
Plant–Microbes Interaction: Exploring the Impact of Cold-Tolerant Bacillus Strains RJGP41 and GBAC46 Volatiles on Tomato Growth Promotion through Different Mechanisms
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacillus Strains, Plant Material, and Culture Conditions
2.2. Production of Indole 3-Acetic Acid (IAA)
2.3. In Vitro Growth Promotion Assay by Bacillus spp.
2.4. In Planta Growth Promotion Assay by Bacillus spp.
2.5. GC-MS Analysis of Bacillus spp. VOCs
2.6. In Vitro Plant Growth Promotion by Pure VOCs
2.7. In Vivo Plant Growth Promotion by Bacillus VOCs
2.8. Determination of Antioxidant Enzyme Activity and Total Protein Contents
2.9. Relative Expression of Growth Promotion Genes Exposed to Pure VOCs
2.10. Statistial Analysis
3. Results
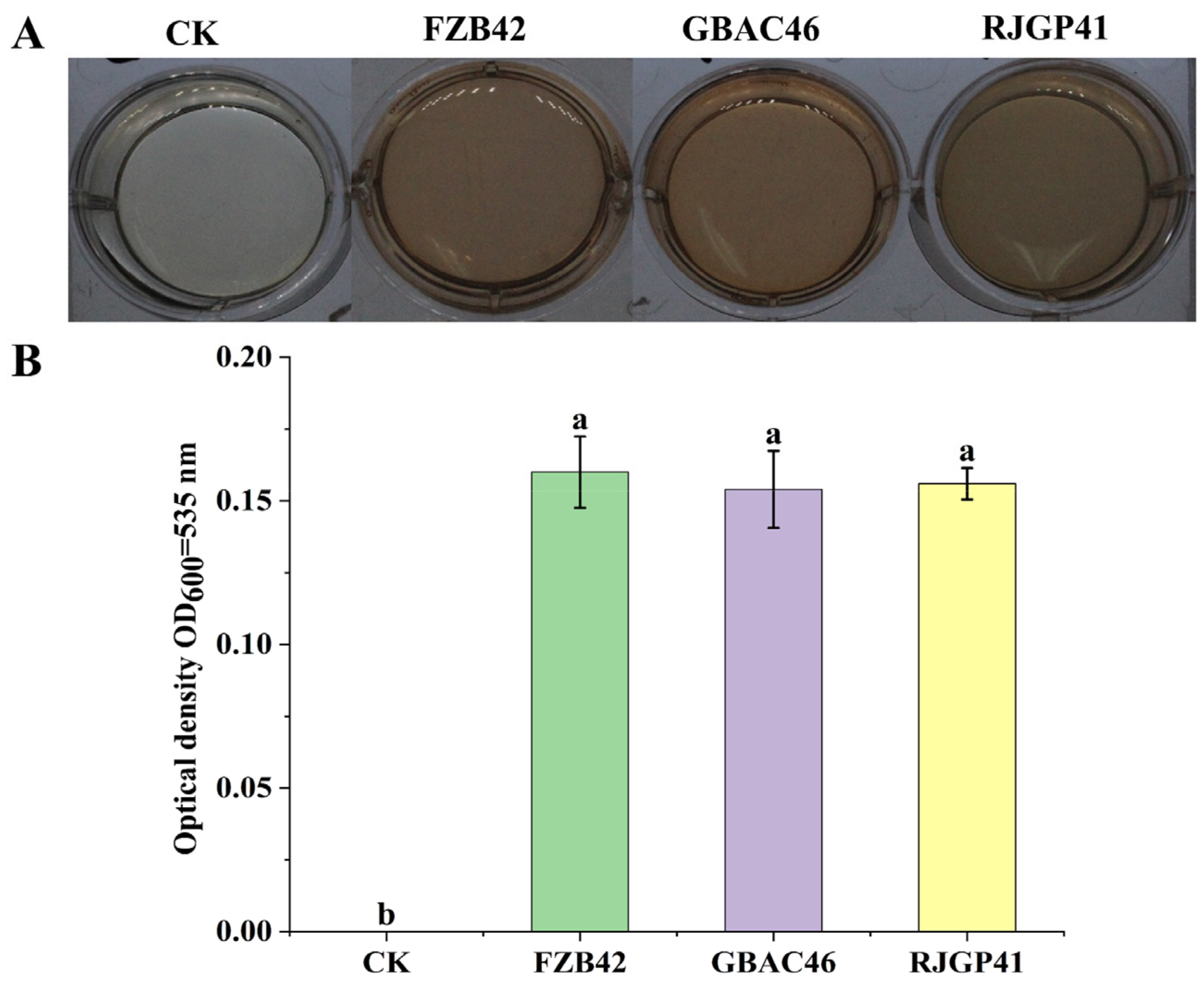
3.1. Production of Indole 3-Acidic Acid (IAA)
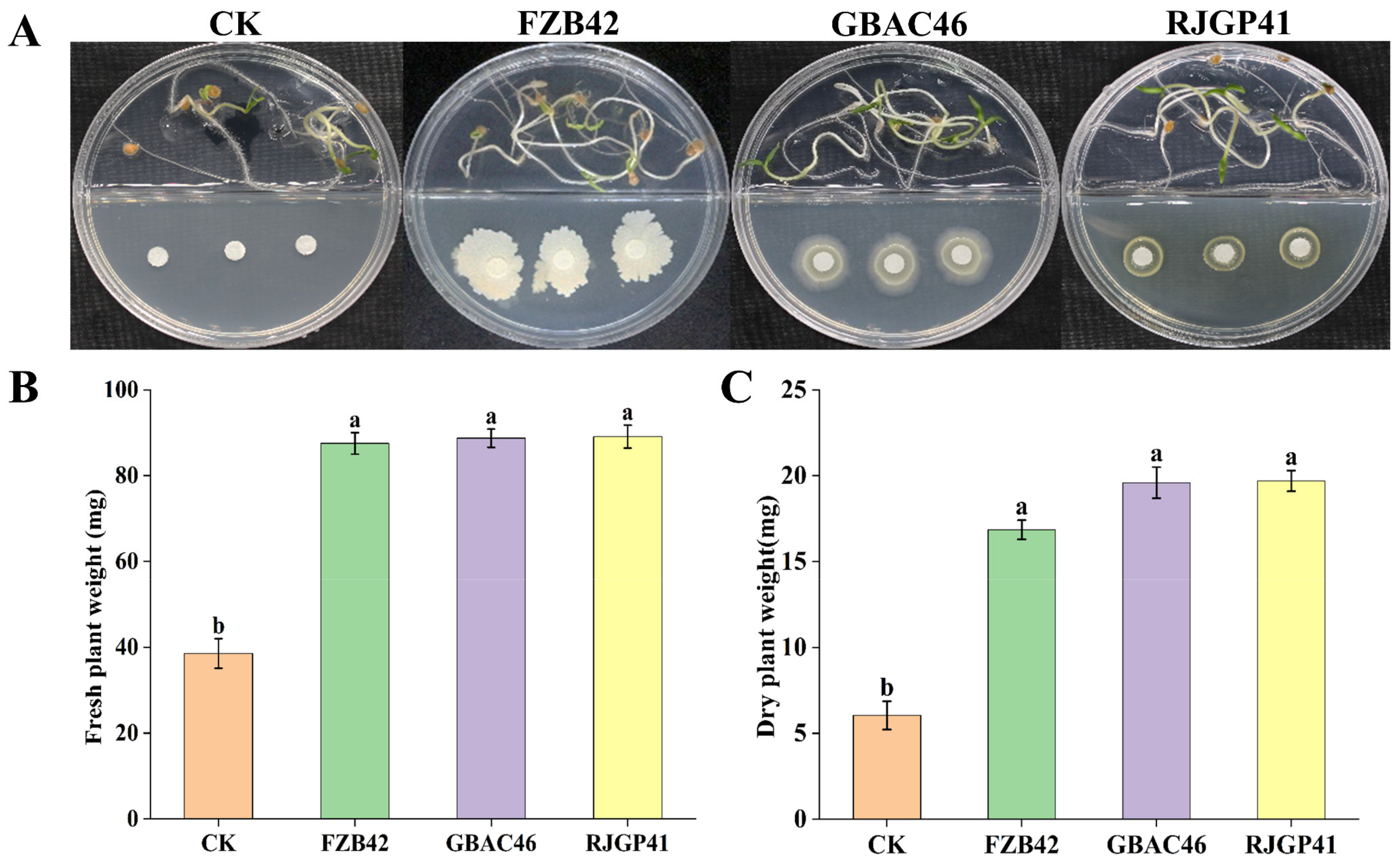
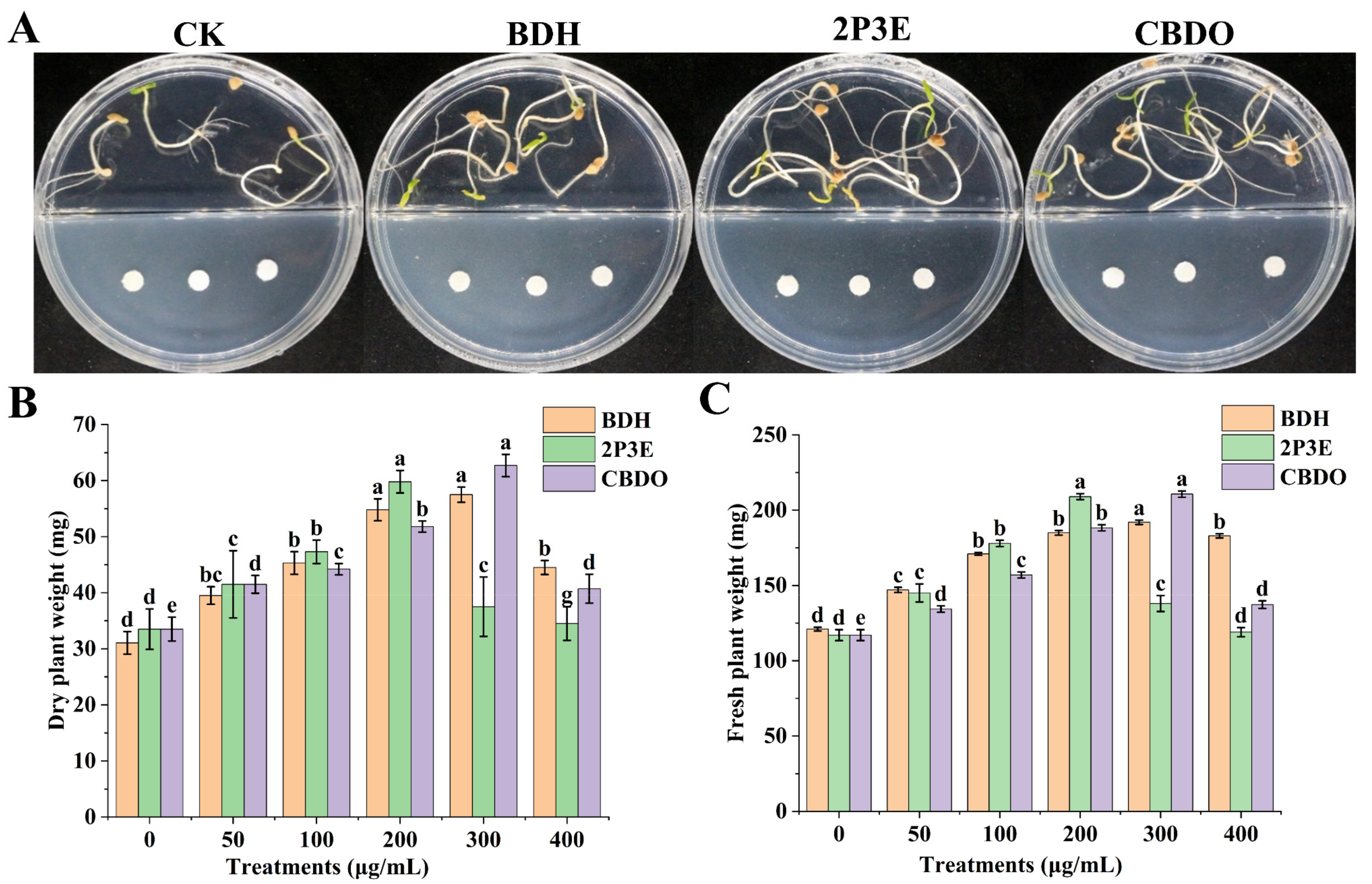
3.2. In Vitro Plant Growth Promotion by Bacillus VOCs
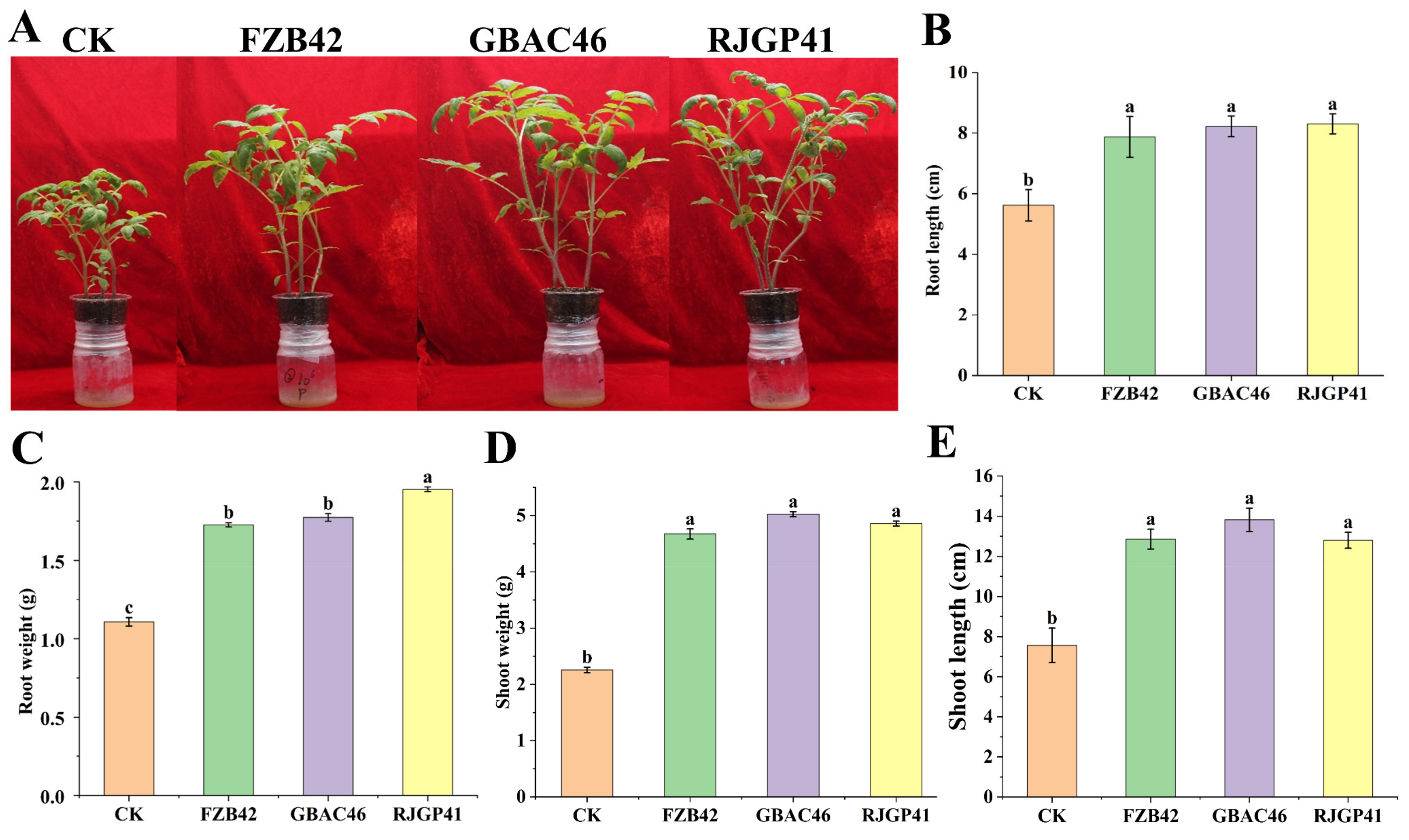
3.3. In Vivo Plant Growth Promotion by Bacillus VOCs
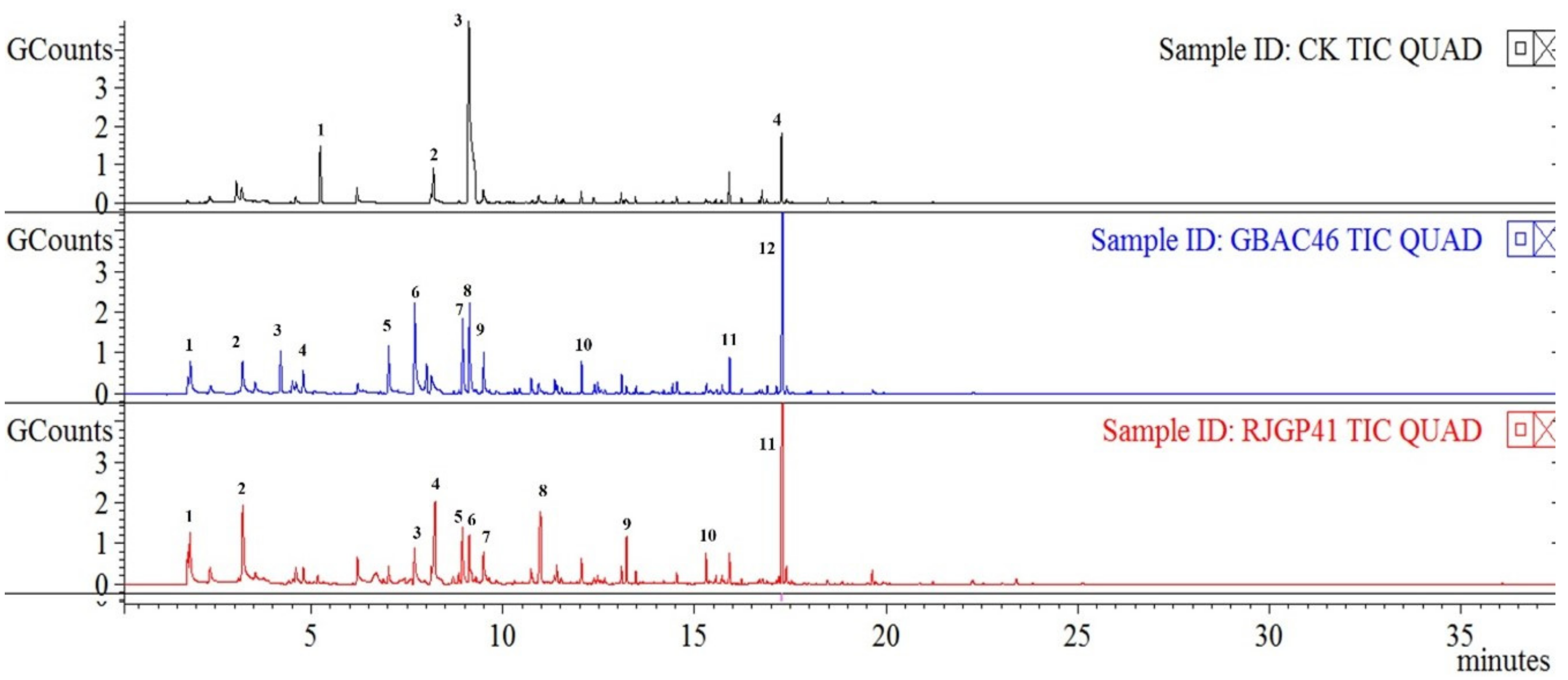
3.4. GC-MS Analysis of Pure VOCs Emitted by Bacillus Strains
3.5. In Vitro Plant Growth Promotion by Pure VOCs
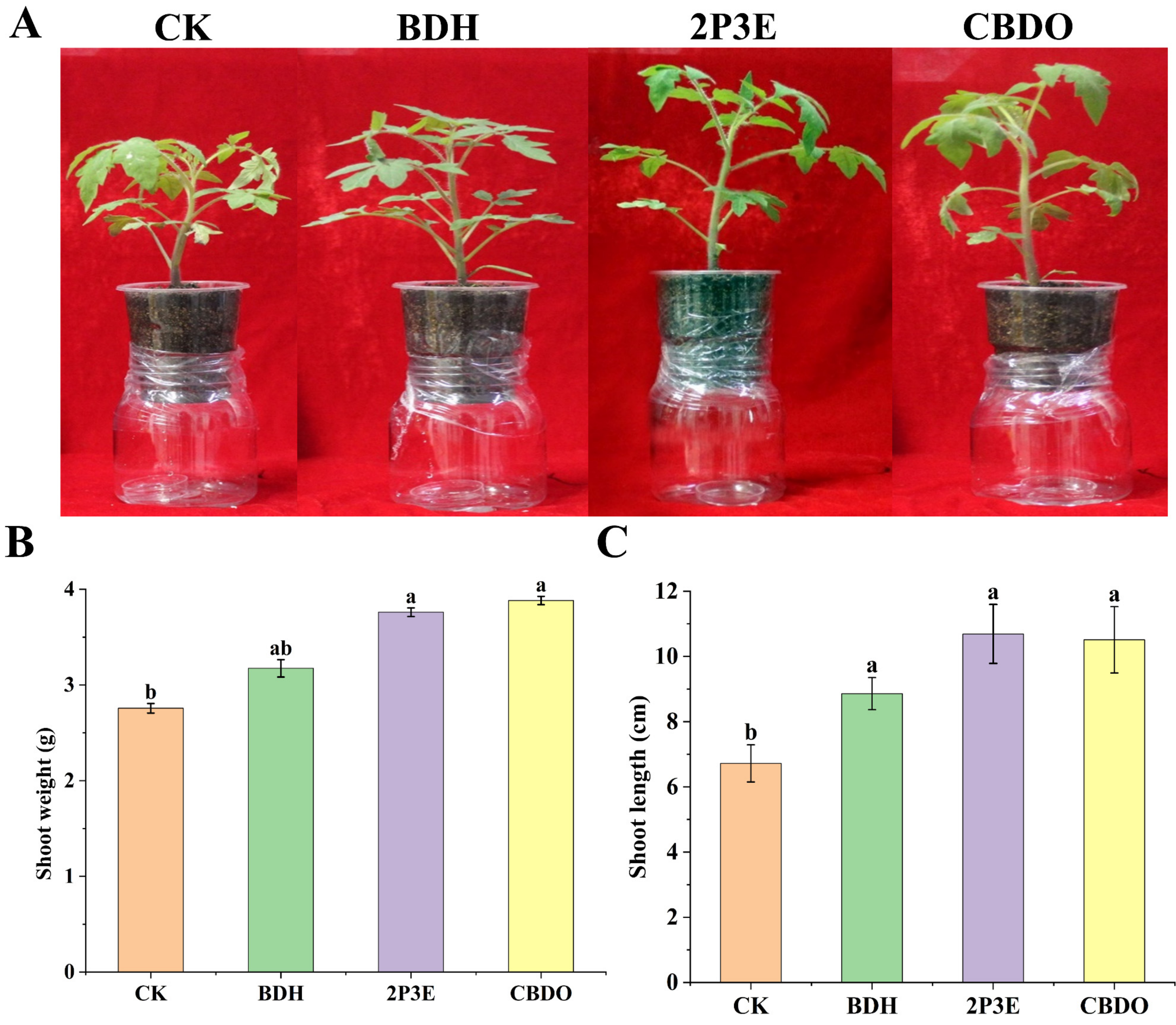
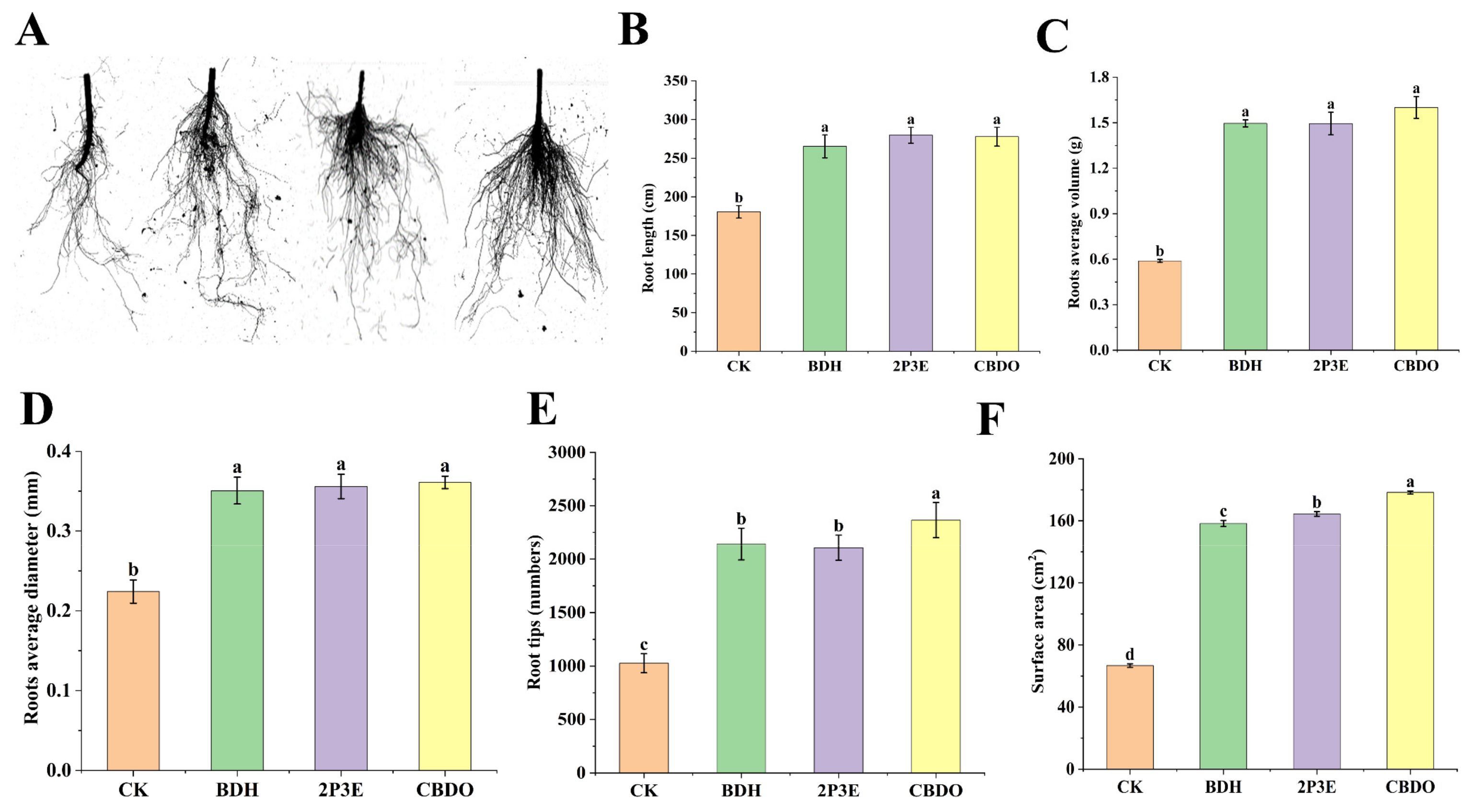
3.6. In Vivo Plant Growth Promotion by Pure VOCs
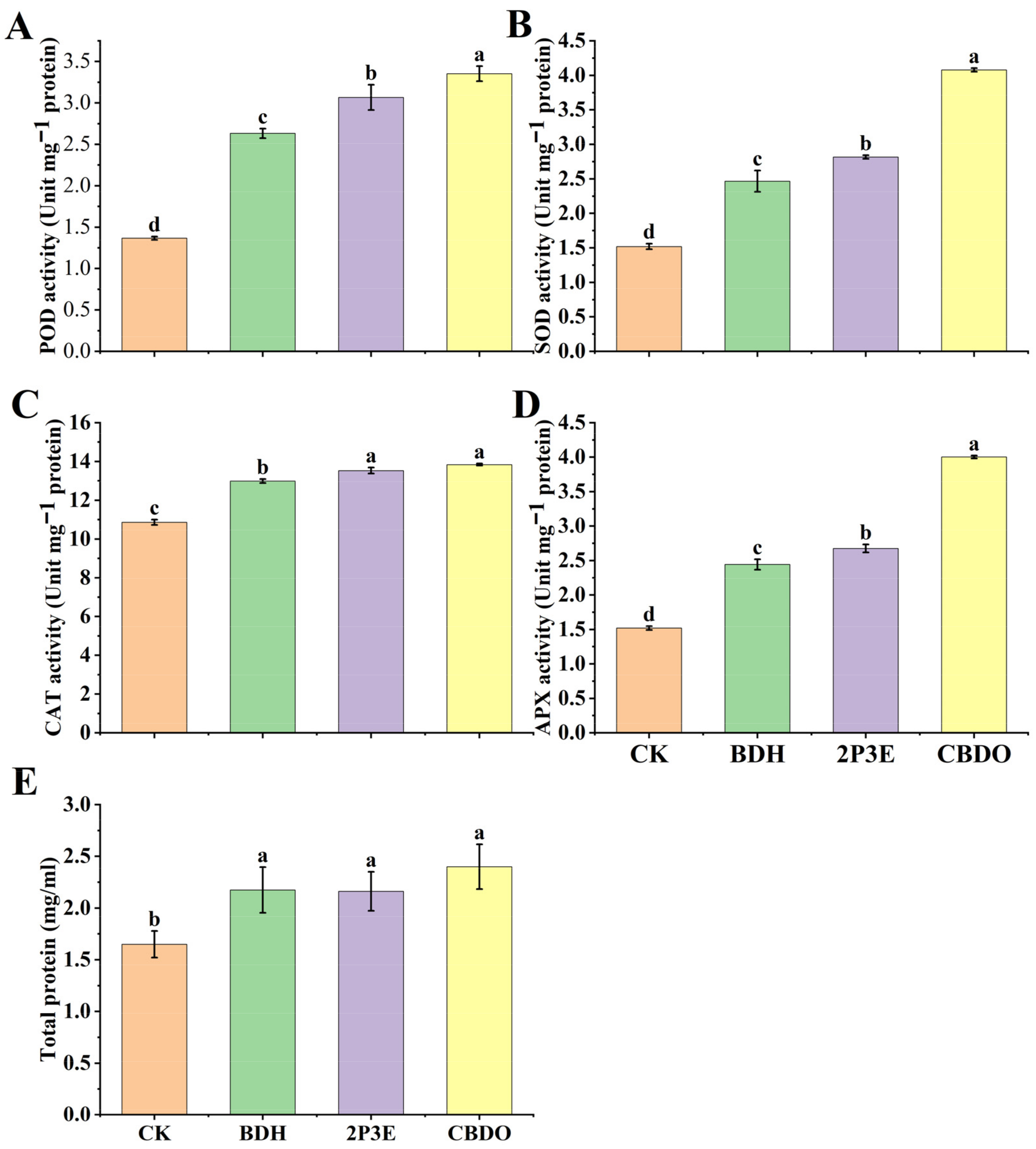
3.7. Antioxidant Activity and Total Protein Contents Post-Exposure to Pure VOCs in Tomato Plants
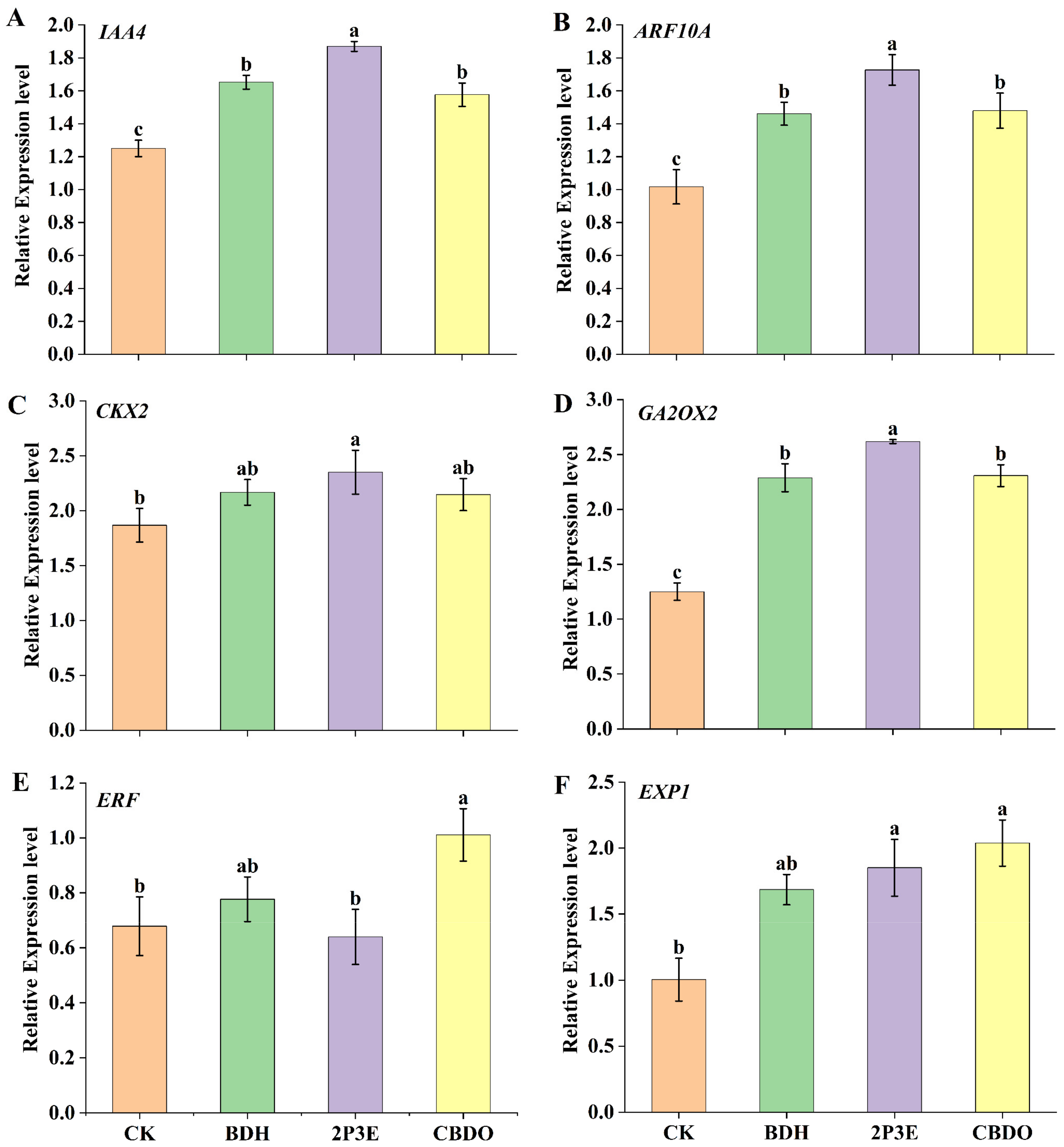
3.8. Effect of Pure VOCs on Plant Growth Promotion Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A Review on the Plant Microbiome: Ecology, Functions, and Emerging Trends in Microbial Application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Yu, C.; Wang, Y.; Sheng, T.; Zhao, X.; Wu, X.; Jing, L.; Gu, Q.; Wu, H.; Gao, X. High Killing Rate of Nematode and Promotion of Rice Growth by Synthetic Volatiles from Bacillus Strains Due to Enhanced Oxidative Stress Response. Physiol. Plant. 2023, 175, e13868. [Google Scholar] [CrossRef] [PubMed]
- Verbon, E.H.; Liberman, L.M. Beneficial Microbes Affect Endogenous Mechanisms Controlling Root Development. Trends Plant Sci. 2016, 21, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Qin, Z.; Wu, S.; Zhao, P.; Zhen, C.; Gao, H. Antifungal Mechanism of Volatile Organic Compounds Produced by Bacillus Subtilis CF-3 on Colletotrichum Gloeosporioides Assessed Using Omics Technology. J. Agric. Food Chem. 2021, 69, 5267–5278. [Google Scholar] [CrossRef]
- Yin, C.; Casa Vargas, J.M.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere Community Selection Reveals Bacteria Associated with Reduced Root Disease. Microbiome 2021, 9, 86. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant Growth Promoting Bacteria as an Alternative Strategy for Salt Tolerance in Plants: A Review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Khan, A.R.; Elkomy, M.H.; Ibrahim, Y.E.; Hamad, Y.K.; Molan, Y.Y.; Saleh, A.A. Organic Management of Tomato Fusarium Wilt Using a Native Bacillus Subtilis Strain and Compost Combination in Saudi Arabia. Int. J. Agric. Biol. 2020, 23, 1003–1012. [Google Scholar] [CrossRef]
- Gómez Expósito, R.; De Bruijn, I.; Postma, J.; Raaijmakers, J.M. Current Insights into the Role of Rhizosphere Bacteria in Disease Suppressive Soils. Front. Microbiol. 2017, 8, 2529. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayilara, M.S.; Ayangbenro, A.S.; Babalola, O.O. Genome Mining of Three Plant Growth-Promoting Bacillus Species from Maize Rhizosphere. Appl. Biochem. Biotechnol. 2021, 193, 3949–3969. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Diversity and Natural Functions of Antibiotics Produced by Beneficial and Plant Pathogenic Bacteria. Annu. Rev. Phytopathol. 2012, 50, 403–424. [Google Scholar] [CrossRef]
- Choub, V.; Won, S.J.; Ajuna, H.B.; Moon, J.H.; Choi, S.I.; Lim, H.I.; Ahn, Y.S. Antifungal Activity of Volatile Organic Compounds from Bacillus Velezensis CE 100 against Colletotrichum Gloeosporioides. Horticulturae 2022, 8, 557. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H. Phytostimulation and Biocontrol Potential of Gram-Positive Endospore-Forming Bacilli. Planta 2021, 254. [Google Scholar] [CrossRef]
- Guimarães Pacifico, M.; Eckstein, B.; Bettiol, W. Screening of Bacillus for the Development of Bioprotectants for the Control of Fusarium oxysporum f. Sp. Vasinfectum and Meloidogye Incognita. Biol. Control 2021, 164, 104764. [Google Scholar] [CrossRef]
- Zerrouk, I.Z.; Rahmoune, B.; Auer, S.; Rößler, S.; Lin, T.; Baluska, F.; Dobrev, P.I.; Motyka, V.; Ludwig-Müller, J. Growth and Aluminum Tolerance of Maize Roots Mediated by Auxin- and Cytokinin-Producing Bacillus Toyonensis Requires Polar Auxin Transport. Environ. Exp. Bot. 2020, 176, 104064. [Google Scholar] [CrossRef]
- Kamilova, F.; Okon, Y.; Weert, S.; Hora, K. Commercialization of Microbes: Manufacturing, Inoculation, Best Practice for Objective Field Testing, and Registration. In Principles of Plant Microbe Interactions. Microbes for Sustainable Agriculture; Lugtenberg, B., Ed.; Springer: Cham, Switzerland, 2015; pp. 319–327. ISBN 978-3-319-08574-6. [Google Scholar]
- Araujo, F.F.; Bonifacio, A.; Bavaresco, L.G.; Mendes, L.W.; Araujo, A.S.F. Bacillus Subtilis Changes the Root Architecture of Soybean Grown on Nutrient-Poor Substrate. Rhizosphere 2021, 18, 100348. [Google Scholar] [CrossRef]
- Sheikh, T.M.M.; Zhou, D.; Haider, M.S.; Hussain, S.; Wang, N.; Chen, S.; Zhao, Y.; Wen, X.; Feng, H.; Wang, X.; et al. Volatile Organic Compounds from Pythium Oligandrum Play a Role in Its Parasitism on Plant-Pathogenic Pythium Myriotylum. Appl. Environ. Microbiol. 2023, 89, e02036-22. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Farzand, A.; Khan, A.R.; Ling, H.; Gao, X. Nematicidal Volatiles from Bacillus Atrophaeus Gbsc56 Promote Growth and Stimulate Induced Systemic Resistance in Tomato against Meloidogyne Incognita. Int. J. Mol. Sci. 2021, 22, 5049. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Niu, Y.; Huo, R.; Gao, X. Bacillus Volatiles Adversely Affect the Physiology and Ultra-Structure of Ralstonia Solanacearum and Induce Systemic Resistance in Tobacco against Bacterial Wilt. Sci. Rep. 2017, 7, 40481. [Google Scholar] [CrossRef]
- Zhao, P.; Li, P.; Wu, S.; Zhou, M.; Zhi, R.; Gao, H. Volatile Organic Compounds (VOCs) from Bacillus Subtilis CF-3 Reduce Anthracnose and Elicit Active Defense Responses in Harvested Litchi Fruits. AMB Express 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile Organic Compounds (VOCs) Produced by Biocontrol Yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef]
- Tilocca, B.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. A Proteomic Investigation of Aspergillus Carbonarius Exposed to Yeast Volatilome or to Its Major Component 2-Phenylethanol Reveals Major Shifts in Fungal Metabolism. Int. J. Food Microbiol. 2019, 306, 108265. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Ryu, C.M. Are Bacterial Volatile Compounds Poisonous Odors to a Fungal Pathogen Botrytis Cinerea, Alarm Signals to Arabidopsis Seedlings for Eliciting Induced Resistance, or Both? Front. Microbiol. 2016, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Kim, M.S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial Volatile Emissions Regulate Auxin Homeostasis and Cell Expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar] [CrossRef]
- Vespermann, A.; Kai, M.; Piechulla, B. Rhizobacterial Volatiles Affect the Growth of Fungi and Arabidopsis Thaliana. Appl. Environ. Microbiol. 2007, 73, 5639–5641. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Haustein, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial Volatiles and Their Action Potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef]
- Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Improved Heat Stress Tolerance of Wheat Seedlings by Bacterial Seed Treatment. Plant Soil 2014, 379, 337–350. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Yu, C.; Wang, Y.; Gu, Q.; Wu, H.; Gao, X. Cadmium Tolerant Microbial Strains Possess Different Mechanisms for Cadmium Biosorption and Immobilization in Rice Seedlings. Chemosphere 2022, 303, 135206. [Google Scholar] [CrossRef]
- Dhouib, H.; Zouari, I.; Abdallah, D.B.; Belbahri, L.; Taktak, W.; Triki, M.A.; Tounsi, S. Potential of a Novel Endophytic Bacillus Velezensis in Tomato Growth Promotion and Protection against Verticillium Wilt Disease. Biol. Control 2019, 139, 104092. [Google Scholar] [CrossRef]
- Wu, H.; Gu, Q.; Xie, Y.; Lou, Z.; Xue, P.; Fang, L.; Yu, C.; Jia, D.; Huang, G.; Zhu, B.; et al. Cold-Adapted Bacilli Isolated from the Qinghai–Tibetan Plateau Are Able to Promote Plant Growth in Extreme Environments. Environ. Microbiol. 2019, 21, 3505–3526. [Google Scholar] [CrossRef]
- Borriss, R.; Wu, H.; Gao, X. Secondary Metabolites of the Plant Growth Promoting Model Rhizobacterium Bacillus Velezensis FZB42 Are Involved in Direct Suppression of Plant Pathogens and in Stimulation of Plant-Induced Systemic Resistance. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Singh, H.B., Keswani, C., Reddy, M.S., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019; pp. 147–168. ISBN 978-981-13-5862-3. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wagi, S.; Ahmed, A. Bacillus Spp.: Potent Microfactories of Bacterial IAA. PeerJ 2019, 7, e7258. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of Plant Growth by Pseudomonas Fluorescens Strain SS101 via Novel Volatile Organic Compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef]
- Farag, M.A.; Ryu, C.M.; Sumner, L.W.; Paré, P.W. GC-MS SPME Profiling of Rhizobacterial Volatiles Reveals Prospective Inducers of Growth Promotion and Induced Systemic Resistance in Plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.; Gao, X. Plant Growth Promotion by Volatile Organic Compounds Produced by Bacillus Subtilis SYST2. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile Organic Compounds from Paenibacillus Polymyxa KM2501-1 Control Meloidogyne Incognita by Multiple Strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.C.P.; Campos, V.P.; Barros, A.F.; Pedroso, L.A.; de Freitas Silva, M.; de Souza, J.T.; Pedroso, M.P.; de Medeiros, F.H.V. Performance of Volatiles Emitted from Different Plant Species against Juveniles and Eggs of Meloidogyne Incognita. Crop Prot. 2019, 116, 196–203. [Google Scholar] [CrossRef]
- Rajer, F.U.; Samma, M.K.; Ali, Q.; Rajar, W.A.; Wu, H.; Raza, W.; Xie, Y.; Tahir, H.A.S.; Gao, X. Bacillus Spp.-Mediated Growth Promotion of Rice Seedlings and Suppression of Bacterial Blight Disease under Greenhouse Conditions. Pathogens 2022, 11, 1251. [Google Scholar] [CrossRef]
- Wu, X.; Fan, Y.; Wang, R.; Zhao, Q.; Ali, Q.; Wu, H.; Gu, Q.; Borriss, R.; Xie, Y.; Gao, X. Bacillus Halotolerans KKD1 Induces Physiological, Metabolic and Molecular Reprogramming in Wheat under Saline Condition. Front. Plant Sci. 2022, 13, 978066. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Mu, G.; Hussain, A.; Yuanyuan, Q.; Yu, C.; Xu, Y.; Manghwar, H.; Gu, Q.; Wu, H.; et al. Revealing Plant Growth-Promoting Mechanisms of Bacillus Strains in Elevating Rice Growth and Its Interaction with Salt Stress. Front. Plant Sci. 2022, 13, 994902. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sikand, K.; Singh, J.; Ebron, J.S.; Shukla, G.C. Housekeeping Gene Selection Advisory: Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and β-Actin Are Targets of MiR-644a. PLoS ONE 2012, 7, e47510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fincheira, P.; Parra, L.; Mutis, A.; Parada, M.; Quiroz, A. Volatiles Emitted by Bacillus Sp. BCT9 Act as Growth Modulating Agents on Lactuca Sativa Seedlings. Microbiol. Res. 2017, 203, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y.-J. Identification of Endophytic Bacillus Velezensis ZSY-1 Strain and Antifungal Activity of Its Volatile Compounds against Alternaria Solani and Botrytis Cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Zang, H.; Jun Wu, H.; Uddin Rajer, F.; Gao, X. Antibacterial Effects of Volatiles Produced by Bacillus Strain D13 against Xanthomonas Oryzae Pv. Oryzae. Mol. Plant Pathol. 2018, 19, 49–58. [Google Scholar] [CrossRef]
- Ben Abdallah, D.; Frikha-Gargouri, O.; Tounsi, S. Rizhospheric Competence, Plant Growth Promotion and Biocontrol Efficacy of Bacillus Amyloliquefaciens Subsp. Plantarum Strain 32a. Biol. Control 2018, 124, 61–67. [Google Scholar] [CrossRef]
- Sukkasem, P.; Kurniawan, A.; Kao, T.C.; Chuang, H.-w. A Multifaceted Rhizobacterium Bacillus Licheniformis Functions as a Fungal Antagonist and a Promoter of Plant Growth and Abiotic Stress Tolerance. Environ. Exp. Bot. 2018, 155, 541–551. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Hernández, A.; Galvan, A.I.; Córdoba, M.G.; Casquete, R.; Serradilla, M.J.; Martín, A. Selection and Application of Antifungal VOCs-Producing Yeasts as Biocontrol Agents of Grey Mould in Fruits. Food Microbiol. 2020, 92, 103556. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial Volatiles Induce Systemic Resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Ramyabharathi, S.A.; Raguchander, T. Induction of Defense Enzymes in Tomato in Response to Treatment with Bacillus Subtilis EPCO 16 Liquid Formulation. J. Mycol. Pl. Pathol. 2014, 44, 122–125. [Google Scholar]
- Asari, S.; Matzén, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple Effects of Bacillus Amyloliquefaciens Volatile Compounds: Plant Growth Promotion and Growth Inhibition of Phytopathogens. FEMS Microbiol. Ecol. 2016, 92, fiw070. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xie, X.; Kim, M.-S.; Kornyeyev, D.A.; Holaday, S.; Paré, P.W. Soil Bacteria Augment Arabidopsis Photosynthesis by Decreasing Glucose Sensing and Abscisic Acid Levels in Planta. Plant J. 2008, 56, 264–273. [Google Scholar] [CrossRef]
- Bensmihen, S. Hormonal Control of Lateral Root and Nodule Development in Legumes. Plants 2015, 4, 523–547. [Google Scholar] [CrossRef]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; De La Cruz, H.R.; Macías-Rodríguez, L. Plant Growth-Promoting Rhizobacteria Modulate Root-System Architecture in Arabidopsis Thaliana through Volatile Organic Compound Emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-Tolerant Plant Growth-Promoting Bacillus Pumilus Strain JPVS11 to Enhance Plant Growth Attributes of Rice and Improve Soil Health under Salinity Stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Jiang, Q.; Wang, R.; Wang, Z.; Mu, G.; Khan, S.A.; Khan, A.R.; Manghwar, H.; Wu, H.; et al. Salt Tolerant Bacillus Strains Improve Plant Growth Traits and Regulation of Phytohormones in Wheat under Salinity Stress. Plants 2022, 11, 2769. [Google Scholar] [CrossRef]
- Mubeen, S.; Shahzadi, I.; Akram, W.; Saeed, W.; Yasin, N.A.; Ahmad, A.; Shah, A.A.; Siddiqui, M.H.; Alamri, S. Calcium Nanoparticles Impregnated With Benzenedicarboxylic Acid: A New Approach to Alleviate Combined Stress of DDT and Cadmium in Brassica Alboglabra by Modulating Bioacummulation, Antioxidative Machinery and Osmoregulators. Front. Plant Sci. 2022, 13, 825829. [Google Scholar] [CrossRef]
- Ghazala, I.; Chiab, N.; Saidi, M.N.; Gargouri-Bouzid, R. The Plant Growth-Promoting Bacteria Strain Bacillus Mojavensis I4 Enhanced Salt Stress Tolerance in Durum Wheat. Curr. Microbiol. 2023, 80, 178. [Google Scholar] [CrossRef]
- Liang, Z.; Ali, Q.; Wang, Y.; Mu, G.; Kan, X.; Ren, Y.; Manghwar, H.; Gu, Q.; Wu, H.; Gao, X. Toxicity of Bacillus Thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides Besseyi. Int. J. Mol. Sci. 2022, 23, 8189. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Ryu, K.C.; Lee, S.; Bee, H.; Soo, K.; Jung, H.; Lee, H.; Lee, K.; Sik, W.; Hee, M.J.; et al. Proteome Analysis of Arabidopsis Seedlings Exposed to Bacterial Volatiles. Planta 2010, 232, 1355–1370. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr.# | GBAC-46 VOCs Detected via GCMS | Probability | MW | CAS# | Formula |
|---|---|---|---|---|---|
| 1 | Undecanal, 2-methyl | 23.51 | 184 | 110-41-8 | C12H24O |
| 2 | 1-4-pentadiene | 18.01 | 68 | 591-93-5 | C5H8 |
| 3 | L-threonine | 28.59 | 119 | 72-19-5 | C4H9NO3 |
| 4 | Amiodarone | 8.54 | 645 | 1951-25-3 | C25H29I2NO3 |
| 5 | Dl-2-Aminoadipic acid | 47.06 | 161 | 542-32-5 | C6H11NO4 |
| 6 | 2-pentanone, 3-methyl | 76.30 | 100 | 565-61-7 | C6H12O |
| 7 | 3-Hexanamine, 3-ethyl | 40.72 | 129 | 56667-17-5 | C8H19N |
| 8 | 1-dodecanamine, N, N dimethyl | 13.95 | 213 | 112-18-5 | C14H31N |
| 9 | 2-Heptanone, 6-methyl | 48.24 | 128 | 928-68-7 | C6H16O |
| 10 | 2-Undecanone | 37.56 | 170 | 112-12-9 | C11H22O |
| 11 | Pentadecane | 16.19 | 212 | 629-62-9 | C15H32 |
| 12 | 2-Tridecanone | 36.07 | 198 | 593-08-08 | C13H26O |
| RJGP41 | |||||
| 1 | Cyclobutane 1,2,3,4-tetramethyl | 31.15 | 112 | 69531-57-3 | C8H16 |
| 2 | 1,3-Cyclobutanediol,2,2,4,4-tetramethyl | 63.31 | 144 | 3010-96-6 | C8H16O2 |
| 3 | 3 Hexanamine, 3-ethyl | 11.68 | 129 | 56667-17-5 | C8H19N |
| 4 | 2 Hexanone, 5-methyl | 26.78 | 114 | 110-12-3 | C7H14O |
| 5 | 2 Heptanone | 23.03 | 114 | 110-43-0 | C7H14O |
| 6 | 2 Heptanone, 6-methyl | 46.25 | 128 | 928-68-7 | C8H16O |
| 7 | Undecanal, 2-methyl-thyl | 30.73 | 184 | 110-41-8 | C12H24O |
| 8 | 1,1,3,3,5,5,7,7,9,9, decamethyl-9 (2 methylpropoxy) pentasiloxane-1-01 | 15.10 | 444 | NONE | C14H40O6Si5 |
| 9 | Choleston-3-one cyclic 1,2-ethane diyl-actal (5-beta) | 22.03 | 430 | 25328-53-4 | C29H50O2 |
| 10 | Tridecane | 12.14 | 184 | 629-50-5 | C13H28 |
| 11 | Octasiloxane,1,1,3,3,5,5,7,7,9,9,11,11,13,13,15,15-hexadecamethyl | 42.41 | 5781 | 19095-24-0 | C16H50O7Si8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.R.; Ali, Q.; Ayaz, M.; Bilal, M.S.; Sheikh, T.M.M.; Gu, Q.; Wu, H.; Gao, X. Plant–Microbes Interaction: Exploring the Impact of Cold-Tolerant Bacillus Strains RJGP41 and GBAC46 Volatiles on Tomato Growth Promotion through Different Mechanisms. Biology 2023, 12, 940. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12070940
Khan AR, Ali Q, Ayaz M, Bilal MS, Sheikh TMM, Gu Q, Wu H, Gao X. Plant–Microbes Interaction: Exploring the Impact of Cold-Tolerant Bacillus Strains RJGP41 and GBAC46 Volatiles on Tomato Growth Promotion through Different Mechanisms. Biology. 2023; 12(7):940. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12070940
Chicago/Turabian StyleKhan, Abdur Rashid, Qurban Ali, Muhammad Ayaz, Muhammad Saqib Bilal, Taha Majid Mahmood Sheikh, Qin Gu, Huijun Wu, and Xuewen Gao. 2023. "Plant–Microbes Interaction: Exploring the Impact of Cold-Tolerant Bacillus Strains RJGP41 and GBAC46 Volatiles on Tomato Growth Promotion through Different Mechanisms" Biology 12, no. 7: 940. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12070940