Biostimulant Capacity of Chlorella and Chlamydopodium Species Produced Using Wastewater and Centrate



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Selected Microorganisms
2.2. Biomass Production
2.3. Assessment of the Biomass Productivity and Nutrient Consumption
2.4. Biomass Processing
2.5. Assessment of the Biostimulant Capacity
2.5.1. Germination Index Assessment of Watercress Seeds (Lepidium sativum L.)
2.5.2. Assessment of the Adventitious Soybean (Glycine max L.) Root Induction
2.5.3. Excised Cucumber (Cucumis sativus L.) Expansion Test
2.5.4. Wheat (Triticum aestivum L.) Chlorophyll Retention Assay
2.6. Statistical Analysis
3. Results and Discussion
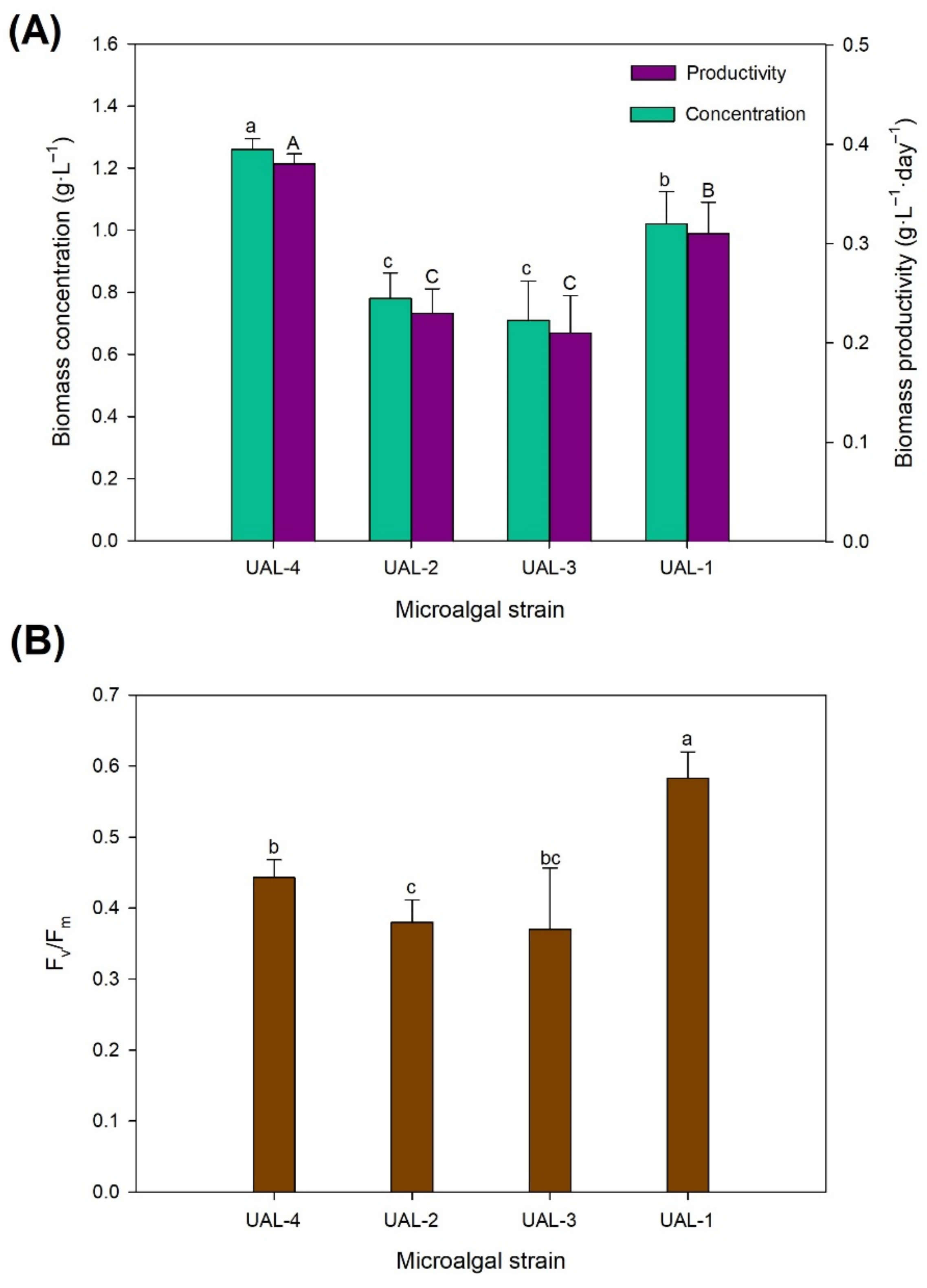
3.1. Biomass Productivity
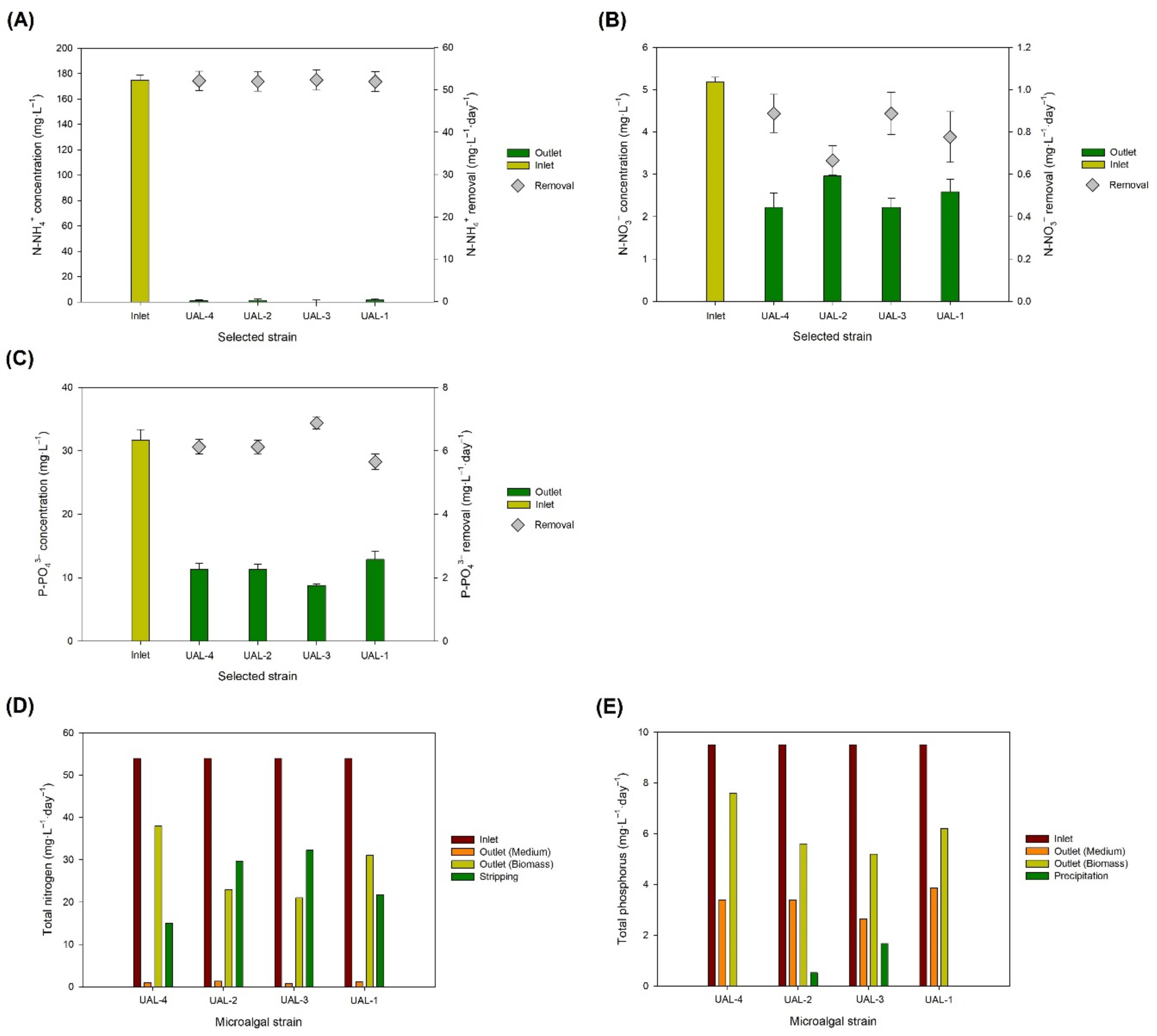
3.2. Nutrient Consumption
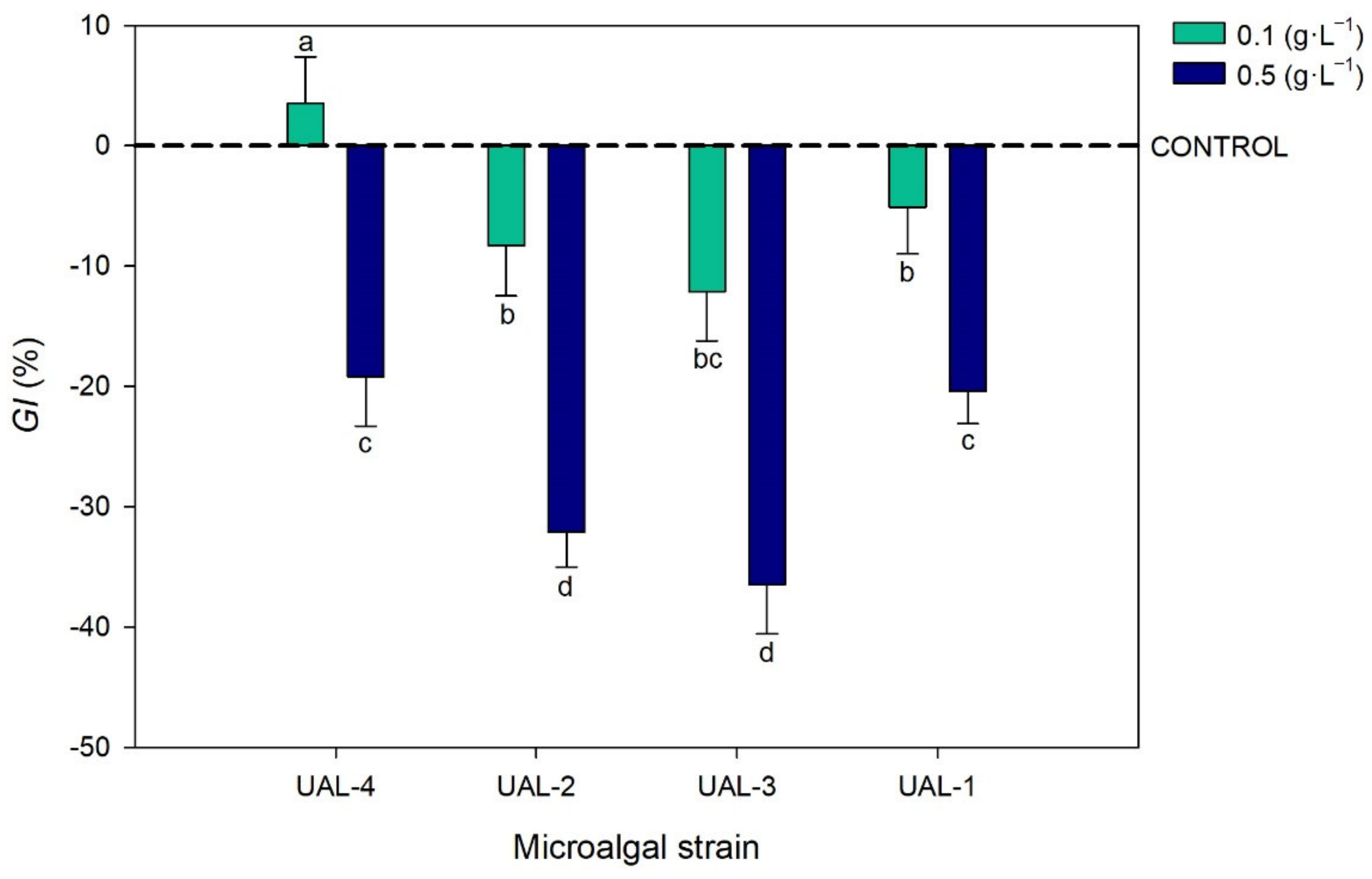
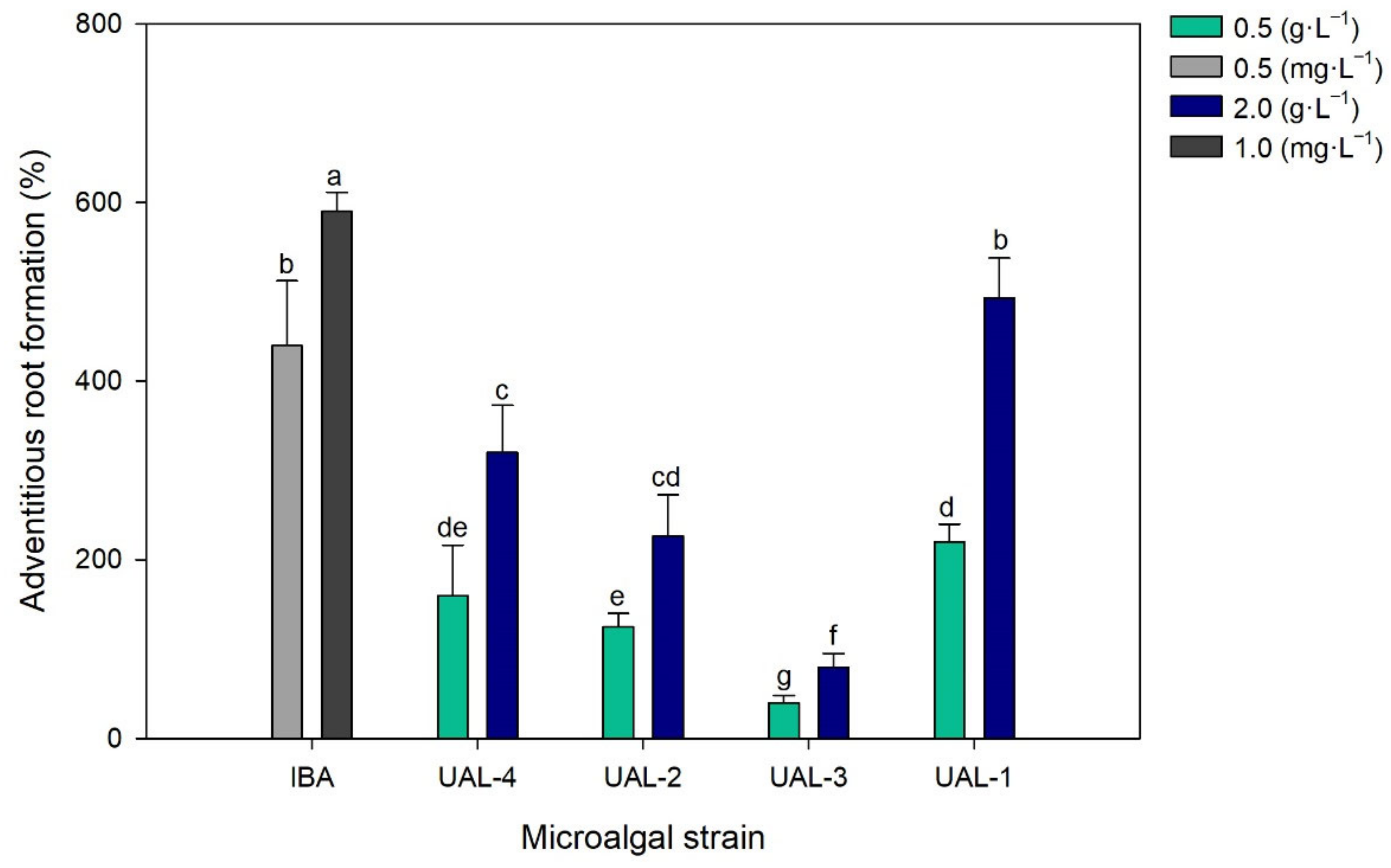
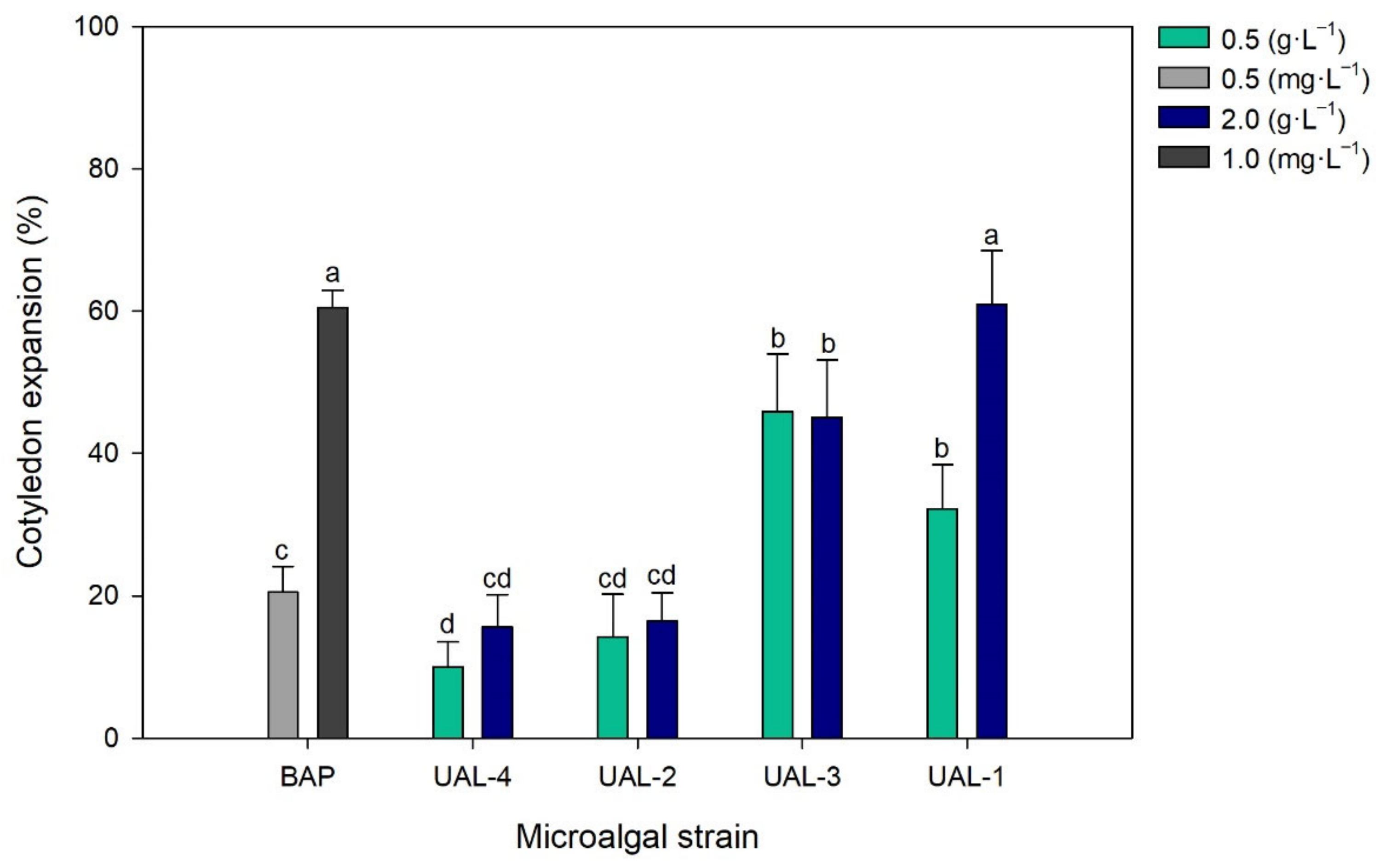
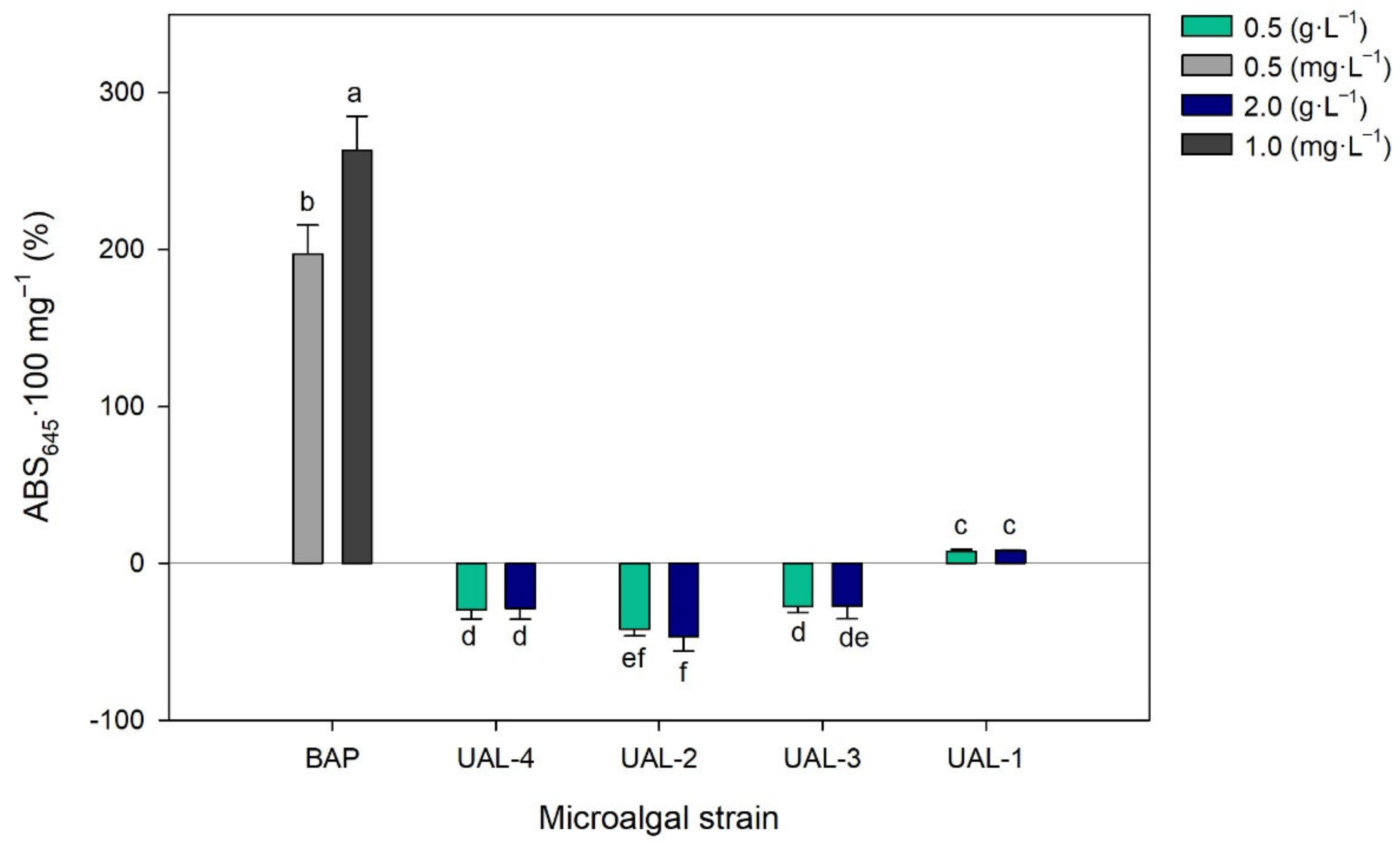
3.3. Biostimulant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Odegard, I.Y.R.; van der Voet, E. The future of food—Scenarios and the effect on natural resource use in agriculture in 2050. Ecol. Econ. 2014, 97, 51–59. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano-Monserrate, M.A.; Carvajal-Lara, C.; Urgilés-Sanchez, R.; Ruano, M.A. Deforestation as an indicator of environmental degradation: Analysis of five European countries. Ecol. Indic. 2018, 90, 1–8. [Google Scholar] [CrossRef]
- Barros, M.V.; Salvador, R.; de Francisco, A.C.; Piekarski, C.M. Mapping of research lines on circular economy practices in agriculture: From waste to energy. Renew. Sustain. Energy Rev. 2020, 131, 109958. [Google Scholar] [CrossRef]
- Povero, G.; Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y. Microalgae: New Source of Plant Biostimulants. Agronomy 2020, 10, 1240. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Elvira, N.-L.; del Carmen Cerón-García, M.; López-Rodríguez, M.; Acién-Fernández, F.G.; Molina-Grima, E. Biostimulants obtained after pilot-scale high-pressure homogenization of Scenedesmus sp. grown in pig manure. Algal Res. 2020, 52, 102123. [Google Scholar] [CrossRef]
- Lafarga, T.; Pieroni, C.; D’Imporzano, G.; Maggioni, L.; Adani, F.; Acién, G. Consumer Attitudes towards Microalgae Production and Microalgae-Based Agricultural Products: The Cases of Almería (Spain) and Livorno (Italy). ChemEngineering 2021, 5, 27. [Google Scholar] [CrossRef]
- Lafarga, T.; Rodríguez-Bermúdez, R.; Morillas-España, A.; Villaró, S.; García-Vaquero, M.; Morán, L.; Sánchez-Zurano, A.; González-López, C.V.; Acién-Fernández, F.G. Consumer knowledge and attitudes towards microalgae as food: The case of Spain. Algal Res. 2021, 54, 102174. [Google Scholar] [CrossRef]
- Llamas, B.; Suárez-Rodríguez, M.C.; González-López, C.V.; Mora, P.; Acién, F.G. Techno-economic analysis of microalgae related processes for CO2 bio-fixation. Algal Res. 2021, 57, 102339. [Google Scholar] [CrossRef]
- Acién Fernández, F.G.; Gómez-Serrano, C.; Fernández-Sevilla, J.M. Recovery of Nutrients from Wastewaters Using Microalgae. Front. Sustain. Food Syst. 2018, 2, 00059. [Google Scholar] [CrossRef] [Green Version]
- Wollmann, F.; Dietze, S.; Ackermann, J.; Bley, T.; Walther, T.; Steingroewer, J.; Krujatz, F. Microalgae wastewater treatment: Biological and technological approaches. Eng. Life Sci. 2019, 19, 860–871. [Google Scholar] [CrossRef] [Green Version]
- Bongiorno, T.; Foglio, L.; Proietti, L.; Vasconi, M.; Lopez, A.; Pizzera, A.; Carminati, D.; Tava, A.; Vizcaíno, A.J.; Alarcón, F.J.; et al. Microalgae from Biorefinery as Potential Protein Source for Siberian Sturgeon (A. baerii) Aquafeed. Sustainability 2020, 12, 8779. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Renuka, N.; Prasanna, R.; Sood, A.; Ahluwalia, A.S.; Bansal, R.; Babu, S.; Singh, R.; Shivay, Y.S.; Nain, L. Exploring the efficacy of wastewater-grown microalgal biomass as a biofertilizer for wheat. Environ. Sci. Pollut. Res. 2016, 23, 6608–6620. [Google Scholar] [CrossRef]
- Morillas-España, A.; Lafarga, T.; Gómez-Serrano, C.; Acién-Fernández, F.G.; González-López, C.V. Year-long production of Scenedesmus almeriensis in pilot-scale raceway and thin-layer cascade photobioreactors. Algal Res. 2020, 51, 102069. [Google Scholar] [CrossRef]
- Morillas-España, A.; Lafarga, T.; Acién-Fernández, F.G.; Gómez-Serrano, C.; González-López, C.V. Annual production of microalgae in wastewater using pilot-scale thin-layer cascade photobioreactors. J. Appl. Phycol. 2021, 33, 3861–3871. [Google Scholar] [CrossRef]
- Gómez, C.; Guzmán-Carrasco, A.; Lafarga, T.; Acién-Fernández, F.G. Optimization of a new culture medium for the large-scale production of protein rich Arthrospira platensis (Oscillatoriales, Cyanophyceae). J. Phycol. 2020, 27, 636–644. [Google Scholar] [CrossRef]
- Zhao, Z.R.; Wu, Z.L.; Huang, G.Q.; Li, G.R. An improved disk bioassay for determining activities of plant growth regulators. J. Plant Growth Regul. 1992, 11, 209–213. [Google Scholar] [CrossRef]
- Kuhnle, J.A.; Fuller, G.; Corse, J.; Mackey, B.E. Antisenescent Activity of Natural Cytokinins. Physiol. Plant. 1977, 41, 14–21. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F.; Acien-Fernandez, F.G.; Molina-Grima, E. Microalgae research worldwide. Algal Res. 2018, 35, 50–60. [Google Scholar] [CrossRef]
- Acién, F.G.; Molina, E.; Fernández-Sevilla, J.M.; Barbosa, M.; Gouveia, L.; Sepúlveda, C.; Bazaes, J.; Arbib, Z. Economics of microalgae production. In Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 485–503. [Google Scholar] [CrossRef]
- Ruiz, J.; Álvarez, P.; Arbib, Z.; Garrido, C.; Barragán, J.; Perales, J.A. Effect of Nitrogen and Phosphorus Concentration on Their Removal Kinetic in Treated Urban Wastewater by Chlorella vulgaris. Int. J. Phytoremediat. 2011, 13, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Mtaki, K.; Kyewalyanga, M.S.; Mtolera, M.S.P. Supplementing wastewater with NPK fertilizer as a cheap source of nutrients in cultivating live food (Chlorella vulgaris). Ann. Microbiol. 2021, 71, 7. [Google Scholar] [CrossRef]
- Touloupakis, E.; Tartari, G.; Zittelli, G.C.; Torzillo, G. Growth and photosynthetic performance of Chlamydopodium fusiforme cells cultivated in BG11 and Bristol media. J. Appl. Phycol. 2019, 32, 145–152. [Google Scholar] [CrossRef]
- Geider, R.J.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Arbib, Z.; Ruiz, J.; Álvarez-Díaz, P.; Garrido-Pérez, C.; Barragan, J.; Perales, J.A. Photobiotreatment: Influence of nitrogen and phosphorus ratio in wastewater on growth kinetics of scenedesmus obliquus. Int. J. Phytoremediat. 2013, 15, 774–788. [Google Scholar] [CrossRef]
- Santabarbara, S.; Villafiorita Monteleone, F.; Remelli, W.; Rizzo, F.; Menin, B.; Casazza, A.P. Comparative excitation-emission dependence of the FV/FM ratio in model green algae and cyanobacterial strains. Physiol. Plant. 2019, 166, 351–364. [Google Scholar] [CrossRef]
- Ledda, C.; Romero Villegas, G.I.; Adani, F.; Acién Fernández, F.G.; Molina Grima, E. Utilization of centrate from wastewater treatment for the outdoor production of Nannochloropsis gaditana biomass at pilot-scale. Algal Res. 2015, 12, 17–25. [Google Scholar] [CrossRef]
- Ciardi, M.; Gómez-Serrano, C.; del Mar Morales-Amaral, M.; Acién, G.; Lafarga, T.; Fernandez-Sevilla, J.M. Optimisation of Scenedesmus almeriensis production using pig slurry as the sole nutrient source. Algal Res. 2022, 61, 102580. [Google Scholar] [CrossRef]
- Plaza, B.M.; Gómez-Serrano, C.; Acién-Fernández, F.G.; Jimenez-Becker, S. Effect of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp.) on Petunia x hybrida growth. J. Appl. Phycol. 2018, 30, 2359–2365. [Google Scholar] [CrossRef]
- Navarro-López, E.; Ruíz-Nieto, A.; Ferreira, A.; Gabriel Acién, F.; Gouveia, L. Biostimulant Potential of Scenedesmus obliquus Grown in Brewery Wastewater. Molecules 2020, 25, 664. [Google Scholar] [CrossRef] [Green Version]
- Niemann, D.I.; Dörffling, K. Growth-inhibitors and growth-promoters in enteromorpha a compress (CHLOROPHYTA). J. Phycol. 1980, 16, 383–389. [Google Scholar] [CrossRef]
- Lafarga, T. Cultured Microalgae and Compounds Derived Thereof for Food Applications: Strain Selection and Cultivation, Drying, and Processing Strategies. Food Rev. Int. 2020, 36, 559–583. [Google Scholar] [CrossRef]
- Supraja, K.V.; Behera, B.; Balasubramanian, P. Efficacy of microalgal extracts as biostimulants through seed treatment and foliar spray for tomato cultivation. Ind. Crops Prod. 2020, 151, 112453. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morillas-España, A.; Ruiz-Nieto, Á.; Lafarga, T.; Acién, G.; Arbib, Z.; González-López, C.V. Biostimulant Capacity of Chlorella and Chlamydopodium Species Produced Using Wastewater and Centrate. Biology 2022, 11, 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11071086
Morillas-España A, Ruiz-Nieto Á, Lafarga T, Acién G, Arbib Z, González-López CV. Biostimulant Capacity of Chlorella and Chlamydopodium Species Produced Using Wastewater and Centrate. Biology. 2022; 11(7):1086. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11071086
Chicago/Turabian StyleMorillas-España, Ainoa, Ángela Ruiz-Nieto, Tomás Lafarga, Gabriel Acién, Zouhayr Arbib, and Cynthia V. González-López. 2022. "Biostimulant Capacity of Chlorella and Chlamydopodium Species Produced Using Wastewater and Centrate" Biology 11, no. 7: 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11071086