
Environmental Status and Geomorphological Characterisation of Seven Black Coral Forests on the Sardinian Continental Shelf (NW Mediterranean Sea)

 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
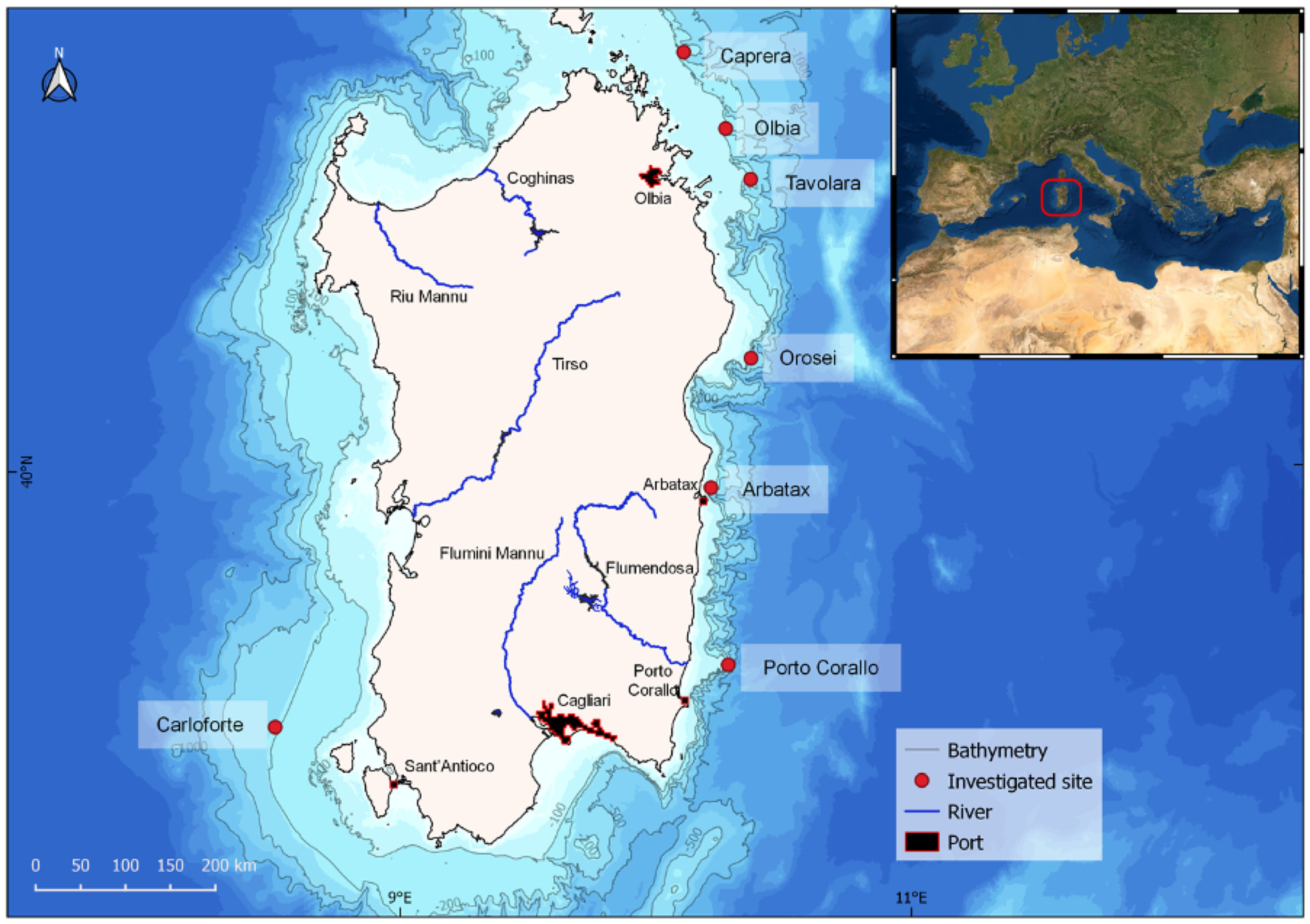
2.1. Study Area
2.2. ROV Surveys—Data Acquisition
2.3. Multibeam Bathymetry
2.4. Index Metrics
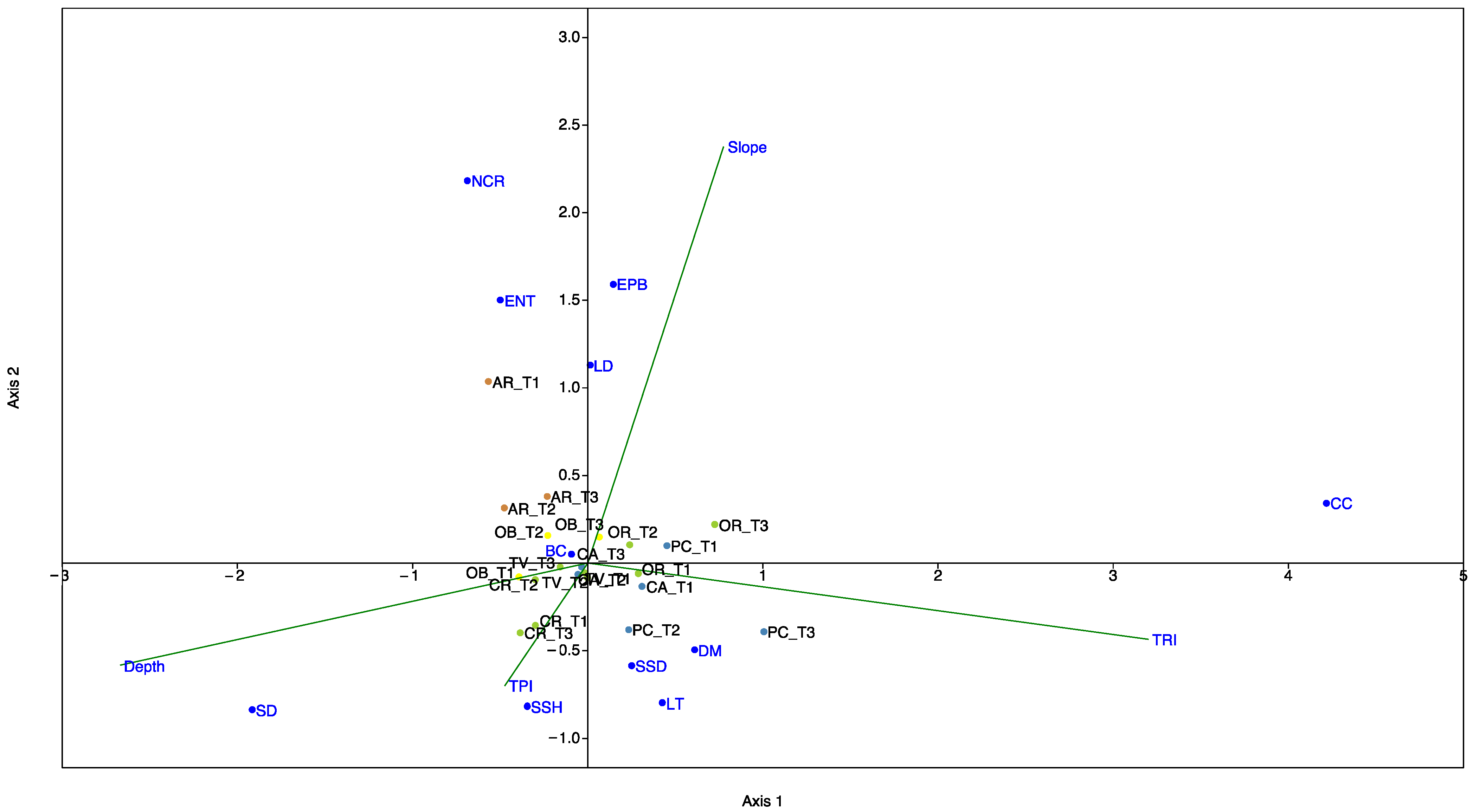
2.5. Statistical Analyses
3. Results
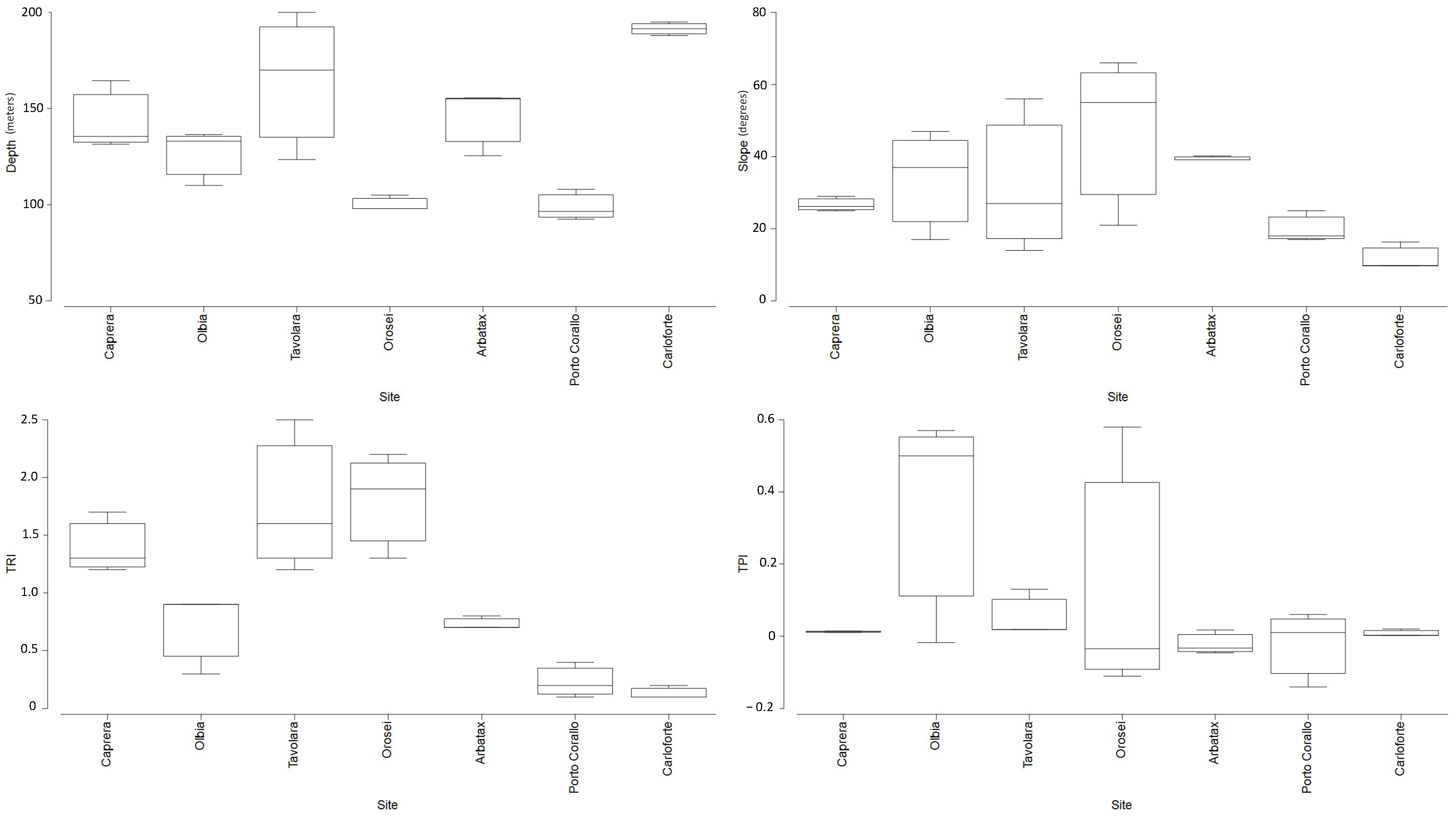
3.1. Geomorphological Setting
Coral Assemblages and Anthropogenic Impacts
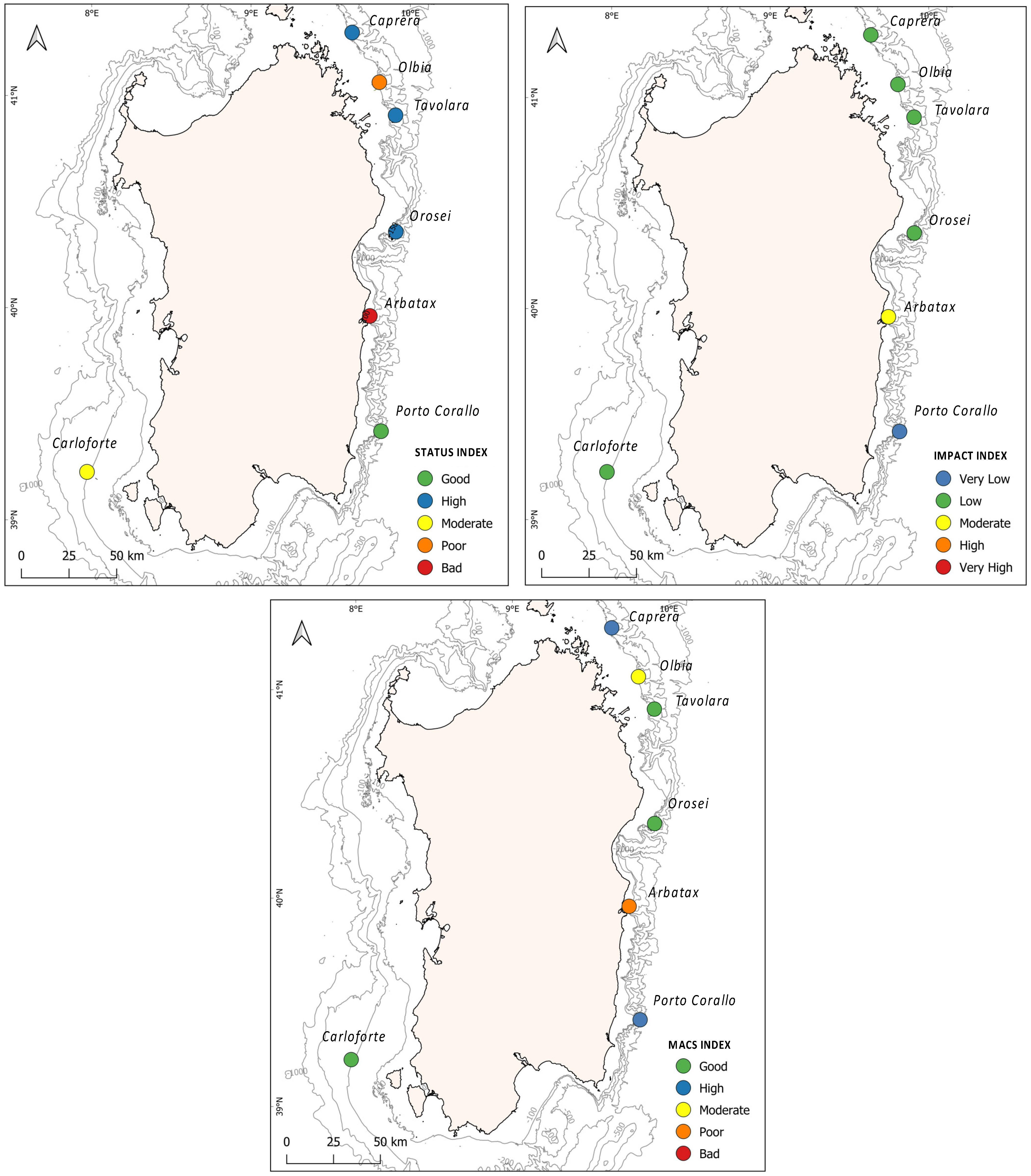
3.2. Index Outcomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rossi, S. The destruction of the “animal forests” in the oceans: Towards an over-simplification of the benthic ecosystems. Ocean Coast. Manag. 2013, 84, 77–85. [Google Scholar] [CrossRef]
- Gori, A.; Grinyó, J.; Dominguez-Carrió, C.; Ambroso, S.; López-González, P.J.; Gili, J.M.; Bavestrello, G.; Bo, M. Gorgonian and black coral assemblages in deep coastal bottoms and continental shelves of the Mediterranean Sea. In Mediterranean Cold-Water Corals: Past, Present and Future. Understanding the Deep-Sea Realms of Coral; Orejas, C., Jiménez, C., Eds.; Springer: Cham, Switzerland, 2019; Volume 9, pp. 245–248. [Google Scholar]
- Gori, A.; Bavestrello, G.; Grinyó, J.; Dominguez-Carrió, C.; Ambroso, S.; Bo, M. Animal Forests in Deep Coastal Bottoms and Continental Shelf of the Mediterranean Sea. In Marine Animal Forests. The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–27. [Google Scholar]
- Chimienti, G.; De Padova, D.; Mossa, M.; Mastrototaro, F. A mesophotic black coral forest in the Adriatic Sea. Sci. Rep. 2020, 10, 8504. [Google Scholar] [CrossRef] [PubMed]
- Prado, E.; Rodríguez-Basalo, A.; Cobo, A.; Ríos, P.; Sánchez, F. 3D fine-scale terrain variables from underwater photogrammetry: A new approach to benthic microhabitat modeling in a circalittoral Rocky shelf. Remote Sens. 2020, 12, 2466. [Google Scholar] [CrossRef]
- Rossi, P.; Ponti, M.; Righi, S.; Castagnetti, C.; Simonini, R.; Mancini, F.; Agrafiotis, P.; Bassani, L.; Bruno, F.; Cerrano, C.; et al. Needs and Gaps in Optical Underwater Technologies and Methods for the Investigation of Marine Animal Forest 3D-Structural Complexity. Front. Mar. Sci. 2021, 8, 171. [Google Scholar] [CrossRef]
- Bo, M.; Bavestrello, G.; Canese, S.; Giusti, M.; Salvati, E.; Angiolillo, M.; Greco, S. Characteristics of a black coral meadow in the twilight zone of the central Mediterranean Sea. Mar. Ecol. Prog. Ser. 2009, 397, 53–61. [Google Scholar] [CrossRef]
- Molodstova, T.N. Deep-sea fauna of European seas: An annotated species check-list of benthic invertebrates living deeper than 2000 m in the seas bordering Europe. Antipatharia. Zool. Bespozvon. 2014, 11, 3–7. [Google Scholar]
- IUCN. 2020. Available online: http://www.iucn.it/scheda.php?id=-512293580 (accessed on 27 August 2020).
- Grinyo’, J.; Gori, A.; Ambroso, S.; Purroy, A.; Calatayud, C.; Dominguez-Carrio, C.; Coppari, M.; Lo Iacono, C.; Lo´pez-Gonzalez, P.; Gili, J. Diversity, distribution and population size structure of deep Mediterranean gorgonian assemblages (Menorca Channel, Western Mediterranean Sea). Prog. Oceanogr. 2016, 145, 42–56. [Google Scholar] [CrossRef]
- Deidun, A.; Andaloro, F.; Bavestrello, G.; Canese, S.; Consoli, P.; Micallef, A.; Romeo, T.; Bo, M. First characterisation of a Leiopathes glaberrima (Cnidaria: Anthozoa: Antipatharia) forest in Maltese exploited fishing grounds. Ital. J. Zool. 2015, 82, 271–280. [Google Scholar]
- Chimienti, G.; Bo, M.; Taviani, M.; Mastrototaro, F. Occurrence and biogeography of Mediterranean cold-water corals. In Mediterranean Cold-Water Corals: Past, Present and Future. Understanding the Deep-Sea Realms of Coral; Orejas, C., Jiménez, C., Eds.; Springer: Cham, Switzerland, 2019; Volume 9, pp. 245–248. [Google Scholar]
- Moccia, D.; Cau, A.; Alvito, A.; Canese, S.; Cannas, R.; Bo, M.; Angiolillo, M.; Follesa, M.C. New sites expanding the “Sardinian cold-water coral province” extension: A new potential cold-water coral network? Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.J. Short-distance dispersal of black coral larvae: Inference from spatial analysis of colony genotypes. Mar. Ecol. Prog. Ser. 1998, 163, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Costantini, F.; Addamo, A.M.; Machordom, A.; Abbiati, M. Genetic connectivity and conservation of temperate and cold-water habitat-forming corals. In Marine Animal Forests. The Ecology of Benthic Biodiversity Hotspot; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 1061–1082. [Google Scholar]
- Costantini, F.; Ferrario, F.; Abbiati, M. Chasing genetic structure in coralligenous reef invertebrates: Patterns, criticalities and conservation issues. Sci. Rep. 2018, 8, 5844. [Google Scholar] [CrossRef] [PubMed]
- Terzin, M.; Paletta, M.G.; Matterson, K.; Coppari, M.; Bavestrello, G.; Abbiati, M.; Bo, M.; Costantini, F. Population genomic structure of the black coral Antipathella subpinnata in Mediterranean Vulnerable Marine Ecosystems. Coral Reefs 2021, 40, 751–766. [Google Scholar] [CrossRef]
- Gori, A.; Rossi, S.; Berganzo, E.; Lluis Pretus, J.; Dale, M.R.T.; Gili, J.M.; Pretus, J.; Dale, M.R.T.; Gili, J.M. Spatial distribution patterns of the gorgonians Eunicella singularis; Paramuricea clavata; Leptogorgia sarmentosa (Cape of Creus, Northwestern Mediterranean Sea). Mar. Biol. 2011, 158, 143–158. [Google Scholar] [CrossRef]
- Cau, A.; Moccia, D.; Follesa, M.C.; Alvito, A.; Canese, S.; Angiolillo, M.; Cuccu, D.; Bo, M.; Cannas, R. Coral forests diversity in the outer shelf of the south Sardinian continental margin. Deep Sea Res. Part I Oceanogr. Res. 2017, 122, 60–70. [Google Scholar] [CrossRef]
- Diaz, R.J.; Solan, M.; Valente, R.M. A review of approaches for classifying benthic habitats and evaluating habitat quality. J. Environ. Manag. 2004, 73, 165–181. [Google Scholar] [CrossRef]
- Rossi, S.; Bramanti, L.; Gori, A.; Orejas, C. Marine Animal Forests. The Ecology of Benthic Biodiversity Hotspots; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Cau, A.; Follesa, M.C.; Moccia, D.; Bellodi, A.; Mulas, A.; Bo, M.; Canese, S.; Angiolillo, M.; Cannas, R. Leiopathes glaberrima millennial forest from SW Sardinia as nursery ground for the small spotted catshark Scyliorhinus canicular. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 731–735. [Google Scholar] [CrossRef]
- Carbonara, P.; Zupa, W.; Follesa, M.C.; Cau, A.; Capezzuto, F.; Chimienti, G.; D’Onghia, G.; Lembo, G.; Pesci, P.; Porcu, C.; et al. Exploring a deep-sea vulnerable marine ecosystem: Isidella elongata (Esper, 1788) species assemblages in the Western and Central Mediterranean. Deep Sea Res. Part I Oceanogr. Res. 2020, 166, 103406. [Google Scholar] [CrossRef]
- Bo, M.; Bava, S.; Canese, S.; Angiolillo, M.; Cattaneo-Vietti, R.; Bavestrello, G. Fishing impact on deep Mediterranean rocky habitats as revealed by ROV investigation. Biol. Conserv. 2014, 171, 167–176. [Google Scholar] [CrossRef]
- Ragnarsson, S.Á.; Burgos, J.M.; Kutti, T.; van den Beld, I.; Egilsdóttir, H.; Arnaud-Haond, S.; Grehan, A. The Impact of Anthropogenic Activity on Cold-Water Corals. In Marine Animal Forests. The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 989–1023. [Google Scholar]
- Bo, M.; Numa, C.; del Mar Otero, M.; Orejas, C.; Garrabou, J.; Cerrano, C.; Kružic, P.; Antoniadou, C.; Aguilar, R.; Kipson, S.; et al. Overview of the Conservation Status of Mediterranean Anthozoa; IUCN: Gland, Switzerland; Malaga, Spain, 2017. [Google Scholar] [CrossRef] [Green Version]
- OSPAR Commission. Case Reports for the OSPAR List of Threatened and/or Declining Species and Habitats; Biodiversity Ecosystems; OSPAR Commission: London, UK, 2008; ISBN 978-1-905859-97-9. [Google Scholar]
- UNEP. Report on the Expert Workshop on Ecological Criteria and Biogeographic Classification System for Marine Areas in Need of Protection; UNEP: Azores, Portugal, 2007. [Google Scholar]
- REC.CM-GFCM/29/2005/1; Management of Certain Fisheries Exploiting Demersal and Deep-Water Species. FAO GFCM: Rome, Italy, 2005.
- REC.CM-GFCM/30/2006/3; Establishment of Fisheries Restricted Areas in Order to Protect the Deep Sea Sensitive Habitats. FAO GFCM: Instanbul, Turkey, 2006.
- Bo, M.; Bavestrello, G.; Angiolillo, M.; Calcagnile, L.; Canese, S.; Cannas, R.; Cau, A.; D’Elia, M.; D’Oriano, F.; Follesa, M.C.; et al. Persistence of Pristine Deep-Sea Coral Gardens in the Mediterranean Sea (SW Sardinia). PLoS ONE 2015, 10, e0119393. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Directive of the European parliament and the council establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive), Directive 2008/56/ EC, OJ L 164. J. Counc. Decis. 2008, 164, 19–40. [Google Scholar]
- Enrichetti, F.; Bo, M.; Morri, C.; Montefalcone, M.; Toma, M.; Bavestrello, G.; Tunesi, L.; Canese, S.; Giusti, M.; Salvati, E.; et al. Assessing the environmental status of temperate mesophotic reefs: A new, integrated methodological approach. Ecol. Indic. 2019, 102, 218–229. [Google Scholar] [CrossRef]
- Piazzi, L.; Bianchi, C.N.; Cecchi, E.; Gatti, G.; Guala, I.; Morri, C.; Sartoretto, S.; Serena, F.; Montefalcone, M. What’s in an index? Comparing the ecological information provided by two indices to assess the status of coralligenous reefs in the NW Mediterranean Sea. Aquat. Conserv. Mar. Fresh. Ecosyst. 2017, 27, 1091–1100. [Google Scholar] [CrossRef]
- Ferrigno, F.; Russo, G.F.; Sandulli, R. Coralligenous Bioconstructions Quality Index (CBQI): A synthetic indicator to assess the status of different types of coralligenous habitats. Ecol. Ind. 2017, 82, 271–279. [Google Scholar] [CrossRef]
- Deter, J.; Descamp, P.; Ballesta, L.; Boissery, P.; Holon, F. A preliminary study toward an index based on coralligenous assemblages for the ecological status assessment of Mediterranean French coastal waters. Ecol. Ind. 2012, 20, 345–352. [Google Scholar] [CrossRef]
- Gatti, G.; Bianchi, C.M.; Morri, C.; Montefalcone, M.; Sartoretto, S. Coralligenous reefs state along anthropized coasts: Application and validation of the COARSE index, based on a rapid visual assessment (RVA) approach. Ecol. Ind. 2015, 52, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Cánovas-Molina, A.; Montefalcone, M.; Bavestrello, G.; Cau, A.; Bianchi, C.N.; Morri, C.; Canese, S.; Bo, M. A new ecological index for the status of mesophotic megabenthic assemblages in the mediterranean based on ROV photography and video footage. Cont. Shelf Res. 2016, 121, 13–20. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Bonifazi, A.; Argenti, L.; Ingrassia, M.; Casalbore, D.; Aguzzi, L.; Viaggiu, E.; Le Foche, M.; Chiocci, F. Geomorphological characterization, spatial distribution and environmental status assessment of coralligenous reefs along the Latium continental shelf. Ecol. Indic. 2021, 131, 108219. [Google Scholar] [CrossRef]
- Elliott, M.; Burdon, D.; Atkins, J.P.; Borja, A.; Cormier, R.; de Jonge, V.N.; Turner, R.K. And DPSIR begat DAPSI(W)R(M)!—A unifying framework for marine environmental management. Mar. Pollut. Bull. 2017, 118, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Würtz, M. Mediterranean Submarine Canyons: Ecology and Governance; IUCN: Gland, Switzerland; Malaga, Spain, 2012; pp. 1–216. [Google Scholar]
- Deiana, G.; Lecca, L.; Melis, R.T.; Soldati, M.; Demurtas, V.; Orrù, P.E. Submarine geomorphology of the southwestern Sardinian continental shelf (Mediterranean sea): Insights into the last glacial maximum sea-level changes and related environments. Water 2021, 13, 155. [Google Scholar] [CrossRef]
- Alvito, A.; Bellodi, A.; Cau, A.; Moccia, D.; Mulas, A.; Palmas, F.; Pesci, P.; Follesa, M.C. Amount and distribution of benthic marine litter along Sardinian fishing grounds (CW Mediterranean Sea). Waste Manag. 2018, 75, 131–140. [Google Scholar] [CrossRef]
- Cau, A.; Franceschini, S.; Moccia, D.; Gorule, P.A.; Agus, B.; Bellodi, A.; Cannas, R.; Carugati, L.; Cuccu, D.; Dessì, C.; et al. Scattered accumulation hotspots of macro-litter on the seafloor: Insights for mitigation actions. Environ. Pollut. 2021, 292, 118338. [Google Scholar] [CrossRef] [PubMed]
- Galgani, F.; Pham, C.K.; Claro, F.; Consoli, P. Marine animal forests as useful indicators of entanglement by marine litter. Mar. Poll. Bull. 2018, 135, 735–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Czechowska, K.; Feldens, P.; Tuya, F.; Cosme de Esteban, M.; Espino, F.; Haroun, R.; Schönke, M.; Otero-Ferrer, F. Testing Side-Scan Sonar and Multibeam Echosounder to Study Black Coral Gardens: A Case Study from Macaronesia. Remote Sens. 2020, 12, 3244. [Google Scholar] [CrossRef]
- Tong, R.; Purser, A.; Unnithan, V.; Guinan, J. Multivariate statistical analysis of distribution of deep-water gorgonian corals in relation to seabed topography on the norwegian margin. PLoS ONE 2012, 7, e43534. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Hammer, H.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeo Electr. 2001, 4, 9. [Google Scholar]
- Grinyó, J.; Lo Iacono, C.; Pierdomenico, M.; Conlon, S.; Corbera, G.; Gràcia, E. Evidences of human impact on megabenthic assemblages of bathyal sediments in the Alboran Sea (western Mediterranean). Deep Sea Res. Part I Oceanogr. Res. 2020, 165, 103369. [Google Scholar] [CrossRef]
- Piazzi, L.; Atzori, F.; Cadoni, N.; Cinti, M.F.; Frau, F.; Pansini, A.; Pinna, F.; Stipcich, P.; Ceccherelli, G. Animal Forest Mortality: Following the Consequences of a Gorgonian Coral Loss on a Mediterranean Coralligenous Assemblage. Diversity 2021, 13, 133. [Google Scholar] [CrossRef]
- Perry, A.L.; Blanco, J.; Fournier, N.; García, S.; Marín, P. Unmanaged = Unprotected: Europe’s Marine Paper Parks. Oceana, Brussels. 2020. Available online: https://europe.oceana.org/en/publications/reports/unmanaged-unprotected-europes-marine-paper-parks (accessed on 10 September 2021).
- Fabri, M.C.; Pedel, L.; Beuck, L.; Galgani, F.; Hebbeln, D.; Freiwald, A. Megafauna of vulnerable marine ecosystems in French mediterranean submarine canyons: Spatial distribution and anthropogenic impacts. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 104, 184–207. [Google Scholar] [CrossRef] [Green Version]
- Moccia, D.; Cau, A.; Bramanti, L.; Carugati, L.; Canese, S.; Follesa, M.C.; Cannas, R. Spatial distribution and habitat characterization of marine animal forest assemblages along nine submarine canyons of Eastern Sardinia (central Mediterranean Sea). Deep Sea Res. Part I Oceanogr. Res. 2021, 167, 103422. [Google Scholar] [CrossRef]
- Ingrassia, M.; Di Bella, L. Black coral distribution in the italian seas: A review. Diversity 2021, 13, 334. [Google Scholar] [CrossRef]
- Altuna, A.; Poliseno, A. Taxonomy, Genetics and Biodiversity of Mediterranean Deep-Sea Corals and Cold-Water Corals. In Mediterranean Cold-Water Corals: Past, Present and Future. Understanding the Deep-Sea Realms of Coral; Orejas, C., Jiménez, C., Eds.; Springer: Cham, Switzerland, 2019; Volume 9, pp. 121–156. [Google Scholar]
- Angeletti, L.; Taviani, M.; Canese, S.; Foglini, F.; Mastrototaro, F.; Argnani, A.; Trincardi, F.; Bakran-Petricioli, T.; Ceregato, A.; Chimienti, G.; et al. New deep-water cnidarian sites in the southern Adriatic Sea. Mediterr. Mar. Sci. 2014, 15, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Grinyo´, J.; Gori, A.; Greenacre, M.; Requena, S.; Canepa, A.; Lo Iacono, C.; Ambroso, S.; Purroy, A.; Gili, J. Megabenthic assemblages in the continental shelf edge and upper slope of the Menorca Channel, Western Mediterranean Sea. Prog. Oceanogr. 2018, 162, 40–51. [Google Scholar] [CrossRef]
- Ingrassia, M.; Macelloni, L.; Bosman, A.; Chiocci, F.; Cerrano, C.; Martorelli, E. Black coral (Anthozoa, Antipatharia) forest near the western Pontine Islands (Tyrrhenian Sea). Mar. Biod. 2016, 46, 285–290. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.A.; Priede, I.G.; Buhl-Mortensen, P.; Gheerardyn, H.; King, N.K.; Raes, M. Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins: Biological structures and biodiversity. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Folkersen, M.V.; Fleming, C.M.; Hasan, S. The economic value of the deep sea: A systematic review and meta-analysis. Mar. Policy 2018, 94, 71–80. [Google Scholar] [CrossRef]
- Cerrano, C.; Danovaro, R.; Gambi, C.; Pusceddu, A.; Riva, A.; Schiaparelli, S. Gold coral (Savalia savaglia) and gorgonian forests enhance benthic biodiversity and ecosystem functioning in the mesophotic zone. Biod. Consev. 2010, 19, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Massi, D.; Vitale, S.; Titone, A.; Milisenda, G.; Gristina, M.; Fiorentino, F. Spatial distribution of the black coral Leiopathes glaberrima (Esper, 1788) (Antipatharia: Leiopathidae) in the Mediterranean: A prerequisite for protection of Vulnerable Marine Ecosystems (VMEs). Eur. Zool. J. 2018, 85, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Thurber, A.R.; Sweetman, A.K.; Narayanaswamy, B.E.; Jones, D.O.B.; Ingels, J.; Hansman, R.L. Ecosystem function and services provided by the deep sea. Biogeosciences 2014, 11, 3941–3963. [Google Scholar] [CrossRef] [Green Version]
- Fabri, M.C.; Vinha, B.; Allais, A.G.; Bouhier, M.E.; Dugornay, O.; Gaillot, A.; Arnaubec, A. Evaluating the ecological status of cold-water coral habitats using non-invasive methods: An example from Cassidaigne canyon, northwestern Mediterranean Sea. Progr. Oceanogr. 2019, 178, 102172. [Google Scholar] [CrossRef] [Green Version]
- Bo, M.; Coppari, M.; Betti, F.; Enrichetti, F.; Bertolino, M.; Massa, F.; Bava, S.; Gay, G.; Cattaneo-Vietti, R.; Bavestrello, G. The high biodiversity and vulnerability of two Mediterranean bathyal seamounts support the need for creating offshore protected areas. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 543–566. [Google Scholar] [CrossRef]
- Lopez-Fernandez, P.; Bianchelli, S.; Pusceddu, A.; Calafat, A.; Danovaro, R.; Canals, M. Bioavailable compounds in sinking particulate organic matter, Blanes Canyon, NW Mediterranean Sea: Effects of a large storm and sea surface biological processes. Progr. Oceanog. 2013, 118, 108–121. [Google Scholar] [CrossRef]
- Robertson, C.M.; Demopoulos, A.W.J.; Bourque, J.R.; Mienis, F.; Duineveld, G.C.A.; Lavaleye, M.S.S.; Koivisto, R.K.K.; Brooke, S.D.; Ross, S.W.; Rhode, M.; et al. Submarine canyons influence macrofaunal diversity and density patterns in the deep-sea benthos. Deep Sea Res. Part I Oceanogr. Res. 2020, 159, 103249. [Google Scholar] [CrossRef]
- Fernandez-Arcaya, U.; Ramirez-Llodra, E.; Aguzzi, J.; Allcock, A.L.; Davies, J.S.; Dissanayake, A.; Harris, P.; Howell, K.; Huvenne, V.A.I.; Macmillan-Lawler, M.; et al. Ecological Role of Submarine Canyons and Need for Canyon Conservation: A Review. Front. Mar. Sci. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- De Leo, F.; Smith, C.; Rowden, A.; Bowden, D.; Clark, M. Submarine canyons: Hotspots of benthic biomass and productivity in the deep sea. Proc. R. Soc. 2010, 277, 2783–2792. [Google Scholar] [CrossRef] [Green Version]
- Revenga Martínez de Pazos, S. The fisheries importance of four submarine canyons in the Spanish Mediterranean Sea: Cap de Creus, Palamós, Cape Tiñoso and Alborán platform canyons. In Mediterranean Submarine Canyons: Ecology and Governance; Würtz, M., Ed.; IUCN: Gland, Switzerland; Malaga, Spain, 2012; pp. 51–55. [Google Scholar]
- Freitas, L.M.; Oliveira, M.D.M.; Leão, Z.M.A.N.; Kikuchi, R.K.P. Effects of turbidity and depth on the bioconstruction of the Abrolhos reefs. Coral Reefs 2019, 38, 241–253. [Google Scholar] [CrossRef]
- Fisher, R.; Bessell-Browne, P.; Jones, R. Synergistic and antagonistic impacts of suspended sediments and thermal stress on corals. Nat. Commun. 2019, 10, 2346. [Google Scholar] [CrossRef] [Green Version]
- Carugati, L.; Moccia, D.; Bramanti, L.; Cannas, R.; Follesa, M.C.; Salvadori, S.; Cau, A. Deep-Dwelling Populations of Mediterranean Corallium rubrum and Eunicella cavolini: Distribution, Demography, and Co-Occurrence. Biology 2022, 11, 333. [Google Scholar] [CrossRef]
- Giusti, M.; Canese, S.; Fourt, M.; Bo, M.; Innocenti, C.; Goujard, A.; Daniel, B.; Angeletti, L.; Taviani, M.; Aquilina, L.; et al. Coral forests and Derelict Fishing Gears in submarine canyon systems of the Ligurian Sea. Progr. Oceanogr. 2019, 178, 102186. [Google Scholar] [CrossRef]
- Cau, A.; Alvito, A.; Moccia, D.; Canese, S.; Pusceddu, A.; Rita, C.; Angiolillo, M.; Follesa, M.C. Submarine canyons along the upper Sardinian slope (Central Western Mediterranean) as repositories for derelict fishing gears. Mar. Poll. Bull. 2017, 123, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Llodra, E.; De Mol, B.; Company, J.B.; Coll, M.; Sardà, F. Effects of natural and anthropogenic processes in the distribution of marine litter in the deep Mediterranean Sea. Progr. Oceanogr. 2013, 118, 273–287. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Status Index (Is) | Impact Index (Ii) | Is | Ii | MACS | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | SR | BC | CC | DM | SSD | SSH | SD | ENT | NCR | EPB | LD | LT | |||
| Caprera | 67 | 78 | 33 | 100 | 67 | 100 | 33 | 11 | 33 | 44 | 33 | 78 | 74 | 39 | 68 |
| Olbia | 67 | 56 | 0 | 44 | 44 | 33 | 56 | 11 | 26 | 33 | 33 | 78 | 41 | 39 | 51 |
| Tavolara | 89 | 89 | 0 | 89 | 78 | 67 | 40 | 33 | 33 | 33 | 33 | 67 | 69 | 40 | 64 |
| Orosei | 56 | 56 | 100 | 78 | 55 | 67 | 29 | 33 | 22 | 44 | 33 | 67 | 69 | 38 | 65 |
| Arbatax | 22 | 44 | 0 | 11 | 22 | 45 | 44 | 11 | 89 | 33 | 44 | 67 | 24 | 48 | 38 |
| Porto Corallo | 67 | 44 | 67 | 56 | 56 | 44 | 0 | 11 | 11 | 11 | 22 | 78 | 56 | 22 | 67 |
| Carloforte | 67 | 67 | 0 | 45 | 44 | 100 | 89 | 22 | 11 | 22 | 22 | 78 | 54 | 41 | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moccia, D.; Carugati, L.; Follesa, M.C.; Cannas, R.; Carbonara, P.; Pusceddu, A.; Cau, A. Environmental Status and Geomorphological Characterisation of Seven Black Coral Forests on the Sardinian Continental Shelf (NW Mediterranean Sea). Biology 2022, 11, 732. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050732
Moccia D, Carugati L, Follesa MC, Cannas R, Carbonara P, Pusceddu A, Cau A. Environmental Status and Geomorphological Characterisation of Seven Black Coral Forests on the Sardinian Continental Shelf (NW Mediterranean Sea). Biology. 2022; 11(5):732. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050732
Chicago/Turabian StyleMoccia, Davide, Laura Carugati, Maria Cristina Follesa, Rita Cannas, Pierluigi Carbonara, Antonio Pusceddu, and Alessandro Cau. 2022. "Environmental Status and Geomorphological Characterisation of Seven Black Coral Forests on the Sardinian Continental Shelf (NW Mediterranean Sea)" Biology 11, no. 5: 732. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050732