
New Bacillus subtilis Strains Isolated from Prosopis glandulosa Rhizosphere for Suppressing Fusarium Spp. and Enhancing Growth of Gossypium hirsutum L.
 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Antagonistic Bacillus spp. from Honey Mesquite Rhizosphere
2.2. Isolation of Pathogenic Fungi
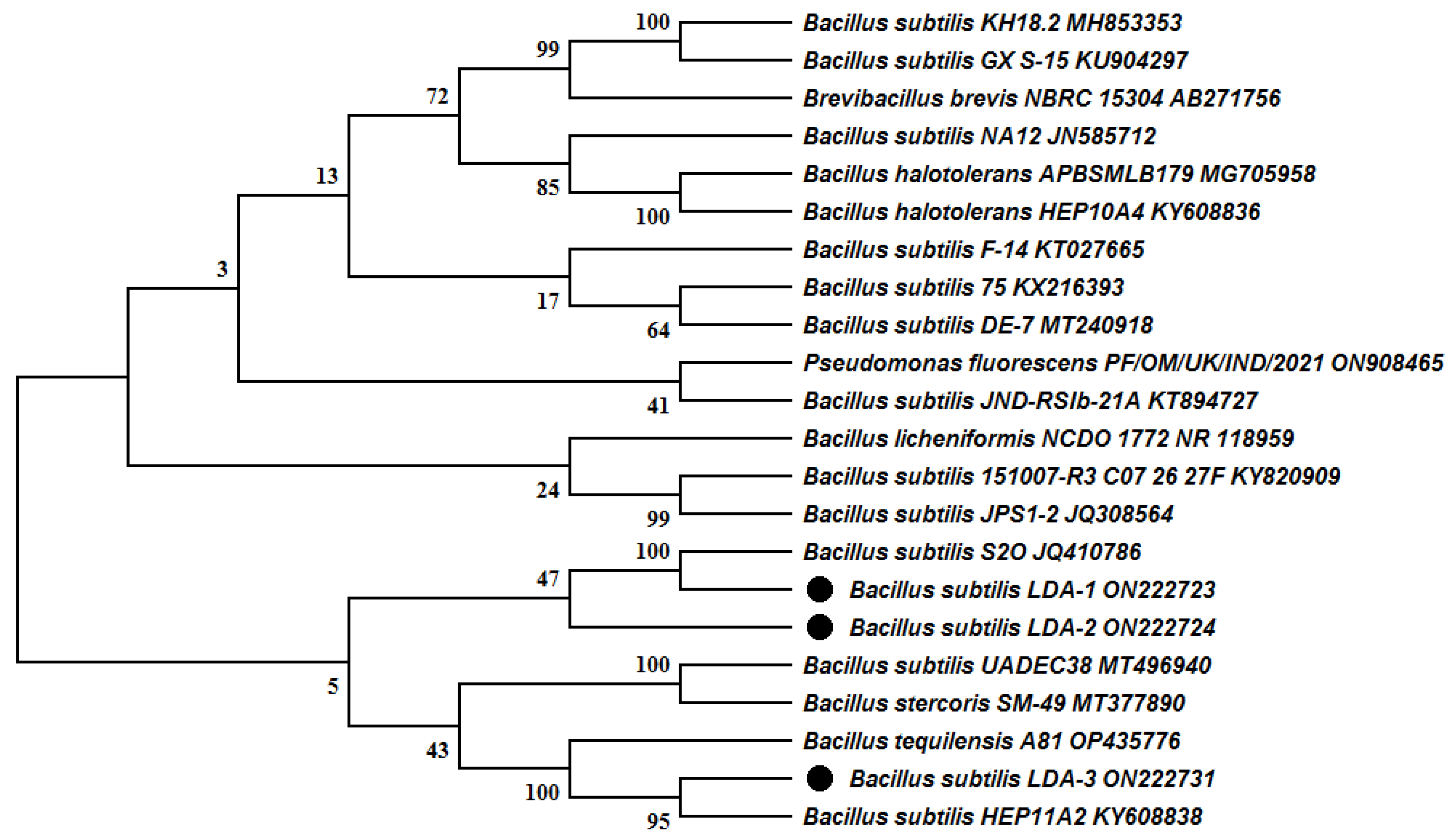
2.3. Molecular Identification of Bacterial Isolates
2.4. Amplification of Lipopeptide Genes
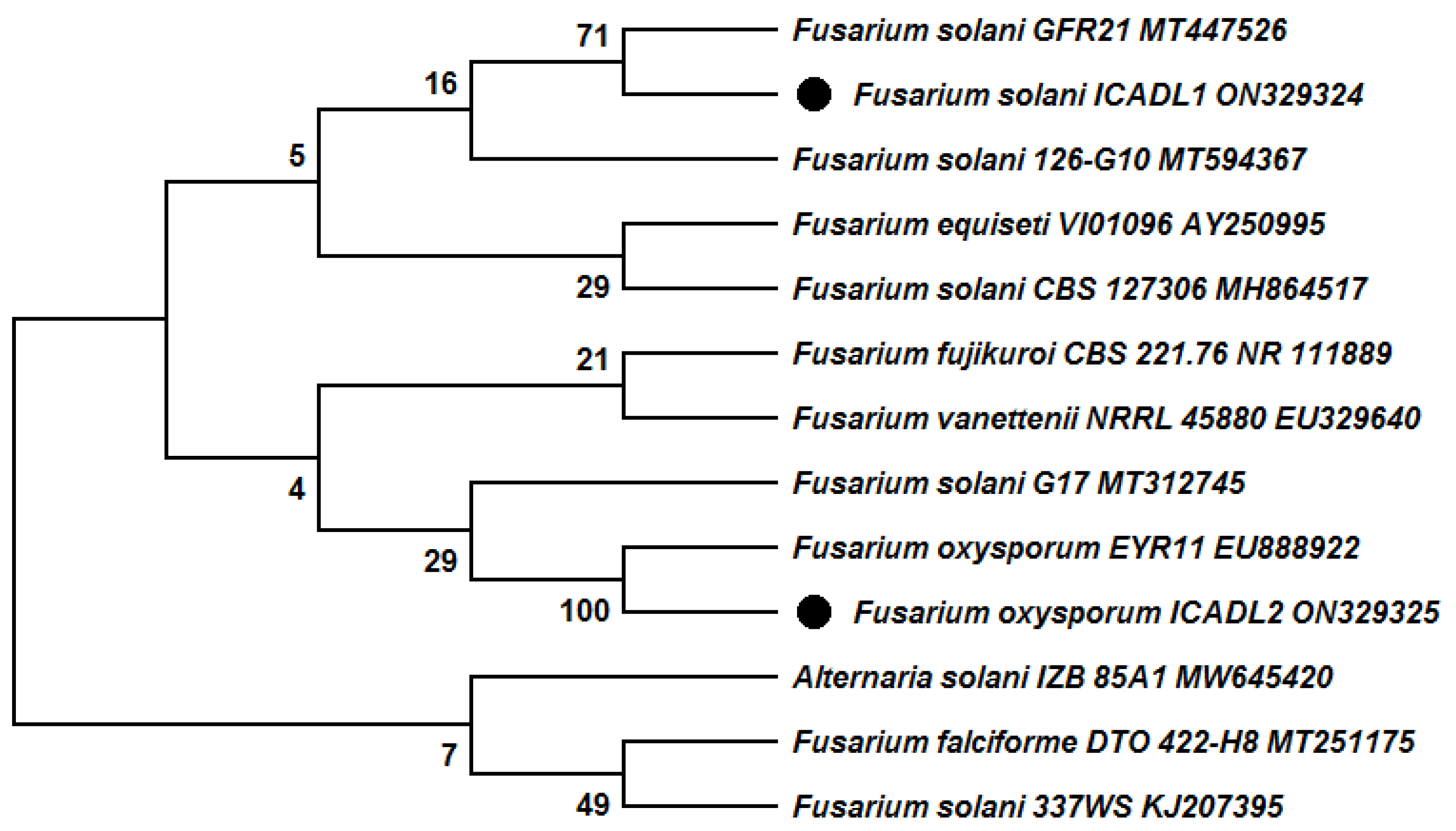
2.5. Molecular Identification of Pathogenic Fungi
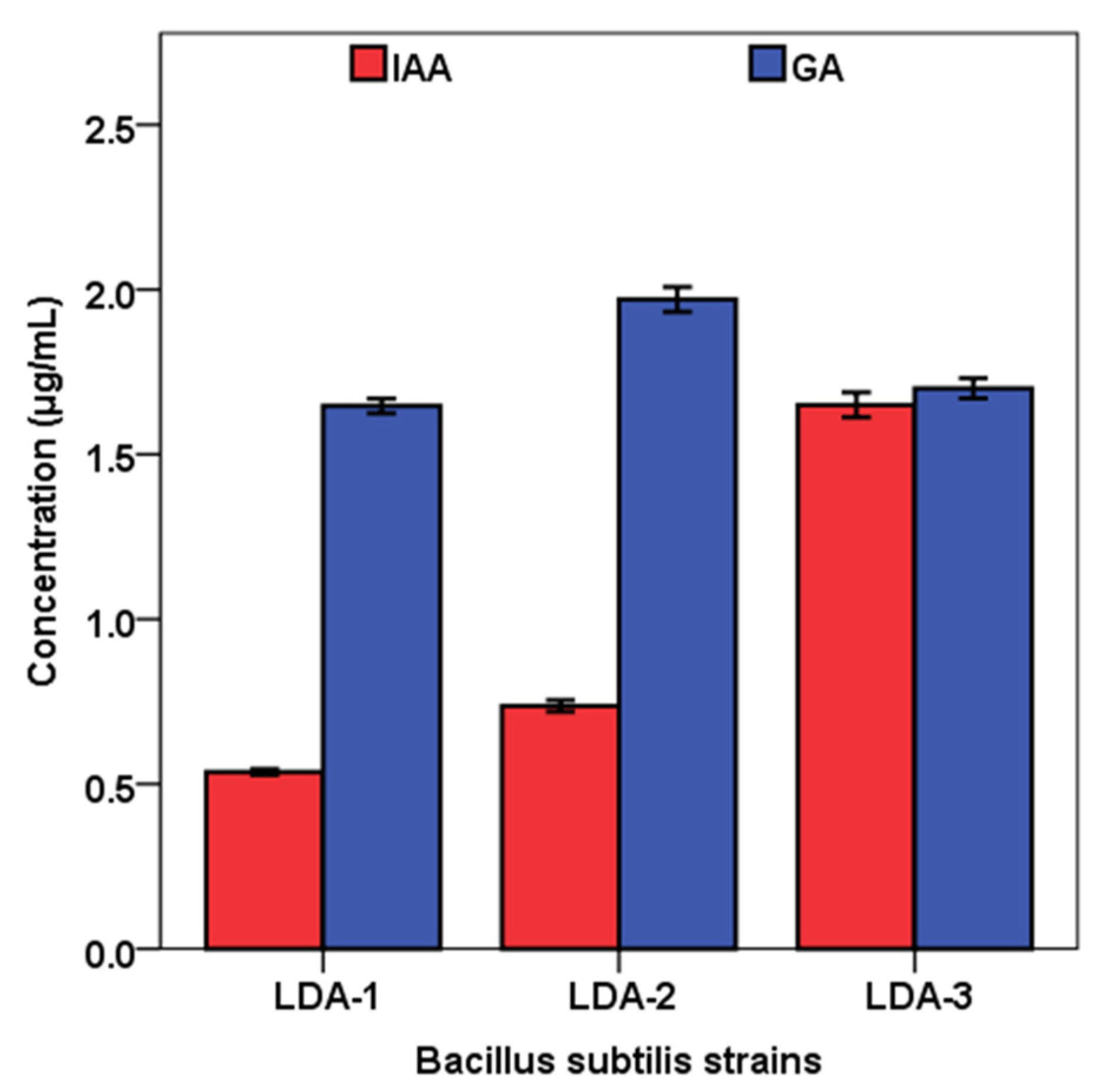
2.6. Indole Acetic Acid (IAA) Production by Bacterial Isolates
2.7. Gibberellic Acid (GA3) Production by Bacterial Isolates
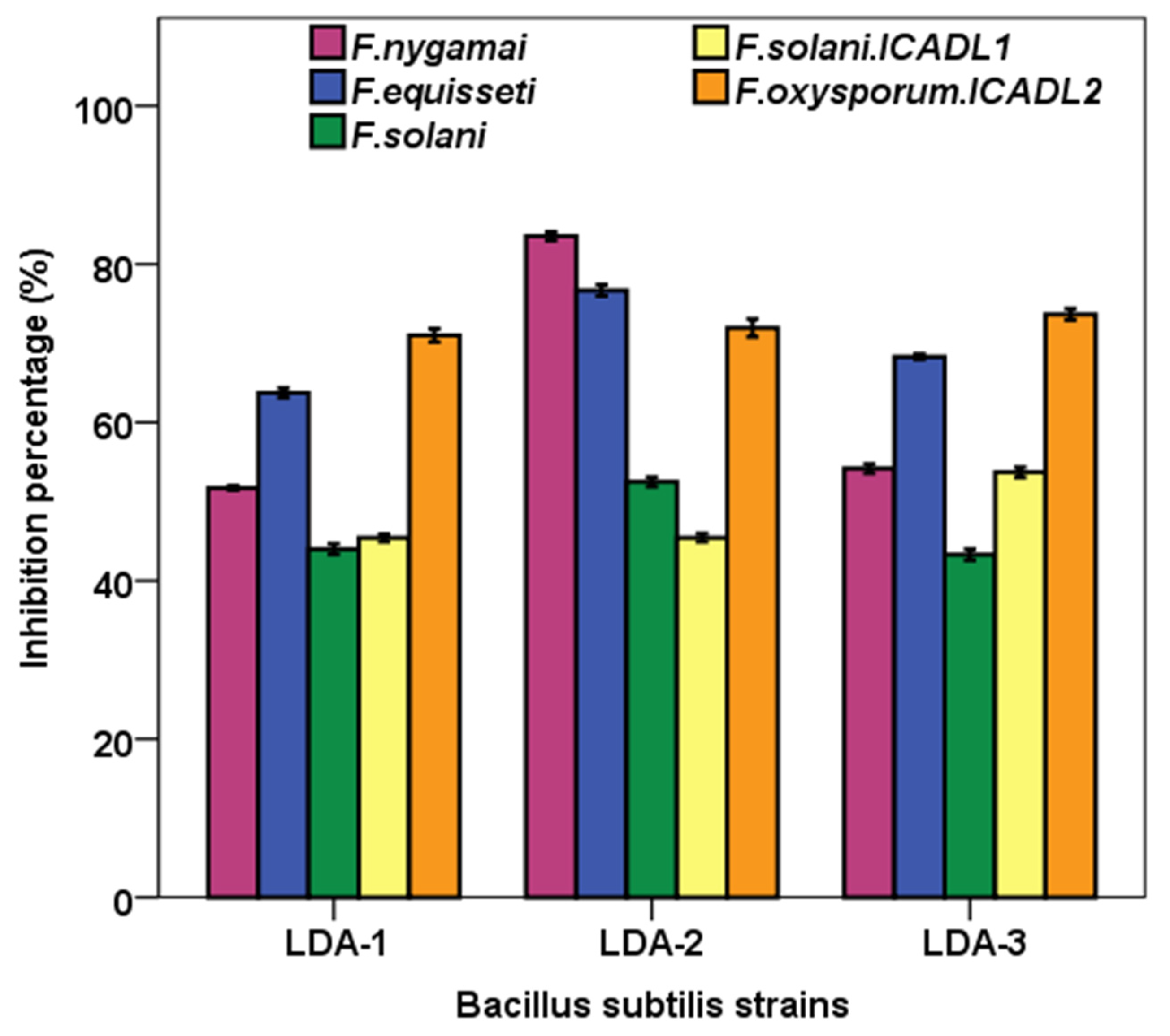
2.8. Antifungal Activity of Bacterial Isolates in Dual-Plate Assay
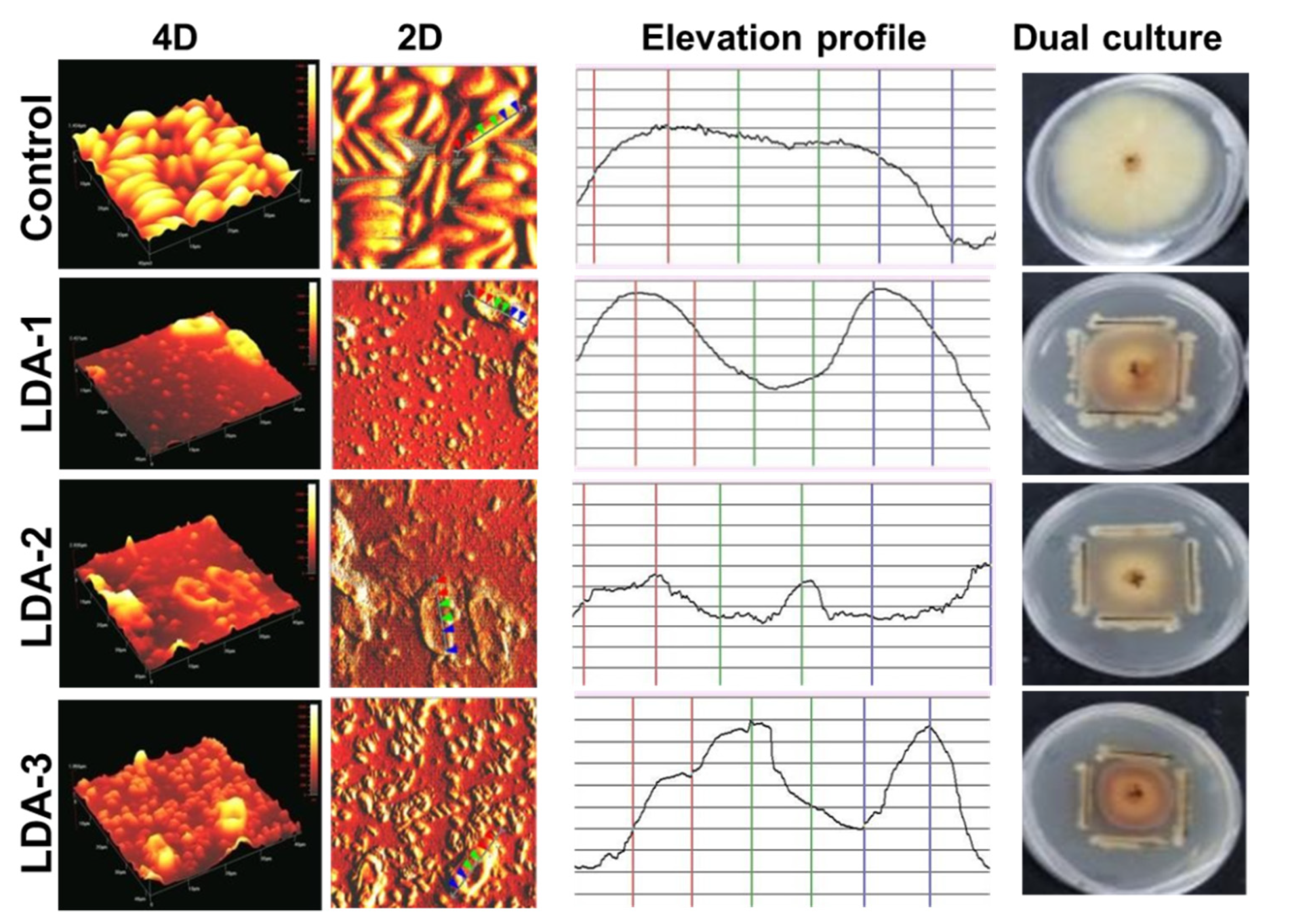
2.9. Morphological Analysis of Phytopathogen in Contact with Antagonistic Bacteria Using Atomic Force Microscopy (AFM)
2.10. Effect of Antagonistic Bacteria on Cotton Seedlings Exposed to Phytopathogen F. solani ICADL1
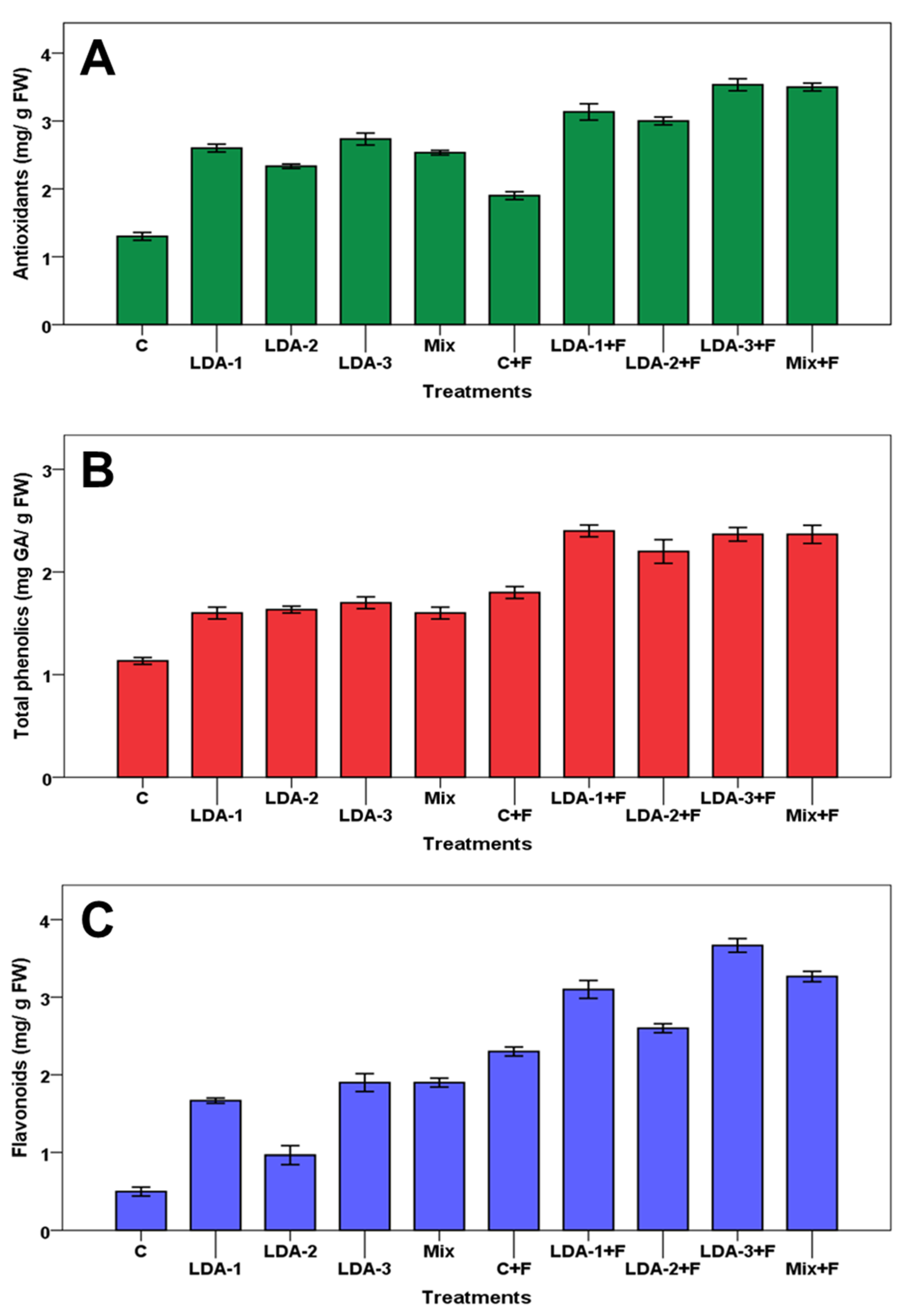
2.11. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Man, M.; Zhu, Y.; Liu, L.; Luo, L.; Han, X.; Qiu, L.; Li, F.; Ren, M.; Xing, Y. Defense Mechanisms of Cotton Fusarium and Verticillium Wilt and Comparison of Pathogenic Response in Cotton and Humans. Int. J. Mol. Sci. 2022, 23, 12217. [Google Scholar] [CrossRef] [PubMed]
- De Farias, O.R.; De Nascimento, L.C.; Lima Cruz, J.M.F.d.; Oliveira Silva, H.A.; Oliveira, M.D.d.M.; Alcântara Bruno, R.d.L.; Castro Arriel, N.H. Biocontrol Potential of Trichoderma and Bacillus Species on Fusarium oxysporum f. sp. vasinfectum. J. Exp. Agric. Int. 2019, 34, 1–11. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Gonzalez-Mendoza, D.; Valdez-Salas, B.; Grimaldo-Juarez, O.; Ceceña-Duran, C. Inhibition of Fusarium solani in Transgenic Insect-Resistant Cotton Plants Treated with Silver Nanoparticles from Prosopis glandulosa and Pluchea sericea. Egypt. J. Biol. Pest Control 2018, 28, 4. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elsalam, K.A.; Omar, M.R.; El-Samawaty, A.R.; Aly, A.A. Response of Commercial Cotton Cultivars to Fusarium solani. Plant Pathol. J. 2007, 23, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Martínez-Hidalgo, P.; Ice, T.A.; Maymon, M.; Humm, E.A.; Nejat, N.; Sanders, E.R.; Kaplan, D.; Hirsch, A.M. Antifungal Activity of Bacillus Species against Fusarium and Analysis of the Potential Mechanisms Used in Biocontrol. Front. Microbiol. 2018, 9, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.; M. El-Meihy, R. Phylogenetic Diversity of Trichoderma Strains and Their Antagonistic Potential against Soil-Borne Pathogens under Stress Conditions. Biology 2020, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Kim, T.Y.; Won, S.-J.; Moon, J.-H.; Ajuna, H.B.; Kim, K.Y.; Ahn, Y.S. Control of Fungal Diseases and Fruit Yield Improvement of Strawberry Using Bacillus velezensis CE 100. Microorganisms 2022, 10, 365. [Google Scholar] [CrossRef]
- Mardanova, A.M.; Hadieva, G.F.; Lutfullin, M.T.; Khilyas, I.V.; Minnullina, L.F.; Gilyazeva, A.G.; Bogomolnaya, L.M.; Sharipova, M.R. Bacillus subtilis Strains with Antifungal Activity against the Phytopathogenic Fungi. Agric. Sci. 2016, 8, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, H.F.A.; Seleiman, M.F.; Al-Saif, A.M.; Alshiekheid, M.A.; Battaglia, M.L.; Taha, R.S. Biological Control of Celery Powdery Mildew Disease Caused by Erysiphe heraclei DC In Vitro and In Vivo Conditions. Plants 2021, 10, 2342. [Google Scholar] [CrossRef]
- Li, E.; Li, Y.; Dai, X.; Yan, W.; Wang, G. Identification of Two Bacillus Strains with Antimicrobial Activity and Preliminary Evaluation of Their Biocontrol Efficiency. Horticulturae 2022, 8, 744. [Google Scholar] [CrossRef]
- Saravanan, R.; Nakkeeran, S.; Saranya, N.; Senthilraja, C.; Renukadevi, P.; Krishnamoorthy, A.S.; El Enshasy, H.A.; El-Adawi, H.; Malathi, V.G.; Salmen, S.H.; et al. Mining the Genome of Bacillus velezensis VB7 (CP047587) for MAMP Genes and Non-Ribosomal Peptide Synthetase Gene Clusters Conferring Antiviral and Antifungal Activity. Microorganisms 2021, 9, 2511. [Google Scholar] [CrossRef]
- Chen, L.; Heng, J.; Qin, S.; Bian, K. A Comprehensive Understanding of the Biocontrol Potential of Bacillus velezensis LM2303 against Fusarium Head Blight. PLoS ONE 2018, 13, e0198560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Quiros, C.; Okechukwu, E.C.; Hong, Y.; Baysal, Ö.; Tör, M. Comparison of Antifungal Activity of Bacillus Strains against Fusarium graminearum In Vitro and In Planta. Plants 2022, 11, 1999. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Radhakrishnan, R.; Kumar, A. Plant Defense Approach of Bacillus subtilis (BERA 71) against Macrophomina phaseolina (Tassi) Goid in Mung Bean. J. Plant Interact. 2017, 12, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Jamil, A. Antifungal and Plant Growth Promoting Activity of Trichoderma Spp. against Fusarium oxysporum f. sp. lycopersici Colonizing Tomato. J. Plant Prot. Res. 2021, 61, 243–253. [Google Scholar] [CrossRef]
- Akram, W.; Ahmad, A.; Yasin, N.A.; Anjum, T.; Ali, B.; Fatima, S.; Ahmed, S.; Simirgiotis, M.J.; Li, G. Mechanical Strengthening and Metabolic Re-Modulations Are Involved in Protection against Fusarium Wilt of Tomato by B. subtilis IAGS174. J. Plant Interact. 2021, 16, 411–421. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Troncoso-Rojas, R.; Gonzalez-Soto, T.; González-Mendoza, D. Antifungical Activity of Autochthonous Bacillus subtilis Isolated from Prosopis juliflora against Phytopathogenic Fungi. Mycobiology 2017, 45, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohil, R.B.; Raval, V.H.; Panchal, R.R.; Rajput, K.N. Plant Growth-Promoting Activity of Bacillus Sp. PG-8 Isolated From Fermented Panchagavya and Its Effect on the Growth of Arachis hypogea. Front. Agron. 2022, 4, 805454. [Google Scholar] [CrossRef]
- Yamamoto, S.; Shiraishi, S.; Suzuki, S. Are Cyclic Lipopeptides Produced by Bacillus amyloliquefaciens S13-3 Responsible for the Plant Defence Response in Strawberry against Colletotrichum gloeosporioides? Lett. Appl. Microbiol. 2015, 60, 379–386. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Valdez-Salas, B.; Ceceña-Duran, C.; Tzintzun-Camacho, O.; Gutiérrez-Miceli, F.; Grimaldo-Juarez, O.; González-Mendoza, D. Silver Nanoparticles from Prosopis glandulosa and Their Potential Application as Biocontrol of Acinetobacter calcoaceticus and Bacillus cereus. Chem. Speciat. Bioavailab. 2017, 29, 1–5. [Google Scholar] [CrossRef]
- Mendez-Trujillo, V.; Moreno-Ramírez, L.; Carrillo-Beltran, M.; González-Mendoza, D. Fast Protocol for DNA Isolation of DNA from Bacterial Isolated from a Hyper-Arid Environment. J. Pure Appl. Microbiol. 2013, 7, 2713–2716. [Google Scholar]
- Zhang, A.-M.; Zhao, G.-Y.; Gao, T.-G.; Wang, W.; Li, J.; Zhang, S.-F.; Zhu, B.-C. Solubilization of Insoluble Potassium and Phosphate by Paenibacillus kribensis CX-7: A Soil Microorganism with Biological Control Potential. Afr. J. Microbiol. Res. 2013, 7, 41–47. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, Z.; Ling, N.; Yuan, Y.; Yang, X.; Chen, L.; Shen, B.; Shen, Q. Isolation and Identification of Lipopeptides Produced by B. subtilis SQR 9 for Suppressing Fusarium Wilt of Cucumber. Sci. Hortic. 2012, 135, 32–39. [Google Scholar] [CrossRef]
- González-Mendoza, D.; Argumedo-Delira, R.; Morales-Trejo, A.; Pulido-Herrera, A.; Cervantes-Díaz, L.; Grimaldo-Juarez, O.; Alarcón, A. A Rapid Method for Isolation of Total DNA from Pathogenic Filamentous Plant Fungi. Genet. Mol. Res. 2010, 9, 162–166. [Google Scholar] [CrossRef]
- Bessai, A.S.; Bensidhoum, L.; Nabti, E. Optimization of IAA Production by Telluric Bacteria Isolated from Northern Algeria. Biocatal. Agric. Biotechnol. 2022, 41, 102319. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; González-Mendoza, D. A Novel Streptomyces Rhizobacteria from Desert Soil with Diverse Anti-Fungal Properties. Rhizosphere 2020, 16, 100243. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Troncoso-Rojas, R.; Tzintzun-Camacho, O.; González-Mendoza, D.; Duran, C.C.; Grimaldo-Juárez, O.; Aviles-Marin, M.; Duran-Hernández, D. Biocontrol of Fusarium spp., Causal Agents of Damping-off in Cotton Plants by Native Bacillus subtilis Isolated from Prosopis juliflora. Int. J. Agric. Biol. 2017, 19, 713–718. [Google Scholar] [CrossRef]
- Ben Slimene, I.; Tabbene, O.; Djebali, N.; Cosette, P.; Schmitter, J.M.; Jouenne, T.; Urdaci, M.C.; Limam, F. Putative Use of a Bacillus subtilis L194 Strain for Biocontrol of Phoma medicaginis in Medicago truncatula Seedlings. Res. Microbiol. 2012, 163, 388–397. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Cisneros, M.F.; Ornelas-Paz, J.d.J.; Olivas-Orozco, G.I.; Acosta-Muñiz, C.H.; Salas-Marina, M.Á.; Molina-Corral, F.J.; Berlanga-Reyes, D.I.; Fernández-Pavía, S.P.; Cambero-Campos, O.J.; Rios-Velasco, C. Effect of Rhizosphere Inoculation with Bacillus Strains and Phytopathogens on the Contents of Volatiles and Human Health-Related Compounds in Tomato Fruits. Food Sci. Technol. 2022, 42, 1–12. [Google Scholar] [CrossRef]
- Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot. Plants 2021, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Ntushelo, K.; Ledwaba, L.K.; Rauwane, M.E.; Adebo, O.A.; Njobeh, P.B. The Mode of Action of Bacillus Species against Fusarium graminearum, Tools for Investigation, and Future Prospects. Toxins 2019, 11, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Liu, D.; Liang, Z.; Huang, K.; Wu, X. Antagonistic Activity of Bacillus subtilis CW14 and Its β-Glucanase against Aspergillus ochraceus. Food Control 2022, 131, 108475. [Google Scholar] [CrossRef]
- Perez, K.J.; Viana, J.d.S.; Lopes, F.C.; Pereira, J.Q.; dos Santos, D.M.; Oliveira, J.S.; Velho, R.V.; Crispim, S.M.; Nicoli, J.R.; Brandelli, A.; et al. Bacillus spp. Isolated from Puba as a Source of Biosurfactants and Antimicrobial Lipopeptides. Front. Microbiol. 2017, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Mora, I.; Cabrefiga, J.; Montesinos, E. Cyclic Lipopeptide Biosynthetic Genes and Products, and Inhibitory Activity of Plant-Associated Bacillus against Phytopathogenic Bacteria. PLoS ONE 2015, 10, e0127738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagar, A.; Yadav, S.S.; Sayyed, R.Z.; Sharma, S.; Ramteke, P.W. Bacillus subtilis: A Multifarious Plant Growth Promoter, Biocontrol Agent, and Bioalleviator of Abiotic Stress. In Bacilli in Agrobiotechnology: Plant Stress Tolerance, Bioremediation, and Bioprospecting; Islam, M.T., Rahman, M., Pandey, P., Eds.; Bacilli in Climate Resilient Agriculture and Bioprospecting; Springer International Publishing: Cham, Switzerland, 2022; pp. 561–580. ISBN 978-3-030-85465-2. [Google Scholar]
- Goswami, M.; Deka, S. Isolation of a Novel Rhizobacteria Having Multiple Plant Growth Promoting Traits and Antifungal Activity against Certain Phytopathogens. Microbiol. Res. 2020, 240, 126516. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for Promoting the Growth of Malus hupehensis Rehd. and Inhibiting Fusarium verticillioides. Front. Microbiol. 2019, 10, 2889. [Google Scholar] [CrossRef]
- Saleemi, M.; Kiani, M.Z.; Sultan, T.; Khalid, A.; Mahmood, S. Integrated Effect of Plant Growth-Promoting Rhizobacteria and Phosphate-Solubilizing Microorganisms on Growth of Wheat (Triticum aestivum L.) under Rainfed Condition. Agric. Food Secur. 2017, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xie, H.; Ku, Y.; Yang, X.; Chen, Y.; Yang, N.; Mei, X.; Cao, C. Chemotaxis of Bacillus cereus YL6 and Its Colonization of Chinese Cabbage Seedlings. Plant Soil 2020, 447, 413–430. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, H.; Xu, W.; Wang, Z. Isolation and Evaluation of the Plant Growth Promoting Rhizobacterium Bacillus methylotrophicus (DD-1) for Growth Enhancement of Rice Seedling. Arch. Microbiol. 2020, 202, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Zhao, D.L.; Shen, L.L.; Jing, C.L.; Zhang, C.S. Application and Mechanisms of Bacillus subtilis in Biological Control of Plant Disease. In Role of Rhizospheric Microbes in Soil: Volume 1: Stress Management and Agricultural Sustainability; Meena, V.S., Ed.; Springer: Singapore, 2018; pp. 225–250. ISBN 978-981-10-8402-7. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Amplification (bp) | Gene | Reference |
|---|---|---|---|---|
| 27F | (5’- AGAGTTTGATCCTGGCTCAG-3’) | 1400 | 16S rDNA | [22] |
| 1495R | (5’- CTACGGCTACCTTGTTACGA-3’) | |||
| ITS4 | (5’-TCCTCCGCTTATTGATATGC-3’) | 650 | ITS Region | [23] |
| ITS5 | (5’-GGAAGTAAAAGTCGTAACAAGG-3’) | |||
| Qk1F | (5’-CTTAAACGTCAGAGGCGGAG-3’) | 704 | Subtilisin | [24] |
| Qk1R | (5’-ATTGTGCAGCTGCTTGTACG-3’) | |||
| Sbo1F | (5’-TCGGTTTGTAAACTTCAACTGC-3’) | 334 | Subtilosin | |
| Sbo1R | (5’-GTCCACTAGACAAGCGGCTC-3’) | |||
| ituA1F | (5’-TGCCAGACAGTATGAGGCAG-3’) | 885 | Iturin | |
| ituA1R | (5’-CATGCCGTATCCACTGTGAC-3’) |
| Number | Treatment Name | Abbreviation of Treatment Name |
|---|---|---|
| 1 | Control (distilled water) | C |
| 2 | B. subtilis LDA-1 | LDA-1 |
| 3 | B. subtilis LDA-2 | LDA-2 |
| 4 | B. subtilis LDA-3 | LDA-3 |
| 5 | LDA-1+ LDA-2+ LDA-3 | Mix |
| 6 | Control + F. solani ICADL1 | C + F |
| 7 | B. subtilis LDA-1 + F. solani ICADL1 | LDA-1 + F |
| 8 | B. subtilis LDA-2 + F. solani ICADL1 | LDA-2 + F |
| 9 | B. subtilis LDA-3 + F. solani ICADL1 | LDA-3 + F |
| 10 | LDA-1+ LDA-2+ LDA-3 + F. solani ICADL1 | Mix + F |
| Treatments | Length | Height | |
|---|---|---|---|
| Maximum | Minimum | ||
| Control | 18.5 ± 0.7 a | 1156 ± 25 c | 822 s± 16 b |
| LDA-1 | 16.0 ± 1.1 a | 2032 ± 49 a | 653 ± 4 c |
| LDA-2 | 16.3 ± 1.0 a | 1514 ± 58 b | 913 ± 34 a |
| LDA-3 | 11.5 ± 0.7 b | 1236 ± 57 c | 211 ± 3 d |
| Significance | * | ** | *** |
| Treatments | IAA (µg/g) | GA (µg/g) | Root Length (cm) | Stem Length (cm) | N. of Secondary Roots |
|---|---|---|---|---|---|
| Control | 5.4 ± 0.06 e | 1.3 ± 0.0005 fg | 22.0 ± 1.0 c | 8.7 ± 0.3 cd | 15.0 ± 0.6 e |
| LDA-1 | 9.8 ± 0.2 b | 1.1 ± 0.0005 h | 14.3 ± 0.3 d | 7.7 ± 0.3 d | 21.3 ± 0.7 d |
| LDA-2 | 6.1 ± 0.24 de | 2.5±0.03 a | 25.3 ± 0.6 b | 13.0 ± 0.6 a | 25.0 ± 0.6 c |
| LDA-3 | 14.7 ± 0.14 a | 1.7±0.03 d | 28.7 ± 0.9 a | 9.0 ± 0.6 cd | 38.7 ± 0.7 a |
| Mix | 6.9 ± 0.18 cd | 0.8 ± 0.0005 i | 23.3 ± 0.9 bc | 11.3 ± 0.7 abc | 29.7 ± 0.3 b |
| Control + F | 5.4 ± 0.09 e | 1.4 ± 0.0005 f | 8.3 ± 0.3 f | 9.3 ± 0.3 bcd | 20.0 ± 0.6 d |
| LDA-1 + F | 6.1 ± 0.07 de | 1.2 ± 0.03 g | 11.7 ± 0.3 de | 12.0 ± 1.1 ab | 8.3 ± 0.3 f |
| LDA-2 + F | 7.4 ± 0.3 c | 1.6 ± 0.02 e | 9.0 ± 0.6 ef | 9.0 ± 0.6 cd | 25.7 ± 0.9 c |
| LDA-3 + F | 6.6 ± 0.03 cd | 2.1 ± 0.0005 b | 9.7 ± 0.3 ef | 8.7 ± 0.3 cd | 26.3 ± 0.9 bc |
| Mix + F | 5.4 ± 0.44 e | 1.9 ± 0.0005 c | 11.3 ± 0.3 def | 11.0 ± 0.6 abc | 29.7 ± 0.9 b |
| Significance | *** | *** | *** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelmoteleb, A.; Moreno-Ramírez, L.; Valdez-Salas, B.; Seleiman, M.F.; El-Hendawy, S.; Aldhuwaib, K.J.; Alotaibi, M.; González-Mendoza, D. New Bacillus subtilis Strains Isolated from Prosopis glandulosa Rhizosphere for Suppressing Fusarium Spp. and Enhancing Growth of Gossypium hirsutum L. Biology 2023, 12, 73. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12010073
Abdelmoteleb A, Moreno-Ramírez L, Valdez-Salas B, Seleiman MF, El-Hendawy S, Aldhuwaib KJ, Alotaibi M, González-Mendoza D. New Bacillus subtilis Strains Isolated from Prosopis glandulosa Rhizosphere for Suppressing Fusarium Spp. and Enhancing Growth of Gossypium hirsutum L. Biology. 2023; 12(1):73. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12010073
Chicago/Turabian StyleAbdelmoteleb, Ali, Lizbeth Moreno-Ramírez, Benjamín Valdez-Salas, Mahmoud F. Seleiman, Salah El-Hendawy, Khalid J. Aldhuwaib, Majed Alotaibi, and Daniel González-Mendoza. 2023. "New Bacillus subtilis Strains Isolated from Prosopis glandulosa Rhizosphere for Suppressing Fusarium Spp. and Enhancing Growth of Gossypium hirsutum L." Biology 12, no. 1: 73. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12010073