Metabolic Signatures of Gestational Weight Gain and Postpartum Weight Loss in a Lifestyle Intervention Study of Overweight and Obese Women

 ,
,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
2.1. 1H NMR Detectable Urine Metabolome Correlates with BMI in Our Study Cohort
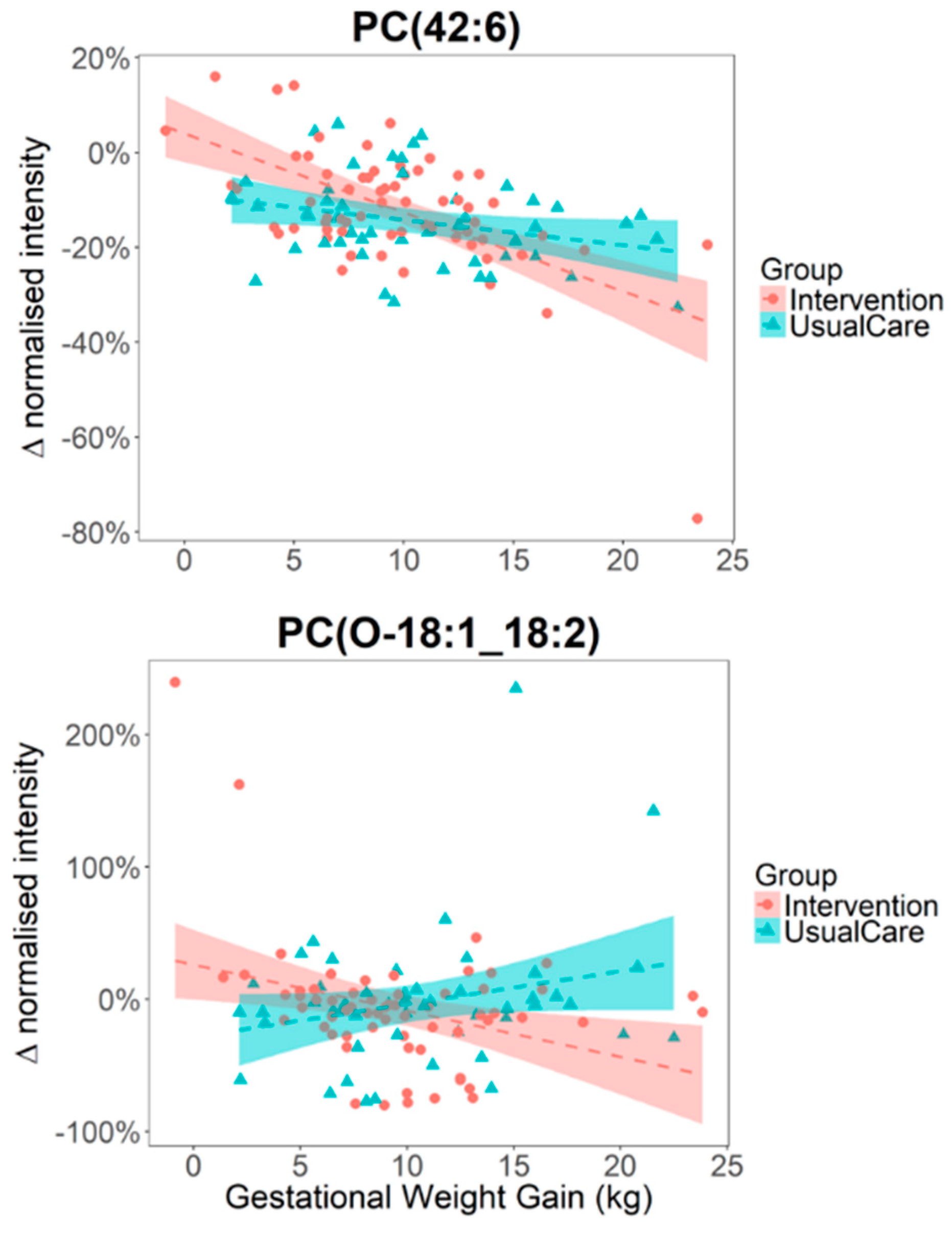
2.2. Alteration in Plasma Phospholipid Levels Are Strongly Associated with GWG and WL
2.3. The Impact of Lifestyle Intervention on Plasma Lipid Levels
3. Discussion
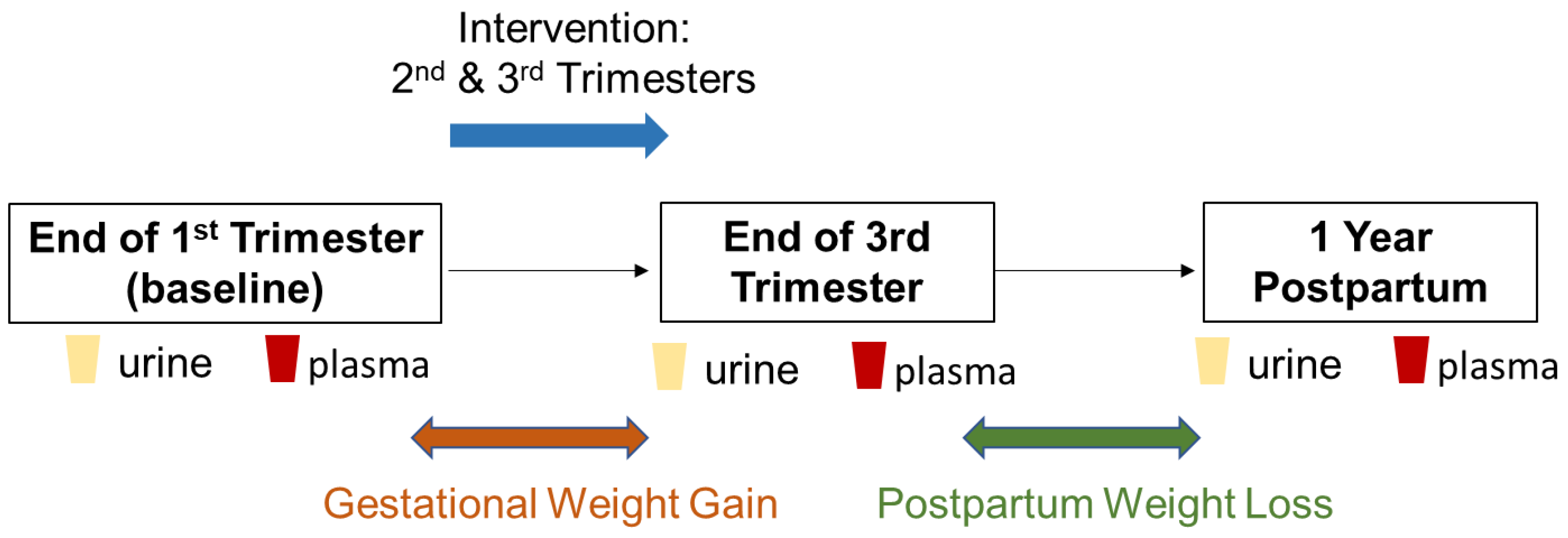
4. Materials and Methods
4.1. Participant Recruitment
4.2. Urinary 1H NMR Metabolite Measurements
4.3. LC-MS Blood Lipids Measurements
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Al-Hamad, D.; Raman, V. Metabolic syndrome in children and adolescents. Transl. Pediatrics 2017, 6, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogev, Y.; Visser, G.H.A. Obesity, gestational diabetes and pregnancy outcome. Semin. Fetal Neonatal Med. 2009, 14, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Morriseau, T.S.; Kereliuk, S.M.; Doucette, C.A.; Wicklow, B.A.; Dolinsky, V.W. Maternal obesity, diabetes during pregnancy and epigenetic mechanisms that influence the developmental origins of cardiometabolic disease in the offspring. Crit. Rev. Clin. Lab. Sci. 2018, 55, 71–101. [Google Scholar] [CrossRef] [PubMed]
- Farpour-Lambert, N.J.; Ells, L.J.; de Tejada, B.M.; Scott, C. Obesity and Weight Gain in Pregnancy and Postpartum: An Evidence Review of Lifestyle Interventions to Inform Maternal and Child Health Policies. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flegal, K.M.; Kruszon-Moran, D.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Trends in Obesity Among Adults in the United States, 2005 to 2014. JAMA J. Am. Med Assoc. 2016, 315, 2284–2291. [Google Scholar] [CrossRef] [Green Version]
- Deputy, N.P.; Dub, B.; Sharma, A.J. Prevalence and Trends in Prepregnancy Normal Weight-48 States, New York City, and District of Columbia, 2011–2015. Mmwr-Morb. Mortal. Wkly. Rep. 2018, 66, 1402–1407. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ Br. Med. J. 2017, 356. [Google Scholar] [CrossRef]
- Mamun, A.A.; Mannan, M.; Doi, S.A.R. Gestational weight gain in relation to offspring obesity over the life course: A systematic review and bias- adjusted meta- analysis. Obes. Rev. 2014, 15, 338–347. [Google Scholar] [CrossRef]
- Nehring, I.; Lehmann, S.; von Kries, R. Gestational weight gain in accordance to the IOM/NRC criteria and the risk for childhood overweight: A meta-analysis. Pediatric Obes. 2013, 8, 218–224. [Google Scholar] [CrossRef]
- Karachaliou, M.; Georgiou, V.; Roumeliotaki, T.; Chalkiadaki, G.; Daraki, V.; Koinaki, S.; Dermitzaki, E.; Sarri, K.; Vassilaki, M.; Kogevinas, M.; et al. Association of trimester-specific gestational weight gain with fetal growth, offspring obesity, and cardiometabolic traits in early childhood. Am. J. Obstet. Gynecol. 2015, 212. [Google Scholar] [CrossRef] [Green Version]
- Olson, C.M.; Strawderman, M.S.; Hinton, P.S.; Pearson, T.A. Gestational weight gain and postpartum behaviors associated with weight change from early pregnancy to 1y postpartum. Int. J. Obes. 2003, 27, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridhar, S.B.; Darbinian, J.; Ehrlich, S.F.; Markman, M.A.; Gunderson, E.P.; Ferrara, A.; Hedderson, M.M. Maternal gestational weight gain and offspring risk for childhood overweight or obesity. Am. J. Obstet. Gynecol. 2014, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelan, S. Pregnancy: A “teachable moment” for weight control and obesity prevention. Am. J. Obstet. Gynecol. 2010, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, A.; Hedderson, M.M.; Brown, S.D.; Ehrlich, S.F.; Tsai, A.L.; Feng, J.R.; Galarce, M.; Marcovina, S.; Catalano, P.; Quesenberry, C.P. A telehealth lifestyle intervention to reduce excess gestational weight gain in pregnant women with overweight or obesity (GLOW): A randomised, parallel-group, controlled trial. Lancet Diabetes Endocrinol. 2020, 8, 490–500. [Google Scholar] [CrossRef]
- Kunath, J.; Gunther, J.; Rauh, K.; Hoffmann, J.; Stecher, L.; Rosenfeld, E.; Kick, L.; Ulm, K.; Hauner, H. Effects of a lifestyle intervention during pregnancy to prevent excessive gestational weight gain in routine care—The cluster-randomised GeliS trial. BMC Med. 2019, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifton, R.G.; Evans, M.; Cahill, A.G.; Franks, P.W.; Gallagher, D.; Phelan, S.; Pomeroy, J.; Redman, L.M.; Van Horn, L.; Grp, L.I.-M.R. Design of lifestyle intervention trials to prevent excessive gestational weight gain in women with overweight or obesity. Obesity 2016, 24, 305–313. [Google Scholar] [CrossRef]
- Poston, L.; Bell, R.; Croker, H.; Flynn, A.C.; Godfrey, K.M.; Goff, L.; Hayes, L.; Khazaezadeh, N.; Nelson, S.M.; Oteng-Ntim, E.; et al. Effect of a behavioural intervention in obese pregnant women (the UPBEAT study): A multicentre, randomised controlled trial. Lancet Diabetes Endocrinol. 2015, 3, 767–777. [Google Scholar] [CrossRef]
- Van Horn, L.; Peaceman, A.; Kwasny, M.; Vincent, E.; Fought, A.; Josefson, J.; Spring, B.; Neff, L.M.; Gernhofer, N. Dietary Approaches to Stop Hypertension Diet and Activity to Limit Gestational Weight: Maternal Offspring Metabolics Family Intervention Trial, a Technology Enhanced Randomized Trial. Am. J. Prev. Med. 2018, 55, 603–614. [Google Scholar] [CrossRef]
- Cirulli, E.T.; Guo, L.N.; Swisher, C.L.; Shah, N.; Huang, L.; Napier, L.A.; Kirkness, E.F.; Spector, T.D.; Caskey, C.T.; Thorens, B.; et al. Profound Perturbation of the Metabolome in Obesity Is Associated with Health Risk. Cell Metab. 2019, 29, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Elliott, P.; Posma, J.M.; Chan, Q.; Garcia-Perez, I.; Wijeyesekera, A.; Bictash, M.; Ebbels, T.M.D.; Ueshima, H.; Zhao, L.C.; van Horn, L.; et al. Urinary metabolic signatures of human adiposity. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Holmes, E.; Loo, R.L.; Stamler, J.; Bictash, M.; Yap, I.K.S.; Chan, Q.; Ebbels, T.; De Iorio, M.; Brown, I.J.; Veselkov, K.A.; et al. Human metabolic phenotype diversity and its association with diet and blood pressure. Nature 2008, 453, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Tzoulaki, I.; Castagne, R.; Boulange, C.L.; Karaman, I.; Chekmeneva, E.; Evangelou, E.; Ebbels, T.; Kaluarachchi, M.R.; Chadeau-Hyam, M.; Mosen, D.; et al. Serum metabolic signatures of coronary and carotid atherosclerosis and subsequent cardiovascular disease. Eur. Heart J. 2019, 40, 2883–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, J.R.; Holmes, E.; Veselkov, K.A.; Lindon, J.C.; Nicholson, J.K. A Unified Conceptual Framework for Metabolic Phenotyping in Diagnosis and Prognosis. Trends Pharmacol. Sci. 2019, 40, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wurtz, P.; Auro, K.; Makinen, V.P.; Kangas, A.J.; Soininen, P.; Tiainen, M.; Tynkkynen, T.; Jokelainen, J.; Santalahti, K.; et al. Metabolic profiling of pregnancy: Cross-sectional and longitudinal evidence. BMC Med. 2016, 14, 205. [Google Scholar] [CrossRef] [Green Version]
- Cecatti, J.G.; Souza, R.T.; Sulek, K.; Costa, M.L.; Kenny, L.C.; McCowan, L.M.; Pacagnella, R.C.; Villas-Boas, S.G.; Mayrink, J.; Passini, R.; et al. Use of metabolomics for the identification and validation of clinical biomarkers for preterm birth: Preterm SAMBA. BMC Pregnancy Childbirth 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Maitre, L.; Fthenou, E.; Athersuch, T.; Coen, M.; Toledano, M.B.; Holmes, E.; Kogevinas, M.; Chatzi, L.; Keun, H.C. Urinary metabolic profiles in early pregnancy are associated with preterm birth and fetal growth restriction in the Rhea mother-child cohort study. BMC Med. 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- White, S.L.; Pasupathy, D.; Sattar, N.; Nelson, S.M.; Lawlor, D.A.; Briley, A.L.; Seed, P.T.; Welsh, P.; Poston, L.; Consortium, U. Metabolic profiling of gestational diabetes in obese women during pregnancy. Diabetologia 2017, 60, 1903–1912. [Google Scholar] [CrossRef] [Green Version]
- Fattuoni, C.; Mando, C.; Palmas, F.; Anelli, G.M.; Novielli, C.; Laudicina, E.P.; Savasi, V.M.; Barberini, L.; Dessi, A.; Pintus, R.; et al. Preliminary metabolomics analysis of placenta in maternal obesity. Placenta 2018, 61, 89–95. [Google Scholar] [CrossRef]
- Kadakia, R.; Nodzenski, M.; Talbot, O.; Kuang, A.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; O’Neal, S.K.; Lowe, L.P.; et al. Maternal metabolites during pregnancy are associated with newborn outcomes and hyperinsulinaemia across ancestries. Diabetologia 2019, 62, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Hellmuth, C.; Lindsay, K.L.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Association of maternal prepregnancy BMI with metabolomic profile across gestation. Int. J. Obes. 2017, 41, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Nikolova, V.; Papacleovoulou, G.; Bellafante, E.; Manna, L.B.; Jansen, E.; Baron, S.; Abu-Hayyeh, S.; Parker, M.; Williamson, C. Changes in LXR signaling influence early-pregnancy lipogenesis and protect against dysregulated fetoplacental lipid homeostasis. Am. J. Physiol.-Endocrinol. Metab. 2017, 313, E463–E472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postle, A.D.; Al, M.D.M.; Burdge, G.C.; Hornstra, G. The Composition of Individual Molecular-Species of Plasma Phosphatidylcholine in Human-Pregnancy. Early Hum. Dev. 1995, 43, 47–58. [Google Scholar] [CrossRef]
- Stirm, L.; Kovarova, M.; Perschbacher, S.; Michlmaier, R.; Fritsche, L.; Siegel-Axel, D.; Schleicher, E.; Peter, A.; Pauluschke-Frohlich, J.; Brucker, S.; et al. BMI-Independent Effects of Gestational Diabetes on Human Placenta. J. Clin. Endocrinol. Metab. 2018, 103, 3299–3309. [Google Scholar] [CrossRef] [PubMed]
- Hellmuth, C.; Lindsay, K.L.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Maternal Metabolomic Profile and Fetal Programming of Offspring Adiposity: Identification of Potentially Protective Lipid Metabolites. Mol. Nutr. Food Res. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Kliewer, S.A.; Sundseth, S.S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C.S.; Devchand, P.; Wahli, W.; Willson, T.M.; Lenhard, J.M.; et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator-activated receptors alpha and gamma. Proc. Natl. Acad. Sci. USA 1997, 94, 4318–4323. [Google Scholar] [CrossRef] [Green Version]
- Guyenet, S.J.; Carlson, S.E. Increase in Adipose Tissue Linoleic Acid of US Adults in the Last Half Century. Adv. Nutr. 2015, 6, 660–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinehr, T.; Wolters, B.; Knop, C.; Lass, N.; Hellmuth, C.; Harder, U.; Peissner, W.; Wahl, S.; Grallert, H.; Adamski, J.; et al. Changes in the serum metabolite profile in obese children with weight loss. Eur. J. Nutr. 2015, 54, 173–181. [Google Scholar] [CrossRef]
- Tonks, K.T.; Coster, A.C.F.; Christopher, M.J.; Chaudhuri, R.; Xu, A.M.; Gagnon-Bartsch, J.; Chisholm, D.J.; James, D.E.; Meikle, P.J.; Greenfield, J.R.; et al. Skeletal muscle and plasma lipidomic signatures of insulin resistance and overweight/obesity in humans. Obesity 2016, 24, 908–916. [Google Scholar] [CrossRef] [Green Version]
- Manna, P.; Jain, S.K. Phosphatidylinositol-3,4,5-Triphosphate and Cellular Signaling: Implications for Obesity and Diabetes. Cell. Physiol. Biochem. 2015, 35, 1253–1275. [Google Scholar] [CrossRef]
- Bergmann, R.L.; Bergmann, K.E.; Haschke-Becher, E.; Richter, R.; Dudenhausen, J.W.; Barclay, D.; Haschke, F. Does maternal docosahexaenoic acid supplementation during pregnancy and lactation lower BMI in late infancy? J. Perinat. Med. 2007, 35, 295–300. [Google Scholar] [CrossRef]
- Maitre, L.; Lau, C.H.E.; Vizcaino, E.; Robinson, O.; Casas, M.; Siskos, A.P.; Want, E.J.; Athersuch, T.; Slama, R.; Vrijheid, M.; et al. Assessment of metabolic phenotypic variability in children’s urine using H-1 NMR spectroscopy. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.O.; Barros, A.S.; Goodfellow, B.J.; Duarte, I.F.; Carreira, I.M.; Galhano, E.; Pita, C.; Almeida, M.D.; Gil, A.M. Following Healthy Pregnancy by Nuclear Magnetic Resonance (NMR) Metabolic Profiling of Human Urine. J. Proteome Res. 2013, 12, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Giskeodegard, G.F.; Davies, S.K.; Revell, V.L.; Keun, H.; Skene, D.J. Diurnal rhythms in the human urine metabolome during sleep and total sleep deprivation. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Xie, J.; Li, W.W.; Bai, S.J.; Wang, W.; Zheng, P.; Xie, P. Age-specific urinary metabolite signatures and functions in patients with major depressive disorder. Aging 2019, 11, 6626–6637. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.H.E.; Siskos, A.P.; Maitre, L.; Robinson, O.; Athersuch, T.J.; Want, E.J.; Urquiza, J.; Casas, M.; Vafeiadi, M.; Roumeliotaki, T.; et al. Determinants of the urinary and serum metabolome in children from six European populations. BMC Med. 2018, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Alexander, P.; Wu, L.J.; Hammer, R.; Cleaver, O.; McKnight, S.L. Dependence of Mouse Embryonic Stem Cells on Threonine Catabolism. Science 2009, 325, 435–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal-Witt, M.J.; Ramon-Krauel, M.; Samino, S.; Llobet, M.; Cuadras, D.; Jimenez-Chillaron, J.C.; Yanes, O.; Lerin, C. Untargeted metabolomics identifies a plasma sphingolipid-related signature associated with lifestyle intervention in prepubertal children with obesity. Int. J. Obes. 2018, 42, 72–78. [Google Scholar] [CrossRef]
- Mills, H.L.; Patel, N.; White, S.L.; Pasupathy, D.; Briley, A.L.; Ferreira, D.L.S.; Seed, P.T.; Nelson, S.M.; Sattar, N.; Tilling, K.; et al. The effect of a lifestyle intervention in obese pregnant women on gestational metabolic profiles: Findings from the UK Pregnancies Better Eating and Activity Trial (UPBEAT) randomised controlled trial. BMC Med. 2019, 17. [Google Scholar] [CrossRef]
- Cloarec, O.; Dumas, M.E.; Craig, A.; Barton, R.H.; Trygg, J.; Hudson, J.; Blancher, C.; Gauguier, D.; Lindon, J.C.; Holmes, E.; et al. Statistical total correlation spectroscopy: An exploratory approach for latent biomarker identification from metabolic H-1 NMR data sets. Anal. Chem. 2005, 77, 1282–1289. [Google Scholar] [CrossRef]
- Dona, A.C.; Kyriakides, M.; Scott, F.; Shephard, E.A.; Varshavi, D.; Veselkov, K.; Everett, J.R. A guide to the identification of metabolites in NMR-based metabonomics/metabolomics experiments. Comput. Struct. Biotechnol. J. 2016, 14, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using Nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN—A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Species | Beta GWG | 95% Confidence | p | Beta WL | 95% Confidence | p |
|---|---|---|---|---|---|---|
| PC (42:6) | −0.011 | (−0.015, −0.007) | 8.9 × 10−8 | 0.009 | (0.001, 0.018) | 3.5 × 10−2 |
| SM (d17:1/24:1) | −0.011 | (−0.015, −0.007) | 5.6 × 10−7 | 0.013 | (0.007, 0.019) | 1.2 × 10−5 |
| PC (O-18:1_20:4) | −0.015 | (−0.022, −0.009) | 2.2 × 10−6 | 0.026 | (0.017, 0.036) | 3.4 × 10−7 |
| PC (O-20:0_20:4) | −0.018 | (−0.025, −0.01) | 6.2 × 10−6 | 0.006 | (0.001, 0.011) | 2.0 × 10−2 |
| PC (40:8) | −0.013 | (−0.019, −0.007) | 2.4 × 10−5 | 0.011 | (0.005, 0.016) | 2.4 × 10−4 |
| PC (16:0_20:4) | 0.009 | (0.005, 0.013) | 2.8 × 10−5 | −0.004 | (−0.007, 0) | 2.8 × 10−2 |
| PC (O-18:1_16:0) | −0.011 | (−0.016, −0.006) | 2.9 × 10−5 | 0.010 | (0.002, 0.017) | 9.7 × 10−3 |
| SM (d18:2/24:1) | −0.014 | (−0.021, −0.008) | 6.4 × 10−5 | 0.007 | (0.002, 0.012) | 6.4 × 10−3 |
| PC (O-24:2_20:4) | −0.009 | (−0.014, −0.005) | 1.2 × 10−4 | 0.006 | (0.002, 0.01) | 5.8 × 10−3 |
| PC (O-22:1_18:1) | −0.014 | (−0.021, −0.007) | 1.5 × 10−4 | 0.009 | (0.005, 0.014) | 9.8 × 10−5 |
| SM (d18:1/24:1) | −0.010 | (−0.016, −0.005) | 4.3 × 10−4 | 0.006 | (0.001, 0.011) | 1.4 × 10−2 |
| PC (O-16:0_22:4) | −0.012 | (−0.019, −0.005) | 9.0 × 10−4 | 0.003 | (−0.001, 0.007) | 1.6 × 10−1 |
| LPC (O-18:1) | −0.014 | (−0.022, −0.006) | 9.7 × 10−4 | 0.047 | (0.02, 0.074) | 9.0 × 10−4 |
| LPC (O-16:1) | −0.011 | (−0.017, −0.004) | 1.2 × 10−3 | 0.082 | (0.034, 0.13) | 9.4 × 10−4 |
| PC (O-24:1_20:4) | −0.008 | (−0.013, −0.003) | 1.2 × 10−3 | 0.007 | (0.003, 0.012) | 2.7 × 10−3 |
| PI (16:0_16:1) | 0.140 | (0.055, 0.226) | 1.5 × 10−3 | −0.016 | (−0.026, −0.006) | 1.3 × 10−3 |
| PC (16:0_22:5) | 0.022 | (0.009, 0.036) | 1.6 × 10−3 | −0.008 | (−0.014, −0.003) | 3.8 × 10−3 |
| SM (d18:2/14:0) | 0.009 | (0.002, 0.016) | 1.2 × 10−2 | −0.013 | (−0.019, −0.007) | 1.1 × 10−4 |
| LPC (O-16:0) | −0.009 | (−0.016, −0.002) | 1.4 × 10−2 | 0.064 | (0.03, 0.098) | 3.3 × 10−4 |
| PC (18:1_18:2) | 0.079 | (0.013, 0.145) | 2.0 × 10−2 | −0.014 | (−0.022, −0.006) | 7.1 × 10−4 |
| PC (16:1_18:2) | 0.013 | (0.002, 0.025) | 2.2 × 10−2 | −0.012 | (−0.017, −0.006) | 1.6 × 10−4 |
| PI (16:0_18:2) | 0.012 | (−0.004, 0.029) | 1.4 × 10−1 | −0.009 | (−0.014, −0.004) | 8.9 × 10−4 |
| LPS(O-18:0) | −0.005 | (−0.012, 0.002) | 1.8 × 10−1 | 0.156 | (0.082, 0.229) | 5.3 × 10−5 |
| DG (18:1_18:2) | −0.013 | (−0.041, 0.015) | 3.4 × 10−1 | −0.010 | (−0.017, −0.004) | 1.5 × 10−3 |
| PC (14:0_18:2) | 0.007 | (−0.017, 0.031) | 5.7 × 10−1 | −0.017 | (−0.025, −0.01) | 1.3 × 10−5 |
| PC (15:0_18:2) | 0.000 | (−0.009, 0.009) | 9.6 × 10−1 | −0.008 | (−0.012, −0.003) | 9.8 × 10−4 |
| Characteristics | Control (n = 50) | Intervention (n = 64) | p Values |
|---|---|---|---|
| Age (y, 15 weeks) | 34 ± 4 | 34 ± 4 | 0.16 |
| Weight (kg, 35 weeks) | 95 ± 12 | 91 ± 13 | 0.07 |
| BMI (kg/m2, 35 weeks) | 35 ± 4 | 34 ± 4 | 0.06 |
| Gestational Weight Gain (kg) | 10.6 ± 5.1 | 9.6 ± 4.6 | 0.14 |
| Weight Loss postpartum (kg) | 9.1 ± 7.2 | 9.4 ± 6.7 | 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, C.-H.E.; Taylor-Bateman, V.; Vorkas, P.A.; Graça, G.; Vu, T.-H.T.; Hou, L.; Chekmeneva, E.; Ebbels, T.M.D.; Chan, Q.; Van Horn, L.; et al. Metabolic Signatures of Gestational Weight Gain and Postpartum Weight Loss in a Lifestyle Intervention Study of Overweight and Obese Women. Metabolites 2020, 10, 498. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10120498
Lau C-HE, Taylor-Bateman V, Vorkas PA, Graça G, Vu T-HT, Hou L, Chekmeneva E, Ebbels TMD, Chan Q, Van Horn L, et al. Metabolic Signatures of Gestational Weight Gain and Postpartum Weight Loss in a Lifestyle Intervention Study of Overweight and Obese Women. Metabolites. 2020; 10(12):498. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10120498
Chicago/Turabian StyleLau, Chung-Ho E., Victoria Taylor-Bateman, Panagiotis A. Vorkas, Gonçalo Graça, Thanh-Huyen T. Vu, Lifang Hou, Elena Chekmeneva, Timothy M. D. Ebbels, Queenie Chan, Linda Van Horn, and et al. 2020. "Metabolic Signatures of Gestational Weight Gain and Postpartum Weight Loss in a Lifestyle Intervention Study of Overweight and Obese Women" Metabolites 10, no. 12: 498. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo10120498