
Phylogenetic Analysis of Pyruvate-Ferredoxin Oxidoreductase, a Redox Enzyme Involved in the Pharmacological Activation of Nitro-Based Prodrugs in Bacteria and Protozoa


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Sequences
2.2. Sequence Alignment
2.3. Phylogenetic Analyses
2.4. Screening Selected Proteomes against HMM Profile Libraries
2.5. Gene Enrichment Analysis
3. Results
3.1. BLAST Pilot Experiment
3.2. Distribution of PFOR and Control Proteins along the Tree of Life
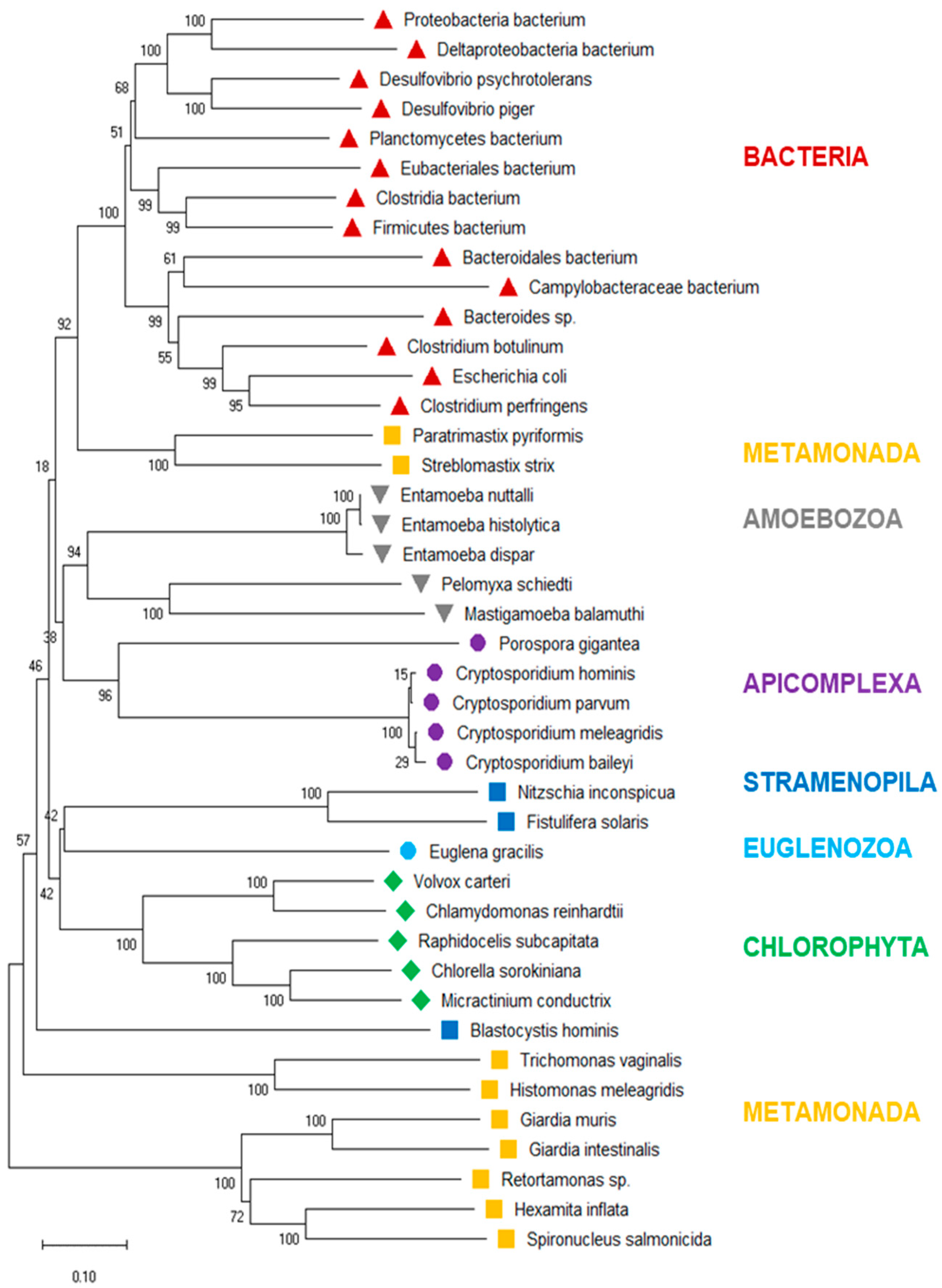
3.3. Phylogenetic Tree of PFOR
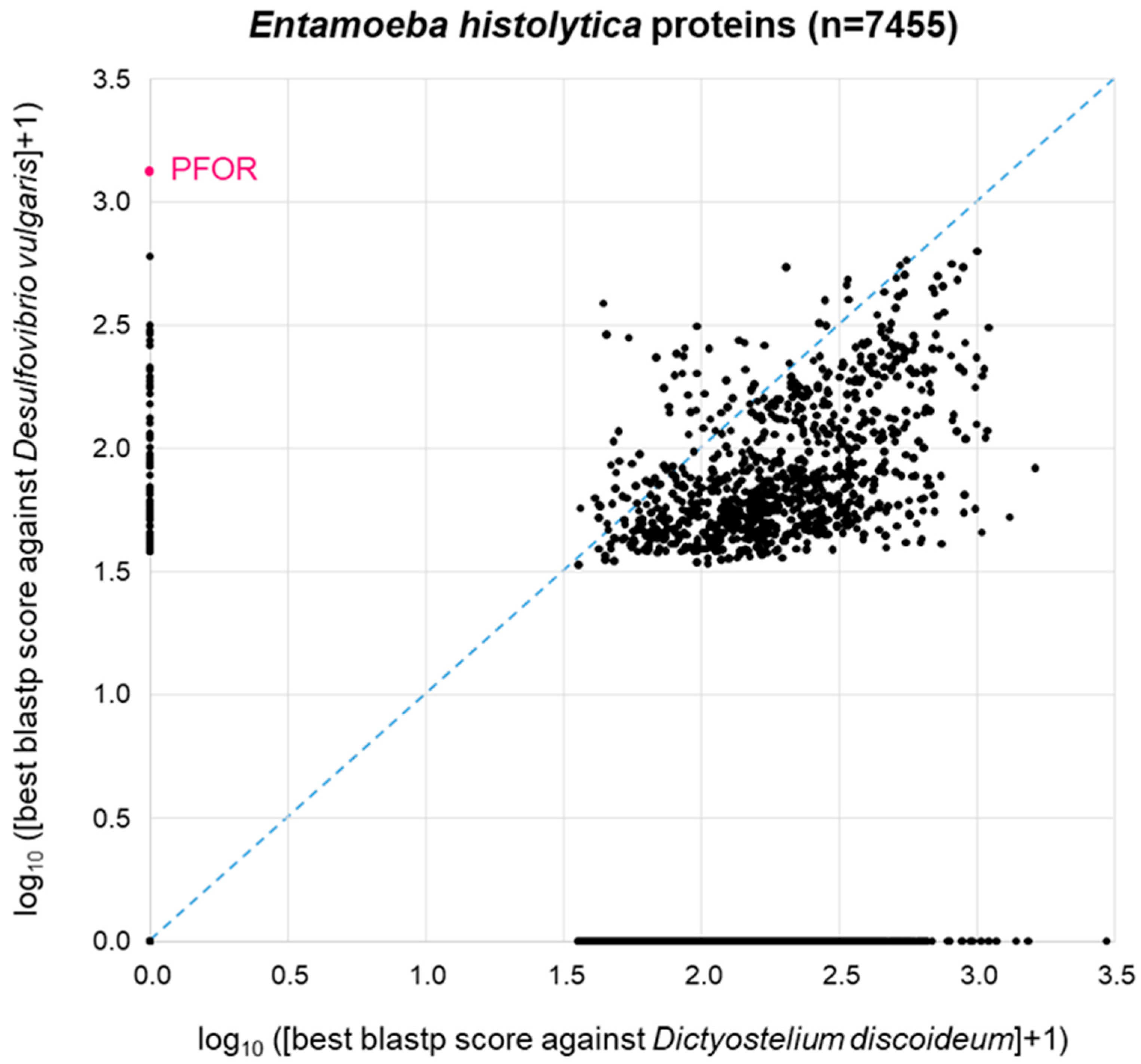
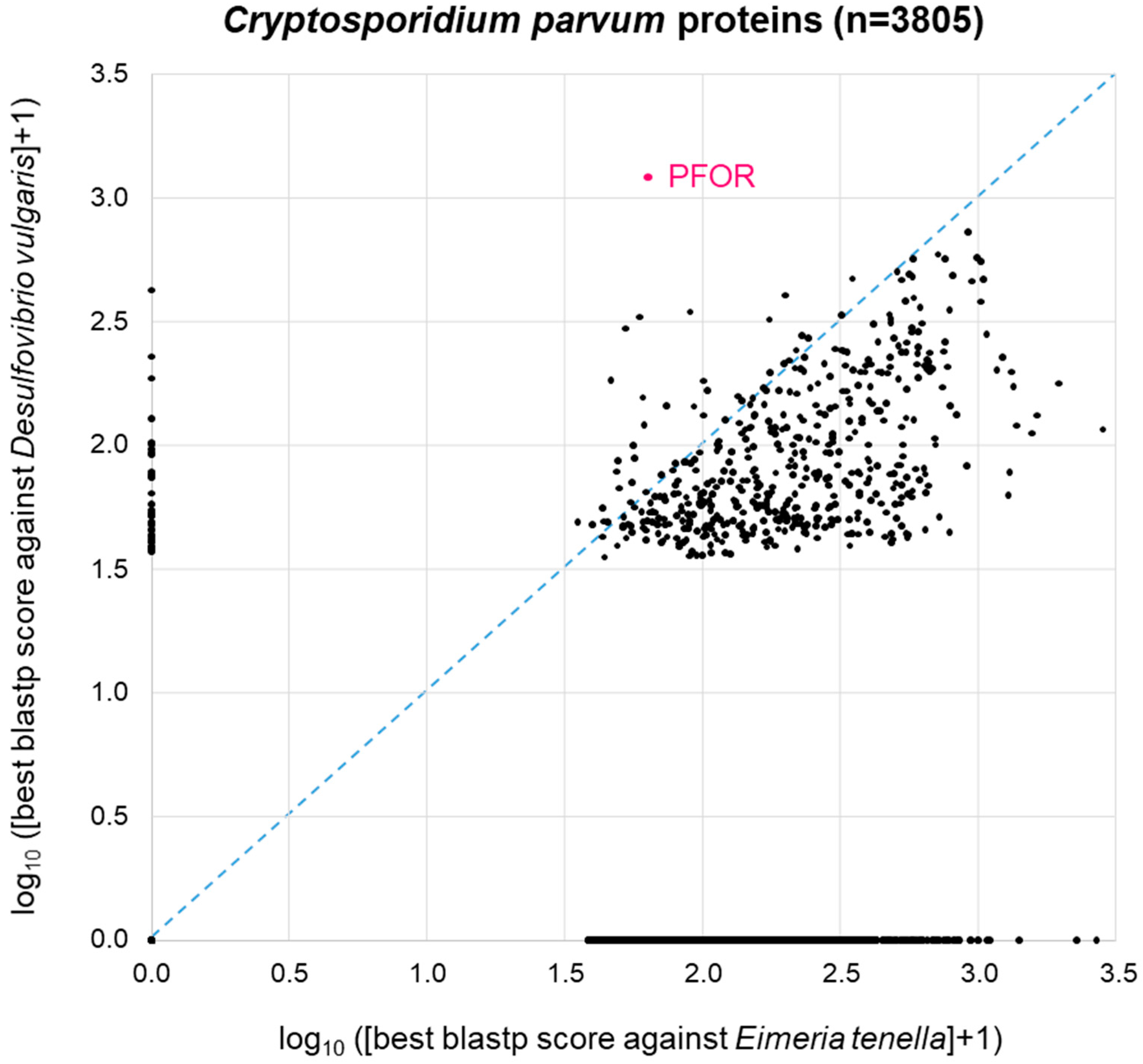
3.4. Proteome-Wide BLAST Surveys
3.5. GO Term Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hucho, F. The Pyruvate Dehydrogenase Multienzyme Complex. Angew. Chem. Int. Ed. 1975, 14, 591–601. [Google Scholar] [CrossRef]
- Chabrière, E.; Charon, M.H.; Volbeda, A.; Pieulle, L.; Hatchikian, E.C.; Fontecilla-Camps, J.C. Crystal structures of the key anaerobic enzyme pyruvate:ferredoxin oxidoreductase, free and in complex with pyruvate. Nat. Struct. Biol. 1999, 6, 182–190. [Google Scholar] [PubMed]
- Charon, M.H.; Volbeda, A.; Chabriere, E.; Pieulle, L.; Fontecilla-Camps, J.C. Structure and electron transfer mechanism of pyruvate:ferredoxin oxidoreductase. Curr. Opin. Struct. Biol. 1999, 9, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.I.; Chen, P.Y.-T.; Drennan, C.L. A structural phylogeny for understanding 2-oxoacid oxidoreductase function. Curr. Opin. Struct. Biol. 2016, 41, 54–61. [Google Scholar] [CrossRef]
- Marczak, R.; Gorrell, T.E.; Müller, M. Hydrogenosomal ferredoxin of the anaerobic protozoon, Tritrichomonas foetus. J. Biol. Chem. 1983, 258, 12427–12433. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.N.; Docampo, R. Mechanism of toxicity of nitro compounds in the chemotherapy of trichomoniasis. Environ. Health Perspect. 1985, 64, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Blamey, J.M.; Adams, M.W. Purification and characterization of pyruvate ferredoxin oxidore-ductase from the hyperthermophilic archaeon Pyrococcus furiosus. Biochim. Biophys. Acta 1993, 1161, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Hrdý, I.; Müller, M. Primary structure and eubacterial relationships of the pyruvate:ferredoxin oxidoreductase of the amitochondriate eukaryote Trichomonas vaginalis. J. Mol. Evol. 1995, 41, 388–396. [Google Scholar] [CrossRef]
- Embley, T.M.; van der Giezen, M.; Horner, D.S.; Dyal, P.L.; Foster, P. Mitochondria and hy-drogenosomes are two forms of the same fundamental organelle. Philos. Trans. R Soc. Lond. B Biol. Sci. 2003, 358, 191–201; discussion 201–192. [Google Scholar] [CrossRef]
- Müller, M. The hydrogenosome. J. Gen. Microbiol. 1993, 139, 2879–2889. [Google Scholar] [CrossRef]
- Kulda, J. Trichomonads, hydrogenosomes and drug resistance. Int. J. Parasitol. 1999, 29, 199–212. [Google Scholar] [CrossRef]
- Benchimol, M. Hydrogenosomes under microscopy. Tissue Cell 2009, 41, 151–168. [Google Scholar] [CrossRef]
- Benchimol, M.; Almeida, J.C.; de Souza, W. Further studies on the organization of the hydrogenosome in Tritrichomonas foetus. Tissue Cell 1996, 28, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Roger, A.J.; Clark, C.G.; Doolittle, W.F. A possible mitochondrial gene in the early-branching amitochondriate protist Trichomonas vaginalis. Proc. Natl. Acad. Sci. USA 1996, 93, 14618–14622. [Google Scholar] [CrossRef] [PubMed]
- Hrdy, I.; Hirt, R.P.; Dolezal, P.; Bardonová, L.; Foster, P.G.; Tachezy, J.; Embley, T.M. Trichomonas hydrogenosomes contain the NADH dehydrogenase module of mitochondrial complex I. Nature 2004, 432, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Makiuchi, T.; Nozaki, T. Highly divergent mitochondrion-related organelles in anaerobic para-sitic protozoa. Biochimie 2014, 100, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Lindmark, D.G.; Müller, M. Hydrogenosome, a Cytoplasmic Organelle of the Anaerobic Flagellate Tritrichomonas foetus, and Its Role in Pyruvate Metabolism. J. Biol. Chem. 1973, 248, 7724–7728. [Google Scholar] [CrossRef] [PubMed]
- Hackstein, J.H.; Akhmanova, A.; Boxma, B.; Harhangi, H.R.; Voncken, F.G. Hydrogenosomes: Eukaryotic adaptations to anaerobic environments. Trends Microbiol. 1999, 7, 441–447. [Google Scholar] [CrossRef]
- Boxma, B.; de Graaf, R.M.; van der Staay, G.W.M.; van Alen, T.A.; Ricard, G.; Gabaldón, T.; van Hoek, A.H.A.M.; der Staay, S.Y.M.-V.; Koopman, W.J.H.; van Hellemond, J.J.; et al. An anaerobic mitochondrion that produces hydrogen. Nature 2005, 434, 74–79. [Google Scholar] [CrossRef]
- Noth, J.; Krawietz, D.; Hemschemeier, A.; Happe, T. Pyruvate:ferredoxin oxidoreductase is cou-pled to light-independent hydrogen production in Chlamydomonas reinhardtii. J. Biol. Chem. 2013, 288, 4368–4377. [Google Scholar] [CrossRef]
- van Lis, R.; Baffert, C.; Couté, Y.; Nitschke, W.; Atteia, A. Chlamydomonas reinhardtii Chloroplasts Contain a Homodimeric Pyruvate:Ferredoxin Oxidoreductase That Functions with FDX1. Plant Physiol. 2012, 161, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Rotte, C.; Stejskal, F.; Zhu, G.; Keithly, J.S.; Martin, W. Pyruvate:NADP Oxidoreductase from the Mitochondrion of Euglena gracilis and from the Apicomplexan Cryptosporidium parvum: A Biochemical Relic Linking Pyruvate Metabolism in Mitochondriate and Amitochondriate Protists. Mol. Biol. Evol. 2001, 18, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Horner, D.S.; Hirt, R.P.; Embley, T.M. A single eubacterial origin of eukaryotic pyruvate: Ferre-doxin oxidoreductase genes: Implications for the evolution of anaerobic eukaryotes. Mol. Biol. Evol. 1999, 16, 1280–1291. [Google Scholar] [CrossRef]
- Juhas, M. Horizontal gene transfer in human pathogens. Crit. Rev. Microbiol. 2015, 41, 101–108. [Google Scholar] [CrossRef]
- Deng, Y.; Xu, H.; Su, Y.; Liu, S.; Xu, L.; Guo, Z.; Wu, J.; Cheng, C.; Feng, J. Horizontal gene transfer contributes to virulence and antibiotic resistance of Vibrio harveyi 345 based on complete genome sequence analysis. BMC Genom. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Wyres, K.L.; Wick, R.R.; Judd, L.M.; Froumine, R.; Tokolyi, A.; Gorrie, C.L.; Lam, M.M.C.; Duchêne, S.; Jenney, A.; Holt, K.E. Distinct evolutionary dynamics of horizontal gene transfer in drug resistant and virulent clones of Klebsiella pneumoniae. PLoS Genet. 2019, 15, e1008114. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef]
- Lacroix, B.; Citovsky, V. Transfer of DNA from Bacteria to Eukaryotes. mBio 2016, 7, e00863-16. [Google Scholar] [CrossRef]
- Lacroix, B.; Citovsky, V. Beyond Agrobacterium-Mediated Transformation: Horizontal Gene Transfer from Bacteria to Eukaryotes. Curr. Top Microbiol. Immunol. 2018, 418, 443–462. [Google Scholar]
- Ricard, G.; McEwan, N.R.; Dutilh, B.E.; Jouany, J.P.; Macheboeuf, D.; Mitsumori, M.; McIntosh, F.M.; Michalowski, T.; Nagamine, T.; Nelson, N.; et al. Horizontal gene transfer from Bacteria to rumen Ciliates indi-cates adaptation to their anaerobic, carbohydrates-rich environment. BMC Genom. 2006, 7, 22. [Google Scholar] [CrossRef]
- Eme, L.; Gentekaki, E.; Curtis, B.; Archibald, J.M.; Roger, A.J. Lateral Gene Transfer in the Adaptation of the Anaerobic Parasite Blastocystis to the Gut. Curr. Biol. 2017, 27, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. You are what you eat: A gene transfer ratchet could account for bacterial genes in eukaryotic nuclear genomes. Trends Genet. 1998, 14, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Field, J.; Rosenthal, B.; Samuelson, J. Early lateral transfer of genes encoding malic enzyme, ace-tyl-CoA synthetase and alcohol dehydrogenases from anaerobic prokaryotes to Entamoeba histolytica. Mol. Microbiol. 2000, 38, 446–455. [Google Scholar] [CrossRef]
- Hedges, S.B.; Chen, H.; Kumar, S.; Wang, D.Y.; Thompson, A.S.; Watanabe, H. A genomic time-scale for the origin of eukaryotes. BMC Evol. Biol. 2001, 1, 4. [Google Scholar] [CrossRef]
- Keeling, P.J. Functional and ecological impacts of horizontal gene transfer in eukaryotes. Curr. Opin. Genet. Dev. 2009, 19, 613–619. [Google Scholar] [CrossRef]
- Schönknecht, G.; Chen, W.H.; Ternes, C.M.; Barbier, G.G.; Shrestha, R.P.; Stanke, M.; Bräutigam, A.; Baker, B.J.; Banfield, J.F.; Garavito, R.M.; et al. Gene transfer from bacteria and archaea facilitated evolution of an extremophilic eukaryote. Science 2013, 339, 1207–1210. [Google Scholar] [CrossRef]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2017, 16, 67–79. [Google Scholar] [CrossRef]
- Sibbald, S.J.; Eme, L.; Archibald, J.M.; Roger, A.J. Lateral Gene Transfer Mechanisms and Pan-genomes in Eukaryotes. Trends Parasitol. 2020, 36, 927–941. [Google Scholar] [CrossRef]
- Van Etten, J.; Bhattacharya, D. Horizontal Gene Transfer in Eukaryotes: Not if, but How Much? Trends Genet. 2020, 36, 915–925. [Google Scholar] [CrossRef]
- Nixon, J.E.; Wang, A.; Field, J.; Morrison, H.G.; McArthur, A.G.; Sogin, M.L.; Loftus, B.J.; Samuelson, J. Evidence for lateral transfer of genes encoding ferredoxins, nitroreductases, NADH oxidase, and alcohol dehydrogenase 3 from anaerobic prokaryotes to Giardia lamblia and Entamoeba histolytica. Eukaryot Cell 2002, 1, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Alsmark, U.C.; Sicheritz-Ponten, T.; Foster, P.G.; Hirt, R.P.; Embley, T.M. Horizontal gene trans-fer in eukaryotic parasites: A case study of Entamoeba histolytica and Trichomonas vaginalis. Methods Mol. Biol. 2009, 532, 489–500. [Google Scholar] [PubMed]
- Wittlin, S.; Mäser, P. From Magic Bullet to Magic Bomb: Reductive Bioactivation of Antiparasitic Agents. ACS Infect. Dis. 2021, 7, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Upcroft, P.; Upcroft, J.A. Drug Targets and Mechanisms of Resistance in the Anaerobic Protozoa. Clin. Microbiol. Rev. 2001, 14, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Banerjee, S.; Cui, J.; Schwartz, A.; Ghosh, S.K.; Samuelson, J. Giardia, Entamoeba, and Tri-chomonas enzymes activate metronidazole (nitroreductases) and inactivate metronidazole (nitroimidazole reductases). Antimicrob. Agents Chemother. 2009, 53, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Lamp, K.C.; Freeman, C.D.; Klutman, N.E.; Lacy, M.K. Pharmacokinetics and pharmacody-namics of the nitroimidazole antimicrobials. Clin. Pharmacokinet. 1999, 36, 353–373. [Google Scholar] [CrossRef]
- A Dingsdag, S.; Hunter, N. Metronidazole: An update on metabolism, structure–cytotoxicity and resistance mechanisms. J. Antimicrob. Chemother. 2018, 73, 265–279. [Google Scholar] [CrossRef]
- Dan, M.; Wang, A.L.; Wang, C.C. Inhibition of pyruvate-ferredoxin oxidoreductase gene expression in Giardia lamblia by a virus-mediated hammerhead ribozyme. Mol. Microbiol. 2000, 36, 447–456. [Google Scholar] [CrossRef]
- Leitsch, D.; Burgess, A.G.; Dunn, L.A.; Krauer, K.G.; Tan, K.; Duchêne, M.; Upcroft, P.; Eckmann, L.; Upcroft, J.A. Pyruvate:ferredoxin oxidoreductase and thioredoxin reductase are involved in 5-nitroimidazole activation while flavin metabolism is linked to 5-nitroimidazole resistance in Giardia lamblia. J. Antimicrob. Chemother. 2011, 66, 1756–1765. [Google Scholar] [CrossRef]
- Ansell, B.R.; Baker, L.; Emery, S.J.; McConville, M.J.; Svärd, S.G.; Gasser, R.B.; Jex, A.R. Tran-scriptomics Indicates Active and Passive Metronidazole Resistance Mechanisms in Three Seminal Giardia Lines. Front. Microbiol. 2017, 8, 398. [Google Scholar] [CrossRef]
- Müller, J.; Braga, S.; Heller, M.; Müller, N. Resistance formation to nitro drugs in Giardia lamblia: No common markers identified by comparative proteomics. Int. J. Parasitol. Drugs Drug Resist. 2019, 9, 112–119. [Google Scholar] [CrossRef]
- Lopes-Oliveira, L.A.P.; Fantinatti, M.; Da-Cruz, A.M. In vitro-induction of metronida-zole-resistant Giardia duodenalis is not associated with nucleotide alterations in the genes involved in pro-drug activation. Mem. Inst. Oswaldo Cruz 2020, 115, e200303. [Google Scholar] [CrossRef]
- Rasoloson, D.; Vanacova, S.; Tomkova, E.; Razga, J.; Hrdy, I.; Tachezy, J.; Kulda, J. Mechanisms of in vitro development of resistance to metronidazole in Trichomonas vaginalis. Microbiology 2002, 148, 2467–2477. [Google Scholar] [CrossRef]
- Hrdý, I.; Cammack, R.; Stopka, P.; Kulda, J.; Tachezy, J. Alternative pathway of metronidazole ac-tivation in Trichomonas vaginalis hydrogenosomes. Antimicrob. Agents Chemother. 2005, 49, 5033–5036. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing evolutionary trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium; Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; et al. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef]
- Saavedra, E.; Encalada, R.; Pineda, E.; Jasso-Chávez, R.; Moreno-Sánchez, R. Glycolysis in Entamoeba histolytica. Biochemical characterization of recombinant glycolytic enzymes and flux control analysis. FEBS J. 2005, 272, 1767–1783. [Google Scholar] [CrossRef] [PubMed]
- Higuera, A.; Villamizar, X.; Herrera, G.; Giraldo, J.C.; Vasquez, A.L.; Urbano, P.; Villalobos, O.; Tovar, C.; Ramírez, J.D. Molecular detection and genotyping of intestinal protozoa from different biogeographical regions of Colombia. PeerJ 2020, 8, e8554. [Google Scholar] [CrossRef] [PubMed]
- Benchimol, M.; Rosa, I.d.A.; Fontes, R.d.S.; Dias, J.B. Trichomonas adhere and phagocytose sperm cells: Adhesion seems to be a prominent stage during interaction. Parasitol. Res. 2008, 102, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Chapwanya, A.; Usman, A.Y.; Irons, P.C. Comparative aspects of immunity and vaccination in human and bovine trichomoniasis: A review. Trop. Anim. Health Prod. 2015, 48, 1–7. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Species | PFOR | GAPDH | TUBa | RPB1 |
|---|---|---|---|---|---|
| Stramenopiles | Blastocystis hominis | 0.89 | 0.91 | 0.82 | 0.59 |
| Phytophthora infestans | 0.22 | 0.97 | 0.99 | 0.94 | |
| Albugo laibachii | 0.01 | 0.95 | 0.97 | 0.92 | |
| Ectocarpus siliculosus | 0.01 | 0.93 | 0.97 | 0.85 | |
| Alveolates | Plasmodium falciparum | 0.01 | 0.92 | 1.00 | 1.00 |
| Cryptosporidium parvum | 0.98 | 0.97 | 0.97 | 0.91 | |
| Toxoplasma gondii | 0.01 | 0.94 | 0.99 | 0.89 | |
| Tetrahymena thermophila | 0.01 | 0.89 | 0.99 | 0.59 | |
| Rhizaria | Plasmodiophora brassicae | 0.01 | 0.84 | 1.00 | 0.87 |
| Reticulomyxa filosa | 0.01 | 0.57 | 0.93 | 0.73 | |
| Euglenozoa | Trypanosoma brucei | 0.01 | 0.95 | 0.98 | 0.72 |
| Phytomonas serpens | 0.00 | 0.89 | 0.97 | 0.69 | |
| Naegleria gruberi | 0.01 | 0.91 | 0.99 | 0.85 | |
| Amoebozoa | Entamoeba invadens | 0.96 | 0.92 | 0.79 | 0.66 |
| Entamoeba histolytica | 0.99 | 0.97 | 0.47 | 0.67 | |
| Dictyostelium purpureum | 0.01 | 0.93 | 0.82 | 0.93 | |
| Dictyostelium discoideum | 0.01 | 0.92 | 0.83 | 0.93 | |
| Metamonada | Giardia lamblia | 0.80 | 0.91 | 0.98 | 0.62 |
| Spironucleus salmonicida | 0.78 | 0.83 | 0.92 | 0.58 | |
| Trichomonas vaginalis | 0.88 | 0.70 | 0.97 | 0.70 | |
| Animalia | Caenorhabditis elegans | 0.01 | 0.98 | 0.97 | 0.93 |
| Schistosoma mansoni | 0.01 | 0.93 | 1.00 | 0.82 | |
| Drosophila melanogaster | 0.01 | 0.95 | 1.00 | 0.93 | |
| Fungi | Sporisorium reilianum | 0.16 | 0.96 | 0.87 | 0.86 |
| Saccharomyces cerevisiae | 0.11 | 0.94 | 0.89 | 0.88 | |
| Aspergillus fumigatus | 0.17 | 0.95 | 0.93 | 0.88 | |
| Embryophyta | Asparagus officinalis | 0.01 | 0.68 | 1.00 | 0.97 |
| Oryza sativa | 0.01 | 0.97 | 0.98 | 0.92 | |
| Pistacia vera | 0.01 | 0.97 | 1.00 | 0.94 | |
| Chlorophyta | Chlamydomonas reinhardtii | 0.98 | 0.92 | 0.99 | 0.86 |
| Ostreococcus lucimarinus | 0.01 | 0.68 | 0.98 | 0.87 | |
| Chlorella pyrenoidosa | 0.90 | 0.96 | 1.00 | 0.19 | |
| Rhodophyta | Chondrus crispus | 0.01 | 0.94 | 0.86 | 0.80 |
| Galdieria sulphuraria | 0.01 | 0.95 | 0.94 | 0.52 |
| Biological Process | Fold Enrichment | p-Value | FDR |
|---|---|---|---|
| Pyruvate metabolic process | 40 | 7.75 × 10−5 | 3.35 × 10−2 |
| Monocarboxylic acid metabolic process | 18 | 8.29 × 10−5 | 3.07 × 10−2 |
| Carboxylic acid metabolic process | 11 | 9.09 × 10−5 | 2.94 × 10−2 |
| Oxoacid metabolic process | 15 | 4.39 × 10−7 | 1.14 × 10−3 |
| Organic acid metabolic process | 15 | 4.63 × 10−7 | 5.99 × 10−4 |
| Small molecule metabolic process | 7.1 | 5.03 × 10−5 | 2.61 × 10−2 |
| Generation of precursor metabolites and energy | 24 | 2.94 × 10−5 | 2.54 × 10−2 |
| Carbohydrate metabolic process | 13 | 4.61 × 10−5 | 2.99 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duwor, S.; Brites, D.; Mäser, P. Phylogenetic Analysis of Pyruvate-Ferredoxin Oxidoreductase, a Redox Enzyme Involved in the Pharmacological Activation of Nitro-Based Prodrugs in Bacteria and Protozoa. Biology 2024, 13, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13030178
Duwor S, Brites D, Mäser P. Phylogenetic Analysis of Pyruvate-Ferredoxin Oxidoreductase, a Redox Enzyme Involved in the Pharmacological Activation of Nitro-Based Prodrugs in Bacteria and Protozoa. Biology. 2024; 13(3):178. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13030178
Chicago/Turabian StyleDuwor, Seth, Daniela Brites, and Pascal Mäser. 2024. "Phylogenetic Analysis of Pyruvate-Ferredoxin Oxidoreductase, a Redox Enzyme Involved in the Pharmacological Activation of Nitro-Based Prodrugs in Bacteria and Protozoa" Biology 13, no. 3: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13030178