Metabolomes and Lipidomes of the Infective Stages of the Gastrointestinal nematodes, Nippostrongylus brasiliensis and Trichuris muris

 ,
,  , ,
, ,
Abstract
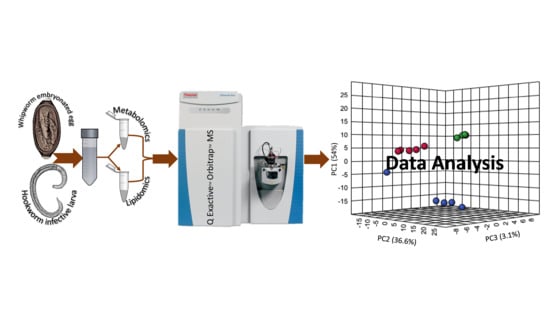
:
1. Introduction
2. Results
2.1. MSI Level-1 Identification of Polar Metabolites Present in the Infective Stages of N. brasiliensis and T. muris
2.2. Metabolic Pathways of Putatively Identified Polar Metabolites
2.3. Chemometric Analysis of the Polar Metabolites of N. brasiliensis L3 and T. muris Embryonated Eggs
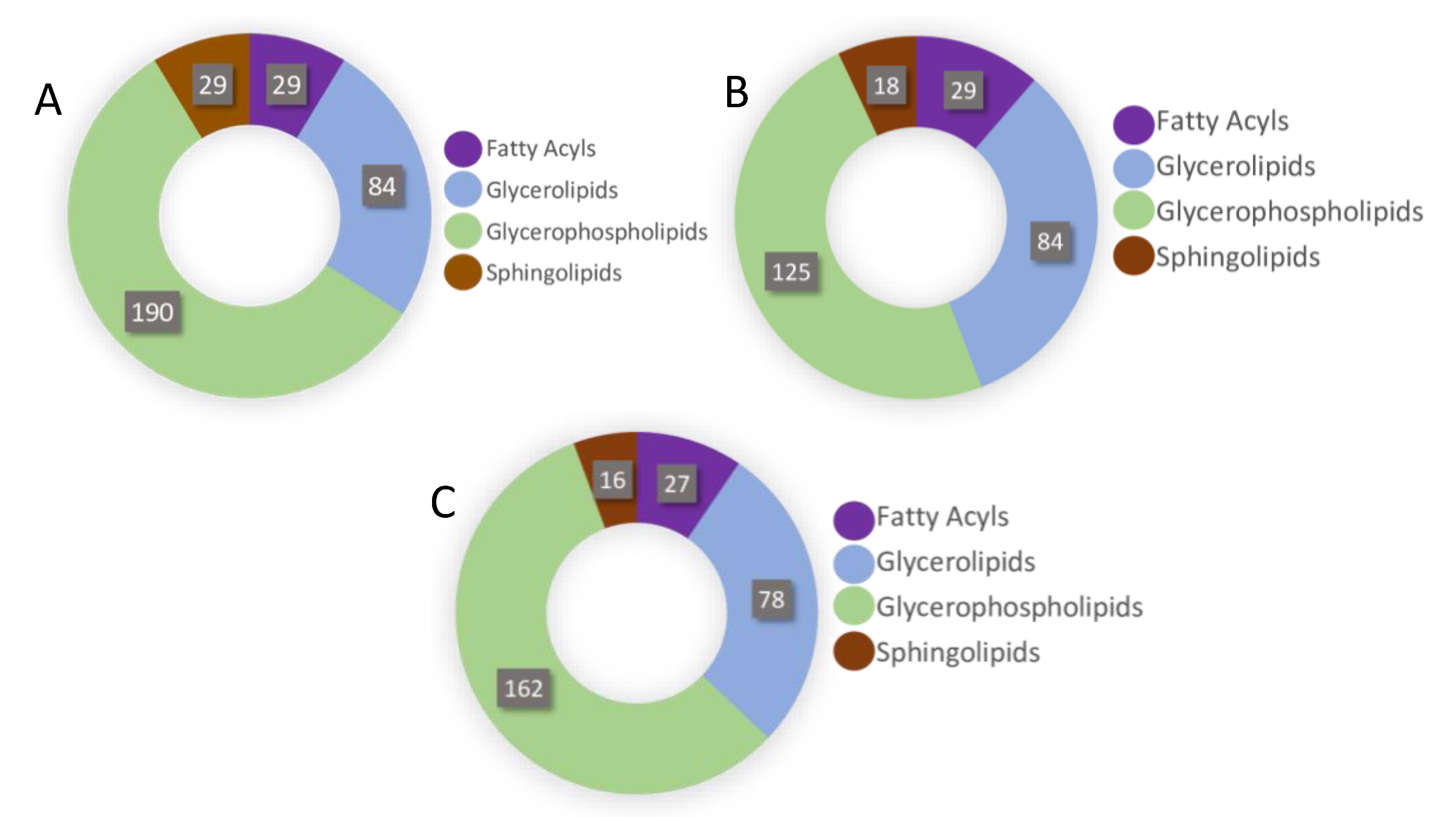
2.4. Lipidomics Analysis of the N. brasiliensis and T. muris Infective Stages
2.5. Lipidomic Pathways of Identified Lipids
2.6. Chemometric Analysis of Putatively Identified Lipids
2.7. Common and Unique Metabolites in N. brasiliensis and T. muris Infective Stages
2.8. Reported Pharmacological Activities of Identified Compounds
3. Discussion
4. Materials and Methods
4.1. Ethics, Source, and Housing of Mice and Rats
4.2. Collection of N. brasiliensis L3 and Its ESP
4.3. Collection of Eggs (Infective Stage) from T. muris Adult Worms
4.4. Somatic Extract Preparation
4.5. ESP Extract Preparation
4.6. Metabolomics LC-MS Data Acquisition
4.7. Lipidomics LC-MS Data Acquisition
4.8. Data Processing Using IDEOM
4.9. Data Analyses and Statistical Interpretation
4.10. Literature Review on Pharmacological Properties of Identified Metabolites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Investing to Overcome the Global Impact of Neglected Tropical Diseases: Third WHO Report on Neglected Tropical Diseases 2015; World Health Organization: Geneva, Switzerland, 2015; pp. 1–191. [Google Scholar]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth infections: The great neglected tropical diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Soil-Transmitted Helminth Infections; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Brooker, S.; Bethony, J.; Hotez, P.J. Human Hookworm Infection in the 21st Century. Adv. Parasitol. 2004, 58, 197–288. [Google Scholar] [PubMed] [Green Version]
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Dunn, J.C.; Bettis, A.A.; Wyine, N.Y.; Lwin, A.M.M.; Tun, A.; Maung, N.S.; Anderson, R.M. Soil-transmitted helminth reinfection four and six months after mass drug administration: Results from the delta region of Myanmar. PLoS Negl. Trop. Dis. 2019, 13, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchuem Tchuente, L.A. Control of soil-transmitted helminths in sub-Saharan Africa: Diagnosis, drug efficacy concerns and challenges. Acta. Trop. 2011, 120S, S4–S11. [Google Scholar] [CrossRef] [PubMed]
- Holden-Dye, L.; Walker, R.J. Anthelmintic drugs. WormBook Ed. C Elegans Res. Community WormBook 2007, 1–13. [Google Scholar] [CrossRef]
- Ok, K.S.; Kim, Y.S.; Song, J.H.; Lee, J.H.; Ryu, S.H.; Lee, J.H.; Moon, J.S.; Whang, D.H.; Lee, H.K. Trichuris trichiura infection diagnosed by colonoscopy: Case reports and review of literature. Korean J. Parasitol. 2009, 47, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Easton, A.V.; Oliveira, R.G.; O’Connell, E.M.; Kepha, S.; Mwandawiro, C.S.; Njenga, S.M.; Kihara, J.H.; Mwatele, C.; Odiere, M.R.; Brooker, S.J.; et al. Multi-parallel qPCR provides increased sensitivity and diagnostic breadth for gastrointestinal parasites of humans: Field-based inferences on the impact of mass deworming. Parasit Vectors 2016, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- WHO. Prevention and Control of Schistosomiasis and Soil-Transmitted Helminthiasis: Report of a WHO Expert Committee; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Knopp, S.; Rinaldi, L.; Khamis, I.S.; Stothard, J.R.; Rollinson, D.; Maurelli, M.P.; Steinmann, P.; Marti, H.; Cringoli, G.; Utzinger, J. A single FLOTAC is more sensitive than triplicate Kato-Katz for the diagnosis of low-intensity soil-transmitted helminth infections. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Coghlan, A.; Tygai, R.; Cotton, J.A. Comparative genomics of the major parasitic worms. Nat. Genet. 2018, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Jex, A.R.; Waeschenbach, A.; Hu, M.; van Wyk, J.A.; Beveridge, I.; Littlewood, D.T.; Gasser, R.B. The mitochondrial genomes of Ancylostoma caninum and Bunostomum phlebotomum—Two hookworms of animal health and zoonotic importance. BMC Genom. 2009, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Meekums, H.; Hawash, M.B.; Sparks, A.M.; Oviedo, Y.; Sandoval, C.; Chico, M.E.; Stothard, J.R.; Cooper, P.J.; Nejsum, P.; Betson, M. A genetic analysis of Trichuris trichiura and Trichuris suis from Ecuador. Parasit. Vectors 2015, 8, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.T.; Gao, X.; Rosa, B.A.; Abubucker, S.; Hallsworth-Pepin, K.; Martin, J.; Tyagi, R.; Heizer, E.; Zhang, X.; Bhonagiri-Palsikar, V.; et al. Genome of the human hookworm Necator americanus. Nat. Genet. 2014, 46, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Foth, B.J.; Tsai, I.J.; Reid, A.J.; Bancroft, A.J.; Nichol, S.; Tracey, A.; Holroyd, N.; Cotton, J.A.; Stanley, E.J.; Zarowiecki, M.; et al. Whipworm genome and dual-species transcriptome analyses provide molecular insights into an intimate host-parasite interaction. Nat. Genet. 2014, 46, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Leroux, L.P.; Nasr, M.; Valanparambil, R.; Tam, M.; Rosa, B.A.; Siciliani, E.; Hill, D.E.; Zarlenga, D.S.; Jaramillo, M.; Weinstock, J.V.; et al. Analysis of the Trichuris suis excretory/secretory proteins as a function of life cycle stage and their immunomodulatory properties. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Mulvenna, J.; Hamilton, B.; Nagaraj, S.H.; Smyth, D.; Loukas, A.; Gorman, J.J. Proteomics analysis of the excretory/secretory component of the blood-feeding stage of the hookworm, Ancylostoma caninum. Mol. Cell Proteom. 2009, 8, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.N.; Gallo, M.B.; Silva, E.S.; Figueiredo, C.A.; Cooper, P.J.; Barreto, M.L.; Loureiro, S.; Pontes-de-Carvalho, L.C.; Alcantara-Neves, N.M. A proteomic approach to identify proteins from Trichuris trichiura extract with immunomodulatory effects. Parasite Immunol. 2013, 35, 188–193. [Google Scholar] [CrossRef]
- Wangchuk, P.; Kouremenos, K.; Eichenberger, R.M.; Pearson, M.; Susianto, A.; Wishart, D.S.; McConville, M.J.; Loukas, A. Metabolomic profiling of the excretory-secretory products of hookworm and whipworm. Metabolomics 2019, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wangchuk, P.; Constantinoiu, C.; Eichenberger, R.M.; Field, M.; Loukas, A. Characterization of tapeworm metabolites and their reported biological activities. Molecules 2019, 24, 1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giera, M.; Kaisar, M.M.M.; Derks, R.J.E.; Steenvoorden, E.; Kruize, Y.C.M.; Hokke, C.H.; Yazdanbakhsh, M.; Everts, B. The Schistosoma mansoni lipidome: Leads for immunomodulation. Anal. Chim. Acta 2018, 1037, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Eichenberger, R.M.; Ryan, S.; Jones, L.; Buitrago, G.; Polster, R.; Montes de Oca, M.; Zuvelek, J.; Giacomin, P.R.; Dent, L.A.; Engwerda, C.R.; et al. Hookworm secreted extracellular vesicles interact with host cells and prevent inducible Colitis in mice. Front. Immunol. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Eichenberger, R.M.; Talukder, M.H.; Field, M.A.; Wangchuk, P.; Giacomin, P.; Loukas, A.; Sotillo, J. Characterization of Trichuris muris secreted proteins and extracellular vesicles provides new insights into host-parasite communication. J. Extracell. Vesicles 2018, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, L.; Korchev, Y.; Bashford, L.; Djamgoz, M.; Wakelin, D.; Ashall, F.; Bundy, D. The major secreted product of the whipworm, Trichuris, is a pore-forming protein. Prec. R. Soc. Lond. B 1994, 257, 255–261. [Google Scholar]
- Bancroft, A.J.; Levy, C.W.; Jowitt, T.A.; Hayes, K.S.; Thompson, S.; McKenzie, E.A.; Ball, M.D.; Dubaissi, E.; France, A.P.; Bellina, B.; et al. The major secreted protein of the whipworm parasite tethers to matrix and inhibits interleukin-13 function. Nat. Commun. 2019, 10, 2344. [Google Scholar] [CrossRef] [Green Version]
- Williamson, A.L.; Brindley, P.J.; Knox, D.P.; Hotez, P.J.; Loukas, A. Digestive proteases of blood-feeding nematodes. Trends Parasitol. 2003, 19, 417–423. [Google Scholar] [CrossRef]
- Stassens, P.; Bergum, P.W.; Gansemans, Y.; Jespers, L.; Laroche, Y.; Huang, S.; Maki, S.; Messens, J.; Lauwereys, M.; Cappello, M.; et al. Anticoagulant repertoire of the hookworm Ancylostoma caninum. Proc. Natl. Acad. Sci. USA 1996, 93, 2149–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, S.; Pickering, D.A.; Ferreira, I.B.; Jones, L.R.S.; Troy, S.; Leech, A.; Hotez, P.J.; Zhan, B.; Laha, T.P.R.; Sparwasser, T.; et al. Hookworm recombinant protein promotes regulatory T cell responses that suppress experimental asthma. Sci. Transl. Med. 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Shepherd, C.; Navarro, S.; Wangchuk, P.; Wilson, D.; Daly, N.L.; Loukas, A. Identifying the immunomodulatory components of helminths. Parasite Immunol. 2015, 37, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Majumder, K.; Zhang, H.; Matsui, T.; Mine, Y. Adenine has an anti-inflammatory effect through the activation of adenine receptor signaling in mouse macrophage. J. Funct. Foods 2017, 28, 235–239. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Y.; Nguyen, H.G.; Koupenova, M.; Chauhan, A.K.; Makitalo, M.; Jones, M.R.; St Hilaire, C.; Seldin, D.C.; Toselli, P.; et al. The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J. Clin. Investig. 2006, 116, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Hasko, G.; Kuhel, D.G.; Nemeth, Z.H.; Mabley, J.G.; Stachlewitz, R.F.; Virag, L.; Lohinai, Z.; Southan, G.J.; Salzman, A.L.; Szabo, C. Inosine inhibits inflammatory cytokine production by a posttranscriptional mechanism and protects against endotoxin-induced shock. J. Immunol. 2000, 164, 1013–1019. [Google Scholar] [CrossRef]
- Liaudet, L.; Mabley, J.G.; Soriano, F.G.; Pacher, P.; Marton, A.; Hasko, G.; Szabo, C. Inosine reduces systemic inflammation and improves survival in septic shock induced by cecal ligation and puncture. Am. J. Respir. Crit. Care Med. 2001, 164, 1213–1220. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.J.; Lin, J.S.; Lin, Y.C.; Lin, P.T. Antiinflammatory effects of l-carnitine supplementation (1000 mg/d) in coronary artery disease patients. Nutrition 2015, 31, 475–479. [Google Scholar] [CrossRef]
- Keskin, E.; Uluisik, D.; Altin, M. Antioxidant effect of l-carnitine in rats fed cholesterol rich diet. Anim. Vet. Sci. 2015, 3, 113–116. [Google Scholar] [CrossRef]
- Detopoulou, P.; Panagiotakos, D.B.; Antonopoulou, S.; Pitsavos, C.; Stefanadis, C. Dietary choline and betaine intakes in relation to concentrations of inflammatory markers in healthy adults: The ATTICA study. Am. J. Clin. Nutr. 2008, 87, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.; Kimball, A.; Schaller, M.; denDekker, A.; Burant, C.; Kunkel, S.L.; Gallagher, K. Modulation of Xanthine metabolism ameliorates inflammation and accelerates diabetic wound healing. J. Immunol. 2018, 200, 170-13. [Google Scholar]
- Afraei, S.; D’Aniello, A.; Sedaghat, R.; Ekhtiari, P.; Azizi, G.; Tabrizian, N.; Magliozzi, L.; Aghazadeh, Z.; Mirshafiey, A. Therapeutic effects of D-aspartate in a mouse model of multiple sclerosis. J. Food. Drug. Anal. 2017, 25, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Boccella, S.; Vacca, V.; Errico, F.; Marinelli, S.; Squillace, M.; Guida, F.; Di Maio, A.; Vitucci, D.; Palazzo, E.; De Novellis, V.; et al. D-aspartate modulates nociceptive-specific neuron activity and pain threshold in inflammatory and neuropathic pain condition in mice. Biomed. Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Littlewood-Evans, A.; Sarret, S.; Apfel, V.; Loesle, P.; Dawson, J.; Zhang, J.; Muller, A.; Tigani, B.; Kneuer, R.; Patel, S.; et al. GPR91 senses extracellular succinate released from inflammatory macrophages and exacerbates rheumatoid arthritis. J. Exp. Med. 2016, 213, 1655–1662. [Google Scholar] [CrossRef]
- Lei, W.; Ren, W.; Ohmoto, M.; Urban, J.F., Jr.; Matsumoto, I.; Margolskee, R.F.; Jiang, P. Activation of intestinal tuft cell-expressed Sucnr1 triggers type 2 immunity in the mouse small intestine. Proc. Natl. Acad. Sci. USA 2018, 115, 5552–5557. [Google Scholar] [CrossRef] [Green Version]
- Fujino, M.; Nishio, Y.; Ito, H.; Tanaka, T.; Li, X.K. 5-Aminolevulinic acid regulates the inflammatory response and alloimmune reaction. Int. Immunopharmacol. 2016, 37, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Maruyama, T.; Urade, Y.; Nagata, S. Immunosuppression via adenosine receptor activation by adenosine monophosphate released from apoptotic cells. Elife 2014, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sag, D.; Carling, D.; Stout, R.D.; Suttles, J. Adenosine 5’-monophosphate-activated protein kinase promotes macrophage polarization to an anti-inflammatory functional phenotype. J. Immunol. 2008, 181, 8633–8641. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Wang, R.X.; Alexeev, E.E.; Lanis, J.M.; Battista, K.D.; Glover, L.E.; Colgan, S.P. Hypoxanthine is a checkpoint stress metabolite in colonic epithelial energy modulation and barrier function. J. Biol. Chem. 2018, 293, 6039–6051. [Google Scholar] [CrossRef] [Green Version]
- Jiao, N.; Wu, Z.; Ji, Y.; Wang, B.; Dai, Z.; Wu, G. l-Glutamate enhances barrier and antioxidative functions in intestinal porcine epithelial cells. J. Nutr. 2015, 145, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Unnikrishnan, M.K.; Rao, M.N.A. Antiinflammatory activity of methionine, methionine sulfoxide and methionine sulfone. Agents Actions 1990, 31, 110–112. [Google Scholar] [CrossRef]
- Derakhshanfar, A.; Bidadkosh, A.; Hashempour Sadeghian, M. l-methionine attenuates gentamicin nephrotoxicity in male Wistar rat: Pathological and biochemical findings. Iran. J. Vet. Res. 2009, 10, 323–328. [Google Scholar]
- Hasegawa, S.; Ichiyama, T.; Sonaka, I.; Ohsaki, A.; Okada, S.; Wakiguchi, H.; Kudo, K.; Kittaka, S.; Hara, M.; Furukawa, S. Cysteine, histidine and glycine exhibit anti-inflammatory effects in human coronary arterial endothelial cells. Clin. Exp. Immunol. 2012, 167, 269–274. [Google Scholar] [CrossRef]
- Winter, A.N.; Brenner, M.C.; Punessen, N.; Snodgrass, M.; Byars, C.; Arora, Y.; Linseman, D.A. Comparison of the neuroprotective and anti-Inflammatory effects of the anthocyanin metabolites, protocatechuic acid and 4-hydroxybenzoic acid. Oxid. Med. Cell Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ullman, B.; Gudas, L.J.; Cohen, A.; Martin, D.W. Deoxyadenosine metabolism and cytotoxicity in cultured mouse T lymphoma cells: A model for immunodeficiency disease. Cell 1978, 14, 365–375. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Huang, N.; Zhou, X.; Tian, J.; Li, T.; Wu, J.; Tian, Y.; Yin, Y.; Yao, K. Alpha-ketoglutarate suppresses the NF-κB-mediated inflammatory pathway and enhances the PXR-regulated detoxification pathway. Oncotarget 2017, 8, 102974–102988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velvizhi, S.; Nagalashmi, T.; Essa, M.M.; Dakshayani, K.B.; Subramanian, P. Effects of—Alpha-ketoglutarate on lipid peroxidation and antioxidant status during chronic ethnol administration in Wistar rats. Pol. J. Pharmacol. 2002, 54, 231–236. [Google Scholar] [PubMed]
- Cavone, L.; Calosi, L.; Cinci, L.; Moroni, F.; Chiarugi, A. Topical mannitol reduces inflammatory edema in a rat model of arthritis. Pharmacology 2012, 89, 18–21. [Google Scholar] [CrossRef]
- Naägeli, M.; Fasshauer, M.; Sommerfeld, J.; Fendel, A.; Brandi, G.; Stover, J.F. Prolonged continuous intravenous infusion of the dipeptide l-alanine- l-glutamine significantly increases plasma glutamine and alanine without elevating brain glutamate in patients with severe traumatic brain injury. Nägeli Crit. Care 2014, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Coqueiro, A.Y.; Raizel, R.; Hypólito, T.M.; Tirapegui, J. Effects of supplementation with l-glutamine and l-alanine in the body composition of rats submitted to resistance exercise. Rev. Bras. Cienc. Esporte 2017, 39, 417–423. [Google Scholar] [CrossRef]
- Raizel, R.; Leite, J.S.; Hypolito, T.M.; Coqueiro, A.Y.; Newsholme, P.; Cruzat, V.F.; Tirapegui, J. Determination of the anti-inflammatory and cytoprotective effects of l-glutamine and l-alanine, or dipeptide, supplementation in rats submitted to resistance exercise. Br. J. Nutr. 2016, 116, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.K.; Sternbach, S.; Fleming, S.; Alkhayer, K.; Shelestak, J.; Popescu, D.; Weaver, A.; Clements, R.; Wasek, B.; Bottiglieri, T.; et al. Betaine restores epigenetic control and supports neuronal mitochondria in the cuprizone mouse model of multiple sclerosis. Epigenetics 2020, 1–16. [Google Scholar] [CrossRef]
- Wu, J.; He, C.; Bu, J.; Luo, Y.; Yang, S.; Ye, C.; Yu, S.; He, B.; Yin, Y.; Yang, X. Betaine attenuates LPS-induced downregulation of Occludin and Claudin-1 and restores intestinal barrier function. BMC Vet. Res. 2020, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodayar, M.J.; Kalantari, H.; Khorsandi, L.; Rashno, M.; Zeidooni, L. Upregulation of Nrf2- related cytoprotective genes expression by acetaminophen-induced acute hepatotoxicity in mice and the protective role of betaine. Hum. Exp. Toxicol. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Wu, C.; Li, P.; Li, N.; Zhang, D.; Zhu, Q.; Ren, W.; Peng, Y. Functions and signaling pathways of amino acids in intestinal inflammation. Biomed. Res. Int. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Han, H.; Yin, J.; Wang, B.; Huang, X.; Yao, J.; Zheng, J.; Fan, W.; Li, T.; Yin, Y. Effects of dietary lysine restriction on inflammatory responses in piglets. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Wang, C.; Ding, L.; Shen, Y.; Cui, H.; Wang, M.; Wang, H. Arginine relieves the inflammatory response and enhances the casein expression in bovine mammary epithelial cells induced by lipopolysaccharide. Mediat. Inflamm. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hnia, K.; Gayraud, J.; Hugon, G.; Ramonatxo, M.; De La Porte, S.; Matecki, S.; Mornet, D. l-arginine decreases inflammation and modulates the nuclear factor-kappaB/matrix metalloproteinase cascade in mdx muscle fibers. Am. J. Pathol. 2008, 172, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Coburn, L.A.; Gong, X.; Singh, K.; Asim, M.; Scull, B.P.; Allaman, M.M.; Williams, C.S.; Rosen, M.J.; Washington, M.K.; Barry, D.P.; et al. l-arginine supplementation improves responses to injury and inflammation in dextran sulfate sodium colitis. PLoS ONE 2012, 7, e33546. [Google Scholar] [CrossRef]
- Alamshah, A.; Spreckley, E.; Norton, M.; Kinsey-Jones, J.S.; Amin, A.; Ramgulam, A.; Cao, Y.; Johnson, R.; Saleh, K.; Akalestou, E.; et al. l-phenylalanine modulates gut hormone release and glucose tolerance, and suppresses food intake through the calcium-sensing receptor in rodents. Int. J. Obes. 2017, 41, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Oldenburger, A.; Roscioni, S.S.; Jansen, E.; Menzen, M.H.; Halayko, A.J.; Timens, W.; Meurs, H.; Maarsingh, H.; Schmidt, M. Anti-inflammatory role of the cAMP effectors Epac and PKA: Implications in chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e31574. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Li, P.; Bi, W.C.; Ma, R.; Yin, Y.; Bi, K.; Li, Q. Plasma N-acetylputrescine, cadaverine and 1,3-diaminopropane: Potential biomarkers of lung cancer used to evaluate the ef cacy of anticancer drugs. Oncotarget 2017, 8, 88575–88585. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.N.; Pendse, V.K.; Khanna, N.K. Anti-inflammatory and analgesic properties of four amino-acids. Indian J. Physiol. Pharmacol. 1984, 28, 299–305. [Google Scholar]
- Kato, H.; Miura, K.; Nakano, S.; Suzuki, K.; Bannai, M.; Inoue, Y. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids 2016, 48, 2145–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safer, D.; Brenes, M.; Dunipace, S.; Schad, S. Urocanic acid is a major chemoattractant for the skin-penetrating parasitic nematode Strongyloides stercoralis. Proc. Natl. Acad. Sci. USA 2007, 104, 1627–1630. [Google Scholar] [CrossRef] [Green Version]
- Mastrofrancesco, A.; Ottaviani, M.; Aspite, N.; Cardinali, G.; Izzo, E.; Graupe, K.; Zouboulis, C.C.; Camera, E.; Picardo, M. Azelaic acid modulates the inflammatory response in normal human keratinocytes through PPARgamma activation. Exp. Dermatol. 2010, 19, 813–820. [Google Scholar] [CrossRef]
- Andersen, F.A. Final report on the safety assessment of Maleic Acid. Int. J. Toxicol. 2007, 26, 125–130. [Google Scholar] [CrossRef]
- Spreafico, A.; Millucci, L.; Ghezzi, L.; Geminiani, M.; Braconi, D.; Amato, L.; Chellini, F.; Frediani, B.; Moretti, E.; Collodel, G.; et al. Antioxidants inhibit SAA formation and pro-inflammatory cytokine release in a human cell model of alkaptonuria. Rheumatology 2013, 52, 1667–1673. [Google Scholar] [CrossRef] [Green Version]
- Hegedus, Z.L.; Nayak, U. Homogentisic acid and structurally related compounds as intermediates in plasma soluble melanin formation and in tissue toxicities. Arch. Int. Physiol. Biochim. Biophys. 1994, 102, 175–181. [Google Scholar]
- Darabi, Z.; Darand, M.; Yari, Z.; Hedayati, M.; Faghihi, A.; Agah, S.; Hekmatdoost, A. Inflammatory markers response to citrulline supplementation in patients with non-alcoholic fatty liver disease: A randomized, double blind, placebo-controlled, clinical trial. BMC Res. Notes 2019, 12, 1–5. [Google Scholar] [CrossRef]
- Ham, D.J.; Gleeson, B.G.; Chee, A.; Baum, D.M.; Caldow, M.K.; Lynch, G.S.; Koopman, R. l-Citrulline protects skeletal muscle cells from cachectic stimuli through an iNOS-dependent mechanism. PLoS ONE 2015, 10, e0141572. [Google Scholar] [CrossRef]
- Lee, Y.C.; Su, Y.T.; Liu, T.Y.; Tsai, C.M.; Chang, C.H.; Yu, H.R. l-arginine and l-citrulline supplementation have different programming effect on regulatory T-cells function of infantile rats. Front. Immunol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Faure, M.; Mettraux, C.; Moennoz, D.; Godin, J.-P.; Vuichoud, J.; Rochat, F.; Breuillé, D.; Obled, C.; Corthésy-Theulaz, I. Specific amino acids increase mucin synthesis and microbiota in dextran sulfate sodium–treated rats. J. Nutr. 2006, 136, 1558–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, E.H.; Bantug, G.; Griss, T.; Condotta, S.; Johnson, R.M.; Samborska, B.; Mainolfi, N.; Suri, V.; Guak, H.; Balmer, M.L.; et al. Serine is an essential metabolite for effector T cell expansion. Cell Metab. 2017, 25, 345–357. [Google Scholar] [CrossRef]
- Gaifem, J.; Goncalves, L.G.; Dinis-Oliveira, R.J.; Cunha, C.; Carvalho, A.; Torrado, E.; Rodrigues, F.; Saraiva, M.; Castro, A.G.; Silvestre, R. l-Threonine supplementation during Colitis onset delays disease recovery. Front. Physiol. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Dong, Y.W.; Jiang, W.D.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; Zhou, X.Q.; et al. Threonine deficiency decreased intestinal immunity and aggravated inflammation associated with NF-kappaB and target of rapamycin signalling pathways in juvenile grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Br. J. Nutr. 2017, 118, 92–108. [Google Scholar] [CrossRef] [Green Version]
- Mine, Y.; Zhang, H. Calcium-sensing receptor (CaSR)-mediated anti-inflammatory effects of l-amino acids in intestinal epithelial cells. J. Agric. Food Chem. 2015, 63, 9987–9995. [Google Scholar] [CrossRef]
- Yue, Y.; Guo, Y.; Yang, Y. Effects of dietary l-tryptophan supplementation on intestinal response to chronic unpredictable stress in broilers. Amino Acids 2017, 49, 1227–1236. [Google Scholar] [CrossRef]
- Islam, J.; Sato, S.; Watanabe, K.; Watanabe, T.; Ardiansyah; Hirahara, K.; Aoyama, Y.; Tomita, S.; Aso, H.; Komai, M.; et al. Dietary tryptophan alleviates dextran sodium sulfate-induced colitis through aryl hydrocarbon receptor in mice. J. Nutr. Biochem. 2017, 42, 43–50. [Google Scholar] [CrossRef]
- Li, X.S.; Wang, Z.; Cajka, T.; Buffa, J.A.; Nemet, I.; Hurd, A.G.; Gu, X.; Skye, S.M.; Roberts, A.B.; Wu, Y.; et al. Untargeted metabolomics identifies trimethyllysine, a TMAO-producing nutrient precursor, as a predictor of incident cardiovascular disease risk. JCI Insight 2018, 3, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, A.; Gieseg, S.P. Pterins as diagnostic markers of exercise-induced stress: A systematic review. J. Sci. Med. Sport. 2020, 23, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pederzolli, C.D.; Mescka, C.P.; Zandona, B.R.; de Moura Coelho, D.; Sgaravatti, A.M.; Sgarbi, M.B.; de Souza Wyse, A.T.; Duval Wannmacher, C.M.; Wajner, M.; Vargas, C.R.; et al. Acute administration of 5-oxoproline induces oxidative damage to lipids and proteins and impairs antioxidant defenses in cerebral cortex and cerebellum of young rats. Metab. Brain Dis. 2010, 25, 145–154. [Google Scholar] [CrossRef]
- Kalinowska, M.; Mazur, L.; Jabłońska-Trypuć, A.; Lewandowski, W. A new calcium 2,5-dihydroxybenzoate: Synthesis, characterization and antioxidant studies and stress mediated cytotoxity in MCF-7 cells. J. Saudi Chem. Soc. 2018, 22, 742–756. [Google Scholar] [CrossRef]
- Pearson, M.S.; Tribolet, L.; Cantacessi, C.; Periago, M.V.; Valero, M.A.; Jariwala, A.R.; Hotez, P.; Diemert, D.; Loukas, A.; Bethony, J. Molecular mechanisms of hookworm disease: Stealth, virulence, and vaccines. J. Allergy Clin. Immunol. 2012, 130, 13–21. [Google Scholar] [CrossRef]
- Rochat, B. Proposed confidence scale and ID score in the identification of known-unknown compounds using high resolution MS data. J. Am. Soc. Mass Spectrom. 2017, 28, 709–723. [Google Scholar] [CrossRef]
- Gika, H.G.; Theodoridis, G.A.; Plumb, R.S.; Wilson, I.D. Current practice of liquid chromatography-mass spectrometry in metabolomics and metabonomics. J. Pharm. Biomed. Anal. 2014, 87, 12–25. [Google Scholar] [CrossRef]
- Cajka, T.; Vaclavikova, M.; Dzuman, Z.; Vaclavik, L.; Ovesna, J.; Hajslova, J. Rapid LC-MS-based metabolomics method to study the Fusarium infection of barley. J. Sep. Sci. 2014, 37, 912–919. [Google Scholar] [CrossRef]
- Barrett, J. Developmental aspects of metabolism in parasites. Int. J. Parasitol. 1987, 17, 105–110. [Google Scholar] [CrossRef]
- Bennuru, S.; Lustigman, S.; Abraham, D.; Nutman, T.B. Metabolite profiling of infection-associated metabolic markers of onchocerciasis. Mol. Biochem. Parasitol. 2017, 215, 58–69. [Google Scholar] [CrossRef]
- Ryan, S.M.; Eichenberger, R.M.; Ruscher, R.; Giacomin, P.R.; Loukas, A. Harnessing helminth-driven immunoregulation in the search for novel therapeutic modalities. PLoS Pathog. 2020, 16, e1008508. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Saric, J.; Li, J.V.; Utzinger, J.; Wang, Y.; Keiser, J.; Dirnhofer, S.; Beckonert, O.; Sharabiani, M.T.; Fonville, J.M.; Nicholson, J.K.; et al. Systems parasitology: Effects of Fasciola hepatica on the neurochemical profile in the rat brain. Mol. Syst. Biol. 2010, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Beis, I.; Barrett, J. Energy metabolism in developing Ascaris lumbricoides eggs II. The steady state content of intermediary metabolites. Dev. Biol. 1975, 42, 188–195. [Google Scholar] [CrossRef]
- Wangchuk, P.; Shepherd, C.; Constantinoiu, C.; Ryan, R.Y.M.; Kouremenos, K.A.; Becker, L.; Jones, L.; Buitrago, G.; Giacomin, P.; Wilson, D.; et al. Hookworm-derived metabolites suppress pathology in a mouse model of colitis and inhibit secretion of key inflammatory cytokines in primary human leukocytes. Infect. Immun. 2019, 87, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmett, M. Acetaminophen toxicity and 5-oxoproline (pyroglutamic acid): A tale of two cycles, one an ATP-depleting futile cycle and the other a useful cycle. Clin. J. Am. Soc. Nephrol. 2014, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Liss, D.B.; Paden, M.S.; Schwarz, E.S.; Mullins, M.E. What is the clinical significance of 5-oxoproline (pyroglutamic acid) in high anion gap metabolic acidosis following paracetamol (acetaminophen) exposure? Clin. Toxicol. 2013, 51, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Loffler, M.; Carrey, E.A.; Zameitat, E. Orotate (orotic acid): An essential and versatile molecule. Nucleos. Nucleot. Nucl. 2016, 35, 566–577. [Google Scholar] [CrossRef]
- Hill, B.; Kilsby, J.; Rogerson, G.W.; Mcintosh, R.T.; Ginger, C.D. The enzymes of pyrimidine biosynthesis in a range of parasitic protozoa and helminths. Mol. Biochem. Parasitol. 1981, 2, 123–134. [Google Scholar] [CrossRef]
- Longo, N. Disorders of biopterin metabolism. J. Inherit. Metab. Dis. 2009, 32, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Arias-Barrau, E.; Olivera, E.R.; Luengo, J.M.; Fernandez, C.; Galan, B.; Garcia, J.L.; Diaz, E.; Minambres, B. The homogentisate pathway: A central catabolic pathway involved in the degradation of l-phenylalanine, l-tyrosine, and 3-hydroxyphenylacetate in Pseudomonas putida. J. Bacteriol. 2004, 186, 5062–5077. [Google Scholar] [CrossRef] [Green Version]
- Sterkel, M.; Perdomo, H.D.; Guizzo, M.G.; Barletta, A.B.; Nunes, R.D.; Dias, F.A.; Sorgine, M.H.; Oliveira, P.L. Tyrosine detoxification is an essential trait in the life history of blood-feeding arthropods. Curr. Biol. 2016, 26, 2188–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.L.; Li, H.T.; Zhang, L.J.; Lin, Z.H.; Kuo, Y.H. Conversion of squid pen to homogentisic acid via Paenibacillus sp. TKU036 and the antioxidant and anti-inflammatory activities of homogentisic acid. Mar. Drugs 2016, 14, 183. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.E.; MacDonald, A.S. Profound suppression of cellular proliferation mediated by the secretions of nematodes. Parasite Immunol. 1998, 20, 241–247. [Google Scholar] [CrossRef]
- Porth, C. Essentials of Pathophysiology: Concepts of Altered Health States; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2011; pp. 1–1248. [Google Scholar]
- Peachey, L.E.; Castro, C.; Molena, R.A.; Jenkins, T.P.; Griffin, J.L.; Cantacessi, C. Dysbiosis associated with acute helminth infections in herbivorous youngstock - observations and implications. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Fotiadis, C.; Adamis, S.; Misiakos, E.P.; Genetzakis, M.; Antonakis, P.T.; Tsekouras, D.K.; Gorgoulis, V.G.; Zografos, G.C.; Papalois, A.; Fotinou, M.; et al. The prophylactic effect of l-arginine in acute ischaemic Colitis in a rat model of ischaemia/reperfusion injury. Acta Chir. Belg. 2016, 107, 192–200. [Google Scholar] [CrossRef]
- Zhu, H.L.; Liu, Y.L.; Xie, X.L.; Huang, J.J.; Hou, Y.Q. Effect of l-arginine on intestinal mucosal immune barrier function in weaned pigs after Escherichia coli LPS challenge. Innate Immun. 2013, 19, 242–252. [Google Scholar] [CrossRef]
- Sato, S.; Hirayama, T.; Hirazawa, N. Lipid content and fatty acid composition of the monogenean Neobenedenia girellae and comparison between the parasite and host fish species. Parasitology 2008, 135, 967–975. [Google Scholar] [CrossRef]
- Zeng, C.; Wen, B.; Hou, G.; Lei, L.; Mei, Z.; Jia, X.; Chen, X.; Zhu, W.; Li, J.; Kuang, Y.; et al. Lipidomics profiling reveals the role of glycerophospholipid metabolism in psoriasis. Gigascience 2017, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.F. Aspects of helminth metabolism. Parasitology 1982, 84, 177–194. [Google Scholar] [CrossRef]
- Wang, T.; Nie, S.; Ma, G.; Korhonen, P.K.; Koehler, A.V.; Ang, C.S.; Reid, G.E.; Williamson, N.A.; Gasser, R.B. The developmental lipidome of Haemonchus contortus. Int. J. Parasitol. 2018, 48, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Henry, P.; Owopetu, O.; Adisa, D.; Nguyen, T.; Anthony, K.; Ijoni-Animadu, D.; Jamadar, S.; Abdel-Rahman, F.; Saleh, M.A. Fatty acids composition of Caenorhabditis elegans using accurate mass GCMS-QTOF. J. Environ. Sci. Health B 2016, 51, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Sinensky, M. Homeoviscous adaptation—A homeostatic process that regulates the viscosity of membrane lipids in Escherichia Coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.K.; Li, Z.; Lu, A.Y.; Sun, F.; Chen, S.; Rothe, M.; Menzel, R.; Sun, F.; Horvitz, H.R. Acyl-CoA Dehydrogenase Drives Heat Adaptation by Sequestering Fatty Acids. Cell 2015, 161, 1152–1163. [Google Scholar] [CrossRef] [Green Version]
- Mangmee, S.; Adisakwattana, P.; Tipthara, P.; Simanon, N.; Sonthayanon, P.; Reamtong, O. Lipid profile of Trichinella papuae muscle-stage larvae. Sci. Rep. 2020, 10, 10125. [Google Scholar] [CrossRef]
- Wewer, V.; Makepeace, B.L.; Tanya, V.N.; Peisker, H.; Pfarr, K.; Hoerauf, A.; Dormann, P. Lipid profiling of the filarial nematodes Onchocerca volvulus, Onchocerca ochengi and Litomosoides sigmodontis reveals the accumulation of nematode-specific ether phospholipids in the host. Int. J. Parasitol. 2017, 47, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.L. Changes in adult Nippostrongylus brasiliensis during the development of immunity to this nematode in rats. 2. Total lipids and neutral lipids. Parasitology 1971, 63, 271–274. [Google Scholar] [CrossRef]
- Barrett, J. Biochemistry of Parasitic Helminths; MacMillan Publishers Ltd.: New York, NY, USA, 1981. [Google Scholar]
- Welty, F.K. How do elevated triglycerides and low HDL-cholesterol affect inflammation and atherothrombosis? Curr. Cardiol. Rep. 2013, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, Z.; Lu, M.; Yonekubo, Y.; Liang, X.; Zhang, Y.; Wu, P.; Zhou, Y.; Grinstein, S.; Hancock, J.F.; et al. Temporal production of the signaling lipid phosphatidic acid by phospholipase D2 determines the output of extracellular signal-regulated kinase signaling in cancer cells. Mol. Cell Biol. 2014, 34, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Modha, J.; Roberts, M.C.; Robertson, W.M.; Sweetman, G.; Powell, K.A.; Kennedy, M.W.; Kusel, J.R. The surface coat of infective larvae of Trichinella spiralis. Parasitology 1999, 118, 509–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nervi, F. Significance of biliary phospholipids for maintenance of the gastrointestinal mucosal barrier and hepatocellular integrity. Gastroenterology 2000, 118, 1265–1267. [Google Scholar] [CrossRef]
- Stremmel, W.; Merle, U.; Zahn, A.; Autschbach, F.; Hinz, U.; Ehehalt, R. Retarded release phosphatidylcholine benefits patients with chronic active ulcerative colitis. Gut 2005, 54, 966–971. [Google Scholar] [CrossRef]
- Treede, I.; Braun, A.; Sparla, R.; Kuhnel, M.; Giese, T.; Turner, J.R.; Anes, E.; Kulaksiz, H.; Fullekrug, J.; Stremmel, W.; et al. Anti-inflammatory effects of phosphatidylcholine. J. Biol. Chem. 2007, 282, 27155–27164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camberis, M.; Le Gros, G.; Urban, J.J. Animal model of Nippostrongylus brasiliensis and Heligmosomoides polygyrus. Curr. Protoc. Immunol. 2003, 55, 19.12.11–19.12.27. [Google Scholar]
- Creek, D.J.; Chua, H.H.; Cobbold, S.A.; Nijagal, B.; MacRae, J.I.; Dickerman, B.K.; Gilson, P.R.; Ralph, S.A.; McConville, M.J. Metabolomics-based screening of the malaria box reveals both novel and established mechanisms of action. Antimicrob. Agents Chemother. 2016, 60, 6650–6663. [Google Scholar] [CrossRef] [Green Version]
- Aurelio, L.; Scullino, C.V.; Pitman, M.R.; Sexton, A.; Oliver, V.; Davies, L.; Rebello, R.J.; Furic, L.; Creek, D.J.; Pitson, S.M.; et al. From sphingosine kinase to dihydroceramide desaturase: A structure-activity relationship (SAR) study of the enzyme inhibitory and anticancer activity of 4-((4-(4-chlorophenyl)thiazol-2-yl)amino)phenol (SKI-II). J. Med. Chem. 2016, 59, 965–984. [Google Scholar] [CrossRef] [PubMed]
- Creek, D.J.; Jankevics, A.; Burgess, K.E.; Breitling, R.; Barrett, M.P. IDEOM: An Excel interface for analysis of LC-MS-based metabolomics data. Bioinformatics 2012, 28, 1048–1049. [Google Scholar] [CrossRef]
- Scheltema, R.A.; Jankevics, A.; Jansen, R.C.; Swertz, M.A.; Breitling, R. PeakML/mzMatch: A file format, Java library, R library, and tool-chain for mass spectrometry data analysis. Anal. Chem. 2011, 83, 2786–2793. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polar Metabolites | Formula a | Mass (m/z) | Rt (min) c | KEGG ID d | Log2(FC) e | Chemical Taxonomy b | Reported Pharmacological Activities | Average Peak Area (mz/rt) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| NB_SE | NB_ESP | TM_SE | ||||||||
| Adenine | C5H5N5 | 135.054 | 8.32 | C00147 | 5.23 | 6-aminopurines | Anti-inflammatory [32] | 103,160,746 | 309,024 | 101,274 |
| Adenosine | C10H13N5O4 | 267.096 | 8.23 | C00212 | 5.21 | Purine nucleosides | Anti-inflammatory [33] | 327,057,005 | 0 | 347,270 |
| Inosine | C10H12N4O5 | 268.080 | 10.32 | C00294 | 4.77 | Purine nucleosides | Anti-inflammatory [34,35] | 14,631,676 | 0 | 18,472 |
| l-Carnitine | C7H15NO3 | 161.105 | 12.32 | C00487 | 4.56 | Carnitines | Anti-inflammatory [36] and anti-oxidant [37] | 81,471,762 | 1,911,162 | 125,132 |
| Choline | C5H13NO | 103.099 | 20.72 | C00114 | 4.53 | Cholines | Anti-inflammatory [38] | 82,418,790 | 0 | 129,808 |
| N(pi)-Methyl-l-histidine | C7H11N3O2 | 169.085 | 11.58 | C01152 | 3.86 | Histidine and derivatives | N/A | 5,268,444 | 2,170,489 | 13,260 |
| Xanthine | C5H4N4O2 | 152.033 | 11.08 | C00385 | 3.62 | Xanthines | Proinflammatory [39] | 37,256,722 | 3,609,773 | 103,264 |
| l-Aspartate | C4H7NO4 | 133.037 | 14.57 | C00402 | 3.52 | Aspartic acid and derivatives | Anti-inflammatory and neuroprotective [40,41] | 3,886,437 | 402,353 | 9321 |
| Succinate | C4H6O4 | 118.026 | 15.15 | C00042 | 3.14 | Dicarboxylic acids and derivatives | Activate inflammatory pathways [42,43] | 29,148,318 | 5,130,431 | 99,396 |
| 5-Aminolevulinate | C5H9NO3 | 131.058 | 13.74 | C00430 | 3.11 | Delta amino acids and derivatives | Anti-inflammatory [44] | 8,231,888 | 0 | 33,627 |
| Adenosine 5′-monophosphate | C10H14N5O7P | 347.063 | 13.03 | C00020 | 3.05 | Purine ribonucleoside monophosphates | Anti-inflammatory [45,46] | 9,485,566 | 0 | 43,938 |
| Hypoxanthine | C5H4N4O | 136.038 | 9.56 | C00262 | 2.84 | Hypoxanthines | Anti-inflammatory and wound healing [47] | 48,567,229 | 425,871 | 230,726 |
| Lactose | C12H22O11 | 342.116 | 15.25 | C00243 | 2.61 | O-glycosyl compounds | N/A | 94,498,840 | 420,550 | 552,215 |
| l-Glutamate | C5H9NO4 | 147.053 | 14.24 | C00025 | 2.30 | Glutamic acid and derivatives | Antioxidant [48] | 16,940,920 | 4,993,538 | 104,717 |
| l-Methionine | C5H11NO2S | 149.051 | 10.81 | C00073 | 2.06 | Methionine and derivatives | Anti-inflammatory [49] and antioxidant [50] | 2,078,174 | 890,947 | 15,017 |
| l-Histidine | C6H9N3O2 | 155.069 | 14.25 | C00135 | 1.82 | Histidine and derivatives | Anti-inflammatory [51] | 14,964,799 | 1,089,738 | 135,916 |
| 4-Hydroxybenzoate | C7H6O3 | 138.031 | 10.37 | C00156 | 1.73 | Hydroxybenzoic acid derivatives | Neuroprotective [52] | 1,566,927 | 1,019,203 | 13,620 |
| l-Tyrosine | C9H11NO3 | 181.074 | 12.44 | C00082 | 1.09 | Tyrosine and derivatives | N/A | 3,708,634 | 6,686,668 | 58,822 |
| Deoxyadenosine | C10H13N5O3 | 251.101 | 7.48 | C00559 | −1.03 | Purine 2′-deoxyribonucleosides | Cell growth inhibitor and cytotoxic [53] | 628,485 | 0 | 39,096 |
| (S)-Malate | C4H6O5 | 134.021 | 16.18 | C00149 | −1.28 | Beta hydroxy acids and derivatives | N/A | 51,344,999 | 43,880,026 | 0 |
| D-Glucose 6-phosphate | C6H13O9P | 260.029 | 15.83 | C00092 | −1.66 | Hexose phosphates | N/A | 402,720 | 0 | 42,982 |
| 2-Oxoglutarate | C5H6O5 | 146.021 | 15.77 | C00026 | −1.80 | Gamma-keto acids and derivatives | Anti-inflammatory [54] and antioxidant [55] | 249,067 | 2,843,652 | 0 |
| l-Pipecolate | C6H11NO2 | 129.079 | 11.42 | C00408 | −1.94 | l-alpha-amino acids | N/A | 139,920 | 0 | 16,319 |
| Mannitol | C6H14O6 | 182.079 | 13.30 | C00392 | −2.08 | Sugar alcohol | Anti-edema [56] | 299,808 | 0 | 37,781 |
| l-Alanine | C3H7NO2 | 89.047 | 14.12 | C00041 | −2.27 | l-alpha-amino acids | Anti-inflammatory [57,58,59] | 3,102,359 | 3,529,005 | 0 |
| Betaine | C5H11NO2 | 117.079 | 10.39 | C00719 | −2.39 | Alpha amino acids | Neuroprotective [60]; improves intestinal barrier function [61]; hepatoprotective [62]; anti-inflammatory [38] | 261,777,360 | 56,072,188 | 0 |
| l-Lysine | C6H14N2O2 | 146.105 | 22.50 | C00047 | −2.45 | l-alpha-amino acids | Anti-inflammatory [63,64] | 2,260,735 | 1,941,102 | 0 |
| l-Arginine | C6H14N4O2 | 174.111 | 24.05 | C00062 | −2.47 | l-alpha-amino acids | Anti-inflammatory [65,66,67] | 30,934,546 | 3,589,173 | 0 |
| l-Glutamine | C5H10N2O3 | 146.069 | 14.44 | C00064 | −2.48 | l-alpha-amino acids | Anti-inflammatory [57,58,59] | 45,487,251 | 39,780,748 | 0 |
| l-2-Aminoadipate | C6H11NO4 | 161.068 | 14.47 | C00956 | −2.49 | l-alpha-amino acids | N/A | 137,122 | 0 | 22,389 |
| l-Phenylalanine | C9H11NO2 | 165.079 | 9.35 | C00079 | −2.50 | l-alpha-amino acids | Anti-diabetic [68] | 1,534,294 | 18,355,000 | 0 |
| 3’,5’-Cyclic AMP | C10H12N5O6P | 329.052 | 8.72 | C00575 | −2.54 | 3′,5′-cyclic purine nucleotides | Anti-inflammatory [69] | 281,873 | 0 | 57,179 |
| N-Acetylputrescine | C6H14N2O | 130.110 | 21.02 | C02714 | −2.58 | Carboximidic acids | Lung cancer biomarker [70] | 125,036 | 0 | 22,355 |
| Thymine | C5H6N2O2 | 126.043 | 6.96 | C00178 | −3.62 | Hydroxypyrimidines | N/A | 169,388 | 0 | 64,297 |
| l-Leucine | C6H13NO2 | 131.094 | 9.97 | C00123 | −4.14 | Leucine and derivatives | Analgesic and anti-inflammatory [71,72] | 1,593,314 | 10,121,672 | 735,972 |
| Urocanate | C6H6N2O2 | 138.043 | 10.49 | C00785 | −4.16 | Imidazolyl carboxylic acids and derivatives | Chemoattractant [73] | 452,897 | 169,049 | 232,883 |
| Azelaic acid | C9H16O4 | 188.105 | 10.69 | C08261 | −4.89 | Medium-chain fatty acids | Anti-inflammatory [74] | 131,028 | 874,645 | 116,635 |
| Maleic acid | C4H4O4 | 116.011 | 12.18 | C01384 | −5.67 | Dicarboxylic acids and derivatives | Inflammatory/Cytotoxic [75] | 98,623 | 0 | 154,549 |
| D-Glycerate | C3H6O4 | 106.026 | 11.85 | C00258 | ns | Sugar acids and derivatives | N/A | 343,824 | 0 | 8833 |
| Homogentisate | C8H8O4 | 168.042 | 9.42 | C00544 | ns | 2(hydroxyphenyl)acetic acids | Pro-inflammatory [76,77] | 0 | 640,416 | 0 |
| Isocitrate | C6H8O7 | 192.027 | 19.04 | C00311 | ns | Tricarboxylic acids and derivatives | N/A | 0 | 37,798,044 | 0 |
| l-Citrulline | C6H13N3O3 | 175.095 | 14.84 | C00327 | ns | l-alpha-amino acids | Anti-inflammatory and antioxidant [78,79,80] | 672,469 | 0 | 21,477 |
| l-Proline | C5H9NO2 | 115.063 | 11.96 | C00148 | ns | Proline and derivatives | Anti-inflammatory [81] | 9,760,345 | 1,106,588 | 269,125 |
| l-Serine | C3H7NO3 | 105.042 | 15.56 | C00065 | ns | Serine and derivatives | Modulates adaptive immunity by controlling T cell proliferative capacity [82]; colon protection and mucosal healing [81] | 337,344 | 45,878 | 6178 |
| l-Threonine | C4H9NO3 | 119.058 | 14.14 | C00188 | ns | l-alpha-amino acids | Anti-inflammatory [83,84] | 425,688 | 0 | 14,375 |
| l-Tryptophan | C11H12N2O2 | 204.090 | 11.03 | C00078 | ns | Indolyl carboxylic acids and derivatives | Anti-inflammatory [85,86,87] | 2,002,647 | 1,435,161 | 127,849 |
| l-Valine | C5H11NO2 | 117.079 | 11.76 | C00183 | ns | Valine and derivatives | Anti-inflammatory [85] | 474,387 | 9,518,904 | 25,791 |
| Ll-2,6-Diaminoheptanedioate | C7H14N2O4 | 190.095 | 17.63 | C00666 | ns | Amino acid | N/A | 0 | 115,608 | 0 |
| N6,N6,N6-Trimethyl-l-lysine | C9H20N2O2 | 188.152 | 21.12 | C03793 | ns | l-alpha-amino acids | Cardiovascular disease biomarker [88] | 462,527 | 208,659 | 12,965 |
| Orotate | C5H4N2O4 | 156.017 | 10.27 | C00295 | ns | Pyrimidinecarboxylic acids | N/A | 0 | 437,266 | 0 |
| Pterin | C6H5N5O | 163.049 | 10.30 | C00715 | ns | Pterins and derivatives | Biomarker of exercise-induced stress [89] | 0 | 460,935 | 0 |
| Pyridoxal | C8H9NO3 | 167.058 | 7.46 | C00250 | ns | Pyridoxals and derivatives | N/A | 323,537 | 0 | 10,226 |
| 5-Oxoproline | C5H7NO3 | 129.042 | 9.82 | C01879 | ns | Alpha amino acids and derivatives | Promotes oxidative stress in neuropathology [90] | 0 | 32,495,730 | 0 |
| 2,5-Dihydroxybenzoate | C7H6O4 | 154.026 | 8.30 | C00628 | ns | Hydroxybenzoic acid derivatives | Anti-cancer activity [91] | 0 | 1,377,617 | 0 |
| 4-Trimethylammoniobutanoate | C7H15NO2 | 145.110 | 12.25 | C01181 | ns | Straight chain fatty acids | N/A | 217,954 | 0 | 2264 |
| Putative Lipids | Formula a | Mass (m/z) | Rt (min) c | Chemical Taxonomy b | LipidMAPS ID d | Peak Areas (mz/rt) | ||
|---|---|---|---|---|---|---|---|---|
| NB_SE | NB_ESP | TM_SE | ||||||
| DG(41:7) | C44H72O5 | 680.536 | 16.05 | Glycerolipids | LMGL02010545 | 124,404 | 0 | 0 |
| FA hydroxy(12:0) dodecanoic acid | C12H24O3 | 238.155 | 2.19 | Fatty Acyls | NA | 0 | 0 | 51,014 |
| LacCer(d38:0) | C50H97NO13 | 919.696 | 18.98 | Sphingolipids | LMSP05010122 | 139,285 | 0 | 0 |
| LacCer(d40:0) | C52H101NO13 | 947.727 | 20.22 | Sphingolipids | LMSP05010124 | 182,175 | 0 | 0 |
| LysoPE(22:2) | C27H52NO7P | 533.350 | 4.84 | Glycerophospholipids | LMGP02050024 | 136,295 | 0 | 0 |
| PE-Cer(d40:1) | C42H85N2O7P | 760.610 | 13.49 | Sphingolipids | LMSP03020086 | 117,114 | 0 | 0 |
| PE-Cer(d38:1) | C40H81N2O7P | 732.579 | 12.26 | Sphingolipids | LMSP03020064 | 112,168 | 0 | 0 |
| PA(25:0) | C28H55O8P | 550.364 | 12.05 | Glycerophospholipids | LMGP10010001 | 0 | 0 | 374,621 |
| PA(26:0) | C29H57O8P | 564.379 | 12.58 | Glycerophospholipids | LMGP10010980 | 0 | 0 | 1,921,006 |
| PC(36:7) | C44H74NO8P | 775.514 | 10.73 | Glycerophospholipids | LMGP01012100 | 105,372 | 0 | 0 |
| PC(P-32:2) | C40H76NO7P | 713.536 | 13.07 | Glycerophospholipids | NA | 657,868 | 0 | 0 |
| PC(P-36:2) | C44H84NO7P | 769.599 | 14.95 | Glycerophospholipids | LMGP01030137 | 138,297 | 0 | 0 |
| PE(28:2) | C33H62NO8P | 631.422 | 7.68 | Glycerophospholipids | LMGP02011238 | 127,324 | 0 | 0 |
| PE(48:2) | C53H102NO8P | 911.734 | 20.75 | Glycerophospholipids | LMGP02010893 | 466,471 | 0 | 0 |
| PE(40:5) | C45H80NO8P | 815.542 | 12.76 | Glycerophospholipids | LMGP02010893 | 483,243 | 0 | 0 |
| PE(48:1) | C53H104NO8P | 913.751 | 21.37 | Glycerophospholipids | NA | 1,154,195 | 0 | 0 |
| PE(O-20:0) | C25H54NO6P | 495.370 | 7.71 | Glycerophospholipids | LMGP02060005 | 108,008 | 0 | 0 |
| PE(P-20:0) | C25H52NO6P | 493.354 | 7.86 | Glycerophospholipids | LMGP02070004 | 375,557 | 0 | 0 |
| PE(P-36:4) | C41H74NO7P | 723.521 | 12.51 | Glycerophospholipids | LMGP02030093 | 195,178 | 0 | 0 |
| PE(P-36:5) | C41H72NO7P | 721.504 | 11.93 | Glycerophospholipids | LMGP02030028 | 87,036 | 0 | 0 |
| PE(P-38:6) | C43H74NO7P | 747.519 | 12.36 | Glycerophospholipids | LMGP02030001 | 308,137 | 0 | 0 |
| PG(36:1) | C42H81O10P | 776.557 | 11.98 | Glycerophospholipids | LMGP04010037 | 699,995 | 0 | 0 |
| PI(37:6) | C46H77O13P | 868.512 | 9.51 | Glycerophospholipids | LMGP06010790 | 119,218 | 0 | 0 |
| PI(38:7) | C47H77O13P | 880.512 | 9.44 | Glycerophospholipids | LMGP06010792 | 168,877 | 0 | 0 |
| PI(P-37:2) | C46H85O12P | 860.576 | 11.78 | Glycerophospholipids | LMGP06030067 | 94,153 | 0 | 0 |
| PS(28:2) | C34H62NO10P | 675.412 | 6.97 | Glycerophospholipids | LMGP03010919 | 253,811 | 0 | 0 |
| PS(36:4) | C42H74NO10P | 783.506 | 10.48 | Glycerophospholipids | LMGP03010038 | 141,937 | 0 | 0 |
| PS(36:5) | C42H72NO10P | 781.491 | 10.06 | Glycerophospholipids | LMGP03010654 | 115,385 | 0 | 0 |
| PS(O-38:0) | C44H88NO9P | 805.621 | 15.40 | Glycerophospholipids | LMGP03020051 | 86,582 | 0 | 0 |
| PS(O-34:0) | C40H80NO9P | 749.558 | 12.46 | Glycerophospholipids | LMGP03020043 | 0 | 0 | 1883 |
| SM(d41:2) | C46H91N2O6P | 844.667 | 14.55 | Sphingolipids | LMSP03010074 | 29,023 | 0 | 0 |
| SM(d42:2) | C47H93N2O6P | 812.676 | 15.24 | Sphingolipids | LMSP03010007 | 78,211 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeshi, K.; Creek, D.J.; Anderson, D.; Ritmejerytė, E.; Becker, L.; Loukas, A.; Wangchuk, P. Metabolomes and Lipidomes of the Infective Stages of the Gastrointestinal nematodes, Nippostrongylus brasiliensis and Trichuris muris. Metabolites 2020, 10, 446. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10110446
Yeshi K, Creek DJ, Anderson D, Ritmejerytė E, Becker L, Loukas A, Wangchuk P. Metabolomes and Lipidomes of the Infective Stages of the Gastrointestinal nematodes, Nippostrongylus brasiliensis and Trichuris muris. Metabolites. 2020; 10(11):446. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10110446
Chicago/Turabian StyleYeshi, Karma, Darren J. Creek, Dovile Anderson, Edita Ritmejerytė, Luke Becker, Alex Loukas, and Phurpa Wangchuk. 2020. "Metabolomes and Lipidomes of the Infective Stages of the Gastrointestinal nematodes, Nippostrongylus brasiliensis and Trichuris muris" Metabolites 10, no. 11: 446. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10110446