant Growth Regul. 2017, 36, 215–226. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Sharma, P.; Nagpal, S.; Gupta, R.K.; Sirari, A.; Nair, R.M.; Bindumadhava, H.; Singh, S. Dual microbial inoculation, a game changer—Bacterial biostimulants with multifunctional growth promoting traits to mitigate salinity stress in spring mungbean. Front. Microbiol. 2021, 11, 600576. [Google Scholar] [CrossRef] [PubMed]
- Latef, A.A.H.; Abu Alhmad, M.F.; Kordrostami, M.; Abo-Baker, A.B.A.E.; Zakir, A. Inoculation with Azospirillum lipoferum or Azotobacter chroococcum reinforces maize growth by improving physiological activities under saline conditions. J. Plant Growth Regul. 2020, 39, 1293–1306. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Viveros, M.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Vassilev, N.; Vassileva, M.; Martos, V.; Garcia Del Moral, L.F.; Kowalska, J.; Tylkowski, B.; Malusá, E. Formulation of microbial inoculants by encapsulation in natural polysaccharides: Focus on beneficial properties of carrier additives and derivatives. Front. Plant Sci. 2020, 1, 270. [Google Scholar] [CrossRef] [PubMed]
- Rubiano-Labrador, C.; Bland, C.; Miotello, G.; Armengaud, J.; Baena, S. Salt stress induced changes in the exoproteome of the halotolerant bacterium Tistlia consotensis deciphered by proteogenomics. PLoS ONE 2015, 10, e0135065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogati, K.; Walczak, M. The impact of drought stress on soil microbial community, enzyme activities and plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.N.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 21, 205. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 23, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mushtaq, Z. PGPR: Present role, mechanism of action and future prospects along bottlenecks in commercialization. EQA-Int. J. Environ. Qual. 2021, 41, 9–15. [Google Scholar]
- Choudhury, A.R.; Choi, J.; Walitang, D.I.; Trivedi, P.; Lee, Y.; Sa, T. ACC deaminase and indole acetic acid producing endophytic bacterial co-inoculation improves physiological traits of red pepper (Capsicum annum L.) under salt stress. J. Plant Physiol. 2021, 267, 153544. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J.; et al. Consortia of Plant-Growth-Promoting Rhizobacteria isolated from halophytes improve response of eight crops to soil salinization and climate change conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- Yañez-Yazlle, M.F.; Romano-Armada, N.; Acreche, M.M.; Rajal, V.B.; Irazusta, V.P. Halotolerant bacteria isolated from extreme environments induce seed germination and growth of chia (Salvia hispanica L.) and quinoa (Chenopodium quinoa Willd.) under saline stress. Ecotoxicol. Environ. Saf. 2021, 218, 112273. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, C.; Sayyed, R.Z.; El Enshasy, H.A.; Vaidya, H.; Sharma, D.; Patel, N.; Malek, R.A.; Syed, A.; Elgorban, A.M.; Ahmad, K.; et al. Halotolerant microbial consortia for sustainable mitigation of salinity stress, growth promotion, and mineral uptake in tomato plants and soil nutrient enrichment. Sustainability 2021, 13, 8369. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl Stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Jabborova, D.; Kannepalli, A.; Davranov, K.; Narimanov, A.; Enakiev, Y.; Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Wirth, S.; Sayyed, R.Z.; et al. Co-inoculation of rhizobacteria promotes growth, yield, and nutrient contents in soybean and improves soil enzymes and nutrients under drought conditions. Sci. Rep. 2021, 11, 22081. [Google Scholar] [CrossRef] [PubMed]
- Mansour, E.; Mahgoub, H.A.M.; Mahgoub, S.A.; El-Sobky, E.-S.E.A.; Abdul-Hamid, M.I.; Kamara, M.M.; AbuQamar, S.F.; El-Rarabily, K.A.; Desoki, E.S.M. Enhancement of drought tolerance in diverse Vicia faba cultivars by inoculation with plant growth-promoting rhizobacteria under newly reclaimed soil conditions. Sci. Rep. 2021, 11, 24142. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Nawaz, F.; Hussain, M.B.; Ikram, R.M. Comparative effects of individual and consortia Plant Growth Promoting Bacteria on physiological and enzymatic mechanisms to confer drought tolerance in maize (Zea mays L.). J. Soil Sci. Plant Nutr. 2021, 21, 3461–3476. [Google Scholar] [CrossRef]
- Cortés-Patiño, S.; Vargas, C.; Álvarez-Flórez, F.; Bonilla, R.; Estrada-Bonilla, G. Potential of Herbaspirillum and Azospirillum consortium to promote growth of perennial ryegrass under water deficit. Microorganisms 2021, 1, 91. [Google Scholar] [CrossRef] [PubMed]
- Igiehon, O.N.; Babalola, O.O. Rhizobium and mycorrhizal fungal species improved soybean yield under drought stress conditions. Curr. Microbiol. 2021, 78, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; Prabhu, S.R.; de-Bashan, L.E.; Kloepper, J.W. Disclosure of exact protocols of fermentation, identity of microorganisms within consortia, formation of advanced consortia with microbe-based products. Biol. Fertil. Soils 2020, 56, 443–445. [Google Scholar] [CrossRef]
- Thomloudi, E.-E.; Tsalgatidou, P.C.; Douka, D.; Spantidos, T.-N.; Dimou, M.; Venieraki, A.; Katinakis, P. Multistrain versus single-strain plant growth promoting microbial inoculants-The compatibility issue. Hell. Plant Prot. J. 2019, 12, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Alori, E.T.; Emmanuel, O.C.; Glick, B.R.; Babalola, O.O. Plant-archaea relationships: A potential means to improve crop production in arid and semiarid regions. World J. Microbiol. Biotechnol. 2020, 36, 133. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Glick, B.R. Halotolerant-plant growth-promoting bacteria: Prospects for mitigating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Larimer, A.L.; Clay, K.; Bever, J.D. Synergism and context dependency of interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie legume. Ecology 2014, 95, 1045–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.I.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Paredes, A.; Valdés, G.; Nuti, M. Ecosystem functions of microbial consortia in sustainable sgriculture. Agronomy 2020, 10, 1902. [Google Scholar] [CrossRef]
- Veselaj, E.; Sallaku, G.; Balliu, A. Tripartite relationships in legume crops are plant-microorganism-specific and strongly influenced by salinity. Agriculture 2018, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, P.G.; Troncoso, M.D.P.; Monfil, V.O.; Estrella, A.H. Trichoderma species: Versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora indica: Potential and significance in plant stress tolerance. Front. Microbiol. 2016, 22, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miceli, A.; Moncada, A.; Vetrano, F. Use of microbial biostimulants to Increase the salinity tolerance of vegetable transplants. Agronomy 2021, 11, 143. [Google Scholar] [CrossRef]
- Liu, B.; Jing, D.; Liu, F.; Ma, H.; Liu, X.; Peng, L. Serendipita indica alleviates drought stress responses in walnut (Juglans regia L.) seedlings by stimulating osmotic adjustment and antioxidant defense system. Appl. Microbiol. Biotechnol. 2021, 105, 8951–8968. [Google Scholar] [CrossRef] [PubMed]
- Jangir, P.; Shekhawat, P.K.; Bishnoi, A.; Ram, H.; Soni, P. Role of Serendipita indica in enhancing drought tolerance in crops, Physiol. Mol. Plant Pathol. 2021, 116, 101691. [Google Scholar] [CrossRef]
- Li, L.; Li, L.; Wang, X.; Zhu, P.; Wu, H.; Qi, S. Plant growth-promoting endophyte Piriformospora indica alleviates salinity stress in Medicago truncatula. Plant Physiol. Biochem. 2017, 119, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Yun, P.; Xu, L.; Wang, S.S.; Shabala, L.; Shabala, S.; Zhang, W.Y. Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoots. Plant Growth Regul. 2018, 86, 323–331. [Google Scholar] [CrossRef]
- Ghorbani, A.; Omran, V.; Razavi, S.M.; Pirdashti, H.; Ranjbar, M. Piriformospora indica confers salinity tolerance on tomato (Lycopersicon esculentum Mill.) through amelioration of nutrient accumulation, K+/Na+ homeostasis and water status. Plant Cell Rep. 2019, 38, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial endophytic bacteria-Serendipita indica interaction for crop enhancement and resistance to phytopathogens. Front. Microbiol. 2019, 19, 2888. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic overview of the consequences of salt and drought stress on plants and the functioning of PGPBs supporting plant growth in these harsh environmental conditions.
Figure 1.
Schematic overview of the consequences of salt and drought stress on plants and the functioning of PGPBs supporting plant growth in these harsh environmental conditions.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gamalero, E.; Glick, B.R. Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress. Biology 2022, 11, 437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11030437
AMA Style
Gamalero E, Glick BR. Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress. Biology. 2022; 11(3):437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11030437
Chicago/Turabian StyleGamalero, Elisa, and Bernard R. Glick. 2022. "Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress" Biology 11, no. 3: 437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11030437
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.