
Colorectal Cancer Chemoprevention by S-Allyl Cysteine–Caffeic Acid Hybrids: In Vitro Biological Activity and In Silico Studies

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
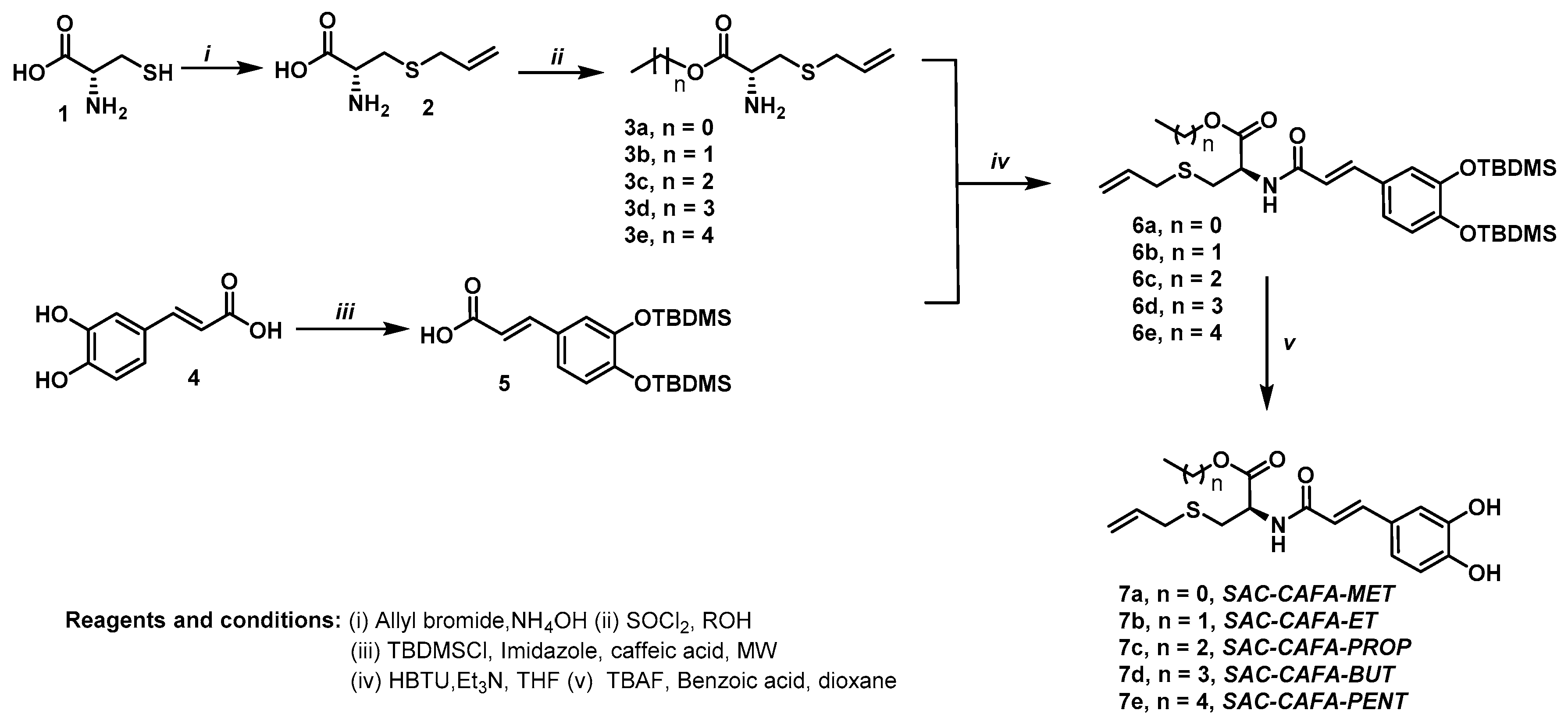
2.1. Source of Hybrid Molecules
2.2. Biological Activity
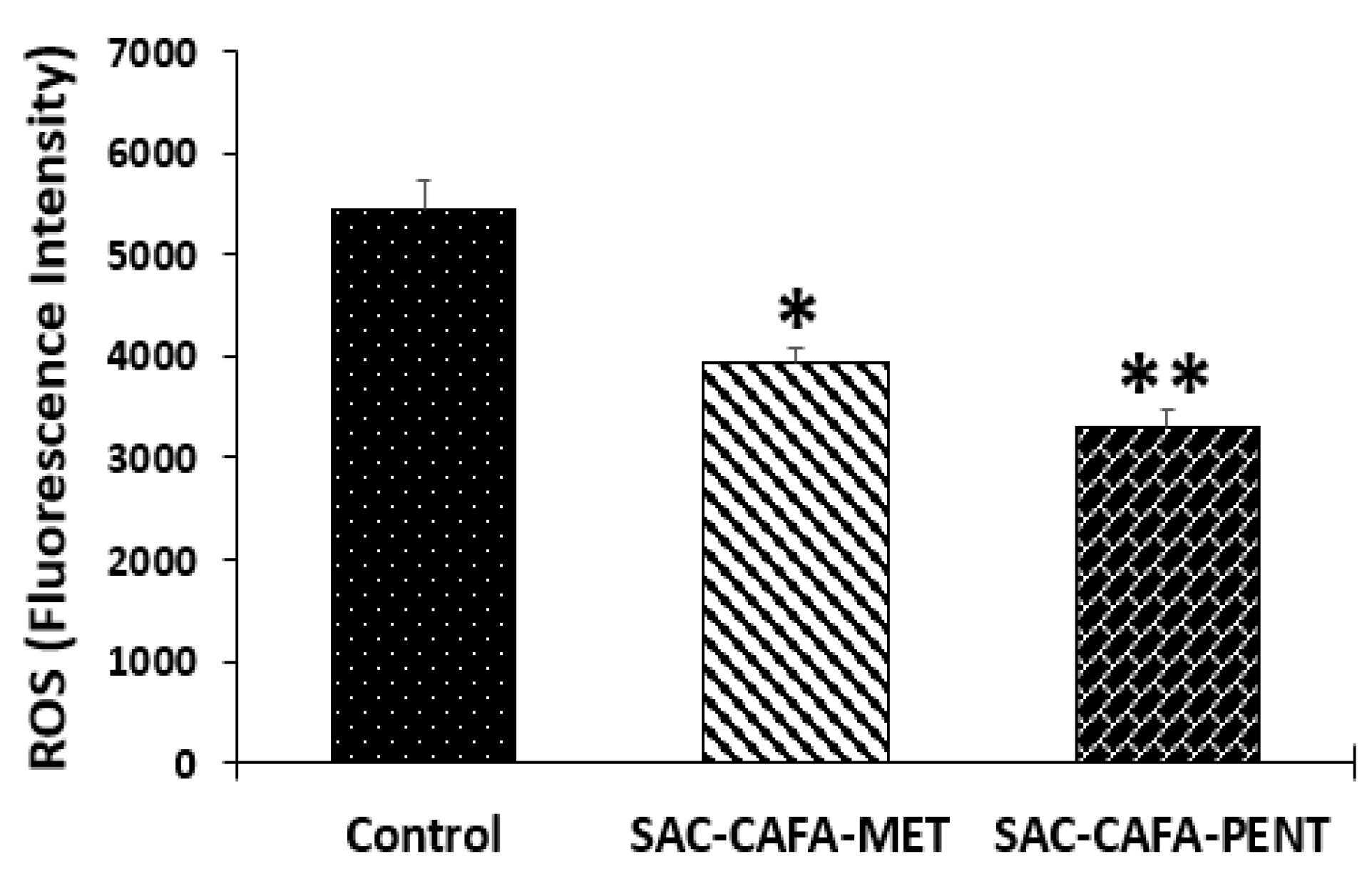
2.2.1. Effect of SAC-CAFA-MET and SAC-CAFA-PENT on Reactive Oxygen Species (ROS)
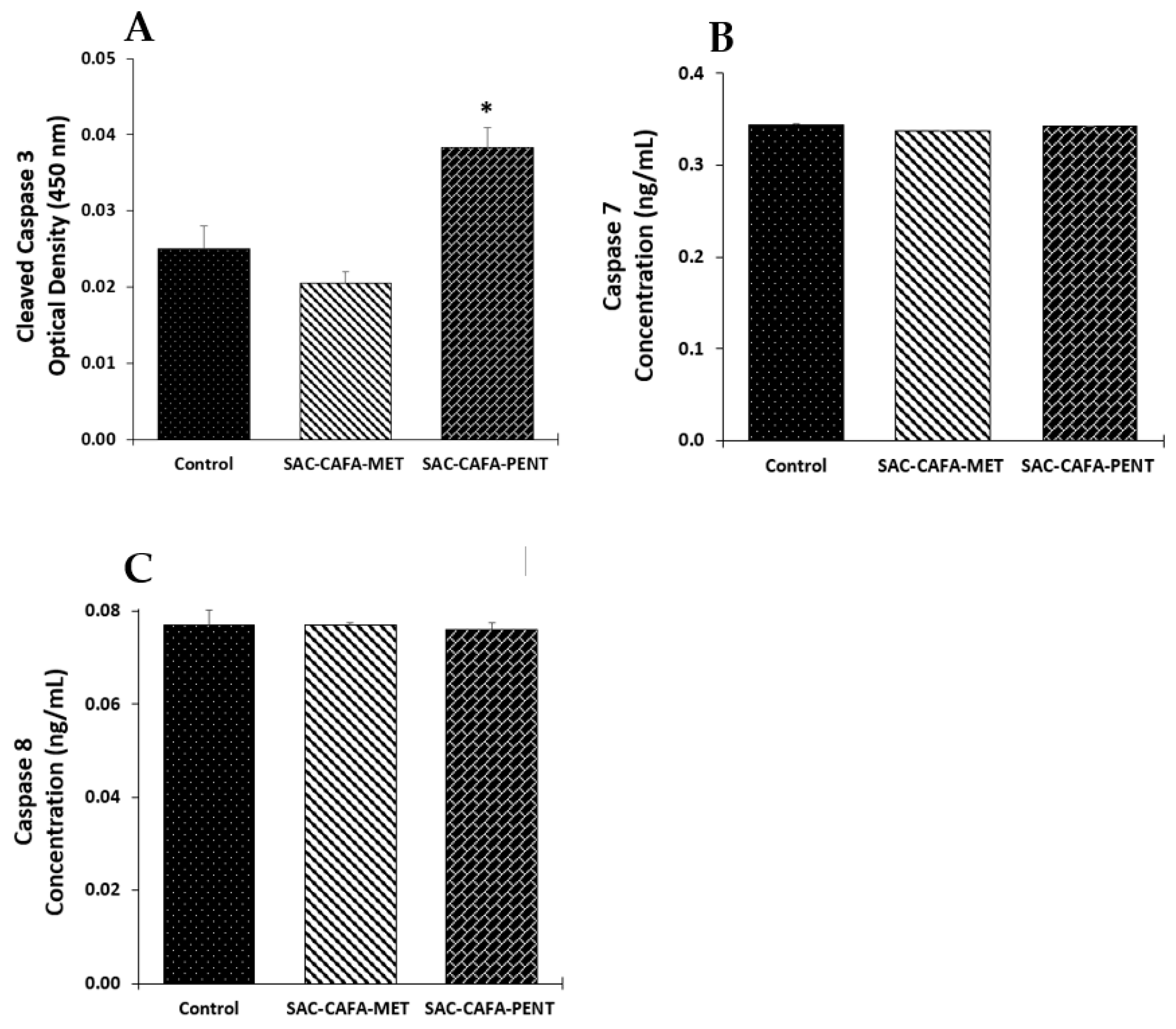
2.2.2. Effects of SAC-CAFA-MET and SAC-CAFA-PENT on the Expression of Caspase-3, -7, and -8
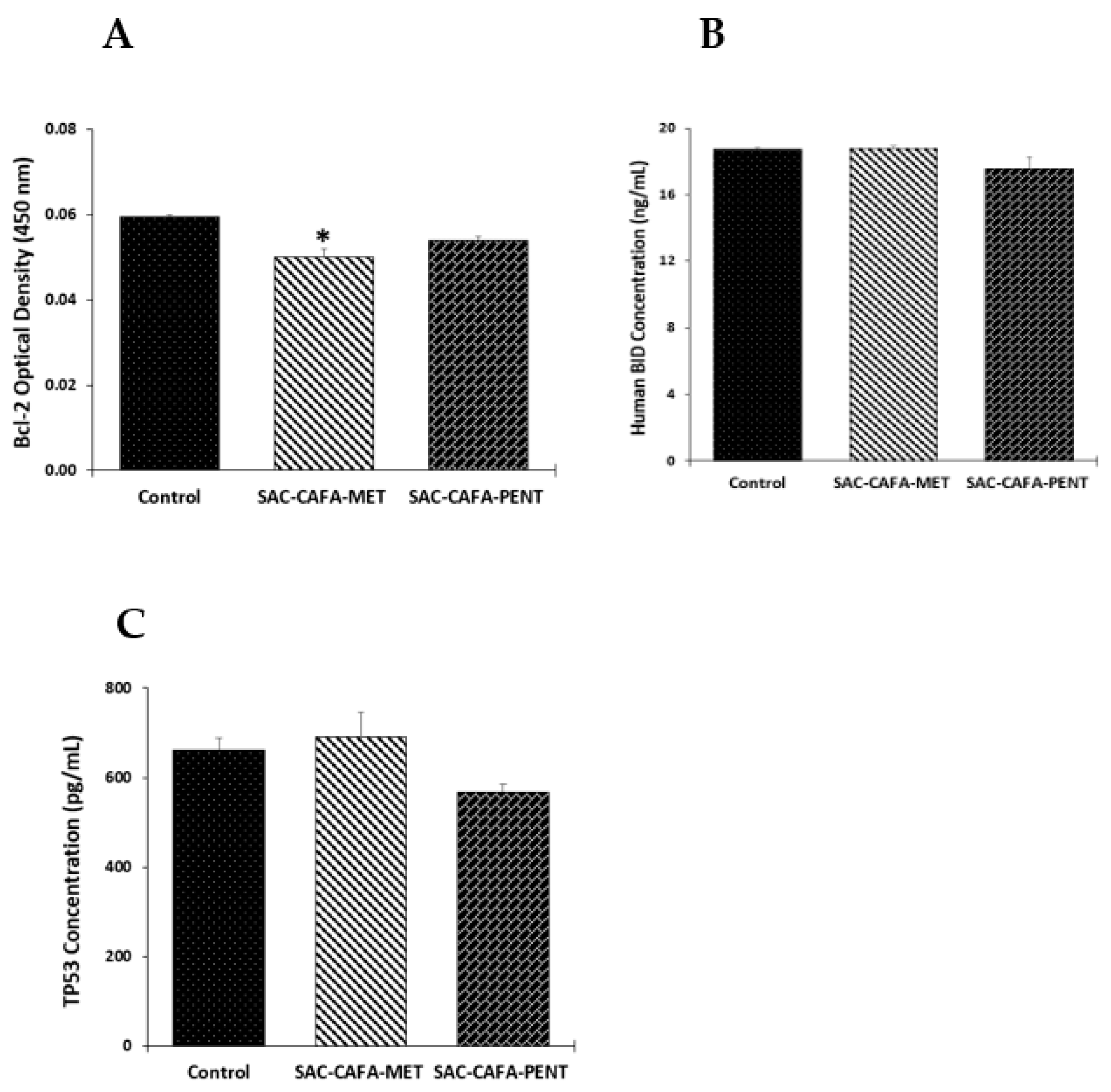
2.2.3. Effects of SAC-CAFA-MET and SAC-CAFA-PENT on the Expression of Apoptotic Biomarkers
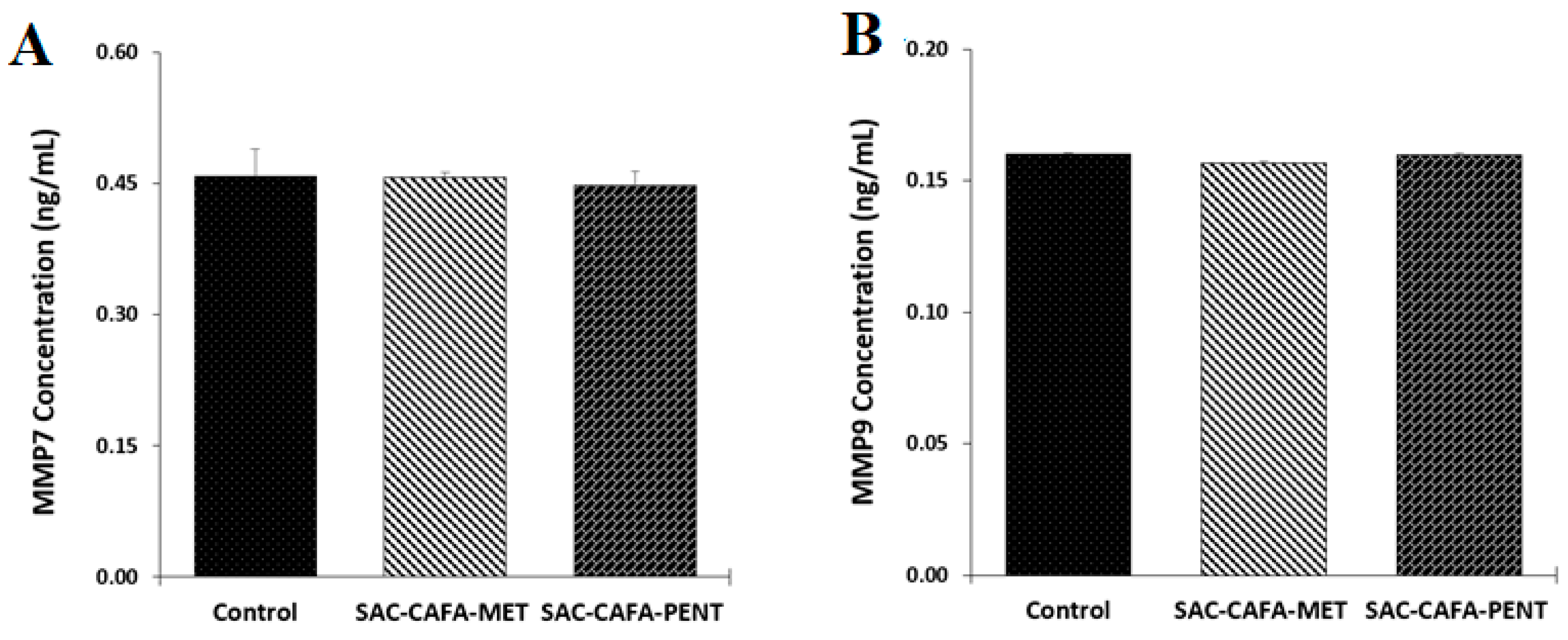
2.2.4. Effects of the Hybrids SAC-CAFA-MET and SAC-CAFA-PENT on the Activity of Matrix Metalloproteinase 7 (MMP7) and MMP9
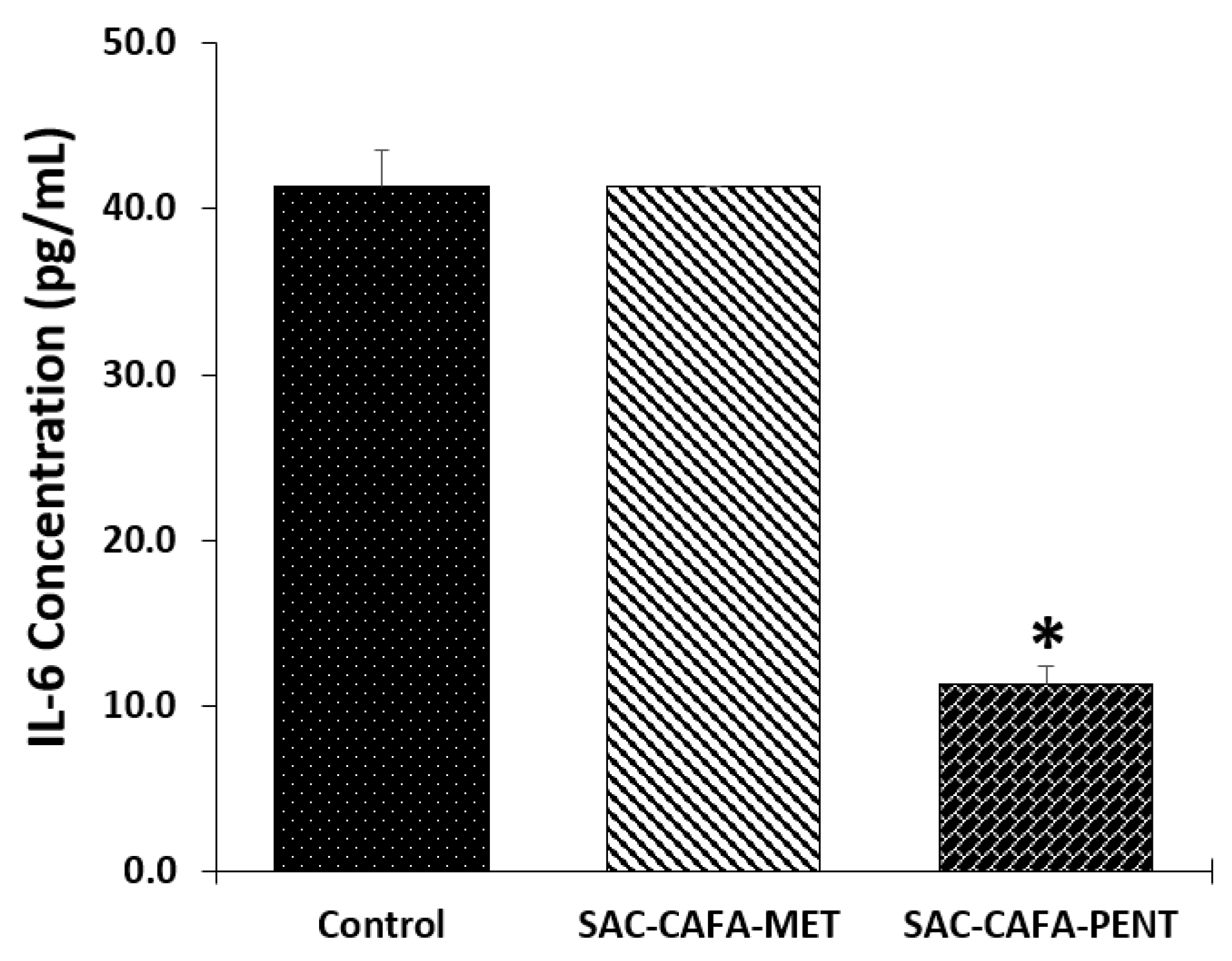
2.2.5. Effects of the hybrids SAC-CAFA-MET and SAC-CAFA-PENT on Expression of Interleukin-6 (IL-6)
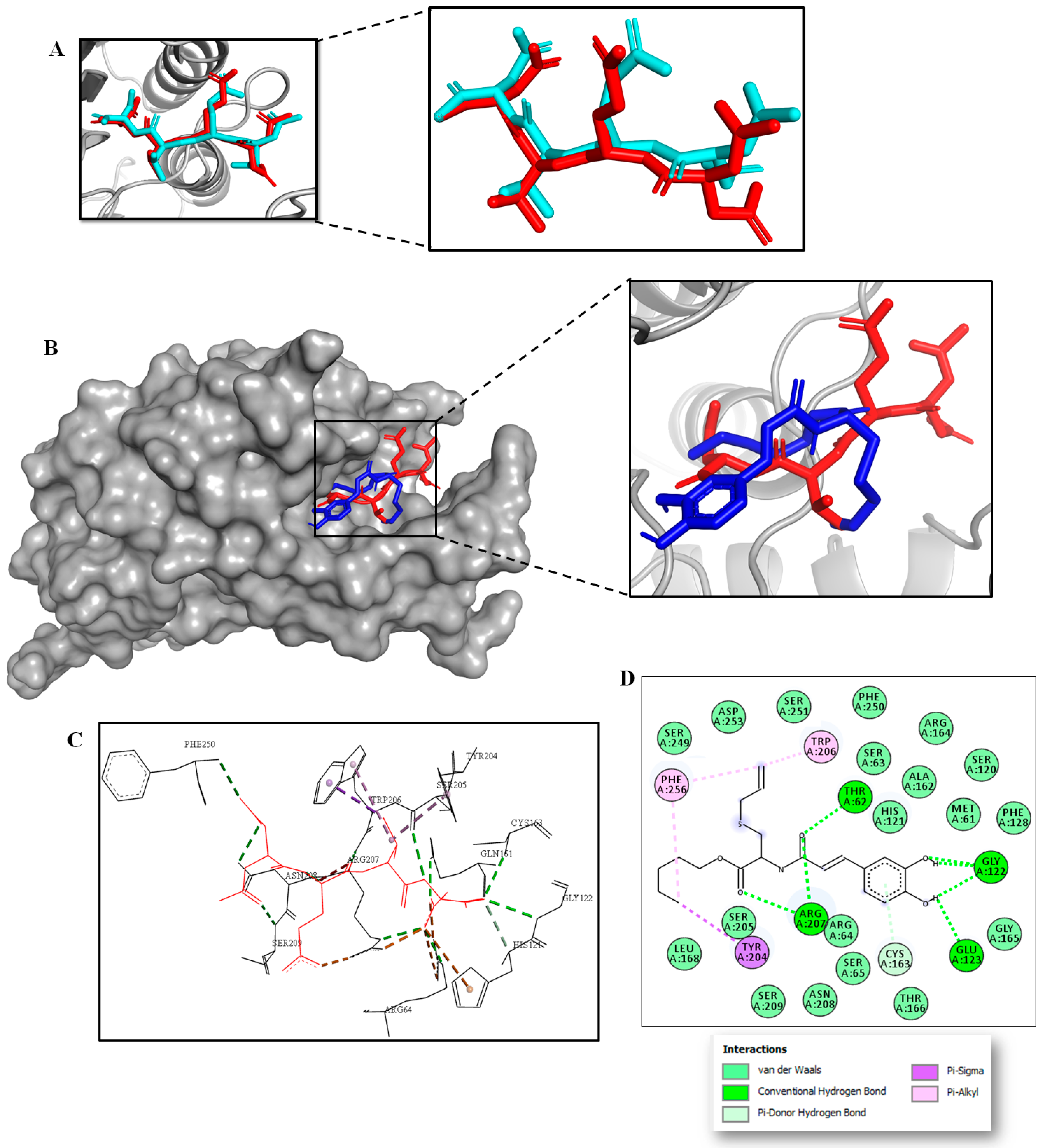
2.3. Multitarget Docking Studies and Prediction of Binding Pose
2.4. DFT Analysis of the Radical-Scavenging Activity of SAC-CAFA-MET and SAC-CAFA-PENT
- (i)
- Hydrogen atom transfer (HAT):
- (ii)
- Sequential proton transfer–electron transfer (SPT–ET):
- (iii)
- Sequential electron transfer–proton transfer (SET–PT):
3. Materials and Methods
3.1. In Vitro Biological Assays
3.1.1. Cell Line, Culture Medium, and Treatment
3.1.2. Determination of ROS
3.1.3. Determination of Inflammatory Cytokines and Apoptotic Proteins
3.1.4. Statistical Analysis
3.2. Computational Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SAC | S-allyl cysteine |
| CA | Caffeic acid |
| CRC | Colorectal cancer |
| DMSO | Dimethyl sulfoxide |
| 5-FU | 5-Fluorouracil |
| ROS | Reactive oxygen species |
References
- Hull, R.; Francies, F.Z.; Oyomno, M.; Dlamini, Z. Colorectal cancer genetics, incidence and risk factors: In search for targeted therapies. Cancer Manag. Res. 2020, 12, 9869–9882. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a preventable disease that requires major lifestyle changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2020, 70, 313. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Alam, W.; Bouferraa, Y.; Haibe, Y.; Mukherji, D.; Shamseddine, A. Management of colorectal cancer in the era of COVID-19_Challenges and suggestions. Sci. Prog. 2021, 104, 104. [Google Scholar] [CrossRef]
- Pointet, A.L.; Taieb, J. Cáncer de colon. EMC-Tratado Med. 2017, 21, 1–7. [Google Scholar] [CrossRef]
- McQuade, R.M.; Bornstein, J.C.; Nurgali, K. Anti-colorectal cancer chemotherapy-induced diarrhoea: Current treatments and side effects. Int. J. Clin. Med. 2014, 5, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [Green Version]
- Ismail, T.; Donati-Zeppa, S.; Akhtar, S.; Turrini, E.; Layla, A.; Sestili, P.; Fimognari, C. Coffee in cancer chemoprevention: An updated review. Expert. Opin. Drug Metab. Toxicol. 2020, 17, 69–85. [Google Scholar] [CrossRef]
- Steward, W.P.; Brown, K. Cancer chemoprevention: A rapidly evolving field. Br. J. Cancer. 2013, 109, 1–7. [Google Scholar] [CrossRef]
- Kerru, N.; Singh, P.; Koorbanally, N.; Raj, R.; Kumar, V. Recent advances (2015–2016) in anticancer hybrids. Eur. J. Med. Chem. 2017, 142, 179–212. [Google Scholar] [CrossRef] [PubMed]
- Nepali, K.; Sharma, S.; Sharma, M.; Bedi, P.M.; Dhar, K.L. Rational approaches, design strategies, structure activity relationship and mechanistic insights for anticancer hybrids. Eur. J. Med. Chem. 2014, 22, 422–487. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Chun, S.B.; Koo, M.S.; Choi, W.J.; Kim, T.W.; Kwon, Y.G.; Chung, H.T.; Billiar, T.R.; Kim, Y.M. Differential regulation of NO availability from macrophages and endothelial cells by the garlic component S-allyl cysteine. Free Radic. Biol. Med. 2001, 30, 747–756. [Google Scholar] [CrossRef]
- Ho, S.E.; Ide, N.; Lau, B.H. S-allyl cysteine reduces oxidant load in cells involved in the atherogenic process. Phytomedicine 2001, 8, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Numagami, Y.; Ohnishi, S.T. S-allyl cysteine inhibits free radical production, lipid peroxidation and neuronal damage in rat brain ischemia. J. Nutr. 2001, 131, 1100S–1105S. [Google Scholar] [CrossRef]
- Numagami, Y.; Sato, S.; Ohnishi, S.T. Attenuation of rat ischemic brain damage by aged garlic extracts: A possible protecting mechanism as antioxidants. Neurochem. Int. 1996, 29, 135–143. [Google Scholar] [CrossRef]
- Mostafa, M.G.; Mima, T.; Ohnishi, S.T.; Mori, K. S-allylcysteine ameliorates doxorubicin toxicity in the heart and liver in mice. Planta Med. 2000, 66, 148–151. [Google Scholar] [CrossRef]
- Welch, C.; Wuarin, L.; Sidell, N. Antiproliferative effect of the garlic compound S-allyl cysteine on human neuroblastoma cells in-vitro. Cancer Lett. 1992, 63, 211–219. [Google Scholar] [CrossRef]
- Takeyama, H.; Hoon, D.S.; Saxton, R.E.; Morton, D.L.; Irie, R.F. Growth inhibition and modulation of cell markers of melanoma by S-allyl cysteine. Oncology 1993, 50, 63–69. [Google Scholar] [CrossRef]
- Hung, C.C.; Tsai, W.J.; Kuo, L.M.; Kuo, Y.H. Evaluation of caffeic acid amide analogues as anti-platelet aggregation and anti-oxidative agents. Bioorg. Med. Chem. 2005, 13, 1791–1797. [Google Scholar] [CrossRef]
- Son, S.; Lewis, B.A. Free radical scavenging and antioxidative activity of caffeic acid amide and ester analogues: Structure-activity relationship. J. Agric. Food Chem. 2002, 50, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Otero, E.; Robledo, S.; Díaz, S.; Carda, M.; Muñoz, D.; Paños, J.; Vélez, I.D.; Cardona, W. Synthesis and leishmanicidal activity of cinnamic acid esters: Structure–activity relationship. Med. Chem. Res. 2014, 23, 1378–1386. [Google Scholar] [CrossRef] [Green Version]
- De, P.; Baltas, M.; Bedos-Belval, F. Cinnamic acid derivatives as anticancer agents—A review. Curr. Med. Chem. 2011, 18, 1672–1703. [Google Scholar] [CrossRef] [PubMed]
- Cardona-G, W.; Herrera-R, A.; Castrillón-L, W.; Ramírez-Malule, H. Chemistry and anticancer activity of hybrid molecules and derivatives based on 5-fluorouracil. Curr. Med. Chem. 2021, 28, 5551–5601. [Google Scholar] [CrossRef] [PubMed]
- Decker, M. Design of hybrid molecules for drug development. In Editorial Project Manager; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 978-0-08-101011-2. [Google Scholar]
- Shaveta, S.M.; Singh, P. Hybrid molecules: The privileged scaffolds for various pharmaceuticals. Eur. J. Med. Chem. 2016, 124, 500–536. [Google Scholar] [CrossRef] [PubMed]
- Tsogoeva, S.B. Recent progress in the development of synthetic hybrids of natural or unnatural bioactive compounds for medicinal chemistry. Mini Rev. Med. Chem. 2010, 10, 773–793. [Google Scholar] [CrossRef]
- Meunier, B. Hybrid molecules with a dual mode of action: Dream or reality? Acc. Chem. Res. 2008, 41, 69–77. [Google Scholar] [CrossRef]
- Castrillón, W.; Herrera-R, A.; Prieto, L.J.; Conesa-Milián, L.; Carda, M.; Naranjo, T.; Maldonado, M.E.; Cardona-G, W. Synthesis and in-vitro Evaluation of S-allyl cysteine ester—Caffeic acid amide hybrids as potential anticancer agents. Iran. J. Pharm. Sci. 2019, 18, 1770–1789. [Google Scholar]
- Hebbar, S.; Knust, E. Reactive oxygen species (ROS) constitute an additional player in regulating epithelial d evelopment. BioEssays 2021, 43, 2100096. [Google Scholar] [CrossRef]
- Sun, Y.; Lu., Y.; Saredy, J.; Wang, X.; Drummer, C., IV; Shao, Y.; Saaoud, F.; Xu, K.; Liu, M.; Yang, W.Y.; et al. ROS systems are a new integrated network for sensing homeostasis and alarming stresses in organelle metabolic processes. Redox Biol. 2020, 37, 101696. [Google Scholar] [CrossRef]
- Nourazarian, A.R.; Kangari, P.; Salmaninejad, A. Roles of oxidative stress in the development and progression of breast cancer. Asian Pac. J. Cancer Prev. 2014, 15, 4745–4751. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ahn, K.S.; Kim, C.; Shanmugam, M.K.; Siveen, K.S.; Arfuso, F.; Samym, R.P.; Deivasigamanim, A.; Lim, L.H.K.; Wang, L.; et al. Nimbolide-induced oxidative stress abrogates STAT3 signaling cascade and inhibits tumor growth in transgenic adenocarcinoma of mouse prostate model. Antioxid. Redox Signal. 2016, 24, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Carini, F.; Mazzola, M.; Rappa, F.; Jurjus, A.; Geagea, A.G.; Al Kattar, S.; Bou-Assi, T.; Jurjus, R.; Damiani, P.; Leone, A.; et al. Colorectal carcinogenesis: Role of oxidative stress and antioxidants. Anticancer Res. 2017, 37, 4759–4766. [Google Scholar] [PubMed] [Green Version]
- Oparka, M.; Walczak, J.; Malinska, D.; van Oppen, L.M.P.E.; Szczepanowska, J.; Koopman, W.J.H.; Wieckowski, M.R. Quantifying ROS levels using CM-H2DCFDA and HyPer. Methods 2016, 109, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.L.; Krishnan, K.; Campbell, S.E.; Palau, V.E. The role of antioxidants and pro-oxidants in colon cancer. World J. Gastrointest. Oncol. 2014, 6, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Mavumengwana, V.; Li, H.B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Lee, J.C.; Chang, N.; Chun, H.S.; Kim, W.K. S-Allyl-L-cysteine attenuates cerebral ischemic injury by scavenging peroxynitrite and inhibiting the activity of extracellular signal-regulated kinase. Free Radic. Res. 2006, 40, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W.; et al. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-κB signaling. PLoS ONE 2013, 8, e73877. [Google Scholar] [CrossRef] [Green Version]
- Ashafaq, M.; Khan, M.M.; Shadab Raza, S.; Ahmad, A.; Khuwaja, G.; Javed, H.; Khan, A.; Islam, F.; Siddiqui, M.S.; Safhi, M.M.; et al. S-allyl cysteine mitigates oxidative damage and improves neurologic deficit in a rat model of focal cerebral ischemia. Nutr. Res. 2012, 32, 133–143. [Google Scholar] [CrossRef]
- Javed, H.; Khan, M.M.; Khan, A.; Vaibhav, K.; Ahmad, A.; Khuwaja, G.; Ahmed, M.E.; Raza, S.S.; Ashafaq, M.; Tabassum, R.; et al. S-allyl cysteine attenuates oxidative stress associated cognitive impairment and neurodegeneration in mouse model of streptozotocin-induced experimental dementia of Alzheimer’s type. Brain Res. 2011, 1389, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Rojas, P.; Serrano-García, N.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Maldonado, P.D.; Ruiz-Sánchez, E. S-Allylcysteine, a garlic compound, protects against oxidative stress 1-methyl-4-phenylpyridinium-induced parkinsonism in mice. J. Nutr. Biochem. 2011, 22, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alotaibi, M.R.; Hassan, Z.K.; Al-Rejaie, S.S.; Alshammari, M.A.; Almutairi, M.M.; Alhoshani, A.R.; Alanazi, W.A.; Hafez, M.M.; Al-Shabanah, O.A. Characterization of apoptosis in a breast cancer cell line after IL-10 silencing. Asian Pac. J. Cancer Prev. 2018, 19, 777–783. [Google Scholar] [PubMed]
- Park, H.H. Structural features of caspase-activating complexes. Int. J. Mol. Sci. 2012, 13, 4807–4818. [Google Scholar] [CrossRef]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [Green Version]
- Bröker, L.E.; Kruyt, F.A.E.; Giaccone, G. Cell death independent of caspases: A review. Clin. Cancer Res. 2005, 11, 3155–3162. [Google Scholar] [CrossRef] [Green Version]
- Constantinou, C.; Papas, K.A.; Constantinou, A.I. Caspase-independent pathways of programmed cell death: The unraveling of new targets of cancer therapy? Curr. Cancer Drug Targets. 2009, 9, 717–728. [Google Scholar] [CrossRef]
- Warren, C.F.A.; Wong-Brown, M.W.; Bowden, N.A. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Akl, H.; Vervloessem, T.; Kiviluoto, S.; Bittremieux, M.; Parys, J.B.; De Smedt, H.; Bultynck, G. A dual role for the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus endoplasmic reticulum. Biochim. Biophys Acta. 2014, 1843, 2240–2252. [Google Scholar] [CrossRef] [Green Version]
- Popgeorgiev, N.; Jabbour, L.; Gillet, G. Subcellular localization and dynamics of the Bcl-2 family of proteins. Front. Cell Dev. Biol. 2018, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatokun, A.A.; Dawson, V.L.; Dawson, T.M. Parthanatos: Mitochondrial-linked mechanisms and therapeuti opportunitis. Br. J. Pharmacol. 2014, 171, 2000–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.J. Biology of mitochondria in neurodegenerative diseases. Prog. Mol. Biol. Transl. Sci. 2012, 107, 355–415. [Google Scholar] [PubMed] [Green Version]
- Galluzzi, L.; Blomgren, K.; Kroemer, G. Mitochondrial membrane permeabilization in neuronal injury. Nat. Rev. Neurosci. 2009, 10, 481–494. [Google Scholar] [CrossRef]
- Liu, Z.; Li, M.; Chen, K.; Yang, J.; Chen, R.; Wang, T.; Liu, J.; Yang, W.; Ye, Z. S-allyl cysteine induces cell cycle arrest and apoptosis in androgen-independent human prostate cancer cells. Mol. Med. Rep. 2012, 5, 439–443. [Google Scholar]
- Kirkin, V.; Joos, S.; Zörnig, M. The role of Bcl-2 family members in tumorigenesis. Biochim. Biophys. Acta 2004, 1644, 229–249. [Google Scholar] [CrossRef]
- Mantovani, F.; Collavin, L.; Del Sal, G. Mutant p53 as a guardian of the cancer cell. Cell Death Differ. 2019, 26, 199–212. [Google Scholar] [CrossRef]
- Xie, S.; Wang, Q.; Wu, H.; Cogswell, J.; Lu, L.; Jhanwar-Uniyal, M.; Dai, W. Reactive oxygen species-induced phosphorylation of p53 on serine 20 is mediated in part by polo-like kinase-3. J. Biol. Chem. 2001, 276, 36194–36199. [Google Scholar] [CrossRef] [Green Version]
- Nomura, M.; Kaji, A.; Ma, W.; Miyamoto, K.; Dong, Z. Suppression of cell transformation and induction of apoptosis by caffeic acid phenethyl ester. Mol. Carcinog. 2001, 31, 83–89. [Google Scholar] [CrossRef]
- Liao, H.Y.; Da, C.M.; Liao, B.; Zhang, H.H. Roles of matrix metalloproteinase-7 (MMP-7) in cancer. Clin. Biochem. 2021, 92, 9–18. [Google Scholar] [CrossRef]
- Said, A.H.; Raufman, J.P.; Xie, G. The role of matrix metalloproteinases in colorectal cancer. Cancers 2014, 6, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Colorectal carcinogenesis: Review of human and experimental animal studies. J. Carcinog. 2009, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; He, Z.; Ye, J.; Liu, Z.; She, X.; Gao, X.; Liang, R. Progress in understanding the IL-6-STAT3 pathway in colorectal cancer. Onco Targets Ther. 2020, 13, 13023–13032. [Google Scholar] [CrossRef] [PubMed]
- Knüpfer, H.; Preiss, R. Serum interleukin-6 levels in colorectal cancer patients—A summary of published results. Int. J. Colorectal Dis. 2009, 25, 135–140. [Google Scholar] [CrossRef]
- Galizia, G.; Orditura, M.; Romano, C.; Lieto, E.; Castellano, P.; Pelosio, L.; Imperatore, V.; Catalano, G.; Pignatelli, C.; De Vita, F. Prognostic significance of circulating IL-10 and IL-6 serum levels in colon cancer patients undergoing surgery. Clin. Immunol. 2002, 102, 169–178. [Google Scholar] [CrossRef]
- You, S.; Nakanishi, E.; Kuwata, H.; Chen, J.; Nakasone, Y.; He, X.; He, J.; Liu, X.; Zhang, S.; Zhang, B.; et al. Inhibitory effects and molecular mechanisms of garlic organosulfur compounds on the production of inflammatory mediators. Mol. Nutr. Food Res. 2013, 57, 2049–2060. [Google Scholar] [CrossRef]
- Zarezadeh, M.; Baluchnejadmojarad, T.; Kiasalari, Z.; Afshin-Majd, S.; Roghani, M. Garlic active constituent S-allyl cysteine protects against lipopolysaccharide-induced cognitive deficits in the rat: Possible involved mechanisms. Eur. J. Pharmacol. 2017, 795, 13–21. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in cell death, inflammation, and disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Maciag, J.; Mackenzie, S.H.; Tucker, M.B.; Schipper, J.L.; Swartz, P.D.; Clark, A.C. Tunable allosteric library of caspase-3 identifies coupling between conserved water molecules and conformational selection. Proc. Natl. Acad. Sci. USA 2016, 113, 6080–6088. [Google Scholar] [CrossRef] [Green Version]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative stress in cancer cell metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell. 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Herrera-R, A.; Castrillón, W.; Otero, E.; Ruiz, E.; Carda, M.; Agut, R.; Naranjo, T.; Moreno, G.; Maldonado, M.E.; Cardona-G, W. Synthesis and antiproliferative activity of 3- and 7-styrylcoumarins. Med. Chem. Res. 2018, 27, 1893–1905. [Google Scholar] [CrossRef]
- Pérez, J.M.; Maldonado, M.E.; Rojano, B.A.; Alzate, F.; Sáez, J.; Cardona, W. Comparative antioxidant, antiproliferative and apoptotic effects of ilex laurina and ilex paraguariensis on colon cancer cells. Trop. J. Pharm. Res. 2014, 13, 1279–1286. [Google Scholar] [CrossRef] [Green Version]
- Massagué, J. G1 cell-cycle control and cancer. Nature 2004, 432, 298–306. [Google Scholar] [CrossRef]
- Herrera-R, A.; Cardona-G, W.; Maldonado, M.E.; Naranjo, T.; Moreno-Q, G. Styrylcoumarin 7-SC2 induces cell death in SW480 human colon adenocarcinoma cells and inhibits azoxymethane-induced aberrant crypt foci formation in BALB/c mice. Med. Chem. Res. 2019, 29, 377–395. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodshell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a lamarckian genetic algorithm and empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 3, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-Class functionals and 12 other function. Theor. Chem. Acc. 2007, 120, 215–241. [Google Scholar] [CrossRef] [Green Version]
- Ditchfield, R.; Hehre, W.J.; Pople, J.A. Self-consistent molecular-orbital methods. IX. An extended gaussian-type basis for molecular-orbital studies of organic molecules. J. Chem. Phys. 1971, 54, 724–728. [Google Scholar] [CrossRef]
- Hehre, W.J.; Ditchfield, R.; Pople, J.A. Self-consistent molecular orbital methods. XII. Further extensions of gaussian—Type basis sets for use in molecular orbital studies of organic molecules. J. Chem. Phys. 1972, 56, 2257–2261. [Google Scholar] [CrossRef]
- Hariharan, P.C.; Pople, J.A. The influence of polarization functions on molecular orbital hydrogenation energies. Theor. Chim. Acta. 1973, 28, 213–222. [Google Scholar] [CrossRef]
- Frisch, M.J.; Pople, J.A.; Binkley, J.S. Self-consistent molecular orbital methods 25. Supplementary functions for gaussian basis sets. J. Chem. Phys. 1984, 80, 3265–3269. [Google Scholar] [CrossRef]
- Tishchenko, O.; Truhlar, D.G. Benchmark Ab initio calculations of the barrier height and transition-state geometry for hydrogen abstraction from a phenolic antioxidant by a peroxy radical and its use to assess the performance of density functionals. J. Phys. Chem. Lett. 2012, 3, 2834–2839. [Google Scholar] [CrossRef]
- Zheng, J.; Zhao, Y.; Truhlar, D.G. Thermochemical kinetics of hydrogen-atom transfers between methyl, methane, ethynyl, ethyne, and hydrogen. J. Phys. Chem. A 2007, 111, 4632–4642. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; et al. General atomic and molecular electronic structure system. J. Comput. Chem. 1993, 14, 1347–1363. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Structure | Target Proteins (Docking Score, kcal·mol−1) | ||
|---|---|---|---|---|
| Caspase-3 (PDB ID: 5I9B) | Bcl-2 b (PDB: 4MAN) | IL-6 c (PDB: 1ALU) | ||
| SAC-CAFA-MET (7a) |  | - | −5.1 | - |
| SAC-CAFA-PENT (7e) |  | −7.3 | - | −5.0 |
| Venetoclax a | - | −10.9 | - | |
| Ac-DEVD-CMK d | −8.1 | - | - | |
| Against ∙OH Radical | |||||
|---|---|---|---|---|---|
| SPT-ET | SET-PT | ||||
| HAT | Step 1 | Step 2 | Step 1 | Step 2 | |
| SAC-CAFA-MET | −39.0 | 49.6 | −88.6 | 8.4 | −47.4 |
| SAC-CAFA-PENT | −39.1 | 51.7 | −90.8 | 4.7 | −43.8 |
| Against O2∙− | |||||
| SAC-CAFA-MET | 10.4 | 7.0 | 3.4 | 70.2 | −59.8 |
| SAC-CAFA-PENT | 10.3 | 9.2 | 1.1 | 66.5 | −56.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Ramirez, A.; Yepes-Pérez, A.F.; Quintero-Saumeth, J.; Moreno-Quintero, G.; Naranjo, T.W.; Cardona-Galeano, W. Colorectal Cancer Chemoprevention by S-Allyl Cysteine–Caffeic Acid Hybrids: In Vitro Biological Activity and In Silico Studies. Sci. Pharm. 2022, 90, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm90030040
Herrera-Ramirez A, Yepes-Pérez AF, Quintero-Saumeth J, Moreno-Quintero G, Naranjo TW, Cardona-Galeano W. Colorectal Cancer Chemoprevention by S-Allyl Cysteine–Caffeic Acid Hybrids: In Vitro Biological Activity and In Silico Studies. Scientia Pharmaceutica. 2022; 90(3):40. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm90030040
Chicago/Turabian StyleHerrera-Ramirez, Angie, Andres F. Yepes-Pérez, Jorge Quintero-Saumeth, Gustavo Moreno-Quintero, Tonny W. Naranjo, and Wilson Cardona-Galeano. 2022. "Colorectal Cancer Chemoprevention by S-Allyl Cysteine–Caffeic Acid Hybrids: In Vitro Biological Activity and In Silico Studies" Scientia Pharmaceutica 90, no. 3: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm90030040