


Usefulness of Surface Electromyography Complexity Analyses to Assess the Effects of Warm-Up and Stretching during Maximal and Sub-Maximal Hamstring Contractions: A Cross-Over, Randomized, Single-Blind Trial



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
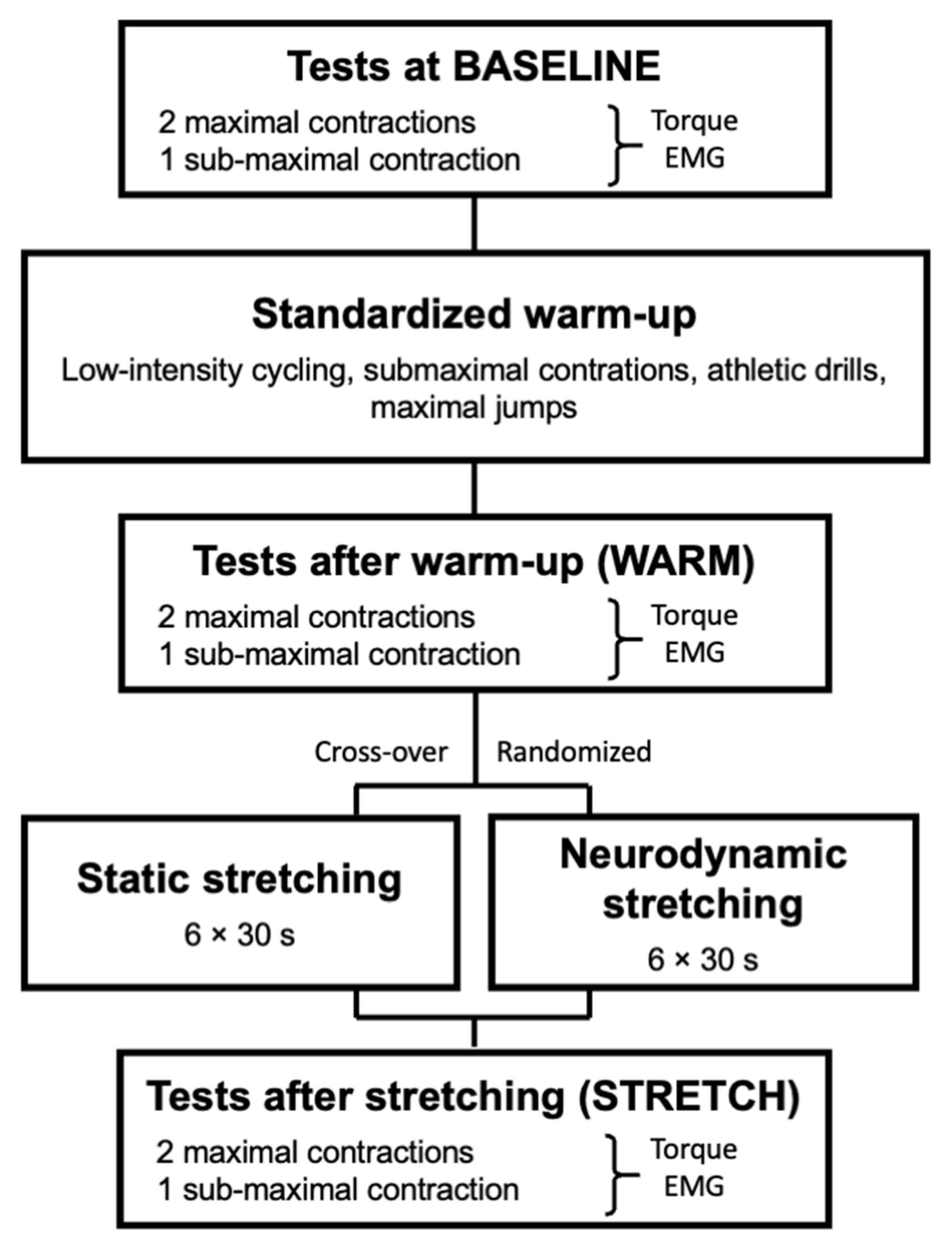
2.1. Study Design
2.2. Participants
2.3. Experimental Sessions
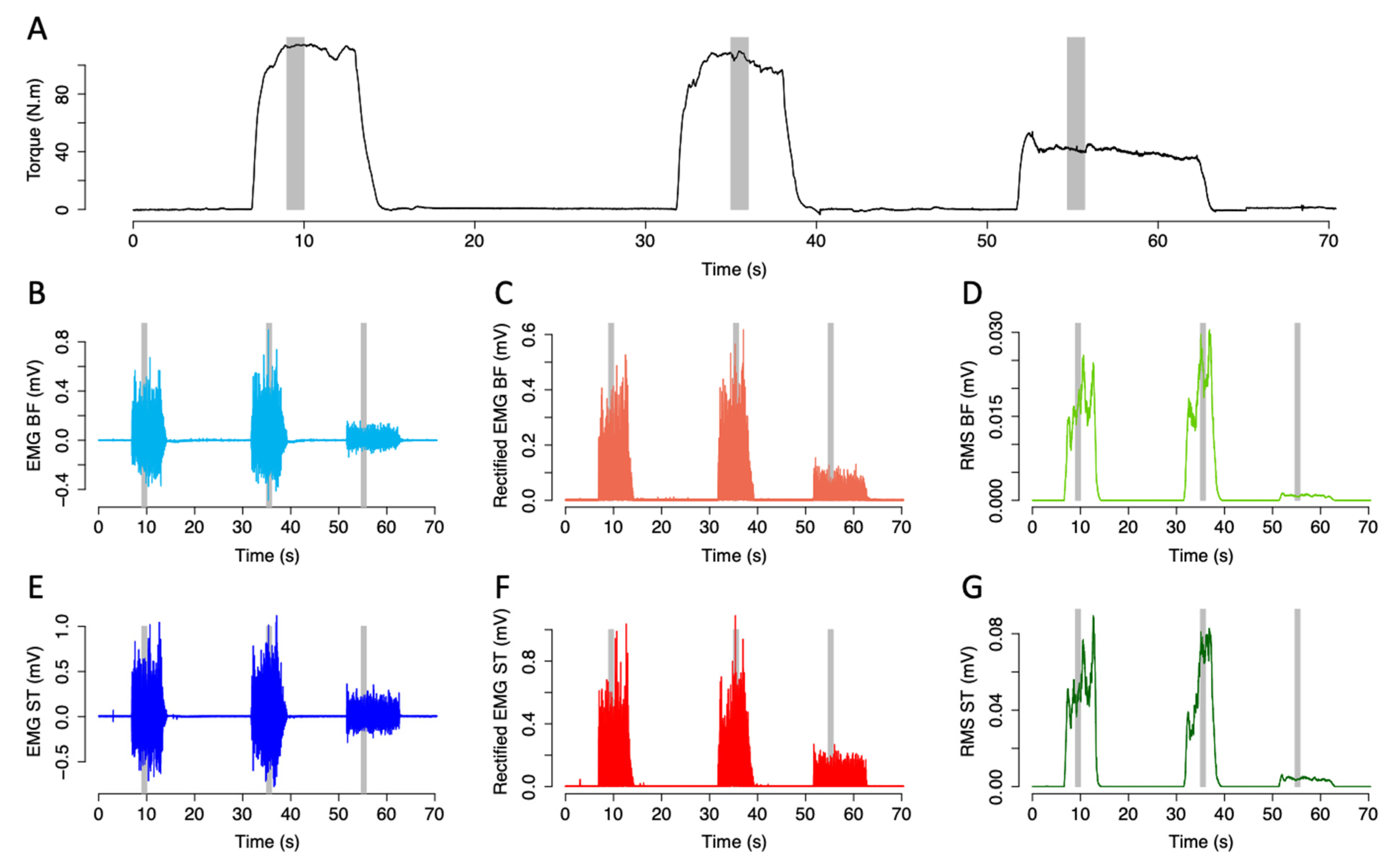
2.4. Data Recording, Preprocessing and Initial Analyses
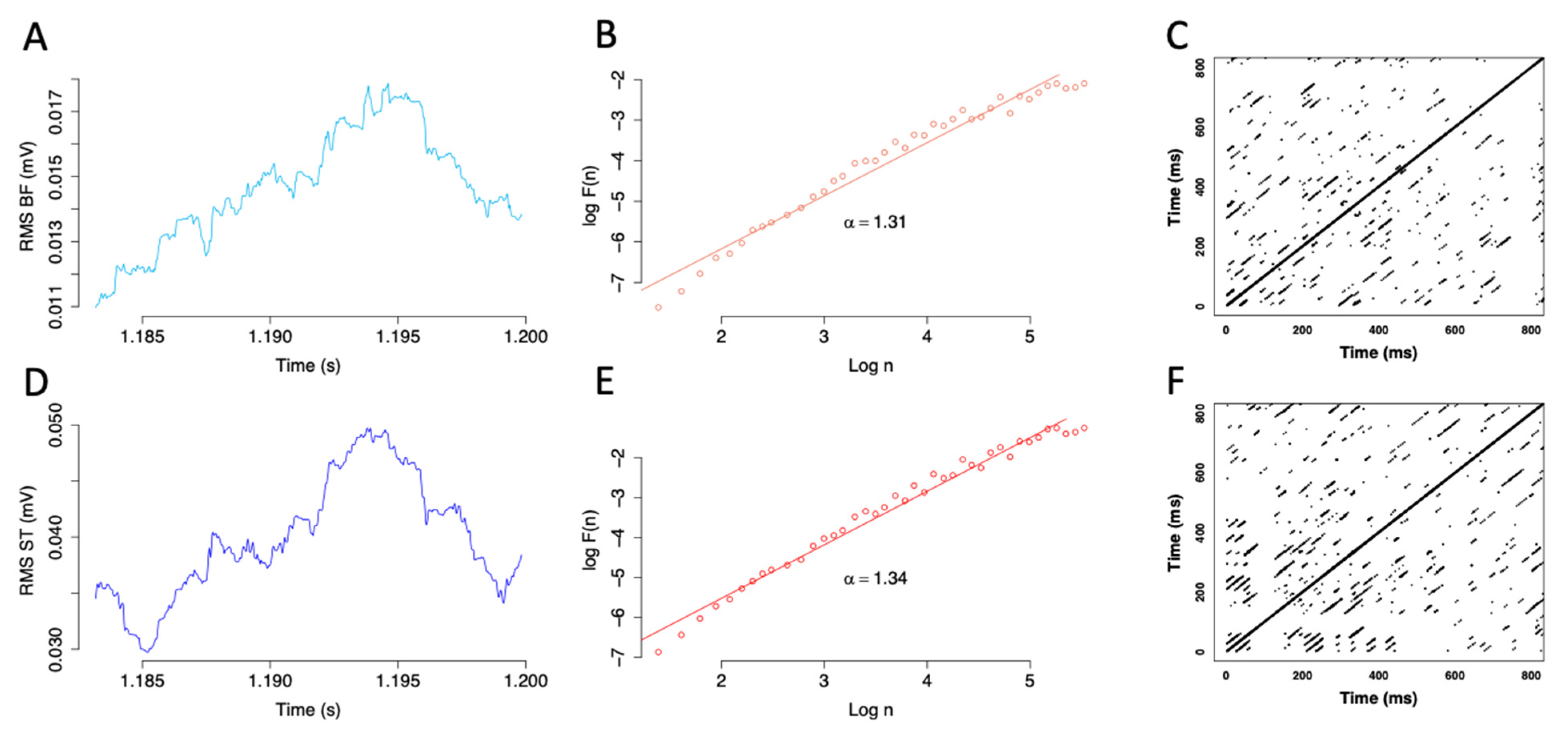
2.5. Sample Entropy
2.6. Recurrence Quantification Analysis
2.7. Detrended Fluctuation Analysis
2.8. Statistical Analyses
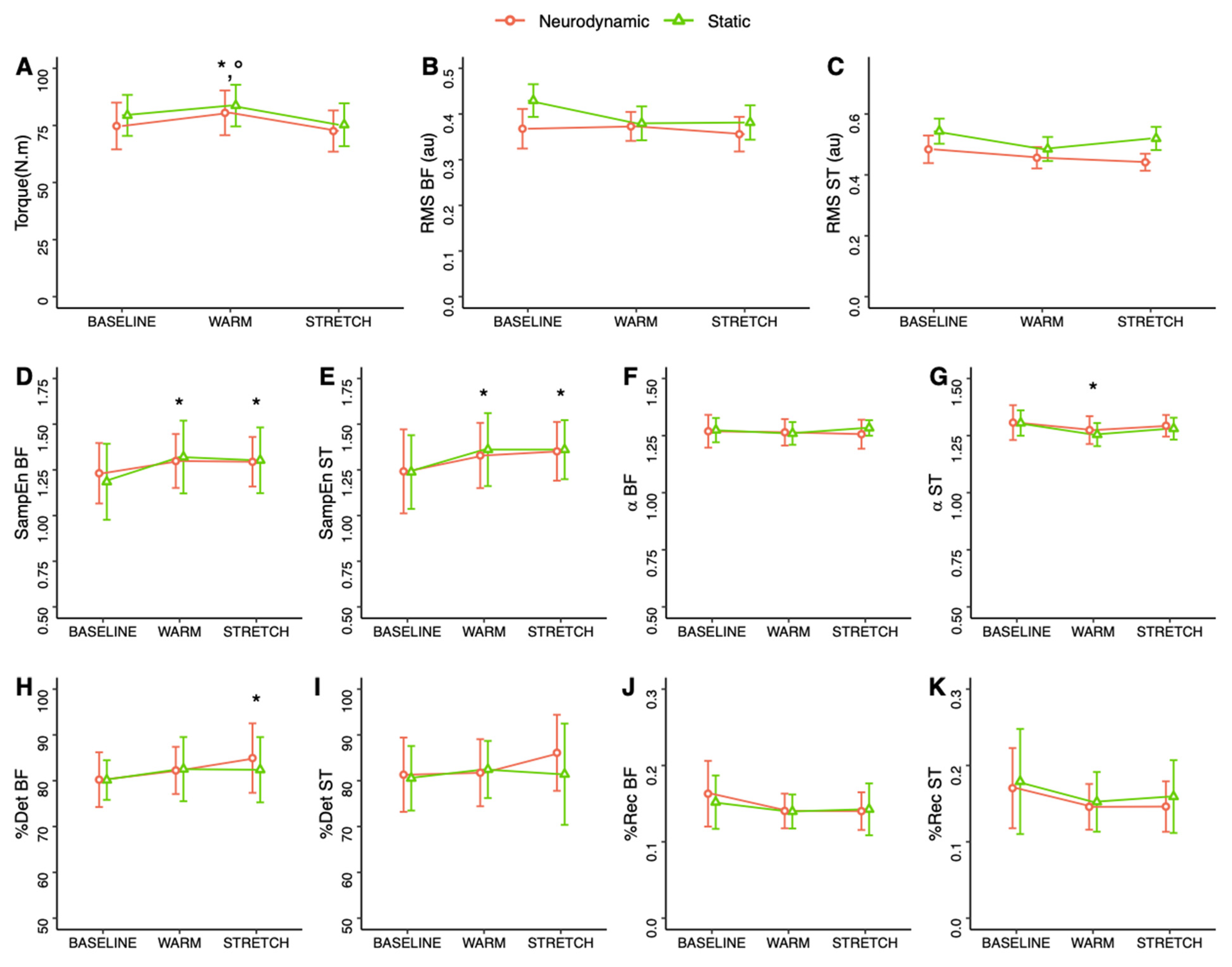
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGavin, R.J. Effect of Different Warm-Up Exercises of Varying Intensities on Speed of Leg Movement. Res. Quarterly. Am. Assoc. Health Phys. Educ. Recreat. 1968, 39, 125–130. [Google Scholar] [CrossRef]
- Mead, W.F.; Hartwig, R. Fitness Evaluation and Exercise Prescription. J. Fam. Prac. 1981, 13, 1039–1050. [Google Scholar]
- Bishop, D. Warm Up I: Potential Mechanisms and the Effects of Passive Warm Up on Exercise Performance. Sports Med. 2003, 33, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Babault, N.; Maffiuletti, N.A.; Pousson, M. Postactivation Potentiation in Human Knee Extensors during Dynamic Passive Movements. Med. Sci. Sports Exerc. 2008, 40, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Babault, N.; Rodot, G.; Champelovier, M.; Cometti, C. A Survey on Stretching Practices in Women and Men from Various Sports or Physical Activity Programs. Int. J. Environ. Res. Public Health 2021, 18, 3928. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.C.; Koslow, R. Comparative Study of Static, Dynamic, and Proprioceptive Neuromuscular Facilitation Stretching Techniques on Flexibility. Percept. Mot. Ski. 1984, 58, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Behm, D.G.; Blazevich, A.J.; Kay, A.D.; McHugh, M. Acute Effects of Muscle Stretching on Physical Performance, Range of Motion, And Injury Incidence in Healthy Active Individuals: A Systematic Review. Appl. Physiol. Nutr. Metab. 2016, 41, 1–11. [Google Scholar] [CrossRef]
- Opplert, J.; Babault, N. Acute Effects of Dynamic Stretching on Muscle Flexibility and Performance: An Analysis of the Current Literature. Sports Med. 2017, 48, 299–325. [Google Scholar] [CrossRef]
- Avela, J.; Kyröläinen, H.; Komi, P.V. Altered Reflex Sensitivity After Repeated and Prolonged Passive Muscle Stretching. J. Appl. Physiol. 1999, 86, 1283–1291. [Google Scholar] [CrossRef]
- Reid, J.C.; Greene, R.; Young, J.D.; Hodgson, D.D.; Blazevich, A.J.; Behm, D.G. The Effects of Different Durations of Static Stretching Within a Comprehensive Warm-Up on Voluntary and Evoked Contractile Properties. Eur. J. Appl. Physiol. 2018, 118, 1427–1445. [Google Scholar] [CrossRef]
- Samson, M.; Button, D.C.; Chaouachi, A.; Behm, D.G. Effects of Dynamic and Static Stretching Within General and Activity Specific Warm-Up Protocols. J. Sports Sci. Med. 2012, 11, 279–285. [Google Scholar] [PubMed]
- Winchester, J.B.; Nelson, A.G.; Kokkonen, J. A Single 30-s Stretch Is Sufficient to Inhibit Maximal Voluntary Strength. Res. Q. Exerc. Sport 2009, 80, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Fortier, J.; Lattier, G.; Babault, N. Acute Effects of Short-Duration Isolated Static Stretching or Combined with Dynamic Exercises on Strength, Jump and Sprint Performance. Sci. Sports 2013, 28, e111–e117. [Google Scholar] [CrossRef]
- Opplert, J.; Paizis, C.; Papitsa, A.; Blazevich, A.J.; Cometti, C.; Babault, N. Static Stretch and Dynamic Muscle Activity Induce Acute Similar Increase in Corticospinal Excitability. PLoS ONE 2020, 15, e0230388. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.F.; Hing, W. Neural Mobilization: A Systematic Review of Randomized Controlled Trials with an Analysis of Therapeutic Efficacy. J. Man. Manip. Ther. 2008, 16, 8–22. [Google Scholar] [CrossRef]
- Satkunskiene, D.; Khair, R.M.; Muanjai, P.; Mickevicius, M.; Kamandulis, S. Immediate Effects of Neurodynamic Nerve Gliding Versus Static Stretching on Hamstring Neuromechanical Properties. Eur. J. Appl. Physiol. 2020, 120, 2127–2135. [Google Scholar] [CrossRef]
- Satkunskiene, D.; Ardekani, M.M.Z.; Khair, R.M.; Kutraite, G.; Venckuniene, K.; Snieckus, A.; Kamandulis, S. Warm-Up and Hamstrings Stiffness, Stress Relaxation, Flexibility, and Knee Proprioception in Young Soccer Players. J. Athl. Train. 2022, 57, 485–493. [Google Scholar] [CrossRef]
- Andrade, R.J.; Freitas, S.R.; Hug, F.; Le Sant, G.; Lacourpaille, L.; Gross, R.; Quillard, J.-B.; McNair, P.J.; Nordez, A. Chronic Effects of Muscle and Nerve-Directed Stretching on Tissue Mechanics. J. Appl. Physiol. 2020, 129, 1011–1023. [Google Scholar] [CrossRef]
- Castellote-Caballero, Y.; Valenza, M.C.; Martín-Martín, L.; Cabrera-Martos, I.; Puentedura, E.J.; Fernández-De-Las-Peñas, C. Effects of a Neurodynamic Sliding Technique on Hamstring Flexibility in Healthy Male Soccer Players. A Pilot Studies. Phys. Ther. Sport 2013, 14, 156–162. [Google Scholar] [CrossRef]
- Fasen, J.M.; O’Connor, A.M.; Schwartz, S.L.; Watson, J.O.; Plastaras, C.T.; Garvan, C.W.; Bulcao, C.; Johnson, S.C.; Akuthota, V. A Randomized Controlled Trial of Hamstring Stretching: Comparison of Four Techniques. J. Strength Cond. Res. 2009, 23, 660–667. [Google Scholar] [CrossRef]
- Behm, D.G.; Button, D.C.; Butt, J.C. Factors Affecting Force Loss with Prolonged Stretching. Can. J. Appl. Physiol. 2001, 26, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Fowles, J.R.; Sale, D.G.; MacDougall, J.D. Reduced Strength After Passive Stretch of the Human plantarflexors. J. Appl. Physiol. 2000, 89, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Herda, T.J.; Cramer, J.; Ryan, E.D.; McHugh, M.P.; Stout, J.R. Acute Effects of Static versus Dynamic Stretching on Isometric Peak Torque, Electromyography, and Mechanomyography of the Biceps Femoris Muscle. J. Strength Cond. Res. 2008, 22, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.J.; Petterson, A.; Cochrane, D.J. Effects of Different Warm-Up Modalities on Power Output During the High Pull. J. Sports Sci. 2016, 35, 976–981. [Google Scholar] [CrossRef]
- Altamirano, K.M.; Coburn, J.W.; Brown, L.E.; Judelson, D.A. Effects of Warm-up on Peak Torque, Rate of Torque Development, and Electromyographic and Mechanomyographic Signals. J. Strength Cond. Res. 2012, 26, 1296–1301. [Google Scholar] [CrossRef] [PubMed]
- Parr, M.; Price, P.D.; Cleather, D.J. Effect of a Gluteal Activation Warm-Up on Explosive Exercise Performance. BMJ Open Sport Exerc. Med. 2017, 3, e000245. [Google Scholar] [CrossRef]
- De Luca, C.J. Physiology and Mathematics of Myoelectric Signals. IEEE Trans. Biomed. Eng. 1979, BME-26, 313–325. [Google Scholar] [CrossRef]
- Rampichini, S.; Vieira, T.M.; Castiglioni, P.; Merati, G. Complexity Analysis of Surface Electromyography for Assessing the Myoelectric Manifestation of Muscle Fatigue: A Review. Entropy 2020, 22, 529. [Google Scholar] [CrossRef]
- Chakraborty, M.; Parbat, D. Fractals, Chaos and Entropy Analysis to Obtain Parametric Features of Surface Electromyography Signals During Dynamic Contraction of Biceps Muscles Under Varying Load. In Proceedings of the 2017 2nd International Conference for Convergence in Technology (I2CT), Mumbai, India, 7–9 April 2017; Institute of Electrical and Electronics Engineers Inc.: Piscataway, NJ, USA, 2017; Volume 2017, pp. 222–229. [Google Scholar]
- Hernandez, L.R.; Camic, C.L. Fatigue-Mediated Loss of Complexity is Contraction-Type Dependent in Vastus Lateralis Electromyographic Signals. Sports 2019, 7, 78. [Google Scholar] [CrossRef]
- Santuz, A.; Ekizos, A.; Kunimasa, Y.; Kijima, K.; Ishikawa, M.; Arampatzis, A. Lower Complexity of Motor Primitives Ensures Robust Control of High-Speed Human Locomotion. Heliyon 2020, 6, e05377. [Google Scholar] [CrossRef]
- Riccetti, M.; Opplert, J.; Durigan, J.L.Q.; Cometti, C.; Babault, N. Acute Static Stretching Results in Muscle-Specific Alterations amongst the Hamstring Muscles. Sports 2020, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.A.; Macias-Diaz, J.E. BiosignalEMG: Tools for Electromyogram Signals (EMG) analysis. Available online: https://cran.r-project.org/web/packages/biosignalEMG/ (accessed on 19 July 2022).
- Richman, J.S.; Moorman, J.R. Physiological Time-Series Analysis Using Approximate Entropy and Sample Entropy. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H2039–H2049. [Google Scholar] [CrossRef]
- Tomcala, J. TSEntropies: Time Series Entropies. R package. 2018. Available online: https://cran.r-project.org/package=TSEntropies (accessed on 19 July 2022).
- Eckmann, J.-P.; Kamphorst, S.O.; Ruelle, D. Recurrence Plots of Dynamical Systems. Eur. Lett. 1987, 4, 973–977. [Google Scholar] [CrossRef]
- Morana, C.; Ramdani, S.; Perrey, S.; Varray, A. Recurrence Quantification Analysis of Surface Electromyographic Signal: Sensitivity to Potentiation and Neuromuscular Fatigue. J. Neurosci. Methods 2009, 177, 73–79. [Google Scholar] [CrossRef]
- Farina, D.; Fattorini, L.; Felici, F.; Filligoi, G. Nonlinear Surface EMG Analysis to Detect Changes of Motor Unit Conduction Velocity and Synchronization. J. Appl. Physiol. 2002, 93, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.; Zhu, X.; Ju, Z.; Liu, H. Dynamical Characteristics of Surface EMG Signals of Hand Grasps via Recurrence Plot. IEEE J. Biomed. Health Inform. 2013, 18, 257–265. [Google Scholar] [CrossRef]
- Marwan, N.; Carmenromano, M.; Thiel, M.; Kurths, J. Recurrence Plots for the Analysis of Complex Systems. Phys. Rep. 2007, 438, 237–329. [Google Scholar] [CrossRef]
- Webber, C.L.; Schmidt, M.A.; Walsh, J.M. Influence of Isometric Loading on Biceps EMG Dynamics As Assessed by Linear and Nonlinear Tools. J. Appl. Physiol. 1995, 78, 814–822. [Google Scholar] [CrossRef]
- Coco, M.; Monster, D.; Leonardi, G.; Dale, R.; Wallot, S.; Dixon, J.; Nash, J. crqa: Recurrence Quantification Analysis for Categorical and Continuous Times-Series. Available online: https://cran.r-project.org/package=crqa (accessed on 19 July 2022).
- Peng, C.-K.; Havlin, S.; Stanley, H.E.; Goldberger, A.L. Quantification of Scaling Exponents and Crossover Phenomena in Nonstationary Heartbeat Time Series. CHAOS 1995, 5, 82–87. [Google Scholar] [CrossRef]
- Mesquita, V.; Rodrigues, P.; Filho, F. DFA: Detrended Fluctuation Analysis. Available online: https://cran.r-project.org/package=DFA (accessed on 21 July 2022).
- Cohen, J. Statistical Power Analysis for The Behavioral Sciences; Routledge: Londan, UK, 2013. [Google Scholar]
- Đurović, M.; Stojanović, N.; Stojiljković, N.; Karaula, D.; Okičić, T. The Effects of Post-Activation Performance Enhancement and Different Warm-Up Protocols on Swim Start Performance. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Wu, C.-C.; Wang, M.-H.; Chang, C.-Y.; Hung, M.-H.; Wang, H.-H.; Chen, K.-C.; Ger, T.-R.; Lin, K.-C. The Acute Effects of Whole-Body Vibration Stimulus Warm-Up on Skill-Related Physical Capabilities in Volleyball Players. Sci. Rep. 2021, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Merino-Marban, R.; Fuentes, V.; Torres, M.; Mayorga-Vega, D. Acute Effect of a Static- And Dynamic-Based Stretching Warm-Up on Standing Long Jump Performance in Primary Schoolchildren. Biol. Sport 2021, 38, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Zagatto, A.M.; Dutra, Y.M.; Claus, G.; Malta, E.D.S.; de Poli, R.A.B.; Brisola, G.M.P.; Boullosa, D. Drop Jumps Improve Repeated Sprint Ability Performances in Professional Basketball Players. Biol. Sport 2022, 39, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Pearce, A.J.; Rowe, G.S.; Whyte, D.G. Neural Conduction and Excitability Following a Simple Warm up. J. Sci. Med. Sport 2012, 15, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Van Der Hoeven, J.H.; Lange, F. Supernormal Muscle Fiber Conduction Velocity During Intermittent Isometric Exercise in Human Muscle. J. Appl. Physiol. 1994, 77, 802–806. [Google Scholar] [CrossRef]
- Nazaroff, B.M.; Pearcey, G.E.P.; Munro, B.; Zehr, E.P. Sensory Enhancement of Warm-Up Amplifies Subsequent Grip Strength and Cycling Performance. Eur. J. Appl. Physiol. 2022, 122, 1695–1707. [Google Scholar] [CrossRef]
- Stevanovic, V.B.; Jelic, M.B.; Milanovic, S.D.; Filipovic, S.R.; Mikic, M.J.; Stojanovic, M.D.M. Sport-Specific Warm-Up Attenuates Static Stretching- Induced Negative Effects on Vertical Jump but Not Neuromuscular Excitability in Basketball Players. J. Sports Sci. Med. 2019, 18, 282–289. [Google Scholar]
- Palmer, T.B.; Pineda, J.G.; Cruz, M.R.; Agu-Udemba, C.C. Duration-Dependent Effects of Passive Static Stretching on Musculotendinous Stiffness and Maximal and Rapid Torque and Surface Electromyography Characteristics of the Hamstrings. J. Strength Cond. Res. 2019, 33, 717–726. [Google Scholar] [CrossRef]
- Blazevich, A.; Babault, N. Post-activation Potentiation Versus Post-Activation Performance Enhancement in Humans: Historical Perspective, Underlying Mechanisms, and Current Issues. Front. Physiol. 2019, 10, 1359. [Google Scholar] [CrossRef]
- Pethick, J.; Winter, S.; Burnley, M. Effects of Ipsilateral and Contralateral Fatigue and Muscle Blood Flow Occlusion on the Complexity of Knee-Extensor Torque Output in Humans. Exp. Physiol. 2018, 103, 956–967. [Google Scholar] [CrossRef]
- Hardstone, R.; Poil, S.-S.; Schiavone, G.; Jansen, R.; Nikulin, V.V.; Mansvelder, H.D.; Linkenkaer-Hansen, K. Detrended Fluctuation Analysis: A Scale-Free View on Neuronal Oscillations. Front. Physiol. 2012, 3, 450. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, Z.-Z.; Ren, X.-M. Classification of Surface EMG Signal with Fractal Dimension. J. Zhejiang Univ. A 2005, 6B, 844–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, N.; Komiya, M.; Nishikawa, Y.; Morikawa, M.; Tsutsumi, S.; Tashiro, T.; Fukui, K.; Kimura, H.; Urabe, Y. Effect of Acute Static Stretching on the Activation Patterns Using High-Density Surface Electromyography of the Gastrocnemius Muscle during Ramp-Up Task. Sensors 2021, 21, 4841. [Google Scholar] [CrossRef] [PubMed]
- Goldberger, A.L.; Peng, C.K.; Lipsitz, L.A. What is Physiologic Complexity and How Does It Change with Aging and Disease? Neurobiol. Aging 2002, 23, 23–26. [Google Scholar] [CrossRef]
- Zhou, J.; Manor, B.; Liu, D.; Hu, K.; Zhang, J.; Fang, J. The Complexity of Standing Postural Control in Older Adults: A Modified Detrended Fluctuation Analysis Based upon the Empirical Mode Decomposition Algorithm. PLoS ONE 2013, 8, e62585. [Google Scholar] [CrossRef]
- Kahl, L.; Hofmann, U. Comparison of Algorithms to Quantify Muscle Fatigue in Upper Limb Muscles Based on sEMG Signals. Med. Eng. Phys. 2016, 38, 1260–1269. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Effect | Maximal Contraction | Sub-Maximal Contraction | ||||
|---|---|---|---|---|---|---|---|
| F | p | ph2 | F | p | ph2 | ||
| Condition | 1.363 | 0.261 | 0.083 | 0.620 | 0.443 | 0.040 | |
| Torque | Time | 11.942 | < 0.001 * | 0.443 | 2.443 | 0.104 | 0.140 |
| Condition x Time | 0.268 | 0.767 | 0.018 | 0.548 | 0.532 | 0.035 | |
| Condition | 1.345 | 0.264 | 0.082 | 1.194 | 0.292 | 0.074 | |
| RMS BF | Time | 0.956 | 0.396 | 0.060 | 2.352 | 0.113 | 0.136 |
| Condition x Time | 0.427 | 0.656 | 0.028 | 1.155 | 0.329 | 0.071 | |
| Condition | 4.650 | 0.048 * | 0.237 | 0.722 | 0.409 | 0.046 | |
| RMS ST | Time | 3.320 | 0.050 * | 0.181 | 2.258 | 0.122 | 0.131 |
| Condition x Time | 0.291 | 0.750 | 0.019 | 1.866 | 0.172 | 0.111 | |
| Condition | 0.031 | 0.863 | 0.002 | 3.865 | 0.068 | 0.205 | |
| SampEn BF | Time | 10.463 | < 0.001 * | 0.411 | 0.097 | 0.907 | 0.006 |
| Condition x Time | 1.794 | 0.184 | 0.107 | 0.854 | 0.436 | 0.054 | |
| Condition | 0.136 | 0.717 | 0.009 | 1.812 | 0.198 | 0.108 | |
| SampEn ST | Time | 19.175 | < 0.001 * | 0.561 | 0.796 | 0.461 | 0.050 |
| Condition x Time | 0.559 | 0.578 | 0.036 | 0.909 | 0.414 | 0.057 | |
| Condition | 1.041 | 0.325 | 0.069 | 0.994 | 0.335 | 0.062 | |
| %Rec BF | Time | 0.952 | 0.346 | 0.064 | 0.930 | 0.350 | 0.058 |
| Condition x Time | 0.965 | 0.343 | 0.064 | 1.046 | 0.323 | 0.065 | |
| Condition | 0.881 | 0.364 | 0.059 | 0.225 | 0.642 | 0.015 | |
| %Rec ST | Time | 0.944 | 0.348 | 0.063 | 0.605 | 0.482 | 0.039 |
| Condition x Time | 0.966 | 0.343 | 0.065 | 1.453 | 0.251 | 0.088 | |
| Condition | 0.837 | 0.375 | 0.053 | 0.416 | 0.529 | 0.027 | |
| %Det BF | Time | 4.638 | 0.018 * | 0.236 | 2.259 | 0.122 | 0.131 |
| Condition x Time | 0.549 | 0.583 | 0.035 | 1.954 | 0.159 | 0.115 | |
| Condition | 1.496 | 0.240 | 0.091 | 2.098 | 0.168 | 0.123 | |
| %Det ST | Time | 1.101 | 0.330 | 0.068 | 1.949 | 0.160 | 0.115 |
| Condition x Time | 1.528 | 0.233 | 0.092 | 0.027 | 0.973 | 0.002 | |
| Condition | 0.557 | 0.467 | 0.036 | 0.096 | 0.761 | 0.006 | |
| a BF | Time | 0.285 | 0.754 | 0.019 | 0.232 | 0.794 | 0.015 |
| Condition x Time | 1.378 | 0.268 | 0.084 | 3.642 | 0.055 | 0.195 | |
| Condition | 0.709 | 0.413 | 0.045 | 0.013 | 0.911 | 0.001 | |
| a ST | Time | 7.623 | 0.002 * | 0.337 | 11.367 | < 0.001 * | 0.431 |
| Condition x Time | 0.394 | 0.615 | 0.026 | 1.118 | 0.340 | 0.069 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babault, N.; Hitier, M.; Cometti, C. Usefulness of Surface Electromyography Complexity Analyses to Assess the Effects of Warm-Up and Stretching during Maximal and Sub-Maximal Hamstring Contractions: A Cross-Over, Randomized, Single-Blind Trial. Biology 2022, 11, 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11091337
Babault N, Hitier M, Cometti C. Usefulness of Surface Electromyography Complexity Analyses to Assess the Effects of Warm-Up and Stretching during Maximal and Sub-Maximal Hamstring Contractions: A Cross-Over, Randomized, Single-Blind Trial. Biology. 2022; 11(9):1337. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11091337
Chicago/Turabian StyleBabault, Nicolas, Marion Hitier, and Carole Cometti. 2022. "Usefulness of Surface Electromyography Complexity Analyses to Assess the Effects of Warm-Up and Stretching during Maximal and Sub-Maximal Hamstring Contractions: A Cross-Over, Randomized, Single-Blind Trial" Biology 11, no. 9: 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11091337