
Anti-Obesity Effect of Daidzein Derived from Pachyrhizus erosus (L.) Urb. Extract via PPAR Pathway in MDI-Induced 3T3-L1 Cell Line

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
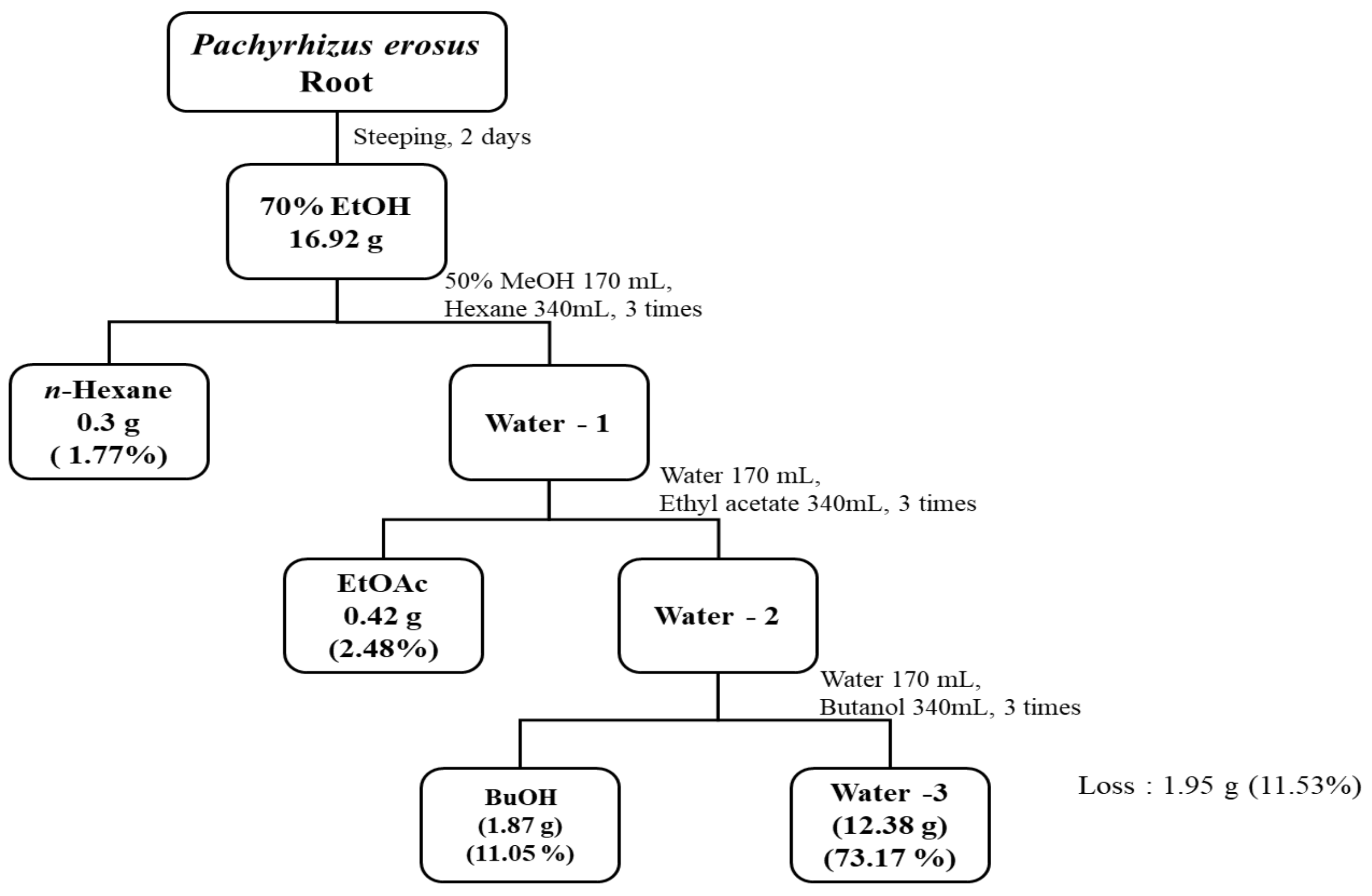
2.2. Preparation of Extracts and Solvent Fractions
2.3. Antioxidants Activity Assay
2.3.1. DPPH Free Radical Activity
2.3.2. ABTS Radical Activity
2.4. Total Polyphenol and Flavonoid Contents
2.4.1. Total Polyphenol Contents (TPC)
2.4.2. Total Flavonoid Contents (TFC)
2.5. High-Performance Liquid Chromatography with Diode-Array Detection (HPLC–DAD) Analysis
2.6. Quantitative Analysis of Active Ingredient Using LC-MS/MS
2.7. Anti-Obesity Enzyme Inhibition Assay
2.7.1. α-Glucosidase Inhibition Assay
2.7.2. Lipase Inhibition Assay
2.8. Inhibition of Adipogenesis and Fatty Acid Synthesis in 3T3-L1 Cells
2.8.1. Cell Culture and Adipocyte Differentiation
2.8.2. Cell Viability Assay
2.8.3. Oil Red O Staining
2.8.4. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Extraction Yield of P. erosus Root Extracts
3.2. Antioxidant Activity of P. erosus Root Extracts
3.3. Total Polyphenol and Total Flavonoid Contents
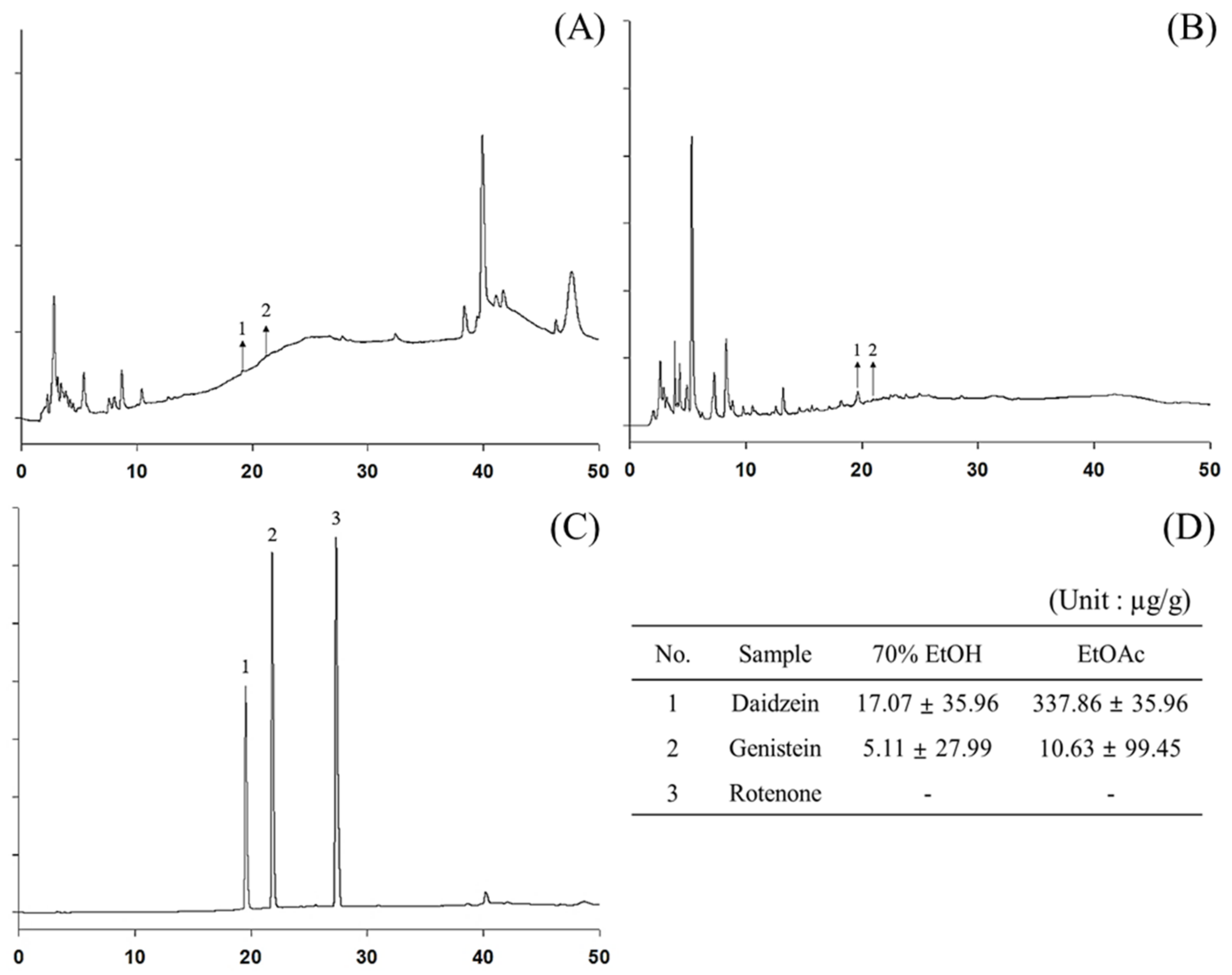
3.4. Analysis of Polyphenol Content Using HPLC and LC-MS/MS
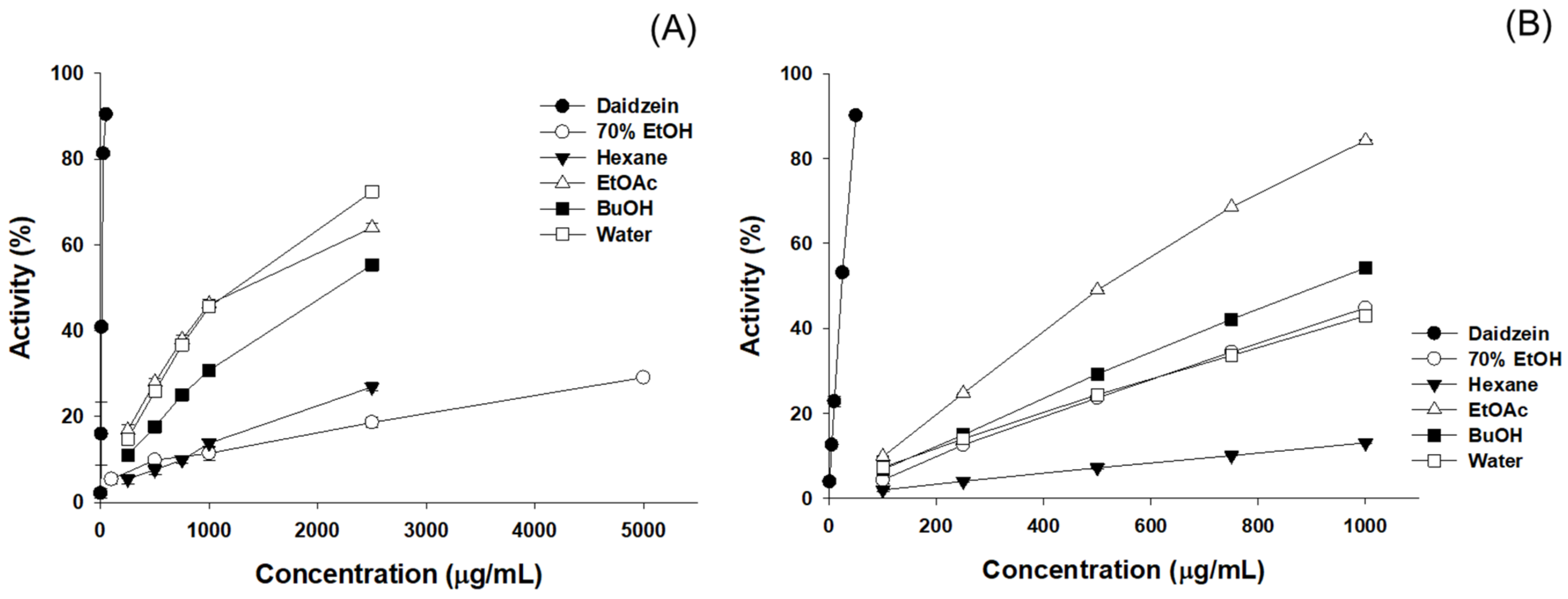
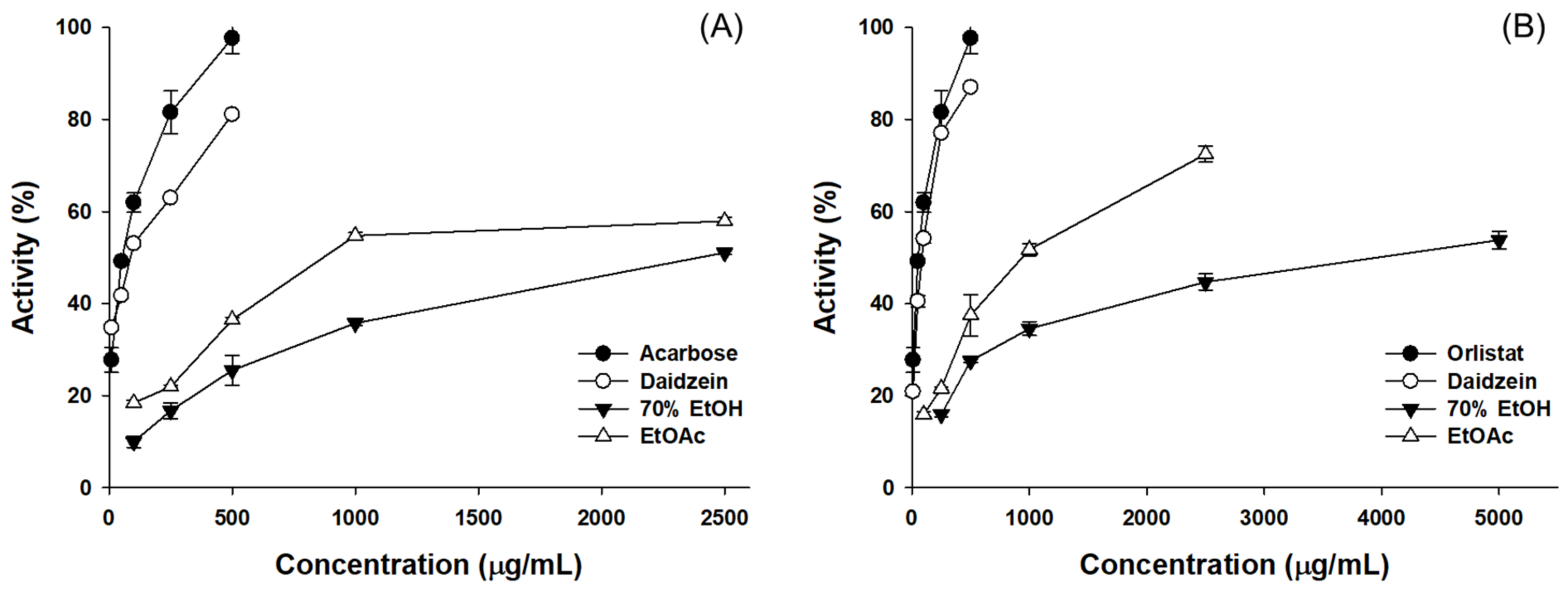
3.5. Results of the Anti-Obesity Enzyme Inhibition Activity
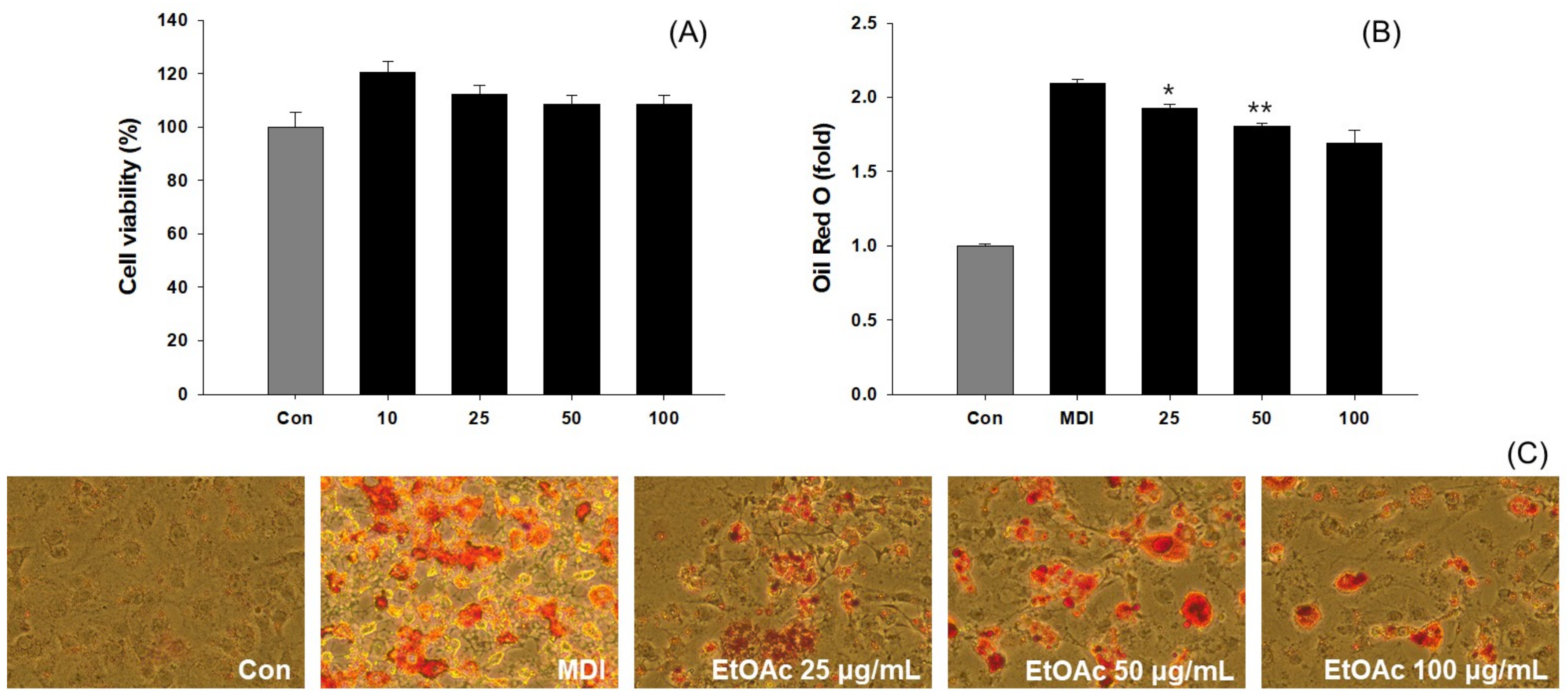
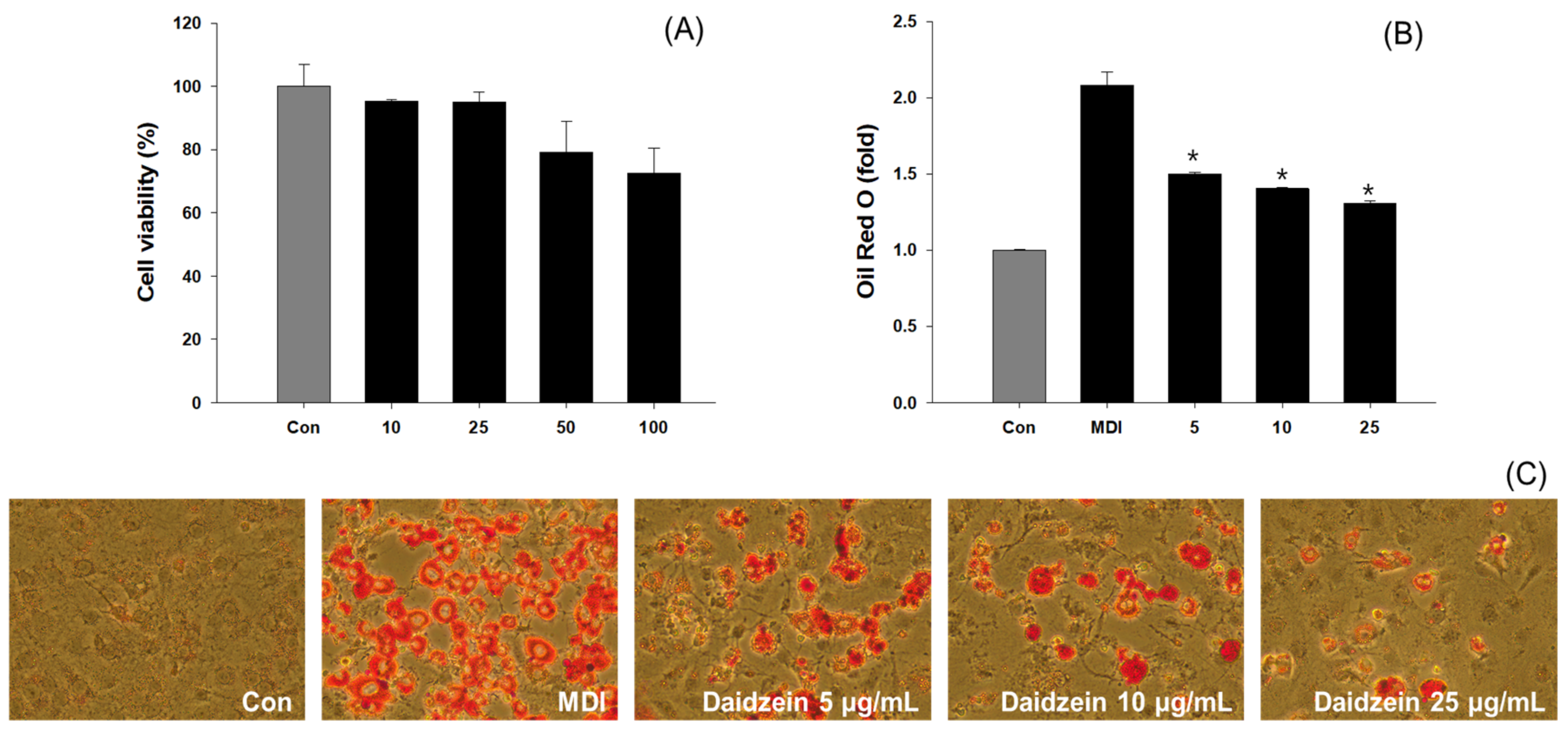
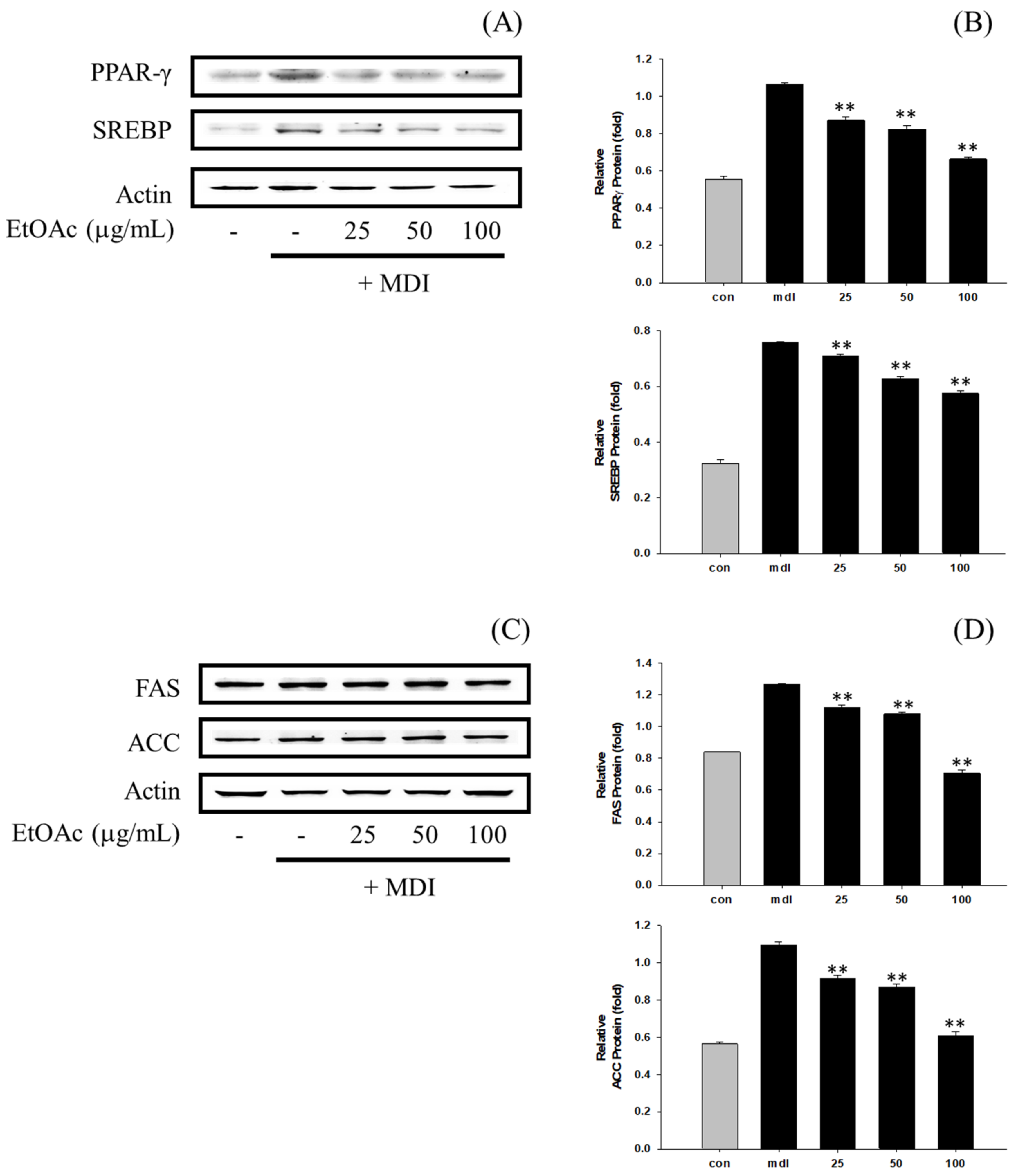
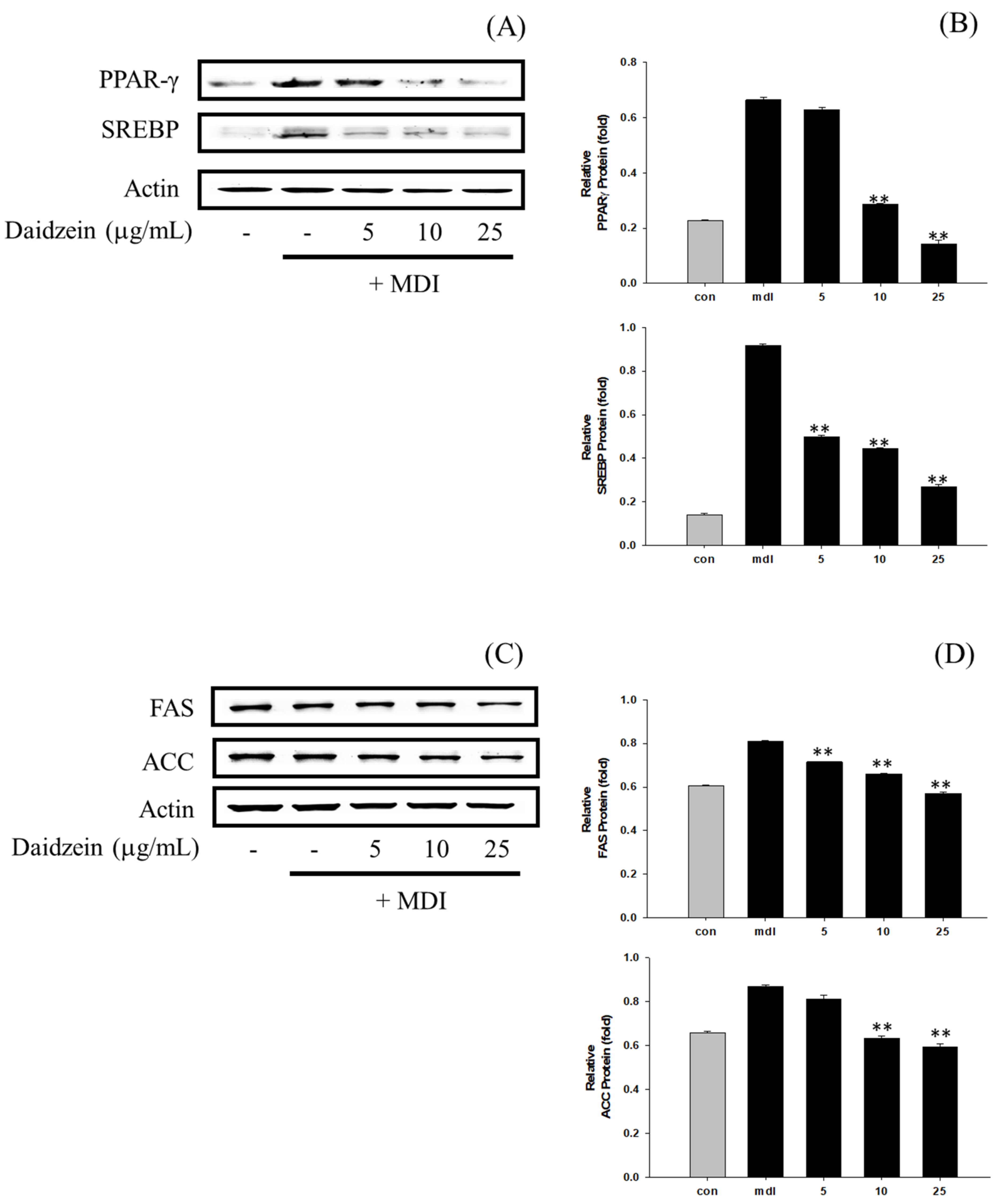
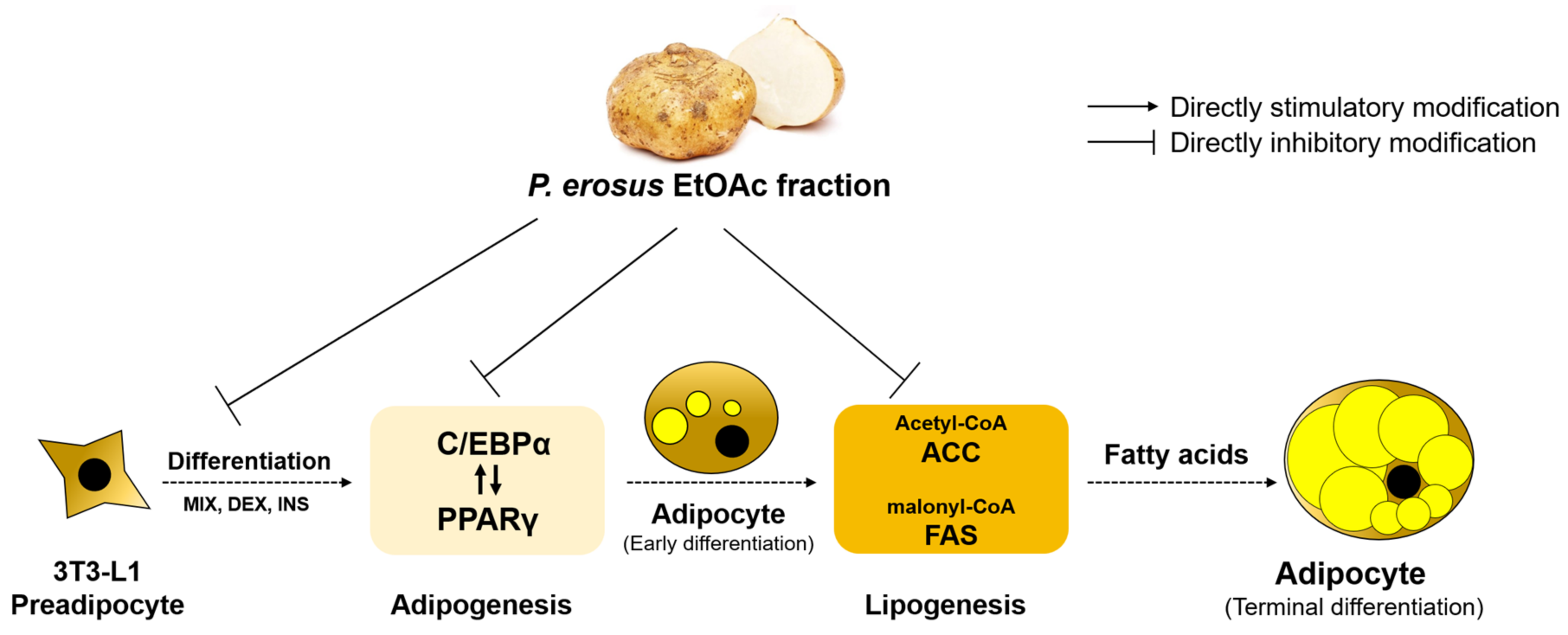
3.6. Inhibition of Adipogenesis and Fatty Acid Synthesis in 3T3-L1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fruh, S.M. Obesity: Risk factors, complications, and strategies for sustainable long-term weight management. J. Am. Assoc. Nurse. Pract. 2017, 29, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Ramos-de-la-Peña, A.M.; Renard, C.M.; Wicker, L.; Montañez, J.; de la Luz Reyes-Vega, M.; Voget, C.; Contreras-Esquivel, J.C. Enzymatic liquefaction of Jicama (Pachyrhizus erosus) tuberous roots and characterization of the cell walls after processing. LWT-Food Sci. Technol. 2012, 49, 257–262. [Google Scholar] [CrossRef]
- Baranowska-Bik, A.; Bik, W. The association of obesity with autoimmune thyroiditis and thyroid function-possible mechanisms of Bilateral Interaction. Int. J. Endocrinol. 2020, 2020, 8894792. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Urbán, C.; Cerón, J.J.; Tecles, F.; Allende, A.; Tomás-Barberán, F.A.; Espín, J.C. Effect of low inulin doses with different polymerisation degree on lipid metabolism, mineral absorption, and intestinal microbiota in rats with fat-supplemented diet. Food Chem. 2009, 113, 1058–1065. [Google Scholar] [CrossRef]
- Nam, Y.R.; Won, S.B.; Chung, Y.S.; Kwak, C.S.; Kwon, Y.H. Inhibitory effects of Doenjang, Korean traditional fermented soybean paste, on oxidative stress and inflammation in adipose tissue of mice fed a high-fat diet. Nutr. Res. Pract. 2015, 9, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, G.F.; Santos, I.B.; de Bem, G.F.; Cordeiro, V.S.C.; da Costa, C.A.; de Carvalho, L.C.R.M.; Ognibene, D.T.; Resende, A.C.; de Moura, R.S. The beneficial effect of anthocyanidin-rich Vitis vinifera L. grape skin extract on metabolic changes induced by high-fat diet in mice involves antiinflammatory and antioxidant actions. Phytother. Res. 2017, 31, 1621–1632. [Google Scholar] [CrossRef] [PubMed]
- Charradi, K.; Elkahoui, S.; Limam, F.; Aouani, E. High-fat diet induced an oxidative stress in white adipose tissue and disturbed plasma transition metals in rat: Prevention by grape seed and skin extract. J. Physiol. Sci. 2013, 63, 445–455. [Google Scholar] [CrossRef]
- Xu, S.P.; Mao, X.Y.; Ren, F.Z.; Che, H.L. Attenuating effect of casein glycomacropeptide on proliferation, differentiation, and lipid accumulation of in vitro Sprague-Dawley rat preadipocytes. J. Dairy Sci. 2011, 94, 676–683. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Cires, M.J.; Gotteland, M. Quercetin and epigallocatechin gallate in the prevention and treatment of obesity: From molecular to clinical studies. J. Med. Food 2019, 22, 753–770. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Wen, X.; Zhang, B.; Wu, B.; Xiao, H.; Li, Z.; Li, R.; Xu, X.; Li, T. Signaling pathways in obesity: Mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.H.; Seo, M.J.; Choi, H.S.; Lee, B.Y. Pycnogenol® inhibits lipid accumulation in 3T3-L1 adipocytes with the modulation of reactive oxygen species (ROS) production associated with antioxidant enzyme responses. Phytother. Res. 2012, 26, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Olejnik, A.; Szwajgier, D.; Olkowicz, M. Inhibitory activity of chokeberry, bilberry, raspberry and cranberry polyphenol-rich extract towards adipogenesis and oxidative stress in differentiated 3T3-L1 adipose cells. PLoS ONE 2017, 12, e0188583. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D.; Burns, K.A.; Hosick, P.A.; McBeth, L.; Nestor-Kalinoski, A.; Drummond, H.A.; AlAmodi, A.A.; Hankins, M.W.; Vanden Heuvel, J.P.; Stec, D.E. Biliverdin reductase A attenuates hepatic steatosis by inhibition of glycogen synthase kinase (GSK) 3β phosphorylation of serine 73 of peroxisome proliferator-activated receptor (PPAR) α. J. Biol. Chem. 2016, 291, 25179–25191. [Google Scholar] [CrossRef]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.; Weickert, M.O. The health benefits of dietary fiber. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- Sabarathinam, S. A glycemic diet improves the understanding of glycemic control in diabetes patients during their follow-up. Future Sci. OA 2023, 9, FSO843. [Google Scholar] [CrossRef]
- Suryana, A.L.; Kristanto, A.A.; Putri, F. Effect consumption of Papaya and Jicama juice to total cholesterol levels in hypercholesterolemia patients. Int. J. Pharma. Bio-Med. Sci. 2023, 3, 152–155. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Mungmai, L.; Kanokwattananon, C.; Thakang, S.; Nakkrathok, A.; Srisuksomwong, P.; Tanamatayarat, P. Physicochemical properties, antioxidant and anti-tyrosinase activities of Durio zibethinus Murray and value added for cosmetic product formulation. Cosmetics 2023, 10, 87. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Ahn, G.Y.; Bae, S.H. Strategies for the safe use of nonsteroidal anti-inflammatory drugs. J. Korean Med. Assoc. 2018, 61, 367–375. [Google Scholar] [CrossRef]
- Sato, K.; Hiraga, Y.; Yamaguchi, Y.; Sakaki, S.; Takenaka, H. Anti-melanogenic and anti-oxidative effects of Nostoc verrucosum (ashitsuki) extracts. Cosmetics 2023, 10, 30. [Google Scholar] [CrossRef]
- Kurzawa, M.; Wilczyńska, E.; Brudzyńska, P.; Sionkowska, A. Total phenolic content, antioxidant capacity and UV Radiation Protection Properties of Marigold (Calendula officinalis), carrot (Daucus carota), tomato (Solanum lycopersicum) and hop (Humulus lupulus) Extracts. Cosmetics 2022, 9, 134. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Rungsang, T.; Srivilai, J.; Rakasawapokin, P.; Rakasawapokin, P.; Mungmai, L.; Saesue, K.; Aoonboontum, P.; Plukham, N.; Siriwipanan, P.; Chaichanathawikit, P.; et al. Assessment of antioxidant, anti-lipid peroxidation, antiglycation, anti-inflammatory and anti-tyrosinase properties of Dendrobium sulcatum Lindl. Cosmetics 2023, 10, 43. [Google Scholar] [CrossRef]
- Phosri, S.; Kiattisin, K.; Intharuksa, A.; Janon, R.; Na Nongkhai, T.; Theansungnoen, T. Anti-aging, anti-acne, and cytotoxic activities of Houttuynia cordata extracts and phytochemicals analysis by LC-MS/MS. Cosmetics 2022, 9, 136. [Google Scholar] [CrossRef]
- Kim, J.M.; Jung, I.A.; Kim, J.M.; Choi, M.H.; Yang, J.H. Anti-Inflammatory Effect of Cinnamomum japonicum Siebold’s Leaf through the Inhibition of p38/JNK/AP-1 Signaling. Pharmaceuticals 2023, 16, 1402. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, S.H.; Yoon, N.Y.; Jung, W.K.; Jeon, Y.J.; Kim, S.K.; Lee, M.S.; Kim, Y.M. α-Glucosidase- and α-amylase-inhibitory activities of phlorotannins from Eisenia bicyclis. J. Sci. Food Agric. 2012, 92, 2084–2090. [Google Scholar] [CrossRef]
- Gupta, R.; Rathi, P.; Gupta, N.; Bradoo, S. Lipase assays for conventional and molecular screening: An overview. Biotechnol. Appl. Biochem. 2003, 37, 63–71. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.J.; Kim, C.; Jung, H.; Kim, Y.O.; Ju, J.Y.; Shin, C.S. Development of lipase inhibitors from various derivatives of monascus pigment produced by Monascus fermentation. Food Chem. 2007, 101, 357–364. [Google Scholar] [CrossRef]
- Tafuri, S.R. Troglitazone enhances differentiation, basal glucose uptake, and Glut1 protein levels in 3T3-L1 adipocytes. Endocrinology 1996, 137, 4706–4712. [Google Scholar] [CrossRef] [PubMed]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Zacarias, J.L.; Castro-Munozledo, F.; Kuri-Harcuch, W. Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429. [Google Scholar]
- Lukitaningsih, E.; Bahi, M.; Holzgrabe, U. Tyrosinase inhibition type of isolated compounds obtained from Pachyrhizus erosus. Aceh Int. J. Sci. Technol. 2013, 2, 98–102. [Google Scholar] [CrossRef]
- Estrella-Parra, E.A.; Gomez-Verjan, J.C.; González-Sánchez, I.; Vázquez-Martínez, E.R.; Vergara-Castañeda, E.; Cerbón, M.A.; Alavez-Solano, D.; Reyes-Chilpa, R. Rotenone isolated from Pachyrhizus erosus displays cytotoxicity and genotoxicity in K562 cells. Nat. Prod. Res. 2014, 28, 1780–1785. [Google Scholar] [CrossRef]
- Sethi, J.K.; Vidal-Puig, A.J. Thematic review series: Adipocyte biology. Adipose tissue function and plasticity orchestrate nutritional adaptation. J. Lipid Res. 2007, 48, 1253–1262. [Google Scholar] [CrossRef]
- Ràfols, M.E. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef]
- Jaiswal, V.; Chauhan, S.; Lee, H.J. The bioactivity and phytochemicals of Pachyrhizus erosus (L.) Urb.: A multifunctional underutilized crop plant. Antioxidants 2021, 11, 58. [Google Scholar] [CrossRef]
- Fu, P.K.; Wang, P.Y. Toxic leukoencephalopathy due to yam bean seeds poisoning. Neurologist 2012, 18, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Levaj, B.; Pelaić, Z.; Galić, K.; Kurek, M.; Ščetar, M.; Poljak, M.; Hunjek, D.D.; Pedisić, S.; Balbino, S.; Čošić, Z.; et al. Maintaining the quality and safety of fresh-cut potatoes (Solanum tuberosum): Overview of recent findings and approaches. Agronomy 2023, 13, 2002. [Google Scholar] [CrossRef]
- Santoso, P.; Maliza, R.; Insani, S.J.; Fadhila, Q.; Rahayu, R. Preventive effect of Jicama (Pachyrhizus erosus) fiber against diabetes development in mice fed with high-fat diet. J. Appl. Pharm. Sci. 2021, 11, 137–143. [Google Scholar]
- Wang, S.; Wang, Y.; Pan, M.H.; Ho, C.T. Anti-obesity molecular mechanism of soy isoflavones: Weaving the way to new therapeutic routes. Food Funct. 2017, 8, 3831–3846. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Shou, Q.; Lu, Y.; Wang, G.; Qiu, J.; Wang, J.; He, L.; Chen, J.; Jiao, J.; Zhang, Y. Arachidonic acid sex-dependently affects obesity through linking gut microbiota-driven inflammation to hypothalamus-adipose-liver axis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2715–2726. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Rong, Y.; Bao, L.; Nie, B.; Ren, G.; Zheng, C.; Amin, R.; Arnold, R.D.; Jeganathan, R.B.; Huggins, K.W. Suppression of adipocyte differentiation and lipid accumulation by stearidonic acid (SDA) in 3T3-L1 cells. Lipids Health Dis. 2017, 16, 181. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Cha, J.Y.; Seok, J.W.; Choi, Y.; Yoon, B.K.; Choi, H.; Yu, J.H.; Song, S.J.; Kim, A.; Lee, H.; et al. Dexras1 links glucocorticoids to insulin-like growth factor-1 signaling in adipogenesis. Sci. Rep. 2016, 6, 28648. [Google Scholar] [CrossRef]
- Han, S.I.; Komatsu, Y.; Murayama, A.; Steffensen, K.R.; Nakagawa, Y.; Nakajima, Y.; Suzuki, M.; Oie, S.; Parini, P.; Vedin, L.L.; et al. Estrogen receptor ligands ameliorate fatty liver through a nonclassical estrogen receptor/Liver X receptor pathway in mice. Hepatology 2014, 59, 1791–1802. [Google Scholar] [CrossRef]
- Wu, C.; Fang, S.; Zhang, H.; Li, X.; Du, Y.; Zhang, Y.; Lin, X.; Wang, L.; Ma, X.; Xue, Y.; et al. Long noncoding RNA XIST regulates brown preadipocytes differentiation and combats high-fat diet induced obesity by targeting C/EBPα. Mol. Med. 2022, 28, 6. [Google Scholar] [CrossRef]
- He, Y.F.; Liu, F.Y.; Zhang, W.X. Tangeritin inhibits adipogenesis by down-regulating C/EBPα, C/EBPβ, and PPARγ expression in 3T3-L1 fat cells. Genet. Mol. Res. 2015, 14, 13642–13648. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part II: PPAR-β/δ and PPAR-γ. Future Cardiol. 2017, 13, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Rayner, M.L.D.; Grillo, A.; Williams, G.R.; Tawfik, E.; Zhang, T.; Volitaki, C.; Craig, D.Q.M.; Healy, J.; Phillips, J.B. Controlled local release of PPARγ agonists from biomaterials to treat peripheral nerve injury. J. Neural Eng. 2020, 17, 046030. [Google Scholar] [CrossRef] [PubMed]
- Baldini, F.; Fabbri, R.; Eberhagen, C.; Voci, A.; Portincasa, P.; Zischka, H.; Vergani, L. Adipocyte hypertrophy parallels alterations of mitochondrial status in a cell model for adipose tissue dysfunction in obesity. Life Sci. 2021, 265, 118812. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Bok, S.H.; Kim, M.H.; Lee, K.S.; Park, D.H. Pachyrhizus erosus inhibits adipogenesis via the Leptin-PPARγ-FAS Pathway in a high-fat diet-induced mouse model. Processes 2023, 11, 735. [Google Scholar] [CrossRef]
- Meriga, B.; Parim, B.; Chunduri, V.R.; Naik, R.R.; Nemani, H.; Suresh, P.; Ganapathy, S.; Uddandrao, V.V. Antiobesity potential of Piperonal: Promising modulation of body composition, lipid profiles and obesogenic marker expression in HFD-induced obese rats. Nutr. Metab. 2017, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Santoso, P.; Amelia, A.; Rahayu, R. Jicama (Pachyrhizus erosus) fiber prevents excessive blood glucose and body weight increase without affecting food intake in mice fed with high-sugar diet. J. Adv. Vet. Anim. Res. 2019, 6, 222. [Google Scholar] [CrossRef]
- Santoso, P.; Maliza, R.; Insani, S.J.; Fadhilah, Q. Effect of Jicama (Pachyrhizus erosus) fiber on energy intake and adipose tissue profiles in mice fed with high-fat diet. J. Phys. Conf. Ser. 2021, 1940, 012055. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, J.S.; Jung, J.W.; Byun, K.W.; Kang, K.S.; Lee, Y.S. Daidzein supplementation prevents non-alcoholic fatty liver disease through alternation of hepatic gene expression profiles and adipocyte metabolism. Int. J. Obes. 2011, 35, 1019–1030. [Google Scholar] [CrossRef]
- Martin, A.E.; Montgomery, P.A. Acarbose: An a-glucosidase inhibitor. Am. J. Health Syst. Pharm. 1996, 53, 2277–2290. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH IC50 (µg/mL) | ABTS IC50 (µg/mL) | TPC (GAE µg/g) | TFC (QUE µg/g) |
|---|---|---|---|---|
| 70% EtOH | 8572.7 ± 356.7 | 1130.1 ± 4.1 | 2674.7 ± 32.8 | 376.9 ± 2.7 |
| n-Hexane | 4890.9 ± 167.5 | 4604.6 ± 7.3 | 1254.7 ± 6.1 | 24.3 ± 2.8 |
| EtOAc | 1054.1 ± 21.5 | 531.8 ± 0.7 | 3105.1 ± 12.3 | 1063.0 ± 12.3 |
| BuOH | 1772.6 ± 16.3 | 907.5 ± 1.4 | 1732.0 ± 16.2 | 80.00 ± 2.8 |
| Water | 1091.2 ± 9.9 | 1191.6 ± 5.7 | 1745.0 ± 6.1 | 68.1 ± 2.8 |
| Daidzein | 14.8 ± 0.4 | 11.3 ± 0.1 | - | - |
| No. | Compound | 70% EtOH | EtOAc |
|---|---|---|---|
| 1 | Allopurinol | 21.1 | 19.3 |
| 2 | Apigenin | - | 18.9 |
| 3 | p-Coumaric acid | - | 6.96 |
| 4 | Daidzein | 14.3 | 319 |
| 5 | t-Ferulic acid | - | 3.34 |
| 6 | Formononetin | - | 0.33 |
| 7 | Fumaric acid | 124 | 90.5 |
| 8 | Genistein | 0.77 | 33.3 |
| 9 | Homogentisic acid | - | 14.1 |
| 10 | 4-Hydroxybenzoic acid | 34.2 | 581 |
| 11 | Hyperoside | - | 7.21 |
| 12 | Naringenin | - | 3.89 |
| 13 | Naringin | - | 12.7 |
| 14 | Nicotinic acid | 1.53 | 20.7 |
| 15 | Protocatechuic acid | 1.61 | 15.7 |
| 16 | Resorcylic acid | - | 23.7 |
| 17 | Rotenone | 0.47 | 0.07 |
| 18 | Rutin | - | 8.92 |
| 19 | Salicylic acid | - | 24.6 |
| 20 | Shikimic acid | 9.26 | 8.51 |
| 21 | Taxifolin | - | 0.73 |
| 22 | Vanillic acid | - | 73.5 |
| Total | 207.24 | 1286.96 | |
| Sample | α-Glucosidase IC50 (µg/mL) | Lipase IC50 (µg/mL) |
|---|---|---|
| 70% EtOH | 2314.1 ± 30.7 | 4069.6 ± 398.3 |
| EtOAc | 873.1 ± 14.5 | 915.0 ± 4.6 |
| Daidzein | 87.6 ± 3.9 | 84.9 ± 2.3 |
| Acarbose | 106.7 ± 4.3 | - |
| Orlistat | - | 63.3 ± 4.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-H.; Yang, S.-H.; Lee, Y.-J.; Sohn, J.H.; Lee, K.S.; Shin, H.-J. Anti-Obesity Effect of Daidzein Derived from Pachyrhizus erosus (L.) Urb. Extract via PPAR Pathway in MDI-Induced 3T3-L1 Cell Line. Cosmetics 2023, 10, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10060164
Choi M-H, Yang S-H, Lee Y-J, Sohn JH, Lee KS, Shin H-J. Anti-Obesity Effect of Daidzein Derived from Pachyrhizus erosus (L.) Urb. Extract via PPAR Pathway in MDI-Induced 3T3-L1 Cell Line. Cosmetics. 2023; 10(6):164. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10060164
Chicago/Turabian StyleChoi, Moon-Hee, Seung-Hwa Yang, Yeo-Jin Lee, Jeong Ho Sohn, Ki Sun Lee, and Hyun-Jae Shin. 2023. "Anti-Obesity Effect of Daidzein Derived from Pachyrhizus erosus (L.) Urb. Extract via PPAR Pathway in MDI-Induced 3T3-L1 Cell Line" Cosmetics 10, no. 6: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10060164